
Contribution au Prodrome des végétations de France : les Elyno myosuroidis – Seslerietea caeruleae Braun-Blanq. 1948
Title
Contribution to the prodromous of French vegetations: the Elyno myosuroidis – Seslerietea caeruleae Braun-Blanq. 1948
Résumé
Dans le cadre du Prodrome des végétations de France, sous l’égide de la Société française de phytosociologie, les auteurs présentent ici la classe des pelouses basiphiles orophiles et arctico-alpines des Elyno myosuroidis – Seslerietea caeruleae : déclinaison au niveau français.
Abstract
Within the framework of the prodromous of French vegetations, under the sponsorship of French Society of plant sociology, the authors present the subalpine basophilous, orophilous and arctic-alpine grasslands class of the Elyno myosuroidis – Seslerietea caeruleae: declination at the French level.
1. Introduction
Les hautes montagnes d’Europe hébergent une grande diversité de pelouses soumises aux particularités des climats locaux alpins et subalpins d’altitude. Les pelouses alpines subordonnées à de très longues périodes d’enneigement occupent une place à part dans la classe des Salicetea herbaceae (récemment synthétisée pour la France par Corriol & Mikolajczak, 2014). Pour le reste, les pelouses développées sur des sols acides se répartissent en deux classes : les Nardetea strictae pour l’étage subalpin (synthétisés pour la France par de Foucault, 2012) et les Caricetea curvulae pour l’étage alpin ; on trouvera dans Corriol & Mikolajczak (2017) une récente synthèse de cette dernière classe pour la France, incluant une présentation synthétique des caractéristiques écologiques de l’étage alpin.
La classification des pelouses basiphiles, beaucoup plus diversifiées, est nettement plus complexe. En fonction des auteurs, elles ont été réparties, en plus des Salicetea herbaceae pour les plus chionophiles, dans une à quatre classes de végétation. Historiquement, la plus ancienne d’entre elles fut nommée Elyno – Seslerietea par Braun-Blanquet (1948). Cette classe ne comprenait alors qu’un seul ordre, les Seslerietalia caeruleae, antérieurement nommé par Braun-Blanquet & Jenny (1926), ordre dans lequel Braun-Blanquet (1948) regroupe toutes les pelouses basiphiles orophiles des Alpes et des Pyrénées.
Suite à une note d’Oberdorfer (1957 : 305) qui s’interroge sur l’opportunité de séparer les pelouses cryophiles arctico-alpines dans une classe distincte des Elyno – Seslerietea, Ohba (1974) propose la nouvelle classe des Carici rupestris – Kobresietea bellardii, dans laquelle il positionne l’alliance de l’Oxytropido – Elynion Braun-Blanq. 1950. Ohba, étendant cette nouvelle classe jusqu’à l’extrême Nord-Est asiatique et à l’Alaska, la considère à répartition circumarctico-alpine. Toutefois, il n’intègre pas dans sa synthèse les syntaxons alpino-subalpins des montagnes d’Europe moyenne, ce qui ne permet pas d’évaluer leur distribution vis-à-vis de la classification strictement arctico-alpine qu’il propose. Par ailleurs, la caractérisation floristique qu’il donne pour sa nouvelle classe ne montre pas un ensemble très cohérent. Seuls Carex rupestris, C. myosuroides et l’annuelle Comastoma tenella semblent pertinents (avec éventuellement Saussurea alpina très sporadique), alors que sont proposées comme caractéristiques de classe une quinzaine d’endémiques absentes de nos massifs (Androsace lehmanniana, Artemisia glomerata, A. trifurcata, Bupleurum triradiatum, Calamagrostis purpurascens, C. vanheurckii, Comastoma pulmonaria, Gentiana algida, Hierochloe alpina, Minuartia arctica, M. macrocarpa, Patrinia sibirica, Ptilagrostis mongolica, Tofieldia coccinea, Zygadenus sibiricus). Par ailleurs sont aussi proposées comme caractéristiques de classe des espèces au moins aussi bien si ce n’est plus largement représentées dans notre synthèse des Elyno – Seslerietea (Aster alpinus, Astragalus alpinus, Dryas octopetala, Minuartia verna, Oxytropis campestris, O. lapponica, Potentilla crantzii).
Cette proposition est suivie notamment par Carrillo & Ninot (1992), Theurillat et al. (1995), Bardat et al. (2004), Rivas-Martínez et al. (2001, 2011), Lancioni et al. (2011), Biondi et al. (2014), Mucina et al. (2016), Julve (2017). Elle est également reprise dans une récente synthèse à l’échelle circumarctique qui, tout comme celle d’Ohba, ne prend pas en compte les syntaxons alpino-subalpins et, qui plus est, ne propose pas de caractérisation floristique de la classe (Walker et al., 2017). En revanche, elle n’est pas adoptée par Gruber (1978, 1979a), Rivas-Martínez et al. (1991a), Rivas-Martínez & Costa (1998), Benito Alonso (2006), Illa et al. (2006), Peyre & Font (2011), Prunier et al. (2019), qui maintiennent ces végétations au sein des Elyno – Seslerietea. L’analyse numérique de Peyre & Font (2011), sur la base de 2 259 relevés en provenance des Pyrénées et des monts Cantabriques, ne s’accorde pas avec la séparation d’une classe à part pour héberger l’Oxytropido – Elynion.
Sur la base d’associations végétales décrites par différents auteurs dans les Alpes maritimes et ligures et les Pyrénées orientales, Barbero (1968) identifie un ensemble de pelouses xérophiles ouvertes sur roches carbonatées des hautes montagnes méridionales dans lesquelles se mêlent des espèces eurosibériennes et des orophytes méditerranéennes et sud-européennes. Il propose de les regrouper dans un ordre nouveau, les Astragaletalia sempervirentis. Il intègre dans cet ordre plusieurs alliances. La première, l’Avenion montanae Barbero 1968, regroupe les pelouses des Alpes occidentales sous influences méditerranéennes les plus orophiles, essentiellement subalpines. À ses côtés, il considère comme son géovicariant pyrénéen le Festucion scopariae Braun-Blanq. 1948, alliance décrite des Pyrénées orientales et intégrée par son auteur à la classe de Elyno – Seslerietea (Braun-Blanquet, 1948). Deux autres alliances, l’Avenion sempervirentis Barbero 1968 et l’Ononidion cenisiae proposé à titre provisoire (finalement validée dans sa thèse en 1972), sont adjointes par cet auteur à sa nouvelle classe, tout en précisant pour cette dernière que sa classification reste problématique. Ces deux alliances regroupent des associations des Alpes du Sud, plus thermophiles, d’altitudes plus modérées. L’année suivante, Barbero et Bonin (1969) introduisent une nouvelle classe de végétation pour accueillir l’ordre des Astragaletalia sempervirentis ainsi qu’un ordre géovicariant, les Seslerietalia tenuifoliae Horvat 1930, des Balkans : les Festuco – Seslerietea. Ce shéma de classification, repris dans Barbero et al. (1971, 1975), est suivi dans les Pyrénées par Gruber (1979b), Ninot (1988) et Montserrat & Villar (1975, 1987). Ces derniers étendent l’aire des Astragaletalia sempervrientis vers les Pyrénées occidentales en décrivant, en plus du Festucion scopariae, deux nouvelles alliances (les Saponarion caespitosae P. Monts. & Villar 1987 et Thymelaeion nivalis nom. inval.). Si, pour des raisons strictement nomenclaturales (voir note en p. 8), le nom de Festuco – Seslerietea caeruleae Barbero & Bonin 1969 sera retenu dans certaines classifications adoptant la séparation des Carici rupestris – Kobresietea, la conception que s’en font leurs auteurs ne fera pourtant pas école.
En effet, Gaultier (1989), dans son travail de synthèse sur les pelouses basiphiles du pourtour méditerranéen, conclut au rattachement de l’Ononidion cenisiae Barbero 1972, incluant partiellement l’Avenion montanae et l’Avenion sempervirentis des Alpes occidentales, à la classe des Festuco – Brometea, dans l’ordre des Ononidetalia striatae Braun-Blanq. 1950, et donc à leur exclusion de la classe des Festuco – Seslerietea. Il maintient par ailleurs le Festucion scopariae pyrénéen dans les Elyno – Seslerietea. Royer (1991 : 113) suit la position de Gaultier dans sa synthèse des Festuco – Brometea, en y intégrant l’Ononidion cenisiae. Cette option est adoptée par Theurillat et al. (1995) dans leur classification pour les Alpes, l’Avenion montanae étant intégré aux Elyno – Seslerietea.
Côté Pyrénées, Rivas-Martínez et al. (1991b) proposent le rattachement du Festucion scopariae à l’ordre des Ononidetalia striatae Braun-Blanq. 1950, positionné dans une nouvelle classe qu’ils nomment Festuco hystricis – Ononidetea striatae, regroupant les pelouses xérophiles basiphiles riches en chaméphytes en coussinets du pourtour méditerranéen (surtout occidental). Cette position est suivie par Ninot (1996), alors que Vigo &et Massales (1996), Carrillo et al. (2000), Soriano (2001), Benito Alonso (2006) et Illa et al. (2006) maintiennent le Festucion scopariae Braun-Blanq. 1948 dans la classe initiale des Elyno – Seslerietea Braun-Blanq. 1948, en accord avec les résultats obtenus par Gaultier (1989).
De même, Valls (2003), dans sa révision des Ononidetalia striatae basée sur des analyses factorielles des correspondances (AFC) de 220 syntaxons élémentaires, obtient l’exclusion rapide (dès la deuxième AFC) du Festucion scopariae Braun-Blanq. 1948 qu’il renvoie aux Elyno – Seslerietea Braun-Blanq. 1948. Il montre que la classe des Festuco hystricis – Ononidetea striatae telle que définie par Rivas-Martínez et al. (1991b) est hétérogène et que le Festucion scopariae Braun-Blanq. 1948 s’y insère mal. Cette même étude rejette l’Ononidion cenisiae Barbero 1972 vers les Festuco – Brometea Braun-Blanq. ex Soó 1947, tout en excluant les Ononidetalia striatae de cette classe.
Peyre et Font (2011), sur la base d’un traitement numérique de 2 259 relevés de pelouses en provenance des Pyrénées et des monts Cantabriques, obtiennent une classification regroupant dans une même classe (Elyno – Seslerietea) toutes les pelouses subalpines de ces massifs, dont le Festucion scopariae Braun-Blanq. 1948, incluant les communautés des Pyrénées centrales et occidentales au rang de sous-alliance [Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart. et al. 1991].
Rivas-Martínez et al. (2002, 2011) et Mucina et al. (2016) à leur suite présentent des classifications qui ne tiennent pas compte des résultas de Valls (2003) ni de Peyre & Font (2011). En revanche, la classification italienne de Biondi et al. (2014) intègre l’ensemble des Astragaletalia aux Elyno – Seslerietea (classe pour laquelle ils retiennent le nom de Festuco – Seslerietea).
Les classifications proposées à l’échelle de la France (Bardat et al., 2004 ; Julve, 2017) rejettent les Festuco – Seslerietea comme classe à part pour les pelouses orophiles méridionales. Elles intègrent l’Avenion montanae aux Seslerietalia caeruleae et, suivant Gaultier (1989), relèguent l’Ononidion cenisiae et en partie l’Avenion sempervirentis aux Ononidetalia striatae dans les Festuco – Brometea. Julve (2017) suit également Gaultier (1989) pour la position du Festucion scopariae dans les Seslerietalia caeruleae. En revanche, Bardat et al. (2004) placent l’alliance pyrénéenne dans les Ononidetalia striatae comme proposé par Rivas-Martínez et al. (1991b), mais sans adopter la nouvelle classe des Festuco hystricis – Ononidetea striatae des auteurs ibériques. On trouvera dans le tableau 1 un comparatif des différentes classifications citées.
2. Matériels et méthodes
L’analyse réalisée pour cette synthèse prend en compte les types de végétation présents en France ou en périphérie immédiate, rattachés classiquement aux alliances suivantes : Avenion montanae Barbero 1968, Festucion scopariae Braun-Blanq. 1948, Ononidion cenisiae Barbero 1972, Avenion sempervirentis Barbero 1968, Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Primulion intricatae Braun-Blanq. ex Vigo 1972, Laserpitio nestleri – Ranunculion thorae Vigo 1979 et Oxytropido – Elynion myosuroidis Braun-Blanq. 1950. Elle s’est faite en deux temps, d’abord sur la base du traitement statistique de 1 112 relevés répartis sur la chaîne pyrénéo-cantabrique, issus de la bibliographie et de la base de données Flora du Conservatoire botanique national des Pyrénées et de Midi-Pyrénées. Les syntaxons élémentaires retenus pour cette chaîne de montagne et les colonnes synthétiques qui y sont présentées sont issus des classifications obtenues à partir de ces relevés (méthode de Ward). Dans un deuxième temps, aux 97 colonnes synthétiques ainsi obtenues, sont adjointes 82 colonnes synthétiques issues de la bibliographie pour les syntaxons des Alpes et du Jura. Le tableau synthétique ainsi obtenu (tableau 2) a été trié à l’aide de l’algorithme Twinspan du logiciel Juice (Tichý, 2002) puis remanié manuellement. Les noms des idiotaxons sont indexés sur le référentiel national Taxref version 7. Deux colonnes synthétiques issues d’une publication pyrénéenne postérieure à notre travail d’analyse statistique ont été ajoutées ultérieurement.
L’article ici présenté contribue à la déclinaison au rang des associations végétales du Prodrome des végétations de France paru en 2004 (Bardat et al., 2004), dans la continuité et la finalisation de nos précédentes contributions sur les végétations de pelouses alpines (Corriol & Mikolaczak, 2014, 2017). Il a rigoureusement et entièrement suivi le processus de publication défini dans le cadre du travail intitulé Prodrome des végétations de France 2 ou « PVF2 » sous l’égide de la Société française de phytosociologie, en coordination avec le Muséum national d’histoire naturelle et les conservatoires botaniques nationaux (Bioret & Royer, 2009 ; Bioret et al., 2013). Le manuscrit et les tableaux phytosociologiques, préalablement relus par Bruno de Foucault, membre du comité scientifique du Conservatoire botanique national des Pyrénées et de Midi-Pyrénées, a été envoyé au coordinateur du Muséum national d’histoire naturelle le 8 janvier 2020 et diffusé au comité de lecture mis en place dans le cadre du PVF2 le 15 janvier 2020. À l’échéance des trois mois dévolus à cette phase de relecture (le 15 avril 2020), nous avons reçu des commentaires, corrections et demandes de compléments de Claude Misset, Emmanuel Catteau et Alexis Mikolajczak. Le manuscrit révisé, prenant en compte l’intégralité de ces retours, a été rendu au comité de lecture dans sa version définitive le 27 avril 2021 et resté depuis lors en attente par défaut d’un support de publication, suite à l’arrêt des Documents phytosociologiques, revue ayant hébergé les dernières contributions au PVF2. Nous prenons la décision de publier ce travail dans le périodique en ligne Carnets botaniques, adapté à la publication de longs manuscrits et tableaux volumineux afin de ne pas différer plus longtemps la diffusion de ce travail.
3. Résultats de la classification des relevés pyrénéo-cantabriques
Épurée de quelques relevés marginaux, exclus d’emblée de la classe (appartenant notamment au Festucetum gautieri caricetosum montanae Vigo & Massales 1996 et à l’Eryngio bourgatii – Plantaginetum mediae seslerietosum caeruleae Vigo 1979) et de quelques relevés non classables, la première classification générale obtenue (figure 1) montre :
– la position très marginale de trois associations, l’Onosmo bubanii – Caricetum humilis Carreras, Carrillo, Ninot, Soriano & Vigo in Ninot 1996, le Teucrio pyrenaici – Astragaletum catalaunici Carrillo & Ninot 1990 et le Gentiano acaulis – Primuletum intricatae Vigo 1972 ; les deux premières peuvent être assez facilement renvoyées vers les Ononidetalia striatae (Festuco – Brometea) et sont donc exclues des Elyno – Seslerietea ; en revanche, la dernière peut difficilement être rattachée à une autre classe qu’aux Elyno – Seslerietea dont elle présente un nombre assez important de taxons caractéristiques ; en conséquence, nous traitons cette association dans cette synthèse, mais dans une position marginale ;
– la séparation très nette d’un Festucion scopariae, incluant les typus du Festucion scopariae et du Saponarion caepistosae P. Monts. & Villar 1987, du reste des relevés ;
– l’identification dans ce dernier groupe de trois branches pouvant correspondre aux alliances de l’Oxytropido – Elynion, du Primulion intricatae et du Salicion pyrenaicae.

La figure 2 illustre une deuxième classification épurée de l’Onosmo bubanii – Caricetum humilis, du Teucrio pyrenaici – Astragaletum catalaunici et du Gentiano acaulis – Primuletum intricatae. La séparation très nette du Festucion scopariae y est confirmée, de même que, pour le reste des relevés, la distinction de trois groupes, attribuables aux Salicion pyrenaicae, Primulion intricatae et Oxytropido – Elynion. Il est intéressant de constater que la position de l’Oxytropido – Elynion est variable en fonction des classifications obtenues. En effet, il se branche ici avec le Primulion intricatae, en opposition au Salicion pyrenaicae alors que la première classification branchait Salicion pyrenaicae et Primulion intricatae ensemble. Ce résultat milite pour le regroupement de ces trois alliances dans la même unité supérieure, en l’occurrence l’ordre des Oxytropido – Elynetalia, dont l’Oxytropido – Elynion constitue le type.
