
Vaccinium gaultherioides Bigelow, une espèce commune du Massif central longtemps confondue avec le rare V. uliginosum L.
Title
Vaccinium gaultherioides Bigelow, a common Massif central species that was long mistaken for the rare, endangered V. uliginosum L.
Résumé
Nous signalons pour la première fois la présence de V. uliginosum dans le Massif central. Cette plante tétraploïde se distingue de V. gaultherioides, qui est diploïde, par ses feuilles odorantes, moins arrondies et généralement plus grandes, sa floraison plus abondante et la formation de très nombreuses réitérations. Les deux espèces se mélangent rarement, V. uliginosum vivant dans les zones les plus humides. V. uliginosum correspond à une nouvelle espèce arctico-alpine pour le Massif central où elle s’y trouve en limite sud de son aire de distribution, ce qui lui confère une grande signification biogéographique.
Abstract
We report the presence of V. uliginosum in the Massif Central for the first time. This tetraploid plant differs from diploid V. gaultherioides by odoriferous, less rounded and generally larger leaves, its flowering is more abundant, and it forms numerous reiterations. The two species rarely mix, with V. uliginosum living in the wettest areas. It is a new arctico-alpine species for the Massif Central, where it is found at the southern limit of its range giving it great biogeographical significance.
1. Introduction
En Scandinavie, V. uliginosum L. est abondant entre 0 et 1 500 m dans presque tous les milieux : berges des lacs, tourbières, sous-bois et lisières forestières, éboulis, escarpements rocheux, etc. (planche 1). Dans les déserts froids de l’est du Groenland, les V. uliginosum s. l. forment souvent des tapis avec des tiges courtes hautes de 2-10 cm : ces plantes ont été décrites par Lange (1890) sous le nom de V. uliginosum subsp. microphyllum (planche 2). Hagerup (1933) a montré qu’il s’agit d’un diploïde autofertile, mais il a également observé au Groenland un cytotype tétraploïde autostérile nettement plus grand dans toutes ses partie et identique au V. uliginosum L. décrit par Linné en Scandinavie.


V. uliginosum L. est un taxon tétraploïde nettement plus grand que le diploïde lorsqu’ils poussent dans des conditions écologiques équivalentes (Löve, 1950 ; Young, 1970). Il présente un certain polymorphisme, notamment des fruits (Shishkin & Bobrov, 1952) qui peuvent même être curieusement pyriformes de 15 mm de long (planche 2).
En Amérique du Nord, V. uliginosum pousse principalement entre 60° et 70° de latitude nord, mais atteint le nord de l’Utah et de la Californie. Young (1970) y distingue cinq sous espèces qui ne sont pas retenues dans la Flora of North America (Vander Kloet, 2009). Cependant, la distinction de deux cytotypes (2x et 4x) a été largement confirmée dans diverses régions d’Europe et d’Amérique du Nord.
Bien que la distribution des deux cytotypes soit relativement bien cernée depuis longtemps, beaucoup de flores et travaux d’écologie suivent les interprétations de Webb (1972) qui considère que la sous espèce microphyllum est uniquement (sub)arctique en Europe et/ou reprennent les cartes et interprétations de Jacquemard (1996) et son équipe qui apportent des observations concernant la biologie et notamment le comportement et l’abondance des bourdons selon l’étendue des populations (Mayer et al., 2012) mais repose malheureusement sur une interprétation partiale de la bibliographie, ne reconnaissant qu’une espèce de V. uliginosum dont le cytotype le plus répandu serait un « autotétraploïde d’origine récente » (planche 3).
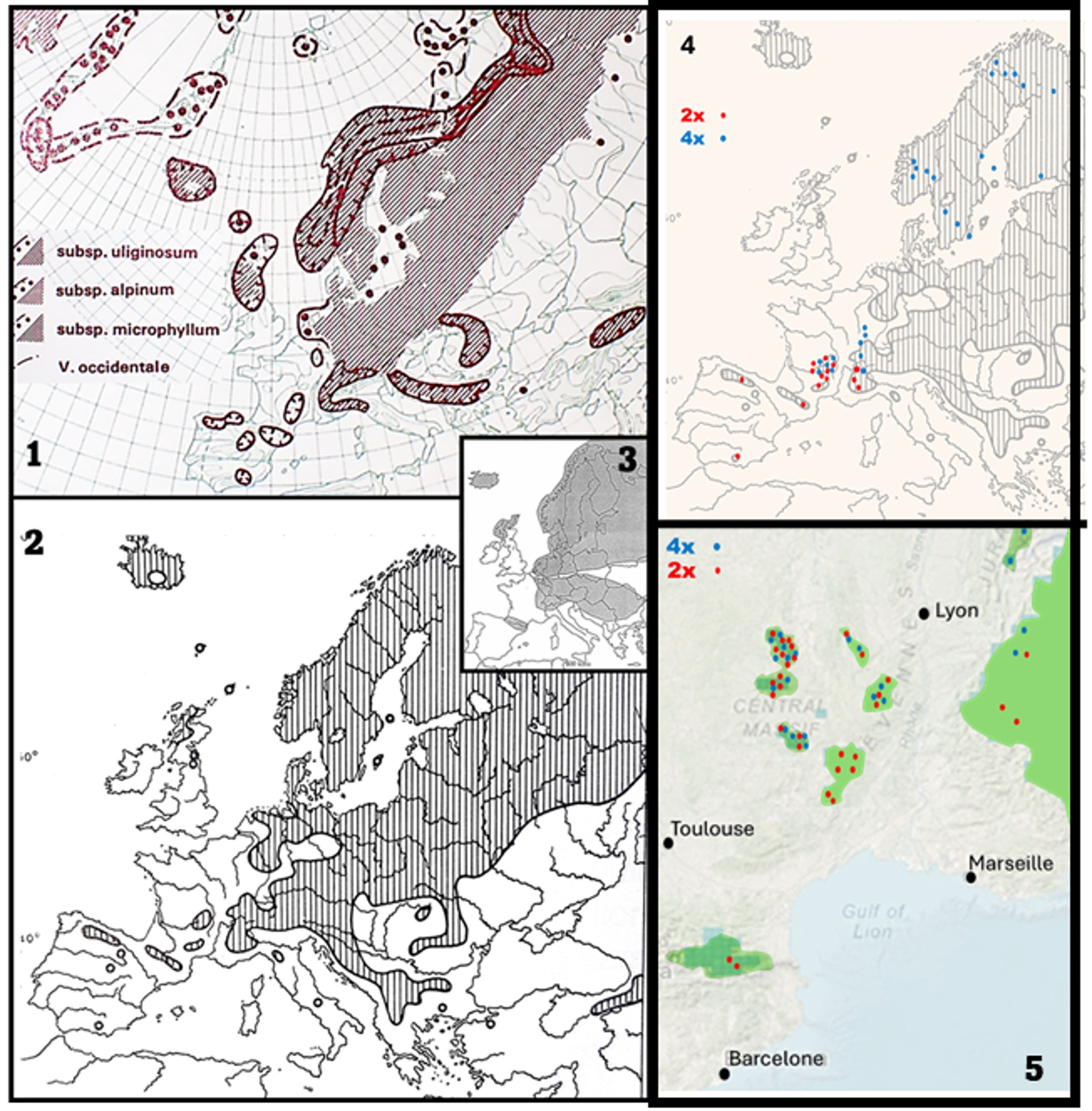
Dans la péninsule Ibérique, le diploïde est signalé (sous les noms de V. uliginosum subsp. microphyllum et subsp. gaultherioides) dans la chaîne pyrénéo-cantabrique (Urritia, 2000), le Système central et la Serra da Estrela (Jansen et al., 2000) avec pour limite sud la Sierra Nevada (figure 3). En Italie, sa limite sud se trouve dans les Apennins centrales (Pietro et al., 2007) plus à l’est, les plantes de Bulgarie signalées sous le nom de V. uliginosum L. entre 1 700 et 2 500 m correspondent également au diploïde comme le montrent leurs petites feuilles arrondies (Kozuharov, 1982).