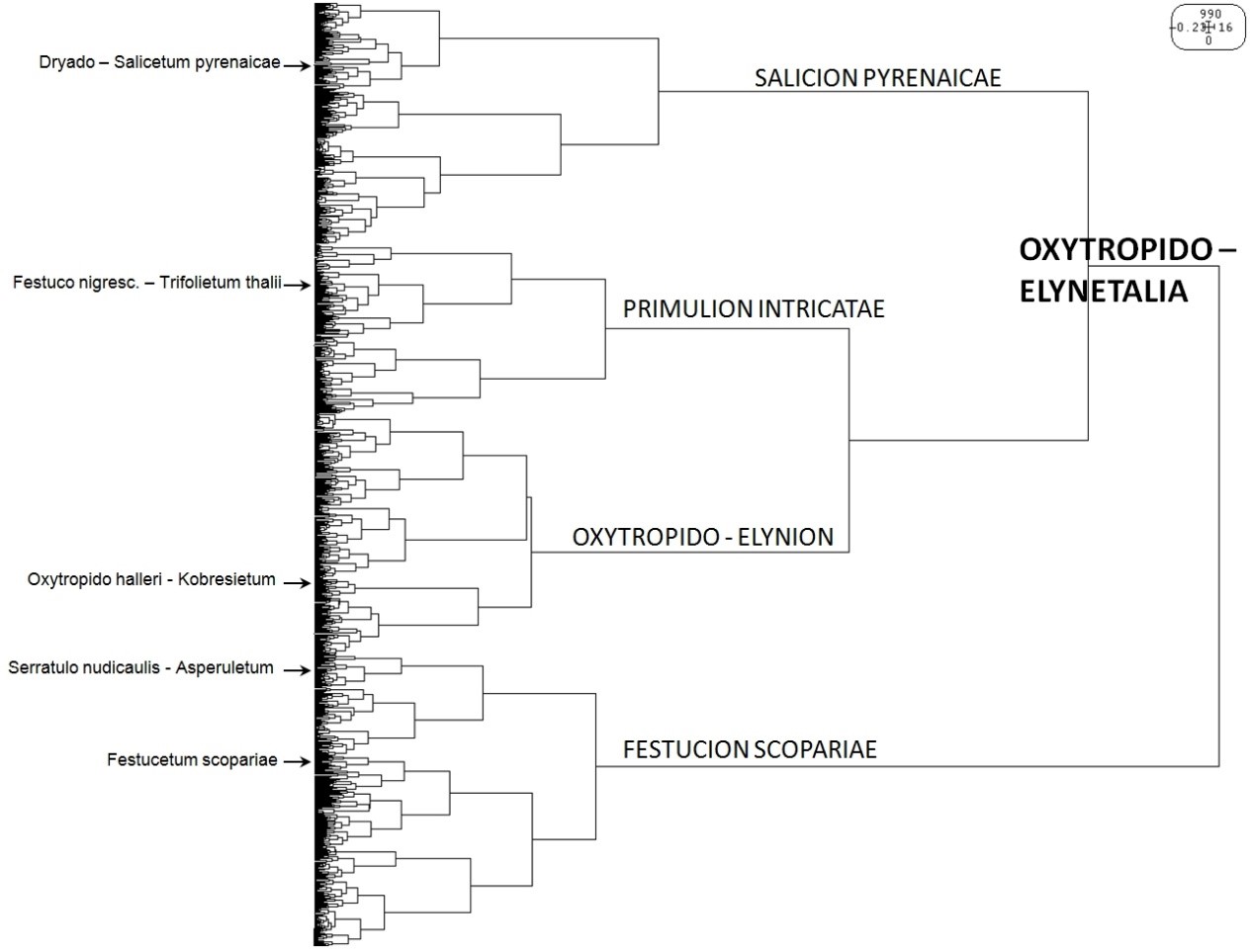
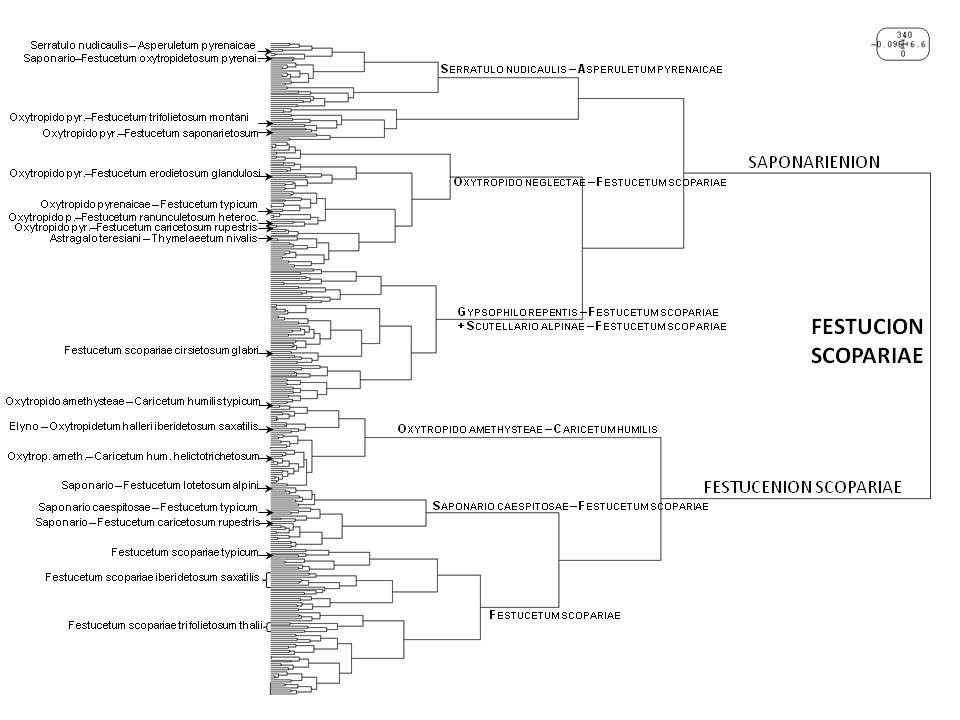
La figure 3 illustre une classification automatique des relevés rattachés à l’alliance du Festucion scopariae et notamment l’existence d’un groupe de relevés qui ne se rattachent à aucune association préalablement décrites (Gypsophilo repentis – Festucetum scopariae et Scutellario alpinae – Festucetum scopariae). Elle semble aussi illustrer l’existence de deux sous-groupes de relevés pouvant correspondre grosso modo aux sous-alliances du Festucenion scopariae et Saponarienion caespitosae, même si, en fonction des classifications obtenues, certaines associations se distribuent différemment.
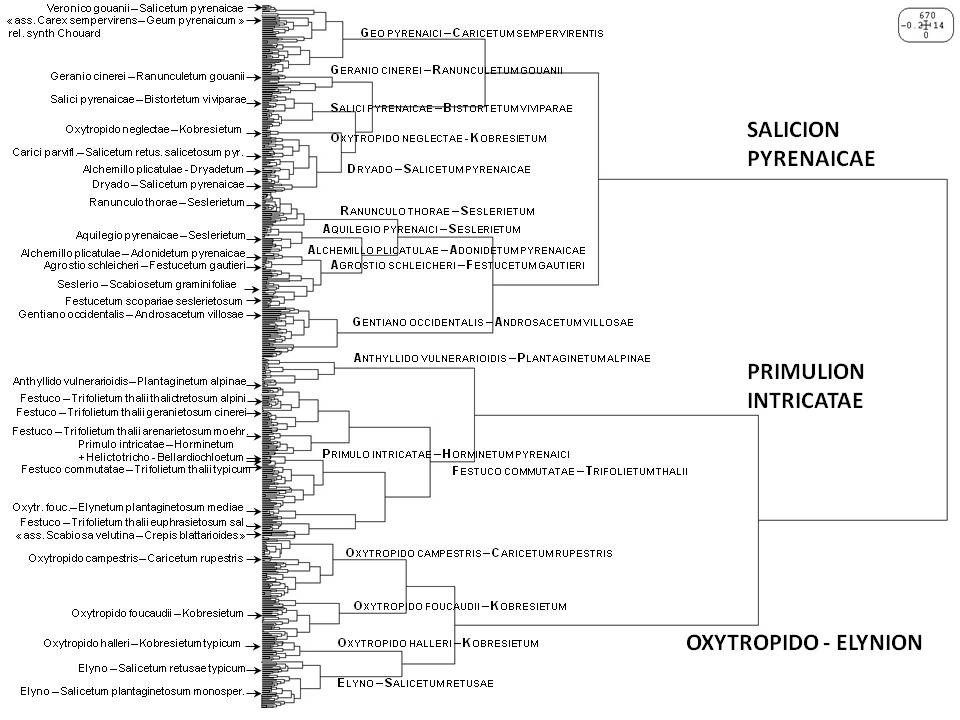
La figure 4 montre une classification automatique obtenue pour les autres relevés pyrénéens, se distribuant nettement en trois groupes de relevés déjà identifiés dans la classification générale (figure 1) et correspondant aux trois alliances du Salicion pyrenaicae, du Primulion intricatae et de l’Oxytropido – Elynion. Toutefois, si la position de la plupart des associations reste stable dans toutes les classifications obtenues, certaines ici placées au sein de l’alliance du Salicion pyrenaicae changent de position en fonction des classifications. Ces associations en limite de plusieurs alliances illustrent les liens existant entre le Salicion pyrenaicae et (i) l’Oxytropido – Elynion (via les Salici pyrenaicae – Bistortetum et Oxytropido neglectae – Kobresietum), (ii) le Festucion scopariae (via le Gentiano occidentalis – Androsacetum villosae) et (iii) le Primulion intricatae (via l’Alchemillo plicatulae – Adonidetum pyrenaicae). Au vu des résultats de la classification générale de la classe et des groupes floristiques caractéristiques et différentiels obtenus (tableau 2), nous avons redistribué ces quatre associations dans les trois alliances citées. La classification interne au Salicion pyrenaicae semble en outre montrer deux sous-groupes de relevés comme déjà identifié par Peyre & Font (2011). Ces auteurs ont proposé de considérer ces deux sous-groupes comme deux sous-alliances, les Laserpitio nestleri – Ranunculenion thorae et Dryado octopetalae – Salicenion pyrenaicae, mais sans en valider formellement les noms. Faisant référence au Laserpitio nestleri – Ranunculion thorae Vigo 1979, alliance invalide (art. 8), pour placer ces deux sous-alliances, la première, autonyme, est invalide. La seconde, également non typifiée, est aussi invalide (art. 5). Lazare (2017) reprend ce schéma et valide le deuxième nom. Toutefois, la distribution des associations entre elles, la circonscription et la caractérisation floristique qu’il donne aux deux sous-unités ne sont que partiellement corroborées par notre analyse. Le Laserpitio – Ranunculenion se trouve essentiellement différencié négativement, et il n’est pas évident de leur attribuer une limite écologique si tranchée.
4. Classification des colonnes synthétiques issues du traitement des relevés pyrénéo-cantabriques et des colonnes synthétiques issues de la bibliographie pour les Alpes et le Jura
La classification automatique (Twinspan) du tableau synthétique ainsi obtenu montre un poids géographique très fort. Elle sépare en tout premier lieu les associations alpiennes et jurassiennes d’un côté, à l’exception de quelques associations alpiennes cryophiles, et de l’autre les associations pyrénéennes, plus ces quelques associations cryophiles. Le poids des espèces à répartition arctico-alpine communes aux deux massifs dans les situations les plus froides et aux plus hautes altitudes explique probablement en partie ce rapprochement, alors que, pour les autres pelouses, la diversification de la flore et de l’endémisme propre à chacun des massifs les sépare plus nettement.
En deuxième niveau de classification, le poids géographique reste assez élevé pour les communautés alpino-jurassiennes, avec une séparation nette des associations du Jura et des Préalpes du Nord qui est en partie liée aux pelouses particulièrement riches en hautes herbes du Caricion ferrugineae, bien représentées dans ces massifs. Se détache aussi nettement un groupe d’associations méridionales xérophiles traité ici dans l’Avenion sempervirentis. Le résidu de relevés, traditionnellement rangés dans l’alliance du Seslerion caeruleae, est moins bien circonscrit.
Côté « pyrénéen », une séparation plus écologique se dessine, avec un groupe xérophile (Festucion scopariae), un groupe cryophile alpien, un groupe cryophile pyrénéen (Oxytropido – Elynion sensu stricto) et deux groupes de relevés pyrénéens correspondant aux alliances du Primulion intricatae et du Salicion pyrenaicae. La subdivision du Salicion pyrenaicae en deux sous-alliances est supportée par notre CAH, mais difficile à circonscrire floristiquement.
5. Choix retenus pour cette synthèse
Nos résultats ne permettent pas de reconnaître statistiquement un ensemble syntaxonomique regroupant l’ensemble des pelouses xérophiles méridionales selon la conception de Barbero & Bonin (1969 ; « Festuco – Seslerietea » au sens original). Nous avons vu en effet que le poids géographique des espèces sépare les communautés des Alpes du Sud de celles des Pyrénées en les classant dans les groupes « alpien » et « pyrénéen » avec les autres associations de pelouses orophiles basiphiles. Par ailleurs, les associations végétales concernées présentent avec une fréquence élevée un lot important de taxons largement représentés dans les pelouses orophiles basiphiles (Sesleria caerulea, Minuartia verna, Gentiana verna, Poa alpina, Carex sempervirens, Potentilla crantzii, Euphrasia salisburgensis, Aster alpinus, etc.), pouvant être considérés comme caractéristiques d’une classe commune à l’ensemble. En conséquence, conformément à la plupart des synthèses récentes, nous ne retiendrons pas une classe à part pour ces communautés.
En revanche, compte-tenu de l’existence de blocs de taxons différentiels communs mis en évidence dans notre tableau, il ne nous paraît pas opportun de distribuer ces associations dans des unités de classification éloignées. C’est pourquoi nous choisissons la position conservatrice de regrouper l’ensemble de ces pelouses dans une unité commune, nous appuyant sur l’ordre existant des Astragaletalia sempervirentis, incluant deux alliances, une pyrénéenne (Festucion scopariae) et une alpienne (Avenion sempervirentis). On constate par ailleurs une cohérence de structure systématique par la ressemblance dans la composition des taxons végétaux (de rangs générique et familial, avec des vicariances) et dans la répartition des types biologiques.
Dans cette alliance alpienne, deux sous-unités peu caractérisées peuvent correspondre à l’Ononidenion cristatae Gaultier 1989 et à l’Ononido cristatae – Helictrotrichenion sempervirentis Gaultier 1989. L’Ononidion cristatae Barbero 1970 prov. est proposé comme synonyme syntaxonomique de l’Avenion sempervirentis, de même que le Festucion dimorphae Bonin 1969. Compte-tenu du lot important d’espèces orophiles des associations classées dans ces unités (Carduus defloratus, Carex sempervirens, Androsace vitaliana, Poa alpina, Helictotrichon sedenense, Antennaria dioica, Minuartia verna, Scutellaria alpina, Astragalus sempervirens, Senecio doronicum, Draba aizoides, Aster alpinus, Campanula scheuchzeri…), elles ont toute leur place au sein de la classe des Elyno – Seslerietea comme définie ici, avis partagé avec Jean-Marie Royer (comm. pers.), coordinateur de la déclinaison des pelouses basiphiles collinéennes à montagnardes (Festuco – Brometea) pour le Prodrome des végétations de la France. L’alliance pyrénéenne peut également être scindée en deux sous-alliances (Festucenion scopariae et Saponarienion caespitosae), comme proposé antérieurement, même si cette séparation est peu nette.
En ce qui concerne les pelouses cryophiles (Oxytropido – Elynion au sens classique), les traitements effectués sur les relevés pyrénéens tout comme le tri du tableau synthétique général des groupements français et limitrophes n’identifient pas une unité cohérente les regroupant et ne s’accordent pas avec la circonscription d’une classe à répartition arctico-alpine à part (concept de Carici rupestris – Kobresietea). Tout en reconnaissant que cette question de classification demanderait à être étudiée à l’échelle circumarctique et eurosibérienne, n’ayant pas obtenu de résultats positifs appuyant cette séparation, nous opterons pour l’option conservatrice de maintenir ces pelouses au sein de la grande classe des Elyno – Seslerietea. En lien avec nos résultats, cette option permet de mieux caractériser et différencier floristiquement les unités retenues à l’échelle nationale, ceci en faisant le choix, en cohérence avec le raisonnement fait pour les Astragaletalia, de ne pas disjoindre les communautés alpiennes et pyrénéennes comme suggéré par le tri automatique du tableau synthétique, mais en les regroupant dans une même alliance pour laquelle on peut identifier un ensemble significatif de taxons caractéristiques. Les espèces bien représentées dans les associations concernées, qui peuvent être par ailleurs considérées comme caractéristiques de classe et ainsi y asseoir leur positionnement, sont notamment Gentiana verna, Poa alpina, Minuartia verna, Potentilla crantzii, Sesleria caerulea, Carex sempervirens, Aster alpinus, Draba aizoides.
Ce faisant, l’ensemble de ces pelouses se trouve, au sein de l’alliance de l’Oxytropido – Elynion, rattaché au groupe des pelouses pyrénéennes psychrophiles (Salicion pyrenaicae) et mésophiles (Primulion intricatae) dans l’ordre des Oxytropido – Elynetalia, ceci dans toutes les classifications automatiques effectuées. À l’intérieur de l’Oxytropido – Elynion, on peut distinguer trois groupes de relevés bien différenciés, un pyrénéen et deux alpiens, considérés ici au rang de sous-alliances.
Les pelouses alpiennes (hormis cryoxérophiles et sous forte influence méditerranéenne) sont toutes regroupées dans les Seselerietalia caeruleae, ordre surtout différencié négativement par rapport aux deux autres. Les pelouses basiphiles plus ou moins psychrophiles sont reprises classiquement dans le Caricion ferrugineae, ayant son optimum dans le Jura et les Préalpes du Nord. Dans cette alliance, deux unités ont été identifiées : une sous-alliance type nettement psychrophile et une sous-alliance d’altitudes plus modérées ou d’expositions plus chaudes, pour laquelle nous proposons le nom de Calamogrostienion variae, qui reprend à ce nouveau rang le Calamagrostion variae Sill. 1932. La pénétration de taxons d’ourlets à valeur différentielle est significative avec la participation des Mulgedio-Aconitetea variegati dans le Caricenion ferrugineae et des Trifolio – Geranietea sanguinei du côté thermophile (Calamagrostienion variae). Ces végétations se montrent ainsi comme des pelouses-ourlets parfois issues d’anciennes pratiques de fauche ou à caractère plus ou moins primaire (couloirs d’avalanche, pentes fortes, vires exiguës…).
Les pelouses subalpines du Seslerion caeruleae montrent deux groupes assez distincts. L’un représente des pelouses thermoxérophiles et l’autre des pelouses mésophiles acidiclinophiles. Déjà identifiées dans le Jura par Béguin (1972) au rang de sous-alliance (respectivement Drabo – Seslerienion caeruleae et Agrostio capillaris – Seslerienion caeruleae), ces syntaxons peuvent être élevés au rang d’alliance en suivant Delarze et al. (1998), ainsi que Prunier et al. (2019). L’alliance du Seslerion caeruleae, centrée alors sur les étages subalpin supérieur et alpin, se retrouve surtout différenciée des autres alliances de l’ordre par la présence d’espèces des Oxytropido – Elynetalia (Dryas octopetala, Silene acaulis, Leontopodium nivale subsp. alpinum, Sedum atratum, Oxytropis campestris…) L’association type de l’Avenion montanae (le Seslerio albicantis – Helictotrichetum sedenensis Lippmaa 1933) possède également ces espèces différentielles plus ou moins cryophiles et ne contient en outre que très peu d’espèces caractéristiques des Astragaletalia sempervirentis. Nous proposons en conséquence d’inclure l’Avenion montanae et son association type dans le Seslerion caeruleae. Les Astragaletalia sempervirentis se retrouvent alors réduits à une seule alliance dans les Alpes françaises, l’Avenion sempervirentis. Les autres associations qui ont pu être classées par certains auteurs dans l’Avenion montanae appartiennent selon nous à l’Avenion sempervirentis. Enfin, une unité de pelouses mésophiles et acidiclinophiles des Alpes du Sud se distingue du reste et justifie la réintroduction de l’alliance de l‘Anthoxantho villosi – Plantaginion brutiae proposée par Barbero & Quézel (1975).
Ainsi définie, la classe des Elyno – Seslerietea inclut des pelouses primaires à déterminisme climatique ou stationnel de l’étage alpin, des pelouses primaires à déterminisme stationnel et des pelouses secondaires en situation d’estives/alpages liées au pastoralisme, à l’étage subalpin et à l’étage montagnard. Elles sont en contact avec des végétations des Salicetea herbaceae en situation plus enneigée, des Caricetea curvulae et Nardetea strictae (Festucetalia spadiceae) sur des sols plus acides et des Thlaspietea rotundifolii sur des sols moins stabilisés ou plus rocheux. Dans le cas des pelouses secondaires, une dynamique peut s’observer vers des ourlets des Mulgedio – Aconitetea variegati (ou plus marginalement les Trifolio – Geranietea sanguinei), des landes des Rhododendro – Vaccinietalia, des fourrés des Betulo carpaticae – Alnetea viridis et des forêts résineuses subalpines des Vacccinio – Piceetea abietis. Des plantes orophiles de pelouses acidiphiles pénètrent les végétations des Elyno – Seslerietea par introgression structurelle (issues d’un stade homologue d’une autre série, selon la définition de Gillet et al., 1991 : 332, ou « structurale » d’après Julve, 1988 : 71), particulièrement dans les situations de stress édapho-climatique extrême favorables à la formation d’humus brut, y compris sur les roches carbonatées. Dans ces mêmes situations existent des liens avec des landines arctico-alpines de l’Arctostaphylo alpini – Cetrarion. On note en particulier dans notre tableau les herbacées acidiphiles suivantes : Pilosella lactucella, Festuca nigrescens, Antennaria dioica, Botrychium lunaria, Campanula scheuchzeri, Plantago alpina, Scorzoneroides pyrenaica, Agrostis rupestris, Euphrasia minima, Juncus trifidus, Luzula nutans, Phyteuma hemisphaericum, Carex ericetorum, Minuartia sedoides…
Les végétations des Elyno – Seslerietea ont été reconnues dans la plupart des hauts massifs d’Europe moyenne et méridionale, des monts Cantabriques à l’ouest (Herrera Gallastegui et al., 1991 ; Jiménez-Alfaro et al., 2014), au moins jusque dans les Carpates à l’est (Tibor & Tamás, 2012 ; Svitková & Šibík, 2013). Vers le sud, hormis la chaîne pyrénéo-cantabrique, on les trouve dans les Apennins du sud (Lancioni et al., 2011) et dans les montagnes balkaniques, jusqu’en Grèce au 40e parallèle (Redžić, 2003). Au nord-ouest, elles sont signalées au nord des îles Britanniques (Shimwell, 1968). Au nord, des végétations rattachées aux Carici – Kobresietea sont signalées jusqu’au 70e parallèle, du nord de la Norvège au nord-ouest de la Sibérie, ainsi qu’au Svalbard au 78e parallèle (Koroleva, 2015). Elles sont également mentionnées au Groenland (Daniëls, 1982), qui toutefois relève du continent américain.
Note nomenclaturale
L’article 35 du code de nomenclature phytosociologioque prévoit le rejet d’un nom de syntaxon composé de deux noms de taxons lorsque ce syntaxon est scindé en deux et que les taxons éponymes se retrouvent séparés chacun dans l’un des deux nouveaux syntaxons. Ainsi pour le nom de la classe des Elyno – Seslerietea amputée des communautés à Elyne des Oxytropido – Elynetalia, constituant la nouvelle classe des Carici – Kobresietea, le plus ancien nom disponible deviendrait celui de Festuco – Seslerietea Barbero & Bonin 1969, dans la mesure où l’on accepte la synonymie syntaxonomique entre les Elyno – Seslerietea et les Festuco – Seslerietea. Ce choix nous semble toutefois discutable pour deux raisons. D’abord parce que, si Carex myosuroides est une caractéristique fortement élective de l’Oxytropido – Elynion, alliance type des Oxytropido – Elynetalia, il n’est pas absent du reste des Elyno – Seslerietea. Ensuite parce que la synonymie syntaxonomique avec les Festuco – Seslerietea implique soit l’intégration des Astragaletalia sempervirentis dans les Elyno – Seslerietea, soit leur mise en synonymie avec les Seslerietalia caeruleae (comme Bardat et al., 2004 ou Biondi et al., 2014). Or, en pratique, les auteurs sont restés fort partagés sur ces options. Rivas-Martínez et al. (2001), par exemple, ont même adopté le nom de Festuco – Seslerietea en place d’Elyno – Seslerietea, sans inclure dans leur schéma aucune unité de la circonscription originale des Festuco – Seslerietea. Quant à Bardat et al. (2004), s’ils citent explicitement les Astragaletalia sempervirentis en synonymie des Seslerietea caeruleae et s’ils intègrent à cet ordre l’Avenion sempervirentis, ils en excluent l’Ononidion cenisiae et le Festucion scopariae et ils ne mentionnent pas l’Avenion montanae. Par ailleurs, à la suite de Gaultier (1989), ils considèrent l’Avenion sempervirentis comme une sous-alliance de l’Ononidion cenisiae. En conséquence, l’Avenion sempervirentis s’y trouve virtuellement dans deux classes différentes : au rang d’alliance dans les Elyno – Seslerietea (nommées Festuco – Seslerietea) et au rang de sous-alliance dans les Festuco – Brometea.
Avertissement : nous avons adopté dans cette synthèse un certain nombre de noms de syntaxons proposés dans des thèses éditées bénéficiant d’une impression propre, lisible et qui sont accessibles dans les principales bibliothèques scientifiques traitant de botanique et de phytosociologie. Nous considérons que ces caractéristiques répondent aux conditions de publication effective définies dans l’article 1 du code de nomenclature phytosociologique ansi qu’à l’esprit de celui-ci : assurer des publications validantes lisibles et accessibles. Nous n’avons en effet pas privilégié une interprétation réductrice qui nous aurait conduits à devoir valider ici de nombreux nouveaux noms. Ainsi en est-il pour les thèses de Barbero (1972), Lejoly (1975), Gruber (1978), Gauthier (1989).
ELYNO MYOSUROIDIS – SESLERIETEA CAERULEAE Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 147)
[syn. : Festuco – Seslerietea caeruleae Barbero & Bonin 1969 (Bull. Soc. Bot. France 116 : 227) (synonyme syntaxonomique inclus)].
Typus classis : Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 225).
Végétations de pelouses basiphiles orophiles et arctico-alpines.
On peut considérer que cette classe est caractérisée en France par Carex sempervirens subsp. sempervirens, Gentiana verna, Potentilla crantzii, Sesleria caerulea, Minuartia verna, Poa alpina, Aster alpinus subsp. alpinus, Bistorta vivipara, Alchemilla alpigena, Euphrasia salisburgensis, Anemone alpina, Draba aizoides, Agrostis alpina, Myosotis alpestris, Bartsia alpina, Erigeron alpinus, Anemone narcissiflora, auxquels on peut ajouter des formes d’altitude de Helianthemum oelandicum (subsp. alpestre, subsp. piloselloides), de Helianthemum nummularium (subsp. tomentosum, subsp. grandiflorum, subsp. pyrenaicum) et peut-être de Trifolium pratense, dont il faudrait vérifier la répartition de la var. villosum (= subsp. nivale). Helictotrichon sedenense est également très bien représenté, à l’exception des communautés les plus septentrionales des Alpes et du Jura. En outre, on note la bonne représentation de Campanula scheuchzeri s. l., Antennaria dioica, Cerastium arvense subsp. strictum, Botrychium lunaria transgressives issues des pelouses orophiles acidiphiles.
La hiérarchie interne de cette classe, nous l’avons vu, est complexe et fortement soumise au poids géographique de la flore. Dans la structuration retenue, à partir du tableau synthétique trié-ordonné (tableau 2), on peut assez bien mettre en évidence d’une part les pelouses xérophiles méridionales, montagnardes à subalpines, des Astragaletalia sempervirentis et d’autre part un groupe de pelouses psychrophiles à cryophiles alpines à subalpines, incluant toutes les pelouses pyrénéennes à l’exclusion du Festucion scopariae, ainsi que les pelouses alpines cryoxérophiles des Alpes et des Pyrénées, constituant les Oxytropido – Elynetalia myosuroidis. Le troisième groupe résiduel, plus hétérogène, comprend les relevés des autres pelouses alpiennes non rattachés aux deux ordres précédents ; ils sont rassemblés par défaut dans l’ordre des Seslerietalia caeruleae.
À l’intérieur des Astragaletalia sempervirentis, on trouve aussi une forte hiérarchie géographique, avec les pelouses des Alpes du Sud d’un côté (Avenion sempervirentis) et des Pyrénées de l’autre (Festucion scopariae). La hiérarchie interne des Oxytropido – Elynetalia est d’abord écologique, isolant d’une part les pelouses cryoxérophiles des Alpes et des Pyrénées (Oxytropido – Elynion), d’autre part les pelouses calcicoles riches en chaméphytes des sols minces (Salicion pyrenaicae) et d’une autre part encore les pelouses plus mésophiles des sols plus évolués (Primulion intricatae).
L’intégralité de la classe des Elyno – Seslerietea telle que traitée dans ce travail correspond aux habitats naturels codés 36.4 dans CORINE biotopes et E4.4 dans EUNIS (Pelouses calcicoles alpines et subalpines). Ces habitats sont tous rattachés au code Eur 28 6170 (annexe I de la directive Habitats-Faune-Flore).
Ordre 1. Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 225)
Typus ordinis : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 225).
Communautés alpiennes et jurassiennes (extension en Europe moyenne à préciser), non cryophiles et non soumises aux influences bioclimatiques oroméditerranéennes.
Taxons caractéristiques : Scabiosa lucida, Galium anisophyllon, Polygala alpestris, Senecio doronicum subsp. doronicum, Hieracium villosum, Bellidiastrum michelii, Festuca violacea, F. laevigata, Thesium alpinum, Gentiana clusii subsp. clusii, Linum alpinum.
Taxons différentiels par rapport aux autres ordres de la classe : Gentiana lutea, Linum catharticum ; ordre essentiellement différencié négativement par rapport aux deux autres.
Alliance 1.1. Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. in G. Braun-Blanq. 1931 (Commun. Stat. Int. Géobot. Médit. Montpellier 13 : 46)
Typus allianciae : Caricetum ferrugineae Lüdi 1921 (Beitr. Geobot. Landesaufn. Schweiz 9 : 244).
Pelouses neutrobasiphiles montagnardes et subalpines particulièrement élevées et riches en phorbes, établies sur des cônes de déjection, versants et couloirs soumis aux avalanches, éboulis stabilisés ou pieds de paroi. Optimum dans le Jura et les Préalpes du Nord.
Taxons caractéristiques : Centaurea scabiosa subsp. alpestris, Campanula thyrsoides, Anemone narcissiflora, Phleum hirsutum, Paradisea liliastrum, Traunsteinera globosa.
Taxons différentiels : Laserpitium latifolium, Geranium sylvaticum, Astrantia major, Cyanus montanus, Pimpinella major, Crepis pyrenaica, Lathyrus occidentalis subsp. occidentalis, Bupleurum longifolium…
Sous-alliance 1.1.1. Eu-Caricenion ferrugineae
Groupements mésophiles à hygroclinophiles de l’étage subalpin.
Taxon caractéristique : Carex ferruginea.
Taxons différentiels : Luzula sylvatica subsp. sieberi, Trollius europaeus, Pedicularis foliosa, Anemone alpina…
- Caricetum ferrugineae Lüdi 1921 (Lüdi, 1921, liste p. 244, non synthétisé ; F 27-01).
- Pulsatillo alpinae – Anemonetum narcissiflorae Béguin 1972 ; a – senecionetosum doronici Béguin 1972 (Béguin, 1972, tab. 7 : rel. 1-7, synthétisés ici) ; b – rhododendretosum ferruginei Béguin 1972 (Béguin, 1972, tab. 7 : col. 8-12, synthétisées ici) ; c – androsacetosum lacteae Béguin 1972 (Béguin, 1972, tab. 7 : col. 13-16, synthétisées ici) (F 27-02).
- Serratulo tinctoriae – Caricetum sempervirentis Berset 1969 ; a – Senecioni – Caricetum sempervirentis J.-L. Rich. 1977 nom. prov. (Richard, 1977, tab. 7 : col. synth.) ; b – Berset (1969, tab. 12, synthétisé ici) (F 27-03).
Sous-alliance 1.1.2. Calamagrostienion variae (Sill. 1932) Sanz, Van Es & Corriol stat. nov. et suball. hoc loco [basionyme : Calamagrostidion variae Sill. 1932 (Rozpr. České Akad. Věd, Tř. 2, Vědy Mat.-Přír. 41 (1931) /16 : 19)]
Typus nominis hoc loco : Calamagrostietum variae Lüdi 1921 (Beitr. Geobot. Landesaufn. Schweiz 9 : 278).
Groupements mésophiles à mésoxérophiles des étages montagnard et subalpin inférieur, caractérisés par Calamagrostis varia et différenciés par des espèces des Trifolio – Geranietea sanguinei.
Taxon caractéristique : Calamagrostis varia.
Taxons différentiels : Buphthalmum salicifolium, Molinia arundinacea, Polygala chamaebuxus, Ranunculus tuberosus, Knautia maxima, Campanula glomerata, Lilium martagon…
- Peucedano austriaci – Laserpitietum latifolii J.-L. Rich. 1977 (Richard, 1977, tab. 5 : col. synth. ; F 27-04).
- Campanulo thyrsoidis – Laserpitietum latifolii Béguin 1972 ; a – faciès sec (Béguin, 1972, tab. 8 : col. 28-36, synthétisées ici) ; b – faciès humide (Béguin, 1972, tab. 8 : col. 37-45, synthétisées ici) (F 27-05).
- Laserpitio latifolii – Calamagrostietum variae Moor in Moor & Schwarz 1957 nom. inval. (liste in https://www.infoflora.ch/fr/milieux/phytosuisse/IV.2.2.2.3-laserpitio_latifoliae-calamagrostietum_variae.html; F 27-06).
- Calamagrostietum variae Lüdi 1921 ; a – Origano – Calamagrostietum variae Lippert ex Thiele 1978 (Thiele, 1978, tab. 1, synthétisé ici) ; b – Lüdi, 1921, liste p. 278, non synthétisée) (F 27-07).
Alliance 1.2. Agrostio capillaris – Seslerion caeruleae (Béguin 1972) Prunier, Greulich, Béguin, Boissezon, Delarze, Hegg, Klötlzi, Pantke, Steffen, Steiger, Vittoz, Sanz, Van Es & Corriol stat. nov. et all. hoc loco
[basionyme : Agrostio capillaris – Seslerienion caeruleae Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 94)].
Typus nominis hoc loco : Alchemillo conjunctae – Seslerietum caeruleae Prunier, Greulich, Béguin, Boissezon, Delarze, Hegg, Klötlzi, Pantke, Steffen, Steiger & Vittoz ex Sanz, Van Es & Corriol (ce travail, fiche F 27-09).
Pelouses secondaires plus ou moins fermées établies sur des pentes faibles à moyennes, sur des sols bruns assez profonds, mésophiles à mésoxérophiles, acidiclinophiles à neutrobasiphiles, mésotrophiles, des étages montagnard supérieur et subalpin, surtout Jura et des Préalpes du Nord. Alliance essentiellement différenciée par des espèces acidiphiles et prairiales.
Taxons différentiels : Agrostis capillaris, Rhinanthus alectorolophus, Veronica officinalis, Hypericum maculatum, Trifolium montanum, Botrychium lunaria, Gentianella campestris, Koeleria pyramidata, Pimpinella saxifraga…
- Plantagini atratae – Caricetum sempervirentis Béguin 1972 (Béguin, 1972, tab. 12, synthétisé ici ; F 27-08).
- Alchemillo conjunctae – Seslerietum caeruleae Prunier, Greulich, Béguin, Boissezon, Delarze, Hegg, Klötlzi, Pantke, Steffen, Steiger & Vittoz ex Sanz, Van Es & Corriol ; a – Aubert & Luquet (1930, tab. 2 : synthétisé ici) ; b – Béguin (1972, tab. 11 : col. 15-27, sub Seslerio – Caricetum jurassicum euphorbietosum verrucosae, synthétisées ici) ; c – Béguin (1972, tab. 11 : col. 8-14, sub Seslerio – Caricetum jurassicum bupleuretosum ranunculoidis, synthétisées ici) ; d – Béguin (1972, tab. 11 : col. 1-7, sub Seslerio – Caricetum jurassicum prunelletosum vulgaris, synthétisées ici) (F 27-09).
- Dichoropetalo carvifoliae – Caricetum sempervirentis Misset 2017 ; a – Misset (2017, tab. 12 : col. A) ; b – typicum (Misset, 2017, tab. 12 : col B) ; c – drymocallidetosum rupestris Misset 2017 (Misset, 2017, tab. 12 : col. C) (F 27-10).
Alliance 1.3. Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol stat. nov. et all. hoc loco
[basionyme : Drabo aizoidis – Seslerienion caeruleae Béguin & Ritter in Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 91) ; syn. : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland 1998 (Guide des milieux naturels de Suisse : 134) nomen nudum (art. 2b)].
Typus nominis hoc loco : Minuartio vernae – Arenarietum multicaulis Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 91).
Pelouses assez ouvertes, calcicoles, mésoxérophiles à xérophiles, thermophiles à mésocryophiles, oligotrophiles, établies sur des substrats stabilisés (calcaires durs notamment) aux étages montagnard supérieur et subalpin du Jura, Préalpes du Nord, Alpes du Sud.
Taxons caractéristiques : Coronilla vaginalis, Veronica fruticulosa, Dianthus saxicola.
Taxons différentiels : Sempervivum tectorum, Sedum album, Laserpitium siler, Teucrium montanum, Globularia cordifolia, Saxifraga paniculata, Gypsophila repens, Kernera saxatilis, Athamanta cretensis, Campanula cochlearifolia, Thalictrum minus…
- Seslerio caeruleae – Laserpitietum sileris Moor ex Béguin 1972 ; a – stipetosum calamagrostis Béguin 1972 (Béguin, 1972, tab. 8 : col. 21-27, synthétisées ici) ; b – typicum (Béguin, 1972, tab. 8 : col. 16-20, synthétisées ici) ; c – arctostaphyletosum uvae-ursi Béguin 1972 (Béguin, 1972, tab. 8 : col. 1-15, synthétisées ici) ; d – (Moor & Schwarz, 1957, liste p. 40, non synthétisée) (F 27-11).
- Veronico fruticulosae – Agrostietum schleicheri Béguin 1972 ; a – sideritidetosum hyssopifoliae Béguin 1972 (Béguin, 1972, tab. 10 : col. 21-37, synthétisées ici) ; b – asplenietosum rutae-murariae Béguin 1972 (Béguin, 1972, tab. 10 : col. 11-20, synthétisées ici) (F 27-12).
- Teucrio montani – Globularietum cordifoliae J.-L. Rich., Bourgnon & Strub. ex Prunier, Sanz & Van Es hoc loco (Richard, 1977, tab. 8 : col. synth. ; F 27-13).
- Teucrio montani – Festucetum macrophyllae Ritter 1972 (Ritter, 1972, tab. 4 : col. synth. ; F 27-14).
- Valeriano tripteridis – Seslerietum caeruleae Oberd. 1957 (Oberdorfer, 1957, tab. 111 : col. 9, non synthétisée ; sud de l’Allemagne).
- Minuartio vernae – Arenarietum multicaulis Béguin 1972 (Béguin, 1972, tab. 10 : col. 1-10, synthétisées ici ; F 27-15).
- Androsaco villosae – Gentianetum angustifoliae Ritter 1972 (Ritter, 1972, tab. 5 : col. synth. ; F 27-16).
Alliance 1.4. Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 225), incl. Avenion montanae Barbero 1968 (Bull. Soc. Bot. France 115 : 223).
Typus allianciae : Seslerio caeruleae – Caricetum sempervirentis Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 236) nom. corr. hoc loco.
Pelouses primaires fermées à ouvertes, basiphiles, mésophiles à mésoxérophiles, thermophiles à mésocryophiles, oligotrophiles à mésotrophiles, établies sur des substrats plus ou moins perturbés par la cryoturbation (ou gelifluxion), aux étages subalpin et alpin des Alpes.
Taxons caractéristiques : Viola calcarata, Oxytropis lapponica, O. jacquini, O. amethystea, Pedicularis rostratocapitata subsp. helvetica…
Taxons différentiels par rapport à l’Agrostio – Seslerion caeruleae et au Drabo – Seslerion caeruleae : Biscutella laevigata, Oxytropis campestris, O. helvetica, Leontopodium alpinum subsp. nivale, Pedicularis verticillata, Astragalus australis, Silene acaulis, Myosotis alpestris, Dryas octopetala, Sedum atratum…
- Seslerio caeruleae – Caricetum sempervirentis Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 nom. corr. propos. hoc loco; a – Seslerio – Semperviretum Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Braun-Blanquet & Jenny, 1926, tab. VIII, synthétisé ici) ; b – Seslerio – Semperviretum Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Quantin & Nétien, 1940, col. synth. p. 36) ; c – Seslerio – Caricetum sempervirentis Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Ritter, 1972, tab. VI : col. synth.) ; d – Seslerio – Semperviretum Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Barbero & Quézel, 1975, tab. 3, col. synth.) (F 27-17).
- Astragalo leontini – Seslerietum caeruleae J.-L. Rich. ex Corriol, Sanz & Van Es hoc loco (Richard, 1985, tab. 2 : col. synth. ; F 27-18).
- Artemisio glacialis – Festucetum pumilae J.-L. Rich. ex Corriol, Sanz & Van Es hoc loco ; a – Richard (1985, tab. 1 : col. synth.) ; b – Béguin (1972, tab. 10 : col. 38-61, sub Festucetum pumilae Braun-Blanq. 1913, synthétisées ici) (F 27-19).
- Gentiano angustifoliae – Seslerietum caeruleae Bannes-Puygiron 1933 (de Bannes-Puygiron, 1933, tab. p. 150: synthétisé ici ; F 27-20).
- Seslerio caeruleae – Helictotrichetum sedenensis Lippmaa 1933 nom corr. propos. ; a – Lippmaa (1933, tab. 7, sub Association à Sesleria caerulea et Avena montana, synthétisé ici) ; b – Association à Sesleria caerulea et Avena montana Lippmaa (Guinochet, 1938, tab. 36, col synth.) ; c – Association à Sesleria caerulea et Avena montana Lippmaa (Barbero, 1972, tab. 11: col. synth.) (F 27-21).
- Hedysaro boutignyani – Caricetum austroalpinae Van Es & Foucaut (ce travail, tab. 3 ; F 27-22).
Alliance 1.5. Anthoxantho odorati – Plantaginion brutiae Barbero & Quézel 1975 (Ecol. Medit. 1 : 20) nom. mut hoc loco
Lectoypus nominis hoc loco : Anthoxantho – Deschampsietum Archiloque, Borel & Lavagne 1971 (Colloq. Intern. Milieux nat. supraforestiers Mont. Bass. Occ. Médit. : 222).
Pelouses subalpines mésophiles à mésohygrophiles, basiphiles à acidiclinophiles des Alpes du Sud.
Taxons caractéristiques : Astragalus danicus, Carex austroalpina, Tephroseris integrifolia, Fritillaria tubiformis.
Taxons différentiels : Bellardiochloa variegata, Ranunculus aduncus, Potentilla grandiflora, Tulipa sylvestris subsp. australis…
- Alchemillo alpigenae – Bellardiochloetum variegatae Lacoste 1965 nom. corr. ; a – typicum (Lacoste, 1965, tab. 1 : col. 1 à 6, synthétisées ici) ; b – laricetosum deciduae (Lacoste, 1965, tab. 1 : col. 13 à 18, synthétisées ici) ; c – Barbero (1972, tab. 14 : col. synth.) (F 27-23).