Naturellement, le port et les dimensions (tiges, feuilles) des diploïdes, dispersés en populations parfois très isolées les unes des autres (Amérique du Nord, Scandinavie, Alpes centrales, Asie, Sibérie, Andalousie), varient énormément. Cependant, toutes les études morphologiques (pilosité, présence et distribution des stomates, des poils et glandes sur les limbes, nombres et dimensions des fleurs) et génétiques (marqueurs nucléaires et chloroplastiques) n’ont pour le moment donné aucun élément tangible qui justifierait la description d’un diploïde européen (V. uliginosum subsp. microphyllum Lange, Fl. Groenl., 1890) distinct d’un diploïde américain (Alsos et al., 2005 ; Gauthier, 2018). Le plus ancien binôme qui s’applique aux populations diploïdes américaines et eurasiatiques serait donc celui de Bigelow (1816) créé pour les airelles des marais qui poussent dans les White Mountain (New Hampshire : limite Canada/nord-est des États Unis). V. gaultherioides renferme trois lignées génétiques (Alsos et al., 2005) qui correspondent à des entités biogéographiques bien connues (amphi-atlantique, Béringie, arctico-alpine). En outre, les données moléculaires suggèrent (Eidesen et al., 2007) que V. uliginosum serait un allopolyploïde entre V. gaultherioides et un taxon proche des endémiques que l’on rencontre encore sur les îles atlantiques (cf. V. cylindraceum Sm. des Açores) ou en Méditerranée nord-orientale (cf. V. arctostaphylos L. qui se trouvent dans quelques massifs de Bulgarie et du nord de l’Anatolie : une distribution assez caractéristique des reliques tertiaires). En Turquie, les citations de V. uliginosum correspondent à la sous espèce diploïde avec également une distribution relictuelle dans les montagnes qui font face à la mer Noire (Stevens, 1978).
L’inflorescence des diploïdes est réduite à 1-2[3] fleurs portées par un court pédicelle, tandis que celle des tétraploïdes présente parfois un axe avec 2-3[5] bractées (forme intermédiaire avec celles, plus primitives et en grappe, des Vaccinium tertiaires qui sont des petits arbustes à grandes feuilles et possèdent des grappes (qui portent souvent plus de 8 fleurs chez les V. arctostaphyllos, planche 4). Soulignons que les Vaccinium arbustifs tropicaux possèdent également des inflorescences fournies (Argent, 2019).

On adoptera donc les deux binômes suivants pour les plantes présentes en France :
- V. gaultherioides Bigelow = V. uliginosum subsp. microphyllum Lange, 1890 : 91, = V. microphyllum (Lange) Hagerup ex Löve, Bot. Not. :49, 1950, = V. uliginosum subsp. gaultherioides (Bigelow) S.B. Young, = V. uliginosum p. p. in Chassagne Fl. Auv., 1957, Sauvage, Fl. France, 1982, = V. uliginosum auct. in Villars 1993, Garraud, 2003, Tison et al., 2014, = V. uliginosum subsp microphyllum (Lange) Tolm. in Bolòs & Vigo,1995, Urrutia, 2000, Le Driant et al., 2022. Soulignons que Blume (1826) avait déjà décrit une plante indonésienne sous le nom de Vaccinium microphyllum (Flora Nederland. Indië 15 : 851) et que l’épithète microphyllum est utilisée dans plusieurs taxons infraspécifiques du genre Vaccinium ;
- V. uliginosum L. est le binôme qui s’applique aux airelles tétraploïdes décrites en Scandinavie et qui sont morphologiquement semblables à celles présentes en France, principalement dans le nord-est du territoire et, désormais, dans le Massif central.