- Anthoxantho odorati – Avenelletum flexuosae Archiloque, Borel & Lavagne 1971[1]. ; a – Anthoxantho – Deschampsietum (Barbero & Quézel, 1975, tab. 10 : col. synth.) ; b – Association à Anthoxanthum villosum et Deschampsia montana (Archiloque, Borel & Lavagne, 1971, tab. III ; col. synth.) ; c – Ranunculo – Fritillarietum (Barbero & Quézel 1975, tab. 12, col. synth.) ; d – Gentiano delphinenis – Ranunculetum pyrenaei (Guinochet, 1975, tab. II, synthétisé ici) (F 27-24).
- Tephroserido integrifoliae – Bellardiochloetum variegatae Barbero & Quézel 1975 nom. corr. propos. (Barbero & Quézel, 1975, tab. 11 : col. synth. ; F 27-25).
Ordre 2. ASTRAGALETALIA SEMPERVIRENTIS Barbero 1968 (Bull. Soc. Bot. France 115 : 221)
Lectotypus ordinis hoc loco : Helictotrichion sempervirentis Barbero 1968 (Bull. Soc. Bot. France 115 : 228) nom. mut. hoc loco.
Végétations de pelouses rases à hautes, ouvertes, essentiellement subalpines, développées sur des sols minces et riches en cailloux (rendosols) ou des éboulis stabilisés, issus de roches mères carbonatées, principalement des adrets et soulanes ou en position de crête, rapidement dégagés de la neige par l’ensoleillement, la topographie et le vent, sous influence macroclimatique méditerranéenne ou continentalisée (vallées abritées ou internes), héliophiles, oligotrophiles, basiphiles, xérophiles à mésoxérophiles, à tendance chionophobe.
Du fait de la combinaison de leur altitude élevée, de leur topographie induisant des déneigements précoces et d’un ensoleillement important, elles subissent pendant de longues périodes des contrastes thermiques intenses, entre froid intense et chaleur, générant des phénomènes périglaciaires d’engel-dégel (gélifraction, cryoturbation), qui affectent mécaniquement le sol, contribuant à maintenir un tapis végétal ouvert prenant souvent l’aspect de guirlandes.
Les pelouses relevant des Astragaletalia sont préférentiellement soumises à des influences macroclimatiques méditerranéennes et sont localisées dans les plus hauts massifs du pourtour du bassin méditerranéen, irradiant un peu plus loin dans les secteurs abrités à influence continentale (climat sec à faible nébulosité), tels que les Alpes internes ou les Pyrénées aragonaises. La végétation y est composée d’espèces xérotolérantes résistantes au froid et à souche puissante permettant un ancrage dans des sols en rhexistasie. Les types biologiques dominants y sont les hémicrytpophytes graminoïdes (notamment cespiteuses), suivies des chaméphytes avec notamment un contingent important de chaméphytes en coussinets et de chaméphytes rampantes (Illa et al., 2006). Notons, au niveau des genres botaniques, certains bien représentés comme Oxytropis, Astragalus, Ononis, Anthyllis, Sideritis, Carex, Helictotrichon.
Taxons caractéristiques : Astragalus sempervirens s. l., Anthyllis montana, Bupleurum ranunculoides var. gramineum, Ononis cristata, Paronychia kapela subsp. serpyllifolia[2], Scutellaria alpina, Iberis sempervirens, Androsace vitaliana s. l…
Taxons différentiels par rapport aux autres ordres : Trinia glauca, Koeleria vallesiana s. l., Juniperus communis subsp. nana, Carex caryophyllea, Asperula cynanchica s. l., Helictochloa pratensis s. l., Teucrium chamaedrys, Potentilla tabernaemontani, Astragalus monspessulanus.
Différentielles négatives : Bartsia alpina, Bistorta vivipara.
Alliance 2.1. Helictotrichion sempervirentis Barbero 1968 (Bull. Soc. Bot. France 115 : 228) nom. mut. hoc loco
[syn. : Festucion dimorphae Bonin 1969 (synonyme syntaxonomique inclus), Ononidion cristatae Barbero 1970 prov., Ononidion cenisiae Barbero 1972 nom. ined. (art. 1), Ononidion cenisiae J.-M. Royer 1991]
Lectotypus allianciae hoc loco : Centaureo triumfettii – Helictotrichetum sempervirentis Barbero 1968 (Bull. Soc. Bot. France 115 : 230).
Pelouses primaires ou secondaires, majoritairement ouvertes, basiphiles, mésoxérophiles à xérophiles, thermophiles, étages montagnard et subalpin des Alpes du Sud.
Taxons caractéristiques : Helictotrichon sempervirens, Minuartia rostrata, Astragalus depressus, Galium obliquum, G. corrudifolium, Knautia collina, Dianthus subacaulis, Erysimum rhaeticum, Eryngium spinalba.
Taxons différentiels par rapport au Festucion scopariae : Linum suffruticosum subsp. appressum, Sempervivum arachnoideum, Lavandula angustifolia, Laserpitium gallicum, Gentiana ligustica, Pedicularis gyroflexa, Plantago serpentina…
Sous-alliance 2.1.1. Ononidenion cristatae Gaultier ex Corriol, Sanz & J.-M. Royer suball. nov. hoc loco
[syn. : Ononidenion cristatae Gaultier in Bardat et al. 2004 nom. inval. (art.3i, 5), Eu-Ononidenion cristatae Gaultier 1989 nom inval. (art. 1, 2d, 3h, 12)]
Typus nominis hoc loco : Astragalo sempervirentis – Ononidetum cenisiae Barbero 1972.
Communautés des sols plus ou moins stabilisés, surtout dominées par des chaméphytes suffrutescents aux étages montagnard et subalpin des Alpes du Sud.
Taxons caractéristiques : Gentiana verna subsp. delphinensis, Dianthus godronianus, Plantago argentea, Genista pilosa.
Taxons différentiels par rapport à l’Eu-Helictotrichenion sempervirentis : Cerastium arvense subsp. suffruticosum, Androsace vitaliana s. l., Cirsium acaulon, Sedum ochroleucum, Stipa gallica, Ononis striata, Pilosella officinarum, Bromopsis erecta, Carex caryophyllea, Coronilla minima, Armeria arenaria…
- Pediculari comosae – Ranunculetum geraniifolii Allier 1971 (Allier, 1971, tab. 1 : col. synth. ; F 27-26).
- Astragalo sempervirentis – Ononidetum cenisiae Barbero 1972 ; a – Barbero (1972, tab. 23: col. synth.) ; b – Lejoly (1975, tab. Mo 3 : col. 10-22, sub Landine à Globularia cordifolia et Lavandula angustifolia synthétisées ici) ; c – Archiloque et al. (1971, tab. 2 : col. synth., sub Association à Astragalus aristatus et Ononis cenisia) ; d – Archiloque et al. (1974, tab. 6, sub Pelouse à Astragalus aristatus, synthétisé ici) (F 27-27).
- Astragalo sempervirentis – Onosmetum fastigiatae Lacoste 1964 ; a – carlinetosum acaulis Lacoste 1964 (Lacoste, 1964, tab. 1 : col. 1 à 5, synthétisées ici) ; b – ononidetosum cristatae Lacoste 1964 (Lacoste, 1964, tab. 1 : col. 8 à 13, synthétisées ici) ; c – Lejoly (1975, tab. Su4 : col. 3, 4, 9-14, 25-28, sub Landine à Astragalus sempervirens et Globularia cordifolia faciès à Avena montana, synthétisées ici) ; d – Lejoly (1975, tab. Su4 : col. 15-24, sub Landine à Astragalus sempervirens et Globularia cordifolia faciès à Astragalus sempervirens, synthétisées ici) ; e – Lejoly (1975, tab. Su4 : col. 1, 2, 5-8, sub Landine à Astragalus sempervirens et Globularia cordifolia faciès à Globularia cordifolia, synthétisées ici) ; f – Chaix (1954, tab XI, sub Pelouse à Astragalus aristatus, synthétisé ici) (F 27-28).
Sous-alliance 2.1.2. Eu-Helictotrichenion sempervirentis
[syn. : Ononido cristatae – Helictrotrichenion sempervirentis Gaultier in Bardat et al. 2004 nom. inval. (art. 3i, 5), Ononido cristatae – Helictrotrichenion sempervirentis Gaultier 1989 nom. ined. (art. 1) (synonymes syntaxonomiques inclus)]
Communautés des sols plus ou moins instables dominées par de grandes graminées cespiteuses et centrées sur les Alpes maritimes.
Taxons caractéristiques : Helictotrichon parlatorei, H. setaceum, Leucopoa dimorpha, Cyanus triumfettii s. l., Hieracium tomentosum, Dianthus scaber…
Taxons différentiels par rapport à l’Ononidenion cristatae : Dianthus furcatus, D. saxicola, Iberis sempervirens, Gentiana ligustica, Knautia collina, Nepeta nepetella, Teucrium lucidum, Tolpis staticifolia, Bunium bulbocastanum, Asperula cynanchica subsp. oreophila, Stachys recta.
- 30. Arenario cinereae – Helictotrichetum sempervirentis Barbero 1968 ; a – Barbero (1972, tab. 27 : col. synth.) ; b – Archiloque et al. (1971 ; tab. 1 : col. synth., sub Pelouse à Avena sempervirens et Koeleria alpicola) ; c – Archiloque et al. (1974, tab. 7, sub Pelouse à Avena sempervirens, synthétisé ici) ; d – Lejoly (1975, tab. Mo 3 : col. 1-6, Pelouse à Avena sempervirens et Lavandula angustifolia, synthétisées ici) (F 27-29).
- Centaureo triumfettii – Helictotrichetum sempervirentis Barbero 1968 ; a – Barbero (1972, tab. 26 : col. synth.) ; b – Pelouse à Avena sempervirens et Gregoria vitaliana (Lejoly, 1975, tab. Su 3, synthétisé ici) (F 27-30).
- Hieracio monregalensis – Avenetum montanae Barbero 1972 ; a – rhamnetosum alpinae (Barbero, 1972, tab. 22 : col. synth.) ; b – leontopodietosum alpini (Barbero, 1972, tab. 22 : col. synth.) (F 27-31).
- Festucetum dimorphae Barbero 1968 ; a – Barbero (1968, tab. 3 : col. synth.) ; b – Barbero (1972, tab. 25 : col. synth.) (F 27-32).
Alliance 2.2. Festucion scopariae Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 150)
Typus allianciae : Festucetum scopariae Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 151).
Communautés pyrénéennes.
Taxons caractéristiques : Festuca gautieri subsp. scoparia, Sideritis hyssopifolia s. l., Astragalus sempervirens subsp. catalaunicus, Asperula cynanchica subsp. pyrenaica, Plantago monosperma, Seseli montanum subsp. nanum, Medicago suffruticosa, Viola rupestris, Vicia pyrenaica, Jurinea humilis.
Taxons différentiels par rapport à l’Helictotrichion sempervirentis : Globularia repens, Galium pyrenaicum, G. pumilum (subsp. marchandii), Saxifraga oppositifolia s. l., Clinopodium alpinum, Eryngium bourgatii, Carduus carlinoides, Pedicularis pyrenaica, Erigeron alpinus, Scorzoneroides pyrenaica, Carex ornithopoda, C. rupestris, Taraxacum pyrenaicum, Polygala alpina, Pilosella lactucella, Oxytropis foucaudii, Botrychium lunaria, Sedum atratum, Ononis striata.
Note – La répartition des associations en deux alliances, telle que proposée par Montserrat & Villar (1987), s’avère problématique ; en fonction des différentes classifications automatiques effectuées, certaines associations basculent de l’une à l’autre d’entre elles, ce qui est rendu plus complexe encore avec la mise en évidence et la prise en compte d’associations en limite d’alliance dans notre travail ; sur la base de notre tableau synthétique général trié-ordonné, on tentera une répartition en deux sous-alliances, telles que proposées par Rivas-Martínez et al. (1991) et adoptées par Rivas-Martínez et al. (2011) puis Peyre & Font (2011), mais en attirant l’attention sur la difficulté d’obtenir une césure nette.
Sous-alliance 2.2.1. Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991(Itinera Geobot. 5 : 509)
≡ Saponarion caespitosae P. Monts. & Villar 1987 (Lazaroa 7 : 10).
= Thymelaeion nivalis P. Monts. & Villar 1975 (Doc. Phytosoc. 9-14 : 211) nom. inval. (art. 17) (synonyme syntaxonomique inclus)
Typus suballianciae : Serratulo nudicaulis – Asperuletum pyrenaicae P. Monts. & Villar 1987 (Lazaroa 7 : 15) désigné par Rivas-Martínez et al. (1991 : 509).
Groupe centro-occidental voire oriental, sous influence continentale (versant méridional surtout).
Taxons caractéristiques : Thymelaea tinctoria subsp. nivalis, Saponaria caespitosa, Erodium glandulosum, Onobrychis pyrenaica, Festuca altopyrenaica.
Taxons différentiels par rapport à l‘Eu-Festucenion scopariae : Oxytropis neglecta, Arenaria purpurascens, Carex ornithopoda, Scorzoneroides pyrenaica, Arenaria multicaulis, Silene acaulis, Gypsophila repens, Teucrium pyrenaicum s. l., Asperula hirta, Crepis pygmaea, Brassica repanda, Ranunculus parnassifolius, Festuca pyrenaica, Serratula nudicaulis, Geranium cinereum, Gentiana occidentalis…
- Gentiano occidentalis – Androsacetum villosae Corriol, F. Prud’homme & Fallour-Rubio ; a – trinietosum glaucae; b – typicum (ce travail, tab. 4) (F 27-33).
- Seslerio caeruleae – Globularietum nudicaulis Nègre 1968 (ce travail, tab. 2b ; F 27-34).
- Gypsophilo repentis – Festucetum scopariae Corriol ; a – seslerietosum caeruleae ; b – astragaletosum catalaunici (ce travail, tab. 5) (F 27-35).
- Serratulo nudicaulis – Asperuletum pyrenaicae P. Monts. & Villar 1987 ; a – typicum ; b – polygaletosum alpinae; c – scorzoneretosum aristatae (ce travail, tab. 6) (F 27-36).
- Oxytropido neglectae – Festucetum scopariae Rivas Mart., Báscones, T.E. Díaz, Fern. Gonz. & Loidi 1991 corr. Rivas Mart. et al. 2002 ; a – saponarietosum caespitosae ; b – helianthemetosum oelandici ; c – caricetosum rupestris; d – erodietosum glandulosi; e – arenarietosum purpurascentis ; f – thymelaeetosum nivalis ; g – typicum ; h – ranunculetosum heterocarpi (ce travail, tab. 7) (F 27-37).
Sous-alliance 2.2.2. Eu-Festucenion scopariae
Groupe oriental sous influence macroclimatique de régime méditerranéen.
Taxons caractéristiques : Anthyllis vulneraria subsp. vulnerariodes, Erysimum duriaei subsp. pyrenaicum, Onosma tricerosperma subsp. fastigiata, O. bubanii.
Taxons différentiels par rapport au Saponarienion caespitosae : Carex humilis, Jasione crispa, Oxytropis amethystea[3], Iberis saxatilis, I. sempervirens, Arenaria tetraquetra, Festuca yvesii, Ranunculus ruscinonensis, R. bulbosus, Polygala calcarea, Daphne cneorum, Dianthus hyssopifolius, Carex brevicollis…
- Festuco scopariae – Seslerietum caeruleae Susplugas 1942 (Susplugas, 1942 : 138 et tab. 9 ; ce travail, tab. 2b ; F 27-38).
- Seslerio caeruleae – Scabiosetum graminifoliae Masalles & Vigo in Farrás, Masalles, E. Velasco & Vigo 1981 (Farrás et al., 1981 : 143, tab. 4) (ce travail, tab. 2) (décrite de la Sierra del Cadi).
- Oxytropido amethysteae – Caricetum humilis Carrillo & Soriano in Carrillo & Vigo 1997 ; a – typicum ; b – asteretosum alpini; c – helictotrichetosum sedenensis ; d – caricetosum rosae (ce travail, tab. 8 ; F 27-39).
- Saponario caespitosae – Festucetum scopariae Gruber 1978 corr. Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991 ; a – typicum ; b – lotetosum alpini (ce travail, tab. 2b, Pré-Pyrénées aragonaises).
- Scutellario alpinae – Festucetum scopariae prov. ; a – adonidetosum pyrenaicae ; b – medicaginetosum suffruticosae ; c – ononidetosum cristatae (ce travail, tab. 9 ; F 27-40).
- Festucetum scopariae Braun-Blanq. 1948 ; a – typicum ; b – oxytropidetosum halleri ; c – astragaletosum monspessulani ; d – trifolietosum thalii ; e – iberidetosum saxatilis ; f – seslerietosum caeruleae (ce travail, tab. 10 ; F 27-41).
Alliance 2.3. Gentiano acaulis – Primulion intricatae Corriol & J.-M. Royer all. nov. hoc loc.
Typus nominis hoc loco : Gentiano acaulis – Primuletum intricatae Vigo 1972 (Pirineos 105 : 51).
Communautés pyrénéennes acidiclinophiles.
Taxons différentiels : Gentiana acaulis, Avenella flexuosa, Calluna vulgaris, Carex umbrosa subsp. huetiana, Hypochaeris maculata, Potentilla erecta, Luzula campestris, Koeleria macrantha, Agrostis capillaris, Gymnadenia nigra, Dactylorhiza viridis…
- Gentiano acaulis – Primuletum intricatae Vigo 1972 (Vigo, 1972 : tab. II ; ce travail, tab. 2b ; F 27-42).
Ordre 3. OXYTROPIDO – ELYNETALIA MYOSUROIDIS Oberd. ex Albrecht 1969 (Diss. Bot. 5 : 12)
Lectotypus ordinis hoc loco : Oxytropido – Elynion myosuroidis Braun-Blanq. 1950 (Vegetatio 2 (1) : 24).
Végétations de pelouses psychrophiles subalpines et cryophiles alpines.
Taxons caractéristiques : Arenaria multicaulis, Dryas octopetala, Carex ornithopoda, Leontopodium nivale subsp. alpinum, Astragalus alpinus, avec en plus, pour les Pyrénées, Arenaria purpurascens, Oxytropis neglecta.
Taxons différentiels par rapport aux autres ordres de la classe : Silene acaulis, Soldanella alpina, Selaginella selaginoides, Scorzoneroides pyrenaica, Saxifraga moschata, Primula integrifolia.
Alliance 3.1. Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002 (Itinera Geobot. 15 (1) : 195)
Typus allianciae : Dryado octopetalae – Salicetum pyrenaicae Chouard 1943 (Bull. Soc. Bot. France 90 : 27) désigné par Rivas-Martínez et al. (2002 : 195).
Landines et pelouses subalpines pyrénéennes psychrophiles à cryophiles, souvent riches en chaméphytes rampantes, des sols minces ou caillouteux, humifères, en exposition fraîche et abritée du soleil.
Taxons caractéristiques : Aquilegia pyrenaica, Ranunculus thora, Gentiana occidentalis, ceux du Salicenion pyrenaicae et, en commun avec le Primulion intricatae, Geranium cinereum, Ranunculus gouanii, Horminum pyrenaicum.
Taxons différentiels par rapport aux autres alliances du même ordre : Anemone hepatica, Globularia nudicaulis s. l., Parnassia palustris, Veronica ponae, Pinguicula grandiflora, Valeriana montana, Agrostis schleicheri, Hypericum nummularium, Saxifraga paniculata, Viola biflora, Rhinanthus pumilus, Rhododendron ferrugineum, Briza media, Arctostaphylos uva-ursi, Polystichum lonchitis, Saxifraga umbrosa, Tofieldia calyculata, Erinus alpinus, Tractema umbellata, Arctostaphylos alpina, Helleborus viridis subsp. occidentalis, Campanula rotundifolia agr., Linum catharticum.
Sous-alliance 3.1.1. Eu-Salicenion pyrenaicae
≡ Dryado octopetalae – Salicenion pyrenaicae Peyre & Font 2011 (Pl. Biosyst. 145, suppl. 1 : 226) nom. inval. (art. 5).
Groupe central, différencié positivement.
Taxons caractéristiques : Salix pyrenaica, Betonica alopecuros, Geum pyrenaicum, Thesium pyrenaicum.
Taxons différentiels par rapport au Ranunculo thorae – Seslerienion caeruleae : Dryas octopetala, Helictotrichon sedenense, Geranium cinereum, Ranunculus gouanii, Saxifraga umbrosa, Luzula nutans, Soldanella alpina, Silene acaule, Selaginella selaginoides, Primula integrifolia, Leontopodium nivale subsp. alpinum, Briza media, Viola biflora, Rhododendron ferrugineum, Arctostaphylos alpina.
- Geo pyrenaici – Caricetum sempervirentis Chouard 1943 ; a – Chouard (1943, p. 25) ; b – typicum ; c – anthyllidetosum boscii ; d – Vanden Berghen (1970, tab. II) ; e – thalictretosum macrocarpi ; f – salicetosum reticulatae (ce travail, tab. 11 ; F 27-43).
- Geranio cinerei – Ranunculetum gouanii Gruber 1978 (ce travail, tab. 2c ; F 27-44).
- Dryado octopetalae – Salicetum pyrenaicae Chouard 1943 ; a – Chouard (1943, p. 27) ; b – var. appauvrie ; c – typicum ; d – salicetosum reticulatae; e – caricetosum rupestris ; f – leontopodietosum alpini ; g – Vanden Berghen (1970, tab. I) (ce travail, tab. 12 ; F 27-45).
Sous-alliance 3.1.2. Ranunculo thorae – Seslerienion caeruleae Peyre & Font ex Lazare 2017 (Botanique 3 : 58).
≡ Ranunculo thorae – Seslerienion caeruleae Vigo ex Soriano 2001 (Acta Bot. Barcinon. 47 : 115) nom. inval. (art. 8, 9).
≡ Laserpitio nestleri – Ranunculion thorae Vigo 1979 (Folia Bot. Misc. 1 : 8) nom. inval. (art. 8).
Typus suballianciae : Ranunculo thorae – Seslerietum caeruleae Vigo ex Rivas Mart. et al. 2002 (Itinera Geobot. 15 (1) : 196).
Groupe regroupant des associations de vires calcaires en ombrée, essentiellement différencié négativement. On peut tout au plus indiquer Agrostis schleicheri comme différentielle positive par rapport au Salicenion pyrenaicae.
- Ranunculo thorae – Seslerietum caeruleae Vigo in Rivas Mart. et al. 2002 (ce travail, tab. 2c ; F 27-46).
- Aquilegio pyrenaicae – Seslerietum caeruleae Herrera Gallastegui, Loidi & F. Prieto 1991 (Herrera Gallastegui et al., 1991, tab. 2, p. 350) ; a – typicum inclus horminetosum Herrera Gallastegui, Loidi & F. Prieto 1991[4] ; b – armerietosum pubinervis Herrera Gallastegui, Loidi & F. Prieto 1991 (ce travail, tab. 2c) (décrite des monts Cantabriques et du Pays basque espagnol).
- Armerio orissonensis – Alchemilletum amphisericeae Lazare 2017 (Lazare, 2017, tab. 6 p. 57 ; F 27-47).
- Agrostio schleicheri – Festucetum gautieri Loidi 1982 (Loidi, 1982 : 81 et tab. 11 p. 85 ; ce travail, tab. 2c) (association du Pays basque espagnol).
Alliance 3.2. Primulion intricatae Braun-Blanq. ex Vigo 1972 (Pirineos 105 : 47)
Typus allianciae : Festuco commutatae – Trifolietum thalii Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 179), lectototype désigné par Rivas-Martínez et al. (2011 : 261).
Pelouses subalpines mésophiles, basiphiles, des sols plus ou moins profonds pouvant être quelque peu acidifiés en surface.
Taxons caractéristiques : Trifolium thalii, Primula elatior subsp. intricata.
Taxons différentiels par rapport aux autres alliances du même ordre : Plantago alpina, Taraxacum pyrenaicum/panalpinnum[5], Clinopodium alpinum, Cerastium arvense subsp. strictum, Alchemilla sect. Flabellata, Plicata et Pubescentes, Phleum alpinum s. l., Geum montanum, Pilosella lactucella, Trifolium alpinum, Gnaphalium supinum, Nardus stricta, Erigeron alpinus…
Sous-alliance 3.2.1. Eu-Primulenion intricatae
Pelouses pyrénéennes.
Taxons différentiels : Ranunculus gouanii, Vicia pyrenaica, Medicago suffruticosa, Geranium cinereum, Horminum pyrenaicum, Carex macrostylon, C. pyrenaica, Luzula nutans, Carduus carlinoides, Galium pumilum subsp. marchandii…
- Festuco commutatae – Trifolietum thalii Braun-Blanq. 1948 ; a – typicum ; b – arenarietosum moehringioidis; c – silenetosum acaulis ; d – thalictretosum alpini; e – geranietosum cinerei ; f – caricetosum myosuroidis ; g – euphrasietosum salisburgensis (ce travail, tab. 13 ; F 27-48).
- Primulo intricatae – Horminetum pyrenaici Lazare & Mauric 1986 ; a – typicum ; b – bellardiochloetosum variegatae ; c – plantaginetosum mediae; d – primuletosum integrifoliae (ce travail, tab. 14 ; F 27-49).
- Alchemillo plicatulae – Adonidetum pyrenaicae Braun-Blanq. ex Gruber 1978 (ce travail, tab. 2c ; F 27-50).
- Anthyllido vulnerarioidis – Plantaginetum alpinae Corriol ; a – typicum ; b – scabiosetosum cinereae ; c – silenetosum acaulis (ce travail, tab. 15 ; F 27-51).
Sous-alliance 3.2.2. Festuco violaceae – Trifolienion thalii Van Es & Sanz suball. nov. hoc loco
Typus nominis hoc loco : Festuco violaceae – Trifolietum thalii Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 244).
Pelouses alpiennes.
Taxons différentiels : Festuca violacea, Viola calcarata, Scabiosa lucida, Hieracium villosum, Pedicularis verticillata, Crepis aurea, Mutellina adonidifolia, Primula veris.
- Festuco violaceae – Trifolietum thalii Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 ; a – Festuco violaceae – Trifolietum thalii Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Braun-Blanq. & Jenny 1926, tab. 9, synthétisé ici) ; b – Festuco violaceae – Trifolietum thalii sous-ass. à Plantago serpentina (Guinochet, 1938, tab. 43, col. synth.) ; c – Festuco violaceae – Trifolietum thalii sous-ass. à Plantago serpentina (Barbero, 1972, tab. 13, col. synth.) (F 27-52).
Alliance 3.3. Oxytropido – Elynion myosuroidis Braun-Blanq. 1950 (Vegetatio 2 (1) : 24)
[syn. : Elynion myosuroidis Gams 1936 nom. inval. (art. 2b), Elynion medioeuropaeum Braun-Blanq. 1948 nom. illeg. (art. 34a).
Lectotypus allianciae hoc loco : Oxytropido halleri – Kobresietum myosuroidis (Braun-Blanq. 1948) Rivas-Mart. et al. 2011 (Itinera Geobot. 18 (1) : 260).
Pelouses primaires alpines, cryophiles, xérophiles, calcicoles, écorchées, des reliefs exposés aux vents.
Taxons caractéristiques : Carex myosuroides, C. rupestris, C. capillaris, C. curvula subsp. rosae, Sedum atratum, Erigeron uniflorus, Antennaria carpatica subsp. helvetica, Oxytropis campestris, Astragalus australis.
Taxons différentiels par rapport aux autres alliances du même ordre : Agrostis rupestris, Luzula spicata aggr., Minuartia sedoides, Carex parviflora, C. ericetorum var. approximata.
Sous-alliance 3.3.1. Eu-Oxytropido – Elynenion myosuroidis
Communautés pyrénéo-cantabriques.
Taxons caractéristiques : Oxytropis halleri, O. foucaudii.
Taxons différentiels : Festuca glacialis, Hornungia alpina, Armeria alpina, Valeriana apula, Potentilla nivalis, Ranunculus alpestris, Poa minor, Artemisia umbelliformis, Juncus trifidus, Poa molinerii, Gentiana nivalis, Thalictrum alpinum, Artemisia eriantha, Cerastium alpinum, Polygala alpina.
- Salici pyrenaicae – Bistortetum viviparae Corriol ; a – typicum ; b – soldanelletosum alpinae (ce travail, tab. 16 ; F 27-53).
- Oxytropido neglectae – Kobresietum myosuroidies Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1984 (Rivas-Martínez et al., 1984, tab. 1, p. 70 ; ce travail tab. 2c) (décrit des Picos de Europa).
- Oxytropido foucaudii – Kobresietum myosuroidis Braun-Blanq. 1948 ; a – association à Elyna spicata et Oxytropis lazica Chouard 1943 (Chouard, 1943, rel. synth. p. 3) ; b – typicum ; c – euphrasietosum minimae ; d – antennarietosum carpaticae (ce travail, tab. 17) ; e – Agrostio alpinae – Caricetum rupestris Lazare 2017 (Lazare, 2017 : 54 et tab. 5 p. 55 ; ce travail, tab. 2c) (F 27-54).
- Oxytropido campestris – Caricetum rupestris Corriol ; a – typicum ; b – potentilletosum nivalis (ce travail, tab. 18 ; F 27-55).
- Oxytropido halleri – Kobresietum myosuroidis (Braun-Blanq. 1948) Rivas Mart. et al. 2011 (Braun-Blanquet, 1948, tab. 20) (ce travail, tab. 2c ; F 27-56).
- Elyno myosuroidis – Salicetum retusae Nègre 1968 ; a – typicum ; b – plantaginetosum monospermae (ce travail, tab. 19 ; F 27-57).
Sous-alliance 3.3.2. Festuco pumilae – Elynenion myosuroidis suball. nov. hoc loco
Typus nominis hoc loco : Carici atratae – Kobresietum myosuroidis Braun-Blanq. 1913 em. Rivas Mart. & Géhu 1978 (Doc. Phytosoc., N.S., III : 396).
= « Elyneta » Gams 1936 p.p.
Communautés alpiennes mésocryophiles à cryophiles établies sur des substrats plus ou moins humifères.
Taxons caractéristiques : Pachypleurum mutellinoides, Gentiana brachyphylla et, en commun avec le Caricenion firmae, Festuca pumila, Carex atrata.
Taxons différentiels : Pedicularis verticillata, Veronica bellidioides, Helictochloa versicolor subsp. versicolor, Festuca halleri, Gentianella campestris, Androsace obtusifolia, Potentilla aurea, Anemone vernalis, Sempervivum montanum et, en commun avec le Caricenion firmae, Salix serpyllifolia, Phyteuma globulariifolium subsp. pedemontanum.
- Carici atratae – Kobresietum myosuroidis Braun-Blanq. 1913 em. Rivas Mart. & Géhu 1978 ; a – Carici atratae – Kobresietum (Delpech, 2003, tab. 13 : col. synth) ; b – Elynetum Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Braun-Blanquet & Jenny, 1926, tab. 10, synthétisé ici) ; c – Elynetum (Braun-Blanq., 1913, tab. p. 101 : col. 1-7, synthétisées ici) (F 27-58).
- Kobresio myosuroidis – Caricetum rosae Erschbamer 1992 ; a – Elynetum Friedel 1956 (Friedel, 1956, tab. F, synthétisé ici) ; b – Groupement à Sesleria varia et Carex rosae (Gensac, 1977, tab. 10, synthétisé ici) ; c – Kobresio myosuroidis – Caricetum rosae (Friedel 1956) Erschbamer 1992 (Gensac, 1977, tab. 9, synthétisé ici) ; d – Kobresio myosuroidis – Caricetum rosae (Friedel 1956) Erschbamer 1992 (Erschbamer 1992, tab. 1, synthétisé ici) (F 27-59).
- Festucetum pumilae Gams 1927 (Gams, 1927, tab. p. 614, synthétisé ici) (F 27-60).
Sous-alliance 3.3.3. Caricenion firmae (Gams 1936) Theurillat in Theurillat, Aeschimann, P. Küpfer & Spichiger 1995 (Colloq. Phytosociol. XXIII : 217)
[corresp. syntax. : Caricion firmae Gams 1936 (Abh. K. K. Zool.-Bot. Ges. Wien : 57)]
Typus suballianciae : Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 227).
Communautés alpiennes cryophiles des sols plus ou moins squelettiques.
Taxons caractéristiques : Carex firma, Saxifraga caesia, Chamorchis alpina et, en commun avec le Festuco pumilae – Elynenion, Festuca pumila, Carex atrata.
Taxons différentiels : Pedicularis rosea subsp. allionii, Leucanthemum coronopifolium, Carex mucronata et, en commun avec le Festuco pumilae – Elynenion, Salix serpyllifolia, Phyteuma globulariifolium subsp. pedemontanum.
- Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 ; a – Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 typicum (Braun-Blanquet & Jenny, 1926, tab. 7, synthétisé ici) ; b – Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 Dryas Fazies (Braun-Blanquet & Jenny, 1926, tab. 7, synthétisé ici) ; c – Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Barbero, 1972, tab. 12 : col. synth.) ; d – Elynetum Barbero 1972 (Barbero, 1972, tab. 15 : col.synth.) (F 27-61).
- Caricetum rupestris Pignatti 1985 (Pignatti, 1985, tab. 1, synth) (association des Dolomites, absente de France).
- Dryadetum octopetalae Rübel 1911 (Rübel, 1911, liste p. 135 non synthétisé) (F 27-62).
FICHE N° 27-01
Association : Caricetum ferrugineae Lüdi 1921 (Beitr. Geobot. Landesaufn. Schweiz 9 : 244).
Synonyme : Ass. à Carex ferruginea et Festuca pulchella G. Braun-Blanq. & Braun-Blanq. 1931.
Unités supérieures : Eu-Caricenion ferrugineae, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. (lectotypus nominis hoc loco) in Lüdi (1921, Beitr. Geobot. Landesaufn. Schweiz 9 : 245-246).
Physionomie : pelouse / prairie dense et assez élevée riche en phorbes et structurée par les feuilles fines de Carex ferruginea.
Combinaison caractéristique d’espèces : Carex ferruginea, Phleum hirsutum, Astragalus frigidus, Hedysarum hedysaroides, Astrantia major, Anemone narcissiflora, Campanula thyrsoides, Leucopoa pulchella subsp. pulchella, Pedicularis foliosa, Cerinthe glabra subsp. glabra, Globularia nudicaulis, Polygala alpestris, Mutellina adonidifolia.
Synécologie : pelouse hygroclinophile des ubacs de l’étage subalpin sur des sols humifères riches en bases, établie sur des éboulis stabilisés, pieds de paroi, couloirs ou versants pouvant être soumis aux dynamiques du manteau neigeux (reptation, avalanche…), subissant donc un déneigement assez tardif pour l’étage considéré et présentant souvent des suintements ou des horizons hydromorphes.
Variations : transition vers le Laserpitio latifolii – Calamagrostietum variae marquée par la présence de Calamagrostis varia aux altitudes inférieures (obs. récente CBNA dans le Haut-Giffre).
Synchorologie :
– territoire d’observation : Préalpes du Nord en Haute-Savoie surtout, un peu moins caracactéristique dans les Préalpes de Savoie et de l’Isère (Chartreuse, Vercors) où Astragalus frigidus et Leucopoa pulchella subsp. pulchella font défaut ; présent mais moins caractéristique également dans les Alpes internes de Savoie (Haute-Tarentaise, Haute-Maurienne) où il trouve probablement une de ses limites d’aire méridionales.
Axes à développer : étude des variations dans les Alpes internes de Savoie ; cité dans le Jura (Béguin, 1972), sa présence dans ce massif serait encore à confirmer car un certain nombre de caractéristiques d’association sont absentes ou restent à confirmer dans ce massif : Astragalus frigidus, Leucopoa pulchella subsp. pulchella, Hedysarum hedysaroides, Cerinthe glabra subsp. glabra, Phleum hirsutum, Globularia nudicaulis, Mutellina adonidifolia.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Boissier J.-M., 2005 ; Braun-Blanquet J., 1948 ; Lüdi W., 1921 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.
FICHE N° 27-02
Association : Pulsatillo alpinae – Anemonetum narcissiflorae Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 66).
Synonymes : -.
Unités supérieures : Eu-Caricenion ferrugineae, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 6 (lectotypus nominis hoc loco) du tab. 7 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 66).
Physionomie : pelouse-ourlet dense, moins élevée que la précédente association, riche en Ranunculacées, notamment Anemone narcissiflora, Pulsatilla alpina, Trollius europaeus… ; Carex ferruginea y est peu représenté et est généralement remplacé par C. sempervirens et Sesleria caerulea ; photo 1.
Combinaison caractéristique d’espèces : Anemone alpina, A. narcissiflora, Trollius europaeus, Ranunculus carinthiacus, R. montanus, R. thora, Luzula sylvatica subsp. sieberi, Gymnadenia conopsea, Bellidiastrum michelii, Homogyne alpina, Bartsia alpina, Bistorta vivipara, Carex sempervirens, C. ferruginea, Sesleria caerulea, Dryas octopetala, Astrantia major, Geranium sylvaticum, Centaurea scabiosa subsp. alpestris.
Synécologie : pelouse-ourlet mésophile des ubacs de l’étage subalpin à enneigement légèrement plus long que l’association précédente (jusqu’à sept mois), établi sur des sols normalement riches en bases, mais pouvant être légèrement acidifiés en surface, ce que traduit la présence de Homogyne alpina, Luzula subsp. sieberi ou Vaccinium myrtillus.
Variations :
– typicum [syn. : senecionetosum doronici Béguin 1972], différencié par Senecio doronicum subsp. doronicum, Calamagrostis varia, Veratrum album subsp. lobelianum, Achillea millefolium, Libanotis pyrenaica, Valeriana montana et lié à des substrats plus nettement marneux ;
– rhododendretosum ferruginei Béguin 1972, différencié par Vaccinium vitis-idaea, Rhododendron ferrugineum, Agrostis capillaris, Vaccinium uliginosum subsp. microphyllum, localisé sur des calcaires plus compacts aux horizons bien décalcifiés ; lectotypus nominis hoc loco : rel. 11 du tab. 7 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 66) ;
– androsacetosum lacteae Béguin 1972, différenciée par Androsace lactea, Gentiana clusii, Ranunculus alpestris, Noccaea montana subsp. montana, Saxifraga paniculata, lectotypus nominis hoc loco : rel. 14 du tab. 7 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 66) ;
– transition vers l’Alchemillo conjunctae – Seslerietum caeruleae : pelouse plus basse observée dans les Préalpes sur les versants ouest notamment (Chartreuse et nord du Vercors) : beaucoup plus appauvrie en grandes herbes telles qu’Astrantia major, Geranium sylvaticum, Centaurea scabiosa subsp. alpestris, Anemone narcissiflora, dominance plus nette de Globularia nudicaulis, Polygala calcarea, Sesleria caerulea, Noccaea montana subsp. villarsiana.
Synchorologie :
– territoire d’observation : Jura ; Préalpes du Nord en Haute-Savoie, Savoie et Isère (surtout Chartreuse et nord du Vercors) ; syntaxon moins caractéristique et probablement en limite d’aire dans les Alpes internes du Nord ;
– sous-associations ou variantes géographiques : la subass. androsacetosum lacteae n’est connue que du Jura ; variante géographique des Préalpes du Nord à Globularia nudicaulis, Hedysarum hedysaroides, Mutellina adonidifolia, Noccaea montana subsp. villarsiana.
Axes à développer : variante des Alpes internes du Nord à étudier (transition vers l’Agrostio – Seslerion).
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C. 1972 ; Bensettiti F. et al., 2002 ; Boissier J.-M., 2005 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.

FICHE N° 27-03
Association : Serratulo tinctoriae – Caricetum sempervirentis Berset 1969 (Comm. Sta. Int. Géobot. Médit. Alp. 183 : 41).
Synonyme : Senecioni – Caricetum sempervirentis J.-L. Rich. 1977 (Bull. Soc. Fribourg. Sci. Nat. 66 (1) : 25) prov. (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Caricenion ferrugineae, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 9 (lectotypus nominis hoc loco) du tab. 12 in Berset (1969, Bull. Soc. Fribourg. Sci. Nat. 66 (1) : 41).
Physionomie : pelouse fermée moyennement élevée, dominée par Carex sempervirens auquel s’associent de nombreuses dicotylédones élevées.