2. Analyse de la ploïdie des plantes du Massif central et comparaison avec d’autres localités européennes
L’originalité de chacune de ces deux espèces est largement reconnue, sous divers rangs taxonomiques, en Suisse, en Italie et au Portugal, mais demeure contestée en Espagne (Villar, 1993) et dans le sud de la France. Pourtant ces deux espèces ont déjà clairement été distinguées dans le sud des Alpes (Gauthier et al., 2002) et même le Massif central (G. Thébaud, comm. pers. & CLF !). Comme chaque espèce possède un nombre chromosomique distinct (diploïdes versus tétraploïdes), nous avons étudié la ploïdie de plus de 250 individus selon les méthodes habituelles (Fridlender et al., 2014, 2015) dans des régions et habitats représentatifs des airelles des marais d’Europe occidentale.
Les localités de prélèvement (planche 3) sont les suivantes :
Plantes tétraploïdes
Massif central. Puy-de-Dôme (63), massifs du Sancy et du Cézallier, Picherande, ca. 1 200 m, ca. 1 600 m pente du Paillaret ; Montbert, ca. 1 235 m ; Chastreix à Rimat ; Besse-et-Saint Anastaise, Montagnoune, ca. 1 185 m ; Egliseneuve-d’Entraigues à Chambedaze, 1 190-1 250 m. Loire (42), Livradois Forez, Jeansagnière, 1 200-1 300 m (la Montagnette, la Litte, le Lignon) ; Loire/Puy-de-Dôme (42/63), crêtes du Forez, les Combes, 1 360 m ; Bazanne, 1 350 m ; Cantal/Lozère/Aveyron (15/48/12), Aubrac ; Nasbinals, Marchastel, 1 040-1 090 m (Pèchouzou, Pourcharesses, Salhiens) ; Ségur-les-Villes, 1 145 m vers la Gazelle ; fossés tourbeux des bois de Laguiole, ca. 1 300 m.
Alpes et Préalpes. Savoie/Haute-Savoie (73/74), Glières 1 400-1 440 m, 1 435 m ; les Saisies, 1 550 m, 1 600 m, 1 700 m ; Jura (39), Les Rousses, ca. 1 060 m ; Doubs (25), Frasne, 845 m ; Vosges (88), La Bresse, 910 m ; Isère (38), Séchilienne, Chamrousse, sous-bois tourbeux, ca. 1 270 m, ca 1 620 m.
Sud de la Scandinavie (provinces floristiques des localités). Suède, Dasland, Västerbotten ; Norvège, Øsfold, Oppland, Sogn og Fjordane, Buskerud, Akershus og Oslo ; Finlande, Uusimaa ;
Nord de la Scandinavie et Laponie. Suède, Torne Lappmark ; Norvège, Troms ; Finlande, Enontekiön Lappi.
Plantes diploïdes
Massif central. Puy-de-Dôme (63), massifs du Sancy et du Cézallier, Le Mont Dore, col de la Croix Morand, tourbières et landes, 1 350 m, 1 420 m ; puys de la Tache et de la Monne, 1 550 m, 1 650 m ; Chambon-sur-Lac, landes de la vallée de Chaudefour, puy Ferrand, montagne de la Platte 1 450 m, 1 530 m, 1 700 m, 1 800 m ; Picherande, landes vers 1 500 m, 1 650 m (puy de Paillaret) ; Loire/Puy-de-Dôme (42/63), Forez : col du Béal, pelouses, 1 400-1 450 m ; landes à Pierre-sur-Haute ca. 1 600m ; Cantal (15) : massif du Puy Mary, 1 400 m, vers Eylac, pelouses de pentes, ca. 1 500 m, landes de crêtes, 1 650 m ; Le Lioran, pelouses et landes du puy du Rocher, 1 400 m, 1 700 m ; Lozère (48) : prairies humides à Finiels, 1 300 m, landes et pelouses des crêtes du mont Lozère, 1 480 m, 1 550 m, 1 600 m, 1 650 m ; mont Aigoual ca. 1 550 m ; Haute-Loire (43), massif du Mézenc, les Estables, Croix de Peccata, lande et prairies, 1 560 m, 1 600 m, rochers et pentes rocailleuses 1 650 m, 1 700 m ; tourbières de Goudoffre 1 350 m ; tourbières de Rouchon 1 440 m.
Alpes et Préalpes. Drôme (26), Luz-la-Croix-Haute, massif de la Tête des Lauzon 1 900 m, 1 960 m ; Isère (38), Chamrousse, landes et pelouses ca. 1 650 m, 1 700 m, 2 000 m ; Savoie/Haute-Savoie (73/74), Chamonix, massif des Aiguilles Rouges, plusieurs individus entre le col des Montets 1 460 m et les pelouses sur dalles rocheuses vers 2 100 m ; Hautes Alpes (05), Orcières Merlette, sentier de Prapic au lac des Pisses (2 080 m, 2 200 m, 2 300 m et 2 500 m).
Massifs pyrénéo-cantabriques et Espagne. Pyrénées Orientales (66), massif du Carlit, 2 400 m ; Corsavy au-dessus de Batere, 2 400 m ; Asturias, Oviedo, à Valgrande ; Sierra Nevada, au Barranco de San Juan ca. 2 400 m.
Nous avons confronté nos observations morphologiques à l’analyse cytogénétique, notamment lorsque les plantes présentaient des caractères foliaires ambigus (planche 5). Sur le terrain, à l’exception de quelques échantillons (souvent broutés ou en conditions écologiques extrêmes), la reconnaissance des deux espèces dans les Alpes et le Massif central ne nous a jamais posé de réelles difficultés et le niveau de ploïdie a toujours été conforme aux identifications morphologiques sur les 250 individus analysés (2x et 4x).
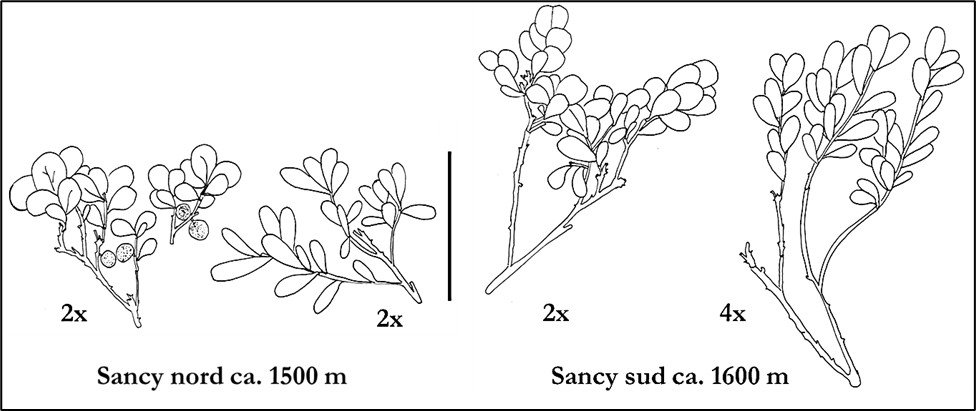
3. Conclusions
V. gaultherioides est l’espèce la plus commune en France. Dans le Massif central, elle est abondante sur les pentes et les crêtes de la plupart des massifs où elle pousse en mélange avec les myrtilles. Les plantes sont parfois géantes dans toutes leurs parties dans le Sancy, le long des ruisselets, tandis qu’elles sont petites dans les tourbières pâturées du Mézenc et souvent prostrées avec des petites feuilles sur les crêtes sèches et/ou ventées (mont Lozère) et dans les localités les plus méridionales (cf. sous espèces « nana » en Andalousie).
V. uliginosum est l’airelle des marais la plus rare et la plus localisée en France où elle ne pousse que dans les secteurs les plus humides (à la différence de ses habitats scandinaves ; planche 1). Elle se reconnaît aisément par plusieurs caractères morphologiques :
- les entrenœuds sont longs et nombreux sur les axes ;
- les réitérations sont abondantes et parfois envahissent l’individu entier ;
- limbes allongés nettement plus grands avec ou sans glandes à la base ;
- feuillage odorant surtout quand il est jeune : si on reste immobile dans un peuplement de uliginosum, il se dégage une puissante odeur d’abord douceâtre et agréable, puis rapidement narcotique, alors que les diploïdes sont souvent presque inodores. En été, en enfermant une branchette de chacune des deux espèces dans des boîtes ou sachets en plastique, après une nuit ou même quelques heures la différence devient flagrante !