Combinaison caractéristique d’espèces : Serratula tinctoria, Carex sempervirens, C. montana, Sesleria caerulea, Potentilla erecta, Anthoxanthum odoratum, Genista sagittalis, Prunella grandiflora, Hieracium gpe lachenali, Laserpitium latifolium, L. siler, Anemone narcissiflora, Geranium sylvaticum, Traunsteinera globosa, Astrantia major, Paradisea liliastrum, Crepis pyrenaica, Lathyrus occidentalis, Phleum hirsutum, Pedicularis foliosa, P. ascendens, Scabiosa lucida.
Synécologie : pelouse d’adret de l’étage subalpin, neutro-basiphile à acidiclinophile, mésophile, établie sur des sols assez profonds à bonne réserve hydrique, riches en bases, issus de substrats préférentiellement marneux ou de calcaires massifs en situations de pied de versant, légèrement décalcifiés en surface.
Variations : transition vers l’Alchemillo conjunctae – Seslerietum caeruleae ou le Seslerio – Caricetum sempervirentis → pelouse plus rase enrichie en espèces mésoxérophiles telles que Helianthemum nummularium, Senecio doronicum, Hieracium villosum, Aster alpinus, Bupleurum ranunculoides, pouvant correspondre au Senecioni – Caricetum sempervirentis J.-L. Rich. 1977 prov. ayant peut-être valeur de sous-association.
Synchorologie :
– territoire d’observation : Préalpes du Nord en Haute-Savoie (Prunier et al., 2019), Bauges et Aravis (A. Mikolajczak comm. pers.) mais présence fort probable jusqu’en Isère (Chartreuse) ;
– sous-associations ou variantes géographiques : -.
Axes à développer :
– présence à confirmer en Chartreuse et éventuellement nord du Vercors ;
– préciser les sous-associations et les liens avec le Senecioni – Caricetum sempervirentis J.-L. Rich. 1977 prov.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Berset J., 1969 ; Prunier P. et al., 2019 ; Richard J.-L., 1977.
FICHE N° 27-04
Association : Peucedano austriaci – Laserpitietum latifolii J.-L. Rich. 1977 (Bull. Soc. Fribourg. Sci. Nat. 66 (1) : 17).
Synonymes : -.
Unités supérieures : Calamagrostienion variae (Sill. 1932) Sanz, Van Es & Corriol, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 ; syntaxon intermédiaire entre le Calamagrostienion variae et le Caricenion ferrugineae.
Type nomenclatural : rel 9 (lectotypus nominis hoc loco) du tab. 5 in Richard (1977, Bull. Soc. Fribourg. Sci. Nat. 66 (1) : 17).
Physionomie : pelouse-ourlet dense et élevé, d’une grande luxuriance et très diversifié, dominé par de grandes ombellifères auxquelles s’associe souvent Calamagrostis varia ; photo 2.
Combinaison caractéristique d’espèces : Calamagrostis varia, Pteroselinum austriacum, Laserpitium latifolium, L. siler, Astrantia major, Pimpinella major, Anemone narcissiflora, Geranium sylvaticum, Traunsteinera globosa, Crepis pyrenaica, Lathyrus occidentalis, Phleum hirsutum, Pedicularis foliosa, Buphthalmum salicifolium, Origanum vulgare, Molinia caerulea subsp. arundinacea, Trifolium medium, Scabiosa lucida, Gentiana lutea, Stachys recta, Lilium martagon, Digitalis grandiflora, Carex flacca, C. sempervirens, Campanula glomerata, Sesleria caerulea.
Synécologie : pelouse d’adret des étages montagnard supérieur et subalpin inférieur, basiphile, mésophile à mésoxérophile, établie sur des sols caillouteux comprenant une matrice fine toujours alimentée en éléments carbonatés (éboulis stabilisés, cônes de déjection…). Établie sur des pentes faibles à moyennes exposées aux dynamiques du manteau neigeux (reptation, avalanche de fonte…), provoquant un déneigement assez tardif pour l’étage et l’exposition considérés.
Variations : -.
Synchorologie :
– territoire d’observation : Préalpes du Nord (Prunier et al., 2019) ; obs. récentes dans le Haut-Giffre (CBNA) et les Bauges, mais dans ce dernier massif sans Pteroselinum austriacum (A. Mikolajczak, comm. pers.) ; présence possible en Chartreuse ;
– sous-associations ou variantes géographiques : -.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Berset J., 1969 ; Prunier P. et al., 2019 ; Richard J.-L., 1977.

FICHE N° 27-05
Association : Campanulo thyrsoidis – Laserpitietum latifolii Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 71).
Synonymes : -.
Unités supérieures : Calamagrostienion variae (Sill. 1932) Sanz, Van Es & Corriol, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 36 (lectotypus nominis hoc loco) du tab. 8 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 77).
Physionomie : pelouse dense et assez élevée, luxuriante, dominée par de grandes ombellifères auxquelles s’associent Sesleria caerulea et Carex sempervirens.
Combinaison caractéristique d’espèces : Laserpitium latifolium, L. siler, Libanotis pyrenaica, Calamagrostis varia, Origanum vulgare, Bupleurum falcatum, B. longifolium, Campanula thyrsoides, Heracleum sphondylium subsp. elegans, Astrantia major, Pimpinella major, Lathyrus occidentalis, Senecio doronicum, Anemone alpina, A. narcissiflora, Crepis pyrenaica, Cyanus montanus, Traunsteinera globosa, Eryngium alpinum, Paradisea liliastrum.
Synécologie : pelouse-ourlet d’adret des étages montagnard supérieur et subalpin inférieur, basiphile à neutrophile (très légère acidification de surface), mésophile à mésoxérophile, établi sur des bas de versants et pentes moyennes sur substrat marno-calcaire donnant des sols assez profonds, plus ou moins caillouteux, comprenant une matrice fine riche en argile.
Variations : deux variations relevées par Béguin ayant probablement valeur de sous-associations :
– « faciès sec », différencié par Anthericum ramosum, Teucrium chamaedrys, Helianthemum oelandicum, Globularia cordifolia, Geranium sanguineum, Vicia sepium faisant transition vers le Laserpitio – Seslerietum variae Moor in Moor & Schwarz 1957 ou vers le Calamagrostietum variae Lüdi 1921 ;
– « faciès humide », différencié par Anemone narcissiflora, Luzula sylvatica subsp. sieberi, Eryngium alpinum, Bupleurum longifolium, Bellidiastrum michelii et faisant transition vers le Pulsatillo alpinae – Anemonetum narcissiflorae Béguin 1972.
Synchorologie :
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al. 2014 ; Prunier et al., 2019) ;
– sous-associations ou variantes géographiques : -.
Axes à développer : statuer sur les deux variations citées.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Bensettiti F. et al. 2002 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019.
FICHE N° 27-06
Association : Laserpitio latifolii – Calamagrostietum variae Moor in Moor & Schwarz 1957 (Beitr. Geobot. Landesaufn. Schweiz 37 : 41) nom inval. (art. 7).
Synonymes : -.
Unités supérieures : Calamagrostienion variae (Sill. 1932) Sanz, Van Es & Corriol, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : non lectotypifiable, l’auteur appuie sa description sur un texte listant les espèces caractéristiques, sans localité ni quantification.
Physionomie : pelouse-ourlet dense et élevé dominé par Calamagrostis varia auquel s’associent de grandes herbes et en particulier Laserpitium latifolium ; photo 3.
Combinaison caractéristique d’espèces : Calamagrostis varia, Laserpitium latifolium, Chaerophyllum hirsutum, Carex flacca, Tofieldia calyculata, Dactylorhiza maculata, Gymnadenia conopsea, G. odoratissima, Angelica sylvestris, Phyteuma spicatum, Ph. orbiculare, Molinia caerulea subsp. arundinacea, Cyanus montanus, Knautia maxima, Anemone narcissiflora, Pimpinella major, Astrantia major, Bellidiastrum michelii, Anthericum ramosum, Gentiana lutea, Buphthalmum salicifolium, Carex sempervirens, Sesleria caerulea.
Synécologie : pelouse-ourlet d’ubac centré sur l’étage montagnard (subalpin inférieur), basiphile à neutrobasiphile, mésophile à hygroclinophile, établi sur des versants préférentiellement marneux, particulièrement suintants et assez instables (glissements de terrain, avalanches, talus de pistes forestières…).
Variations : -.
Synchorologie :
– territoire d’observation : Jura, Préalpes du Nord jusqu’au Dévoluy isèrois (Villaret, 2007 ; Mikolajczak et al. 2014) ;
– sous-associations ou variantes géographiques : -.
Axes à développer : identifier un lectotype parmi le matériel original, sinon désigner un néotype acceptable dans la région princeps (Béguin, 1972 : 71 cite des relevés de Müller que nous n’avons pas identifiés).
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Mikolajczak A. et al., 2014 ; Moor M. & Schwarz U., 1957 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.

FICHE N° 27-07
Association : Calamagrostietum variae Lüdi 1921 (Beitr. Geobot. Landesaufn. Schweiz 9 : 278).
Synonyme : Origano – Calamagrostietum variae Lippert ex Thiele 1978 (syn. syntaxonomique inclus).
Unités supérieures : Calamagrostienion variae (Sill. 1932) Sanz, Van Es & Corriol, Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. 1931, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. (lectotypus nominis hoc loco) in Lüdi (1921, Beitr. Geobot. Landesaufn. Schweiz 9 : 279).
Physionomie : pelouse-ourlet dense largement dominé par Calamagrostis varia auquel s’associent différentes espèces de grandes herbes.
Combinaison caractéristique d’espèces : Calamagrostis varia, Origanum vulgare, Buphthalmum salicifolium, Carex flacca, C. sempervirens, Leontodon hispidus, Hippocrepis comosa, Brachypodium rupestre, Anthericum ramosum, Campanula glomerata, Polygala chamaebuxus, Sesleria caerulea, Scabiosa lucida, Bellidiastrum michelii, Tolpis staticifolia, Teucrium chamaedrys, Potentilla erecta.
Synécologie : pelouse-ourlet d’adret de l’étage montagnard, basiphile à neutrobasiphile, mésophile à mésoxérophile, thermophile, établi sur des versants parcourus par des avalanches ou sur des talus, clairières et coupes forestières ensoleillées sur substrat riche en particules fines plutôt argileuses.
Variations : -.
Synchorologie :
– territoire d’observation : Préalpes du Nord, observé en Haute-Savoie (Prunier et al., 2019 ; obs. récentes CBNA dans le Haut-Giffre), groupements très proches cités en Isère (Villaret, 2007).
Axes à développer : répartition à préciser dans les Préalpes de Savoie et d’Isère ; à confirmer dans le Jura où il est fortement probable et étudier sa présence dans les Alpes internes du Nord (Haute-Tarentaise et Haute-Maurienne notamment), ainsi que dans les Alpes du Sud.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Lippert W., 1966 ; Lüdi W., 1921 ; Prunier P. et al., 2019 ; Thiele K., 1978 ; Villaret J.-C., 2007.
FICHE N° 27-08
Association : Plantagini atratae – Caricetum sempervirentis Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 99).
Synonymes : -.
Unités supérieures : Agrostio capillaris – Seslerion caeruleae (Béguin 1972) Prunier et al., Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 2 (lectotypus nominis hoc loco) du tab. 12 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 104).
Physionomie : pelouse généralement fermée et dominée par Carex sempervirens.
Combinaison caractéristique d’espèces : Carex sempervirens, Phleum rhaeticum, Plantago media, P. atrata, Trifolium montanum, Agrostis capillaris, Pimpinella saxifraga, Festuca rubra, Dactylorhiza sambucina, Gnaphalium sylvaticum, Hypericum maculatum, Pilosella lactucella, Luzula campestris, Antennaria dioica, Gentiana lutea, Scabiosa lucida, Galium anisophyllon, Alchemilla conjuncta, Polygala alpestris, Potentilla erecta, P. aurea.
Synécologie : pelouse secondaire mésophile, mésotrophile, des étages montagnards supérieurs et subalpins, neutrobasiphile à acidiclinophile, établie sur des pentes faibles à moyennes sur des sols brunifiés, riches en particules fines et décalcifiés en surface.
Variations : variante évoluant vers des prairies plus eutrophiles du Veratro lobeliani – Cirsietum eriophori Béguin 1972 ou du Poion alpinae (notamment le Luzulo multiflorae – Koelerietum pyramidatae Béguin 1972 dans le Jura), différenciée par Alchemilla monticola, Cirsium eriophorum, Veratrum album, Phleum rhaeticum, Dactylis glomerata, Lathyrus pratensis…
Synchorologie :
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al., 2014) ; Préalpes du Nord (Mikolajczak et al., 2014).
Axes à développer : étudier et préciser la répartition de cette pelouse dans les Préalpes du Nord et ses sous-associations.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.
FICHE N° 27-09
Association : Alchemillo conjunctae – Seslerietum caeruleae Prunier, Greulich, Béguin, Boissezon, Delarze, Hegg, Klötlzi, Pantke, Steffen, Steiger & Vittoz ex Sanz, Van Es & Corriol ass. nov. loc loco.
Synonymes :
– « Peuplements à Carex sempervirens du mont Tendre » in Aubert & Luquet (1930 : 518) nom inval. (art. 3c,d) ;
– Seslerio – Caricetum jurassicum (Braun-Blanq. & H. Jenny 1926) Béguin 1972 nom. illeg. (art. 34a).
Unités supérieures : Agrostio capillaris – Seslerion caeruleae (Béguin 1972) Prunier et al., Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 9 (typus nominis hoc loco) du tab. II in Aubert & Luquet (1930, Rev. Géogr. Alpine 18 (3) : 518).
Physionomie : pelouse généralement fermée et dominée par Carex sempervirens et Sesleria caerulea, vivement colorée de petites dicotylédones ; photo 4.
Combinaison caractéristique d’espèces : Alchemilla conjuncta, Carex sempervirens, Sesleria caerulea, Aster alpinus, Anthyllis vulneraria subsp. alpestris, Helianthemum nummularium, Globularia cordifolia, Carlina acaulis subsp. caulescens, Gentianella campestris, Soldanella alpina, Dactylorhiza viridis, Bellidiastrum michelii, Hypericum richeri, Leucanthemum adustum, Agrostis capillaris, Gymnadenia nigra, Rhinanthus alectorolophus.
Synécologie : pelouse mésophile à mésoxérophile, basiphile, oligotrophile à mésotrophile, des étages montagnard supérieur et subalpin, établie sur des pentes moyennes à fortes ou sur des zones d’affleurements calcaires, en situation chaude et sur des sols peu épais, rocailleux, alcalins.
Variations :
– typicum : optimum pour Alchemilla conjuncta ;
– prunelletosum vulgaris Béguin 1972 : pelouse secondaire de l’étage montagnard supérieur en lien avec des hêtraies, différenciée par Prunella vulgaris, Euphrasia officinalis, Selaginella selaginoides, Luzula spicata, Ranunculus acris, Bellis perennis, lectotypus nominis hoc loco : rel. 1 du tab. 11 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 94) ; il existe une variante de cette sous-association évoluant vers des prairies plus eutrophiles du Veratro lobeliani – Cirsietum eriophori Béguin 1972 différenciée par Cirsium eriophorum, Dactylis glomerata, Lathyrus pratensis, Digitalis grandiflora… ;
– bupleuretosum ranunculoidis Béguin 1972 : pelouse subalpine plus psychrophile que le type et les deux autres sous-associations, probablement primaire, différenciée par Bupleurum ranunculoides, Bistorta vivipara, Luzula alpina, Ranunculus thora, Trollius europaeus, Scabiosa lucida auxquels on peut ajouter Ranunculus alpestris, Salix retusa, S. reticulata ou encore Pyrola minor (A. Mikolajczak, comm. pers.), lectotypus nominis hoc loco : rel. 10 du tab. 11 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 97) ;
– euphorbietosum verrucosae Béguin 1972 : pelouse secondaire de l’étage montagnard inférieur à Euphorbia verrucosa, Aconitum anthora, Koeleria pyramidata, Bromus erectus, de transition vers les Arrhenatheretea elatioris, lectotypus nominis hoc loco : rel. 15 du tab. 11 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 98).
Synchorologie :
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al., 2014), Préalpes du Nord (obs. CBNA). Les pelouses du « Seslerio-Caricetum auct. » des Préalpes du Nord, dans leur structure, leur composition floristique mais également leur contexte mésoclimatique, présentent bien plus d’affinités avec ce groupement décrit du Jura qu’avec le Seslerio – Caricetum de Braun-Blanquet décrit des Grandes Alpes, qui trouve son optimum à l’étage alpin et qui est lié à des versants exposés à la solifluxion sur des substratums généralement autres que des calcaires durs ;
– sous-associations ou variantes géographiques : sous-associations connues du Jura, à préciser pour les Préalpes du Nord.
Axes à développer : étudier et préciser la répartition de cette pelouse dans les Préalpes du Nord et ses sous-associations.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Aubert S. & Luquet A., 1930 ; Béguin C., 1972 ; Braun-Blanquet J. & Jenny H., 1926 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.

FICHE N° 27-10
Association : Dichoropetalo carvifoliae – Caricetum sempervirentis Misset 2017 (Psychodrômia, Bull. Soc. Bot. Drôme 2 : 60)
Synonymes : -.
Unités supérieures : Agrostio capillaris – Seslerion caeruleae (Béguin 1972) Prunier et al., Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 13 du tab. 12 in Misset (2017, Psychodrômia, Bull. Soc. Bot. Drôme 2 : 84).
Physionomie : pelouse très fermée et diversifiée, marquée par les floraisons de Hypericum richeri subsp. r., Carduus defloratus, Drymocallis rupestris, Dianthus hyssopifolius, Orchis mascula ; Globularia nudicaulis peut être très recouvrant, comme Festuca laevigata subsp. l., Carex sempervirens ou Agrostis capillaris.
Combinaison caractéristique d’espèces : Carex sempervirens, Festuca laevigata subsp. l., Agrostis capillaris, Hypericum richeri subsp. r., Trifolium montanum, Carduus defloratus, Plantago atrata subsp. a., Nardus stricta, Dichoropetalum carvifolia, Globularia nudicaulis.
Synécologie : pelouse acidiclinophile des étages montagnard et subalpin du Vercors occupant des pentes faibles ou nulles aux expositions surtout ouest et sud-ouest, sur sols modérément secs, constitués d’une matrice argileuse associée ou non à des cailloux calcaires, établis sur des argiles de décarbonatation ou des marnes.
Variations :
– typicum sur sols plus ou moins mésoxérophiles, assez pauvres en bases et en éléments nutritifs, acidiclines, sans doute plus riches en cailloux calcaires dans la matrice argileuse, avec Alchemilla alpigena, Dianthus hyssopifolius, Orchis mascula, Avenella flexuosa, Dactylorhiza viridis, Cirsium acaulon, Galium pumilum, Gentiana angustifolia, Thymus polytrichus, Polygala calcarea, Anthoxanthum odoratum ;
– drymocallidetum rupestris Misset 2017, sur pentes moins fortes et sols modérément secs, à bonne réserve en eau, différenciée par Poa alpina subsp. a., Euphrasia officinalis subsp. rostkoviana, Drymocallis rupestris, Veronica spicata subsp. s., Aster alpinus, Arabis ciliata, Alchemilla glabra, A. monticola. sur pentes moins fortes et sols modérément secs, à bonne réserve en eau, différenciée par Poa alpina subsp. a., Euphrasia officinalis subsp. rostkoviana, Drymocallis rupestris, Veronica spicata subsp. s., Aster alpinus, Arabis ciliata, Alchemilla glabra, A. monticola ; type nomenclatural : rel. 6 du tab. 12 in Misset (2017, Psychodrômia, Bull. Soc. Bot. de la Drôme 2 : 60).
Synchorologie : Vercors méridional.
Axes à développer : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Misset C., 2017.
FICHE N° 27-11
Association : Seslerio caeruleae – Laserpitietum sileris Moor ex Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 79)
Synonyme : Laserpitio sileris – Seslerietum variae Moor in Moor & Schwarz 1957 (Beitr. Geobot. Landesaufn. Schweiz 37 : 40) nom inval. (art. 7).
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : Moor et Schwarz (1957) appuient leur description sur un texte listant les espèces caractéristiques, sans localité ni quantification ; en revanche, Béguin (1972) inclut dans son travail des relevés de Moor, parmi lesquels nous désignons un lectotype : rel. 17 (lectotypus nominis hoc loco) du tab. 8 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 76).
Physionomie : pelouse-ourlet plus ou moins fermé largement dominé par Laserpitium siler et dans une moindre mesure par Sesleria caerulea ; photo 5.