- inflorescences portant souvent plus de 2-3 fleurs ; le nombre et la densité des inflorescences dépendent de l’âge : les relais végétatifs d’un jeune axe portent souvent une seule inflorescence (sub)terminale alors qu’en fin de vie ses relais sont progressivement envahis par la sexualité. Fréquemment la plupart des entrenœuds de ces relais portent des inflorescences et parfois aucune feuille ! Chez les deux espèces les axes âgés sont nettement plus florifères que les jeunes axes de sorte qu’un axe âgé de gaultherioides sera plus fructifère qu’une jeune tige vigoureuse de V. uliginosum. Il faut donc comparer des axes de même âge ;
- les fruits sont plus abondants et souvent plus gros que ceux du gaultherioides mais la texture comme le goût (chair blanche, légèrement gélatineuse) sont semblables.
Les plantes signalées dans le Massif central sous les noms de V. uliginosum et V. uliginosum subsp. uliginosum correspondent presque toutes à V. gaultherioides, très rarement à V. uliginosum L. puisque jusqu’à présent les deux cytotypes étaient confondus (sub V. uliginosum) et n’étaient pas considérés comme deux espèces distinctes.
Le Puy-de-Dôme et le Forez abritent de nombreuses populations de V. uliginosum ; elles sont plus dispersées dans l’Aubrac et le Cantal où elles sont d’autant plus menacées qu’elles se rencontrent à plus faible altitude (principalement en dessous de 1 300 m). Dans les Préalpes, les peuplements sont isolés les uns des autres et la limite sud atteinte par le tétraploïde se situe apparemment au niveau du massif de Belledonne (Isère) où nous n’en avons observé que des petits peuplements relictuels, principalement en sous-bois à basse altitude, alors que V. gaultherioides y est abondant jusqu’aux sommets. Partout, l’habituelle synergie drainages, enrésinement, surpâturage les menace et a vraisemblablement fait disparaître de nombreuses populations.
Les deux cytotypes sont particulièrement bien représentés dans le Cézallier où nous avons également mis en évidence la présence de trois espèces de canneberges (Fridlender, 2023). Ce petit territoire s’avère exceptionnel en Europe puisqu’au cours d’une journée d’herborisation on y observera facilement sept espèces de Vaccinium : trois assez communes dans tous le Massif central, V. vitis-idaea, V. myrtillus, V. gaultherioides, et quatre beaucoup plus localisées ou rares qu’il faudrait activement protéger (V. uliginosum, V. majuscula, V. microcarpus, V. oxycoccus) en préservant leurs habitats tous menacés par le tourisme, le surpâturage, les drainages et leurs conséquences (eutrophisation, piétinement…).
Bibliographie
Alsos I.G., Engelskjon T., Gielly L., Taberlet P. & Brochlman C., 2005. Impact of ices ages on circumpolar molecular diversity: insights from an ecological key species. Molecular Ecology 14 : 2739-2753.
Argent G., 2019. Rigiolepis and Vaccinium (Ericaceae) in Borneo. Edinburgh Journal of Botany 76 (1) : 55-172.
Eidesen P.B., Alsos G., Popp M., Stensrud O., Suda J. & Brochmann C., 2007. Nuclear vs. plastid data: complex Pleistocene history of a circumpolar key species. Molecular Ecology 16 : 3902-3925.
Fridlender A., 2023 Vaccinium majuscula (Chassagne) Fridl. in French Massif central and its related Scandinavian Oxycoccus palustris var major Lange. Biom 4 (1) : 35-54.
Fridlender A., Pustahija F., Šolić M.E., Abadžić S., Bourge M., Pech N., Šiljak-Yakovlev S. & Brown S.C., 2014. Is it possible to identify Colchicum neapolitanum s.l. and C. autumnale s.l. in vegetative stage? Biometry and flow cytometry approaches. Botanica Serbica 38 (1) : 21-34.