Combinaison caractéristique d’espèces : Sesleria caerulea, Laserpitium siler, Libanotis pyrenaica, Coronilla vaginalis, Sedum album, Anthericum ramosum, Orobanche laserpitii-sileris, Leucanthemum adustum, Dianthus saxicola, Globularia cordifolia, Scabiosa lucida, Galium anisophyllon, Teucrium montanum, T. chamaedrys, Carduus defloratus subsp. defloratus, Thesium alpinum, Aster alpinus, Euphrasia salisburgensis, Carex sempervirens, Calamagrostis varia, Origanum vulgare, Laserpitium latifolium.
Synécologie : pelouse mésoxérophile thermophile des étages montagnard supérieur et subalpin inférieur, calcicole, établie sur des affleurements calcaires, sur des sols de type rendzine.
Variations :
– typicum ;
– arctostaphyletosum uvae-ursi Béguin 1972, lié aux vires et différencié par des espèces saxicoles telles que Sempervivum tectorum, Asplenium ruta-muraria, Kernera saxatilis, Sideritis hyssopifolia, Erinus alpinus ou Arctostaphylos uva-ursi ; lectotypus nominis hoc loco : rel. 4 du tab. 8 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 79) ;
– stipetosum calamagrostis Béguin 1972, transition vers les éboulis de matrice plus ou moins fine, différencié par Achnatherum calamagrostis (= Stipa calamagrostis), Thalictrum minus subsp. saxatile, Vincetoxicum hirundinaria ; lectotypus nominis hoc loco : rel. 24 du tab. 8 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 80).
Synchorologie :
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al., 2014 ; Prunier et al., 2019), Préalpes du Nord jusqu’au Dévoluy isèrois (Villaret, 2007 ; Mikolajczak et al., 2014) ;
– sous-associations décrites du Jura mais potentiellement présentes sur toute l’aire connue.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Mikolajczak A. et al., 2014 ; Moor M. & Schwarz U., 1957 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.

FICHE N° 27-12
Association : Veronico fruticulosae – Agrostietum schleicheri Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 91).
Synonymes : -.
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 23 (lectotypus nominis hoc loco) du tab. 10 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 93).
Physionomie : pelouse ouverte riche en chaméphytes suffrutescents.
Combinaison caractéristique d’espèces : Coronilla vaginalis, Sedum album, Dianthus saxicola, Veronica fruticulosa, Agrostis schleicheri, Athamanta cretensis, Globularia cordifolia, Galium anisophyllon, Erinus alpinus, Hippocrepis comosa, Draba aizoides, Aster alpinus, Helianthemum oelandicum, Minuartia capillacea, Kernera saxatilis.
Synécologie : pelouse calcicole mésoxérophile à xérophile, thermophile, des étages montagnard supérieur et subalpin établie sur des lithosols ou des rendzines, en situation de vires de paroi ou sur des pentes moyennes à fortes stabilisées par des affleurements de calcaires durs plus ou moins verticaux, pouvant alterner avec des horizons plus caillouteux et ébouleux.
Variations :
– typicum [syn. : sideritidetosum hyssopifoliae Béguin 1972], type de l’association, sur sols de type rendzine, en contexte plus rocailleux et ébouleux, moins rupicole, avec Laserpitium siler, Sideritis hyssopifolia, Anthyllis montana, Campanula thyrsoides, Teucrium montanum, Leucopoa pulchella subsp. jurana ;
– asplenietosum rutae-murariae Béguin 1972, lié aux sols squelettiques stabilisés ou en contexte de paroi, différencié par Asplenium ruta-muraria, Sedum dasyphyllum, Saxifraga moschata, Sedum acre, Carex ornithopoda subsp. ornithopoda ; lectotypus nominis hoc loco : rel. 11 du tab. 10 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 93).
Synchorologie
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al., 2014), Préalpes du Nord (Villaret, 2007 ; Mikolajczak et al., 2014) ; groupements proches observés sur des affleurements calcaires dans les Écrins (Valjouffrey, obs. CBNA).
Axes à développer : étudier la présence de ce syntaxon dans les massifs calcaires plus internes (Écrins, Vanoise notamment) et préciser les relations avec le syntaxon suivant.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.
FICHE N° 27-13
Association : Teucrio montani – Globularietum cordifoliae J.-L. Rich., Bourgnon & Strub. ex Prunier, Sanz & Van Es ass. nov. hoc loco.
Synonymes : Teucrio – Globularietum prov. J.-L. Rich. et al. 1977.
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 4 (typus nominis hoc loco) du tab. 8 in Richard et al. (1977, Bull. Soc. Fribourg. Sci. Nat. 66 (1) : 25).
Physionomie : pelouse ouverte riche en chaméphytes suffrutescents dominée par les deux espèces éponymes et Festuca laevigata.
Combinaison caractéristique d’espèces : Globularia cordifolia, G. nudicaulis, Teucrium montanum, Festuca laevigata, Gypsophila repens, Dianthus saxicola, Sempervivum tectorum, Carex humilis, Astragalus sempervirens, Bupleurum ranunculoides, Draba aizoides, Saxifraga paniculata, Aster alpinus, Sesleria caerulea, Leontopodium nivale subsp. alpinum, Kernera saxatilis, Allium lusitanicum, Primula auricula.
Synécologie : pelouse calcicole xérophile établie sur les sols superficiels au contact d’affleurements rocheux calcaires, sur des pentes variées mais toujours bien ensoleillées, aux étages montagnard supérieur et subalpin inférieur.
Variations : -.
Synchorologie :
– territoire d’observation : Jura (Prunier et al., 2019), Préalpes du Nord en Suisse et Haute-Savoie (Richard, 1977 ; Prunier et al., 2019), Alpes internes du Nord dans le Haut-Valais (Richard, 1984) ; pelouses affines dans les Alpes internes et intermédiaires du Sud.
Axes à développer : présence attestée dans les Bauges, les Aravis, la Vanoise et la Chartreuse ; étudier sa présence ailleurs dans les massifs calcaires des Alpes internes françaises du Nord et du Sud, et préciser les relations avec le syntaxon précédent.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Prunier P. et al., 2019 ; Richard J.-L., 1977, 1984.
FICHE N° 27-14
Association : Teucrio montani – Festucetum macrophyllae Ritter 1972 (Vegetatio 24 : 329).
Synonymes : -.
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 2 (lectotypus nominis hoc loco) du tab. 4 in Ritter (1972, Vegetatio 24 : 331).
Physionomie : pelouse ouverte riche en chaméphytes suffrutescents dominée par les deux espèces éponymes.
Combinaison caractéristique d’espèces : Globularia cordifolia, G. bisnagarica, G. nudicaulis, Teucrium montanum, Festuca laevigata (= F. glauca var. macrophylla, F. ovina var. macrophylla), Sideritis hyssopifolia, Coronilla vaginalis, Helictotrichon sedenense, Bupleurum petraeum, Gentiana angustifolia, Tulipa sylvestris subsp. australis, Sedum ochroleucum, Antennaria dioica, Alchemilla alpigena, Botrychium lunaria, Ranunculus kuepferi, Veronica spicata, Valeriana montana, Saxifraga paniculata.
Synécologie : pelouse calcicole xéroclinophile à xérophile, thermophile, établie sur des sols peu profonds pouvant être légèrement décalcifiés en surface ou sur des sols plus superficiels au contact d’affleurements rocheux calcaires. Étages montagnard supérieur et subalpin inférieur.
Variations : -.
Synchorologie :
– territoire d’observation : Vercors, Dévoluy (Ritter, 1972 ; Villaret, 2007).
Axes à développer : confirmer sa présence hautement probable dans les zones calcaires des Écrins.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Bensetitti F. et al., 2002 ; Ritter J., 1972 ; Villaret J.-C., 2007.
FICHE N° 27-15
Association : Minuartio vernae – Arenarietum multicaulis Béguin 1972 (Matér. Levé Géobot. Suisse 54 : 91).
Synonymes : -.
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 4 (lectotypus nominis hoc loco) du tab. 10 in Béguin (1972, Matér. Levé Géobot. Suisse 54 : 93).
Physionomie : pelouse ouverte riche en chaméphytes suffrutescents.
Combinaison caractéristique d’espèces : Minuartia verna, Arenaria multicaulis, Sedum album, Dianthus saxicola, Athamanta cretensis, Sempervivum tectorum, Globularia cordifolia, Gypsophila repens, Trinia glauca, Plantago media, Bromopsis erecta, Hippocrepis comosa, Saxifraga paniculata, Gentiana verna, Antennaria dioica, Poa molinerii, Sedum atratum, Gentianella campestris, Kernera saxatilis, Sesleria caerulea, Carex sempervirens, Helianthemum nummularium.
Synécologie : pelouse calcicole xérophile des bombements et croupes exposés au vent, pouvant être déneigée en hiver, sur des substrats calcaires stabilisés mais néanmoins ensoleillés, aux étages montagnard supérieur et subalpin inférieur.
Variations : -.
Synchorologie :
– territoire d’observation : Jura (Béguin, 1972 ; Mikolajczak et al., 2014 ; Prunier et al., 2019).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.
FICHE N° 27-16
Association : Androsaco villosae – Gentianetum angustifoliae Ritter 1972 (Vegetatio 24 : 337).
Synonymes : -.
Unités supérieures : Drabo aizoidis – Seslerion caeruleae (Béguin & Ritter in Béguin 1972) Delarze, Gonseth & Galland ex Sanz, Van Es & Corriol, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 25 (lectotypus nominis hoc loco) du tab. 4 in Ritter (1972, Vegetatio 24 : 338).
Physionomie : pelouse ouverte dominée par de petites graminées à feuilles fines et divers chaméphytes suffruescents formant des placages plus ou moins étendus ; photo 6.
Combinaison caractéristique d’espèces : Androsace villosa, Gentiana angustifolia, Globularia cordifolia, Dryas octopetala, Anthyllis montana, Thymus gpe serpyllum, Arenaria multicaulis, Silene acaulis, Oxytropis amethystea, Leontopodium nivale subsp. alpinum, Saxifraga paniculata, Antennaria dioica, Minuartia verna, Aster alpinus, Helianthemum oelandicum, Helictotrichon sedenense, Sesleria caerulea, Athamantha cretensis, Bupleurum petraeum.
Synécologie : pelouse calcicole pionnière xérophile et mésocryophile établie sur des buttes et hauts de versant exposés à des vents forts, suceptibles d’être déneigés en hiver, sur des substrats stabilisés par l’affleurement de calcaires durs.
Variations :
– typicum ;
– anthyllidetosum Allier & Ritter 1971, sous-association sous influence provençale faisant transition avec le Genistetum villarsii, différenciée par Anthyllis montana, Carex humilis, Teucrium montanum et par l’absence d’espèces des Oxytropido – Elynetalia telles que Dryas octopetala, Arenaria multicaulis, Leontopodium nivale subsp. alpinum, Minuartia verna ; lectotypus nominis hoc loco : rel. 1 du tab. III in Allier et Ritter (1971, Ann. Litt. Univ. Besançon 125 : 292).
Synchorologie :
– territoire d’observation : Préalpes du Sud, depuis le Vercors méridional (Ritter, 1972) jusqu’aux Préalpes de Grasse et de Castellane, Baronnies septentrionales (Allier & Ritter, 1970), Ventoux (Barbero, 1972).
Axes à développer : variabilité et chorologie dans les Alpes du Sud.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Allier C. & Ritter J., 1971 ; Barbero M., 1972 ; Barbero M. & Quézel P., 1975 ; Bensettitti F. et al., 2002 ; Mikolajczak A. et al., 2014 ; Ritter J., 1972 ; Villaret J.-C., 2007.

FICHE N° 27-17
Association : Seslerio caeruleae – Caricetum sempervirentis Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 nom. corr. hoc loco.
Synonyme : Seslerieto – Semperviretum Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 236).
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 2 (lectotypus nominis hoc loco) du tab. 8 in Braun-Blanq. & Jenny (1926, Denksch. Schweiz. Naturf. Ges. 63 (2) : 237).
Physionomie : pelouse plus ou moins ouverte, pouvant former des « gradins » caractéristiques, dominée par Carex sempervirens et Sesleria caerulea ; photo 7.
Combinaison caractéristique d’espèces : Carex sempervirens, Sesleria caerulea, Aster alpinus, Helianthemum oelandicum, H. nummularium, Oxytropis jacquinii, O. campestris, Gentiana clusii, Leontopodium nivale subsp. alpinum, Pedicularis verticillata, Myosotis alpestris, Agrostis alpina, Minuartia verna, Sedum atratum, Hieracium villosum, Globularia cordifolia, Biscutella laevigata subsp. laevigata, Viola calcarata, Homogyne alpina, Bartsia alpina, Bistorta vivipara.
Synécologie : pelouse primaire supraforestière des étages subalpin et alpin, neutrocalcicole à calcicole, mésoxérophile, thermophile, établie sur des sols assez humifères et carbonatés (rendzines) sur des versants bien ensoleillés, soumis à des déneigements précoces et à des phénomènes plus ou moins intenses d’érosion ou de cryoturbation.
Variations : nombreuses, en fonction notamment de la stabilité du substrat, de l’humidité édaphique et de la géographie :
– variante des éboulis fixés et alluvions torrentielles, différenciée par la présence de lithophytes migrateurs ;
– variante des vires rocheuses, différenciée par la présence de chasmophytes, notamment Athamanta cretensis, Saxifraga paniculata ;
– variante des situations les plus fraîches, différenciée par des espèces du Primulion intricatae, notamment Festuca violacea ;
– variante géographique des Alpes du Nord externes, différenciée par la présence de Globularia nudicaulis.
Synchorologie :
– territoire d’observation : Alpes du Nord (Villaret, 2007 ; Mikolajczak et al., 2014 ; Prunier et al., 2019…) ; cité ailleurs dans de nombreuses publications mais semble plutôt être remplacé par le Seslerio albicantis – Helictotrichetum sedenensis dans les Alpes du Sud, avec une large zone de transition située au niveau Vercors méridional – Écrins – Lautaret – plateau du Mont-Cenis.
Axes à développer :
– répartition à préciser dans les Préalpes de Savoie et d’Isère sur les plus hauts sommets ;
– poursuivre l’étude des variations.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Bensettiti F. et al., 2002 ; Braun-Blanquet J. & Jenny H., 1926 ; Mikolajczak A. et al., 2014 ; Mikolajczak A., 2018 ; Prunier P. et al., 2019 ; Quantin A. & Nétien G., 1940 ; Villaret J.-C., 2007.

FICHE N° 27-18
Association : Astragalo leontini – Seslerietum caeruleae J.-L. Rich. ex Corriol, Sanz & Van Es ass. nov. hoc. loco.
Synonyme : Astragalo – Seslerietum J.-L. Rich. 1985 (Bot. Helv. 95 (2) : 200) nom. inval. (art. 5).
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 29 (typus nominis hoc loco) du tab. 2 in Richard (1985, Bot. Helv. 95 (2) : 201).
Physionomie : pelouse plus ou moins ouverte à Sesleria caerulea, Agrostis alpina et Carex humilis.
Combinaison caractéristique d’espèces : Sesleria caerulea, Astragalus leontinus, Carex humilis, Dianthus saxicola, Globularia cordifolia, Gypsophila repens, Teucrium montanum, Leucanthemum coronopifolium, Astragalus australis, Oxytropis campestris, Festuca pumila, Leontopodium nivale subsp. alpinum, Bupleurum ranunculoides, Saxifraga paniculata.
Synécologie : pelouse primaire de l’étage alpin inférieur en situation d’adret, calcicole, mésoxérophile à xérophile, établie sur des sols secs superficiels en adrets (moins humifères que ceux de l’association précédente, ce qui explique la raréfaction de Carex sempervirens), différenciée par des espèces xérophiles pionnières du Drabo – Seslerion.
Variations :
– typicum [syn. : piloselletosum officinarum J.-L. Rich. 1985 nom. inval.) sur substrats plus ou moins acides (ophiolithes et gneiss), différencié par Pilosella officinarum, Sempervivum arachnoideum, Anemone halleri, Euphrasia alpina, Bellardiochloa variegata, Veronica fruticans, Cerastium arvense subsp. strictum, Pedicularis tuberosa, Myosotis stricta, Trifolium pratense var. villosum et Poa perconcinna ;
– linarietosum alpinae J.-L. Rich. ex Corriol, Sanz & Van Es subass. nov. hoc loco, sur substrat à pH neutre (calcschistes et parfois dolomie), différencié par Euphrasia salisburgensis, Astragalus monspessulanus, Trisetum distichophyllum, Leontodon hyoseroides, Linaria alpina et Herniaria alpina ; typus nominis hoc loco : rel 40 du tab 2 in Richard (1985, Bot. Helv. 95 (2) : 201).
Synchorologie :
– territoire d’observation : Haut-Valais (Richard, 1985), Mont-Cenis (Prunier et al., 2019) ; groupements proches récemment observés en Vanoise dans le massif de la Sassière et le vallon de la Rocheure (obs. CBNA).
Axes à développer : préciser sa répartition en Vanoise.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Prunier P. et al., 2019 ; Richard J.-L., 1985.
FICHE N° 27-19
Association : Artemisio glacialis – Festucetum pumilae J.-L. Rich. ex Corriol, Sanz & Van Es ass. nov. hoc. loco.
Synonymes :
– Festucetum pumilae sensu Béguin 1972 ;
– Artemisio glacialis – Festucetum pumilae J.-L. Rich. 1985 nom. inval. (art. 5).
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 6 (typus nominis hoc loco) du tab. 1 in Richard (1985, Bot. Helv. 95 (2) : 198).
Physionomie : pelouse ouverte pionnière où codominent Festuca pumila et de petits chaméphytes.
Combinaison caractéristique d’espèces : Festuca pumila, Oxytropis helvetica, O. campestris, Artemisia glacialis, Androsace vitaliana, Astragalus sempervirens, Alyssum alpestre, Herniaria alpina, Viola rupestris, Minuartia verna, Leontopodium nivale subsp. alpinum, Saxifraga moschata, Comastoma tenellum, Carex curvula subsp. rosae, C. myosuroides, Linaria alpina, Poa perconcinna.
Synécologie : pelouse primaire pionnière de l’étage alpin supérieur en situation d’adret, calcicole, xérophile, établie sur des sols secs superficiels pauvres en humus, en situation de vires caillouteuses et hauts de versant assez stabilisés, sur roches mères préférentiellement schisteuses (variées mais hors calcaire dur) ; association à la charnière entre Seslerion caeruleae (par son caractère thermoxérophile) et Oxytropido – Elynion (par la présence des espèces cryotolérantes).
Variations : -.
Synchorologie :
– territoire d’observation : Haut-Valais, probable dans toutes les Alpes internes sud-occidentales (Richard, 1985) ; groupements très proches observés en Vanoise, Queyras (obs. CBNA) et Ubaye, mais sans Poa perconcinna.
Axes à développer : préciser sa répartition dans les Alpes internes françaises.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972, Prunier P. et al., 2019 ; Richard J.-L., 1985.
FICHE N° 27-20
Association : Gentiano angustifoliae – Seslerietum caeruleae Bannes-Puygiron 1933 (Trav. Inst. Bot. Montpellier : 151).
Synonyme : Seslerietum prealpinum Archiloque, Borel & Lavagne 1970.
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel.1 (lectotypus nominis hoc loco) du tab. p. 150 in Bannes-Puygiron (1933, Trav. Inst. Bot. Montpellier : 150).
Physionomie : pelouse plus ou moins fermée à Sesleria caerulea et Carex sempervirens riche en petites dicotylédones colorées ; photo 8.
Combinaison caractéristique d’espèces : Gentiana angustifolia, Pedicularis gyroflexa, Oxytropis amethystea, Arenaria multicaulis, Dianthus gratianopolitanus, Alchemilla alpigena, Hieracium villosum, Armeria alpina, Minuartia verna, Scabiosa lucida, Aster alpinus, Agrostis alpina, Helictochloa pratensis, Helictotrichon sedenense, Eryngium spinalba, Globularia cordifolia, Antennaria dioica, Saxifraga paniculata, (Oxytropis campestris, Astragalus australis).
Synécologie : pelouse calcicole mésoxérophile plutôt thermophile établie sur des versants assez stabilisés sur des substrats de type rendzine des étages subalpin et alpin inférieur.
Variations : -.
Synchorologie :
– territoire d’observation : Haut-Diois, Vercors, Chartreuse méridionale, Ventoux. Décrite par Bannes-Puygiron en 1933 probablement en une limite d’aire et en situation un peu particulière de « prés suspendus » sur les sommets du Diois, cette association semble pourtant bien correspondre aux pelouses mésoxérophiles à Sesleria caerulea et Carex sempervirens subalpines des sommets du Vercors et du Dévoluy (obs. CBNA), ainsi que du sud de la Chartreuse. Elle peut être vue comme une vicariante méridionale relayant dans ces massifs l’Alchemillo conjunctae – Seslerietum caeruleae Aubert & Luquet 1930 des Préalpes du Nord et du Jura, particulièrement sa sous-association alticole bupleuretosum.