Fridlender A. & Brown S. 2015. La citometría de flujo como ayuda para identificación florísticas: confirmación de Colchicum lusitanum y C. multiflorum (Colchicaceae) en Asturias. Flora Montiberica 61 : 7-15.
Garraud L., 2003. Flore de la Drôme. Atlas écologique et floristique. Conservatoire botanique national alpin de Gap-Charance, 925 p.
Gauthier R., 2018. Ericaceae In S. Payette (ed.), Flore nordique du Québec et du Labrador, Presses de l’Université de Laval, Québec, 3 : 225-330.
Gauthier R., Polidori J.-L. & Gervais C. 2002. Vaccinium gaultherioides Bigelow (Ericaceae) en Haute Tinée, Alpes Maritimes. Le Monde des plantes 477 : 1922.
Hagerup O., 1933. Studies on polyploid ecotypes in Vaccinium uliginosum L. Hereditas 18 : 122-128.
Hultén E. & Fries M., 1986. Atlas of North European Vascular Plants. North of the Tropic of Cancer. I-III (1 text & 2 map-volumes), Koeltz Scientific Books.
Jacquemart A.-L., 1996. Vaccinium uliginosum L. In Biological Flora of the British Isles, Journal of ecology 84 : 771-785.
Jansen J., Nijs H.C.M. & Paiva J.A.R., 2000. Some notes on Vaccinium uliginosum subsp. gaultherioides (Bigelow) S.B. Young, a new taxon to the flora of Portugal. Portugaliae Acta Biologica 19 : 177-186.
Kozuharov S, 1982. Ericaceae. In D. Jordanov et al. (eds), Flora Reipublicae popularis bulgaricae VIII : 288-326.
Lange J., 1890. Florae Groenlandicae. Andet Oplag, Kjohenhavn : 90-91.
Le Driant F., Ferrus L. & Pellicier P., 2022. Plantes de montagne, Alpes, Massif central, Jura et Vosges. Biotope, Mèze, 1 144 p.
Löve A., 1950. Some innovations and nomenclatural suggestions in the Icelandic flora. Botaniska Notiser 1 : 45-60.
Mayer C., Michez D., Chyzy A., Brédat E. & Jacquemart A.-L., 2012. The abundance and pollen foraging behaviour of bumble bees in relation to population size of whortleberry (Vaccinium uliginosum). Plos One 7 (11) : 10 p.
Pietro R., Castonica C. & Copiz R., 2007. Sulla presenza di Vaccinium gaultherioides Bigelow in Italia centrale. Biogeographia 28 : 275-285.
Sauvage C., 1982. Ericales In M. Guinochet & R. de Vilmorin (eds), Flore de France, CNRS, Paris, 4 : 1322-1353.
Shishkin B.K. & Bobrov E.G., 1952. Genus 1111. Vaccinium, In Flora C.C.C.P. XVIII : 94-102.
Stevens P.F., 1978. Vaccinium L. In P.H. Davis (ed.), Flora of Turkey, University Press of Edinburgh 6 :100-104.
Urritia P.M., 2000. Ericaceae. In I. Aizpuru et al. (eds.), Claves ilustradas de la flora del País Vasco y territorios limítrofes. Servicio Central de Publicaciones del Gobierno Vasco, Vitoria Gasteiz : 233-236.
Vander Kloet S.P., 2009. Vaccinium L. In Flora of North America committee (ed.), Flora of North America, north of Mexico, Oxford University Press, New York, 8 : 515-529.
Villar L., 1993. Vaccinium L. In S. Castroviejo et al. (eds.), Flora Iberica, CSIC, Madrid, 4 : 520-523.
Webb D.A., 1972. Vaccinium L. In T.G. Tutin et al. (eds.), Flora Europaea, Cambridge University Press 3 : 12-13.
Young S.B., 1970. On the taxonomy and distribution of Vaccinium uliginosum. Rhodora 72 (792) : 439-459.
Remerciements
Je remercie M. Bourge, K. Madionna (Gif), G. Thébault (CLF !) et O. Bablee pour leur aide notamment concernant les analyses et les herbiers.