Axes à développer : à étudier dans le Dévoluy.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Archiloque A. et al., 1970 ; de Bannes-Puygiron G., 1933 ; Barbero M., 1968 ; Barbero M. & Quézel P., 1975 ; Ritter J., 1972 ; Villaret J.-C., 2007.

FICHE N° 27-21
Association : Seslerio caeruleae – Helictotrichetum sedenensis Lippmaa 1933 (Acta Inst. Horti. Bot. Tartu. 3 : 52) nom. corr. propos. hoc loco.
Synonymes :
– association à Sesleria caerulea et Avena montana Lippmaa 1933 ;
– Seslerieto – Avenetum montanae Braun-Blanq. 1954.
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 3 (lectotypus nominis hoc loco) du tab. 7 in Lippmaa (1933, Acta Inst. Horti. Bot. Tartu. 3 : 52).
Physionomie : pelouse ouverte formant optimalement des guirlandes, structurée par les deux espèces éponymes et plus ou moins riche en chaméphytes, notamment Astragalus sempervirens ; photo 9.
Combinaison caractéristique d’espèces : Sesleria caerulea, Helictotrichon sedenense, Astragalus sempervirens, A. australis, Alyssum alpestre, Androsace vitaliana, Plantago maritima subsp. serpentina, Koeleria vallesiana, Onobrychis montana, Arenaria multicaulis, Leontopodium nivale subsp. alpinum, Oxytropis campestris, Silene acaulis, Bistorta vivipara, Aster alpinus, Antennaria dioica, Globularia cordifolia, Helianthemum oelandicum, Carex sempervirens, Euphrasia alpina.
Synécologie : pelouse calcicole, mésoxérophile et thermophile développée sur des sols peu humifères, située en adret aux étages subalpin supérieur et alpin, sur des versants soumis à un déneigement précoce et plus ou moins perturbés par des phénomènes de cryoturbation.
Variations : -.
Synchorologie :
– territoire d’observation : Alpes du Sud plutôt internes à partir des Écrins, du Lautaret…
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Bensettiti F. et al., 2002 ; Braun-Blanquet J., 1954 ; Dalmas J.-P., 1972 ; Guinochet M., 1938 ; Lacoste A., 1975 ; Lippmaa T., 1933 ; Mikolajczak A. et al., 2014 ; Van Es J., 2013 ; Villaret J.-C., 2007.

FICHE N° 27-22
Association : Hedysaro boutignyani – Caricetum austroalpinae Van Es & L. Foucaut ass nov hoc loco.
Synonymes :
– pelouse subalpine, neutro-basophile, mésophile à Carex austroalpina Van Es 2003 (Étude de la végétation du site Natura 2000 « le Mercantour » : 124) ;
– couloirs herbeux à Carex austroalpina L. Foucaut 2006 (Courr. Sci. Parc Nat. Rég. Verdon : 26).
Unités supérieures : Seslerion caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 390366 (typus nominis hoc loco) du tab. 3 hoc loco retranscrit ici
J. Van Es, Uvernet-Fours (04), 2 150 m, 2/VIII/2002, 41 taxons
Hedysarum boutignyanum 3, Carex austroalpina 2, Sesleria caerulea 2, Festuca violacea 2, Leucanthemum coronopifolium 2, Lotus corniculatus s. l. 2, Helictotrichon sedenense *sedenense 1, Campanula scheuchzeri *scheuchzeri 1, Poa alpina 1, Hieracium gr. caesioides 1, Solidago virgaurea *minuta 1, Ranunculus gr. montanus 1, Anthyllis vulneraria *alpestris 1, Homogyne alpina 1, Anemone baldensis 1, Trifolium pratense 1, Soldanella alpina 1, Festuca pumila 1, Alchemilla alpigena 1, Valeriana montana +, Myosotis alpestris +, Euphrasia salisburgensis +, Arenaria multicaulis +, Anemone alpina +, Leontodon hispidus +, Hieracium villosum +, Galium obliquum +, Adenostyles alpina +, Achillea millefolium +, Cerastium arvense +, Helianthemum italicum +, Botrychium lunaria +, Coristospermum ferulaceum +, Polygala alpina +, Minuartia verna +, Astragalus australis +, Imperatoria ostruthium +, Silene acaulis +, Bupleurum petraeum +, Poa cenisia +, Dactylorhiza viridis +.
Physionomie : pelouse riche en grandes herbes, ouverte à fermée et marquée par les touffes de Carex austroalpina.
Combinaison caractéristique d’espèces : Carex austroalpina, Hedysarum boutignyanum, Linum alpinum, Valeriana montana, Helictotrichon sedenense subsp. sedenense, Leucanthemum atratum subsp. coronopifolium, Bellidiastrum michelii.
Synécologie : pelouse basiphile, mésophile à hygroclinophile liée à des versants rocailleux ou ébouleux d’ubacs de l’étage (montagnard) subalpin.
Variations :
– variante sur pentes plus faibles ou stabilisées, à sol plus profond, différenciée par des espèces de milieux plus stables, Astragalus danicus, Plantago maritima subsp. serpentina, Gentiana lutea ;
– variante des pentes mieux exposées de la Haute-Ubaye, différenciée par Helictotrichon parlatorei.
Synchorologie :
– territoire d’observation : décrit du Mercantour (Van Es, 2003) et des Préalpes du Verdon (Foucaut, 2006) sous une forme plus montagnarde ; présent dans toutes les Alpes du Sud.
Axes à développer :
– préciser les variations chorologiques dans les Alpes du Sud et préciser la limite nord dans les Alpes françaises ;
– formalisation des variantes à poursuivre ;
– confirmer le rattachement phytosociologique pour cette association tendant également vers le Caricion ferrugineae.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Foucaut L. & Béthouse J.-E., 2006 ; Mikolajczak A. et al., 2014 ;Van Es J., 2003.
FICHE N° 27-23
Association : Alchemillo alpigenae – Bellardiochloetum variegatae Lacoste ex Sanz, Van Es & Corriol ass. nov. hoc. loco.
Synonyme : Alchemilleto[6] – Poetum violaceae Lacoste 1965 (Rev. Gén. Bot. 72 : 606) nom. inval. (art. 3b).
Unités supérieures : Anthoxantho odorati – Plantaginion brutiae Barbero & Quézel 1975, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 3 (typus nominis hoc loco) du tab. 1 in Lacoste (1965, Rev. Gén. Bot. 72 h. t.).
Physionomie : pelouse / prairie assez fermée plus ou moins riche en hautes herbes.
Combinaison caractéristique d’espèces : Sesleria caerulea, Bellardiochloa violacea, Astragalus danicus, A. sempervirens, Galium obliquum, Helictotrichon sedenense, Alchemilla alpigena, Helianthemum nummularium, Carlina acaulis subsp. caulescens, Anthyllis vulneraria, Bellidiastrum michelii, Plantago maritima subsp. serpentina, Carex sempervirens, Myosotis alpestris, Onobrychis montana, Achillea millefolium, Lotus corniculatus s. l., Anthoxanthum odoratum s. l., Trifolium pratense, T. montanum.
Synécologie : pelouse secondaire basiphile, mésophile, de l’étage subalpin, sur sols relativement profonds pouvant être décalcifiés en surface, généralement établie sur des faibles pentes plutôt en expositions fraîches, évoluant en lien avec des mélézins sur substrat calcaire.
Variations : -.
Synchorologie :
– territoire d’observation : Alpes du Sud (Lacoste, 1965 ; Barbero, 1972 ; Van Es et al., 2014).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Lacoste A., 1965 ; Van Es J. et al., 2014.
FICHE N° 27-24
Association : Anthoxantho odorati – Avenelletum flexuosae Archiloque, Borel & Lavagne 1971 nom. corr. propos. hoc loco.
Synonymes :
– association à Anthoxanthum villosum et Deschampsia montana Archiloque, Borel & Lavagne 1971 (Colloq. Intern. Mil. Nat. supraforestiers Mont. Bass. Occ. Médit. : 222) ;
– Ranunculo – Fritillarietum Barbero & Quézel 1975 (Ecol. Medit. 1 : 24) (synonyme syntaxonomique inclus) ;
– Anthoxantho-Deschampsietum Barbero & Quézel 1975 (Ecol. Medit. 1 : 21) ;
– Gentianeto – Ranunculetum pyrenaei Guin. 1975 (Anal. Inst. Bot. Cavanilles 32 : 1293) (synonyme syntaxonomique inclus).
Unités supérieures : Anthoxantho odorati – Plantaginion brutiae Barbero & Quézel 1975, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : non lectotypifiable, les auteurs appuyant leur description sur une liste synthétique ; neotypus nominis hoc loco : rel. 10 du tab. 10 in Barbero & Quézel 1975 (Ecol. Medit. 1 : 22).
Physionomie : pelouse fermée dominée par diverses petites graminées (Anthoxanthum odoratum, Avenella flexuosa, Agrostis capillaris) et des géophytes.
Combinaison caractéristique d’espèces : Ranunculus kuepferi, Fritillaria tubiformis, Bulbocodium vernum, Crocus albiflorus, Coeloglossum viride, Dactylorhiza sambucina, Botrychium lunaria, Alchemilla gpe flabellata, Tulipa sylvestris subsp. australis, Gentiana verna subsp. delphinensis, Astragalus danicus, Gentianella campestris, Trifolium thalii, Leontopodium alpinum subsp. nivale, Festuca violacea, Anthoxanthum odoratum s. l., Avenella flexuosa.
Synécologie : pelouse mésophile, neutrophile à acidiclinophile, chionoclinophile, developpée dans des combes et bas de versants à faibles pentes sur substrat calcaire, aux étages montagnards supérieur et subalpin.
Variations :
– variante des secteurs de stationnements prolongés des troupeaux, différenciée par la présence d’espèces eutrophiles : Poa alpina, Taraxacum spp. (Barbero & Quézel, 1975) ;
– variante des plus hautes altitudes enrichie en espèces plus chionophiles, différenciée par Soldanella alpina, Linum alpinum, Viola calcarata, etc.
Synchorologie :
– territoire d’observation : Préalpes du Sud, depuis le mont Ventoux jusqu’aux Préalpes de Castellane et Grasse (Barbero & Quézel ,1975 ; Noble, 2008).
Axes à développer : étude des vaiations à poursuivre.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Archiloque A. et al., 1971 ; Barbero M. & Quézel P., 1975 ; Guinochet M., 1975 ; Noble V., 2008.
FICHE N° 27-25
Association : Tephroserido integrifoliae – Bellardiochloetum variegatae Barbero & Quézel 1975 nom. corr. propos. hoc loco.
Synonyme : Senecioni campestris – Poetum violaceae Barbero & Quézel 1975 (Ecol. Medit. 1 : 23).
Unités supérieures : Anthoxantho odorati – Plantaginion brutiae Barbero & Quézel 1975, Seslerietalia caeruleae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926.
Type nomenclatural : rel. 4 (lectotypus nominis hoc loco) du tab. 11 in Barbero & Quézel (1975, Ecol. Medit. 1 : 23).
Physionomie : pelouse fermée à Bellardiochloa variegata et diverses graminées autres que Sesleria caerulea.
Combinaison caractéristique d’espèces : Anthoxanthum odoratum s. l., Bellardiochloa violacea, Luzula alpina, Tephroseris integrifolia subsp. integrifolia, Astragalus danicus, Plantago media, Tulipa sylvestris subsp. australis, Gentianella campestris, Helictochloa pratensis, Trifolium pratense, Antennaria dioica, Festuca violacea, Myosotis alpestris, Plantago maritima subsp. serpentina, Alchemilla alpigena, Euphrasia salisburgensis, Potentilla crantzii, Pedicularis gyroflexa, Botrychium lunaria.
Synécologie : pelouse mésophile acidiclinophile de l’étage subalpin, liée à des expositions plutôt fraîches sur des substrats gréseux préalpins.
Variations : -.
Synchorologie :
– territoire d’observation : Préalpes du Sud, entre Tinée et Verdon (Barbero & Quézel, 1975 ; Van Es et al., 2014).
Axes à développer : comme le précisent les auteurs, ce groupement est à la charnière des Festucetalia spadiceae et paraît constituer une transition entre l’Alchemillo hoppeanae – Bellardiochloetum variegatae Lacoste 1965 et le Phyteumato michelii – Poetum violaceae Barbero 1970.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M. & Quézel, 1975 ; Van Es J. et al., 2014.
FICHE N° 27-26
Association : Pediculari comosae – Ranunculetum geraniifolii Allier 1971 (Colloq. Intern. Mil. Nat. supraforestiers Mont. Bass. Occ. Médit. : 178).
Synonymes : -.
Unités supérieures : Ononidenion cristatae Gaultier ex Corriol, Sanz & J.-M. Royer, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : non lectotypifiable, l’auteur appuie sa description sur une colonne synthétique.
Physionomie : pelouse fermée dominée par Sesleria caerulea et Carex sempervirens.
Combinaison caractéristique d’espèces : Sesleria caerulea, Carex sempervirens, Pedicularis comosa, Ranunculus aduncus, Alchemilla alpigena, Botrychium lunaria, Primula veris subsp. columnae, Thesium alpinum, Lilium martagon, Trifolium montanum, Linum suffruticosum subsp. appressum, Lavandula angustifolia, Bromopsis erecta, Knautia collina, Carex humilis, Polygala calcarea, Anemone alpina, Helianthemum nummularium, Aster alpinus.
Synécologie : pelouse mésophile à mésoxérophile plutôt psychrophile développée sur des sols humocalciques édifiés sur des pentes moyennes à fortes d’ubac, à l’étage montagard.
Variations : -.
Synchorologie :
– territoire d’observation : Baronnies drômoises (Allier, 1971) et haut-alpines.
Axes à développer :
– répartition à préciser dans les Préalpes du Sud, fort probable dans le Dévoluy notamment. Cette pelouse présente des affinités écologique et floristique avec celles de l’Alchemillo hoppeanae – Bellardiochloion variegatae ;
– néotypification.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Allier C., 1971.
FICHE N° 27-27
Association : Astragalo sempervirentis – Ononidetum cenisiae Barbero 1972 (Études phytosociologiques et écologiques comparées des végétations orophiles… 1 : 218).
Synonymes :
– association à Astragalus aristatus et Ononis cenisia Archiloque, Borel & Lavagne 1971 (Colloq. Intern. Mil. Nat. supraforestiers Mont. Bass. Occ. Médit. : 218) (synonyme syntaxonomique inclus) ;
– pelouse à Astragalus aristatus Archiloque, Borel & Devaux 1974 (Coll. Bull. Carte Vég. Provence Alpes Sud 1 : 113) (synonyme syntaxonomique inclus) ;
– landine à Globularia cordifolia et Lavandula angustifolia Lejoly 1975 (Phytosociologie écologique en moyenne montagne méditerranéenne… 2 : 320) (synonyme syntaxonomique inclus).
Unités supérieures : Ononidenion cristatae Gaultier ex Corriol, Sanz & J.-M. Royer, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 6 (lectotypus nominis hoc loco) du tab. 23 in Barbero (1972, Études phytosociologiques et écologiques comparées des végétations orophiles… 2 : 111).
Physionomie : pelouse écorchée dominée par des chaméphytes suffrutescents.
Combinaison caractéristique d’espèces : Astragalus sempervirens, Ononis cenisia, Linum suffruticosum subsp. appressum, Globularia cordifolia, Galium obliquum, Astragalus depressus, Armeria arenaria subsp. bupleuroides, Eryngium spinalba, Lavandula angustifolia subsp. angustifolia, Dianthus subacaulis, Coronilla minima, Bromopsis erecta, Iberis saxatilis, Helictotrichon sedenense, Sesleria caerulea, Aster alpinus, Ononis striata, Myosotis stricta, Minuartia capillacea, Valeriana tuberosa.
Synécologie : pelouse mésoxérophile à xérophile centrée sur l’étage montagnard sur des rendzines initiales (calcaires durs ou marneux) en situation d’adret sur des versants stabilisés ou des replats en situation de plateau.
Variations : variante géographique dans les Préalpes de Grasse et de Castellane marquée par la présence de Minuartia glomerata subsp. burnatii et Leucanthemum burnati.
Synchorologie :
– territoire d’observation : Préalpes du Sud.
Axes à développer :
– variantes géographiques à étudier dans le Gapençais, le Dévoluy et le Vercors notamment ;
– variantes altitudinales à étudier (transition avec le Gentiano angustifoliae – Seslerietum caeruleae dans le Dévoluy ou le Vercors) ;
– dans sa thèse sur le mont Ventoux, Gontard (1955) propose une association, l’Aveno – Galietum villarsii, qu’il place initialement dans le Thlaspion rotundifolii (classe des Thlaspietea rotundifolii) ; par la suite, Barbero et Quézel (1975) la retiennent dans les Astragaletalia sempervirentis. La difficulté d’accès à cette thèse ne nous a pas permis d’intégrer les relevés originaux au tableau des Elyno – Seslerietea et de confirmer cette position synsystématique ; étude et typification à poursuivre.
Correspondances : IC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Archiloque A. et al., 1970 ; Archiloque A. et al., 1971 ; Archiloque A. et al., 1974 ; Barbero M., 1968, 1972 ; Barbero M. & Quézel P., 1975 ; Chaix G., 1954 ; Gontard P., 1955 ; Lejoly J., 1975 ; Noble V., 2008.
FICHE N° 27-28
Association : Astragalo sempervirentis – Onosmetum fastigiatae Lacoste 1964 (Bull. Soc. Bot. France 111 : 61).
Synonymes :
– pelouse à Astragalus aristatus Chaix 1954 (Étude phytogéographique des vallées supérieures de la Romanche… : 95) (synonyme syntaxonomique inclus) ;
– landine à Astragalus sempervirens et Globularia cordifolia Lejoly 1975 (Phytosociologie écologique en moyenne montagne méditerranéenne… 2 : 476) (synonyme syntaxonomique inclus) ;
– Globulario cordifoliae – Astragaletum sempervirentis (Lejoly 1975) Julve 1993 prov.
Unités supérieures : Ononidenion cristatae Gaultier ex Corriol, Sanz & J.-M. Royer, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 6 (lectotypus nominis hoc loco) du tab. 1 in Lacoste (1964, Bull. Soc. Bot. France 111 : 64).
Physionomie : pelouse écorchée dominée par des chaméphytes suffrutescents.
Combinaison caractéristique d’espèces : Astragalus sempervirens, Bupleurum ranunculoides, Onosma tricerosperma subsp. fastigiata, Leucanthemum adustum, Ononis cristata, Onobrychis montana, Plantago maritima subsp. serpentina, Teucrium montanum, Linum suffruticosum subsp. appressum, Galium obliquum, Potentilla tabernaemontani, Astragalus monspessulanus, Dianthus godronianus, D. pavonius, Scabiosa columbaria, Globularia cordifolia, Senecio doronicum, Bromopsis erecta, Aster alpinus, Sesleria caerulea, Draba aizoides.
Synécologie : pelouse calcicole mésoxérophile à xérophile développée sur des pentes raides en adret de l’étage subalpin.
Variations :
– typicum [syn. : ononidetosum cristatae Lacoste 1964], sur sol peu épais, différencié par Ononis cristata, Leucanthemum adustum, Carlina acanthifolia subsp. acanthifolia, Euphrasia salisburgensis ;
– carlinetosum acaulis Lacoste 1964, sur sol plus profond avec Carlina acaulis subsp. caulescens, Poa alpina, Trifolium montanum ; lectotypus nominis hoc loco : rel. 7 du tab. 1 in Lacoste (1964, Bull. Soc. Bot. France 111 : 64).
Synchorologie :
– territoire d’observation : Alpes du Sud intermédiaires et internes dans le Haut-Var, le Haut-Verdon, Haute-Roya, Tinée, Ubaye, Queyras et Briançonnais.
Axes à développer : préciser les variations géographiques entre les massifs intermédiaires (Haut-Verdon, Haut-Var) et internes (Queyras, Briançonnais, Ubaye).
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Chaix G., 1954 ; Lacoste A., 1964 ; Lejoly J., 1975, Van Es J. et al., 2014.
FICHE N° 27-29
Association : Arenario cinereae – Helictotrichetum sempervirentis Barbero 1968 (Bull. Soc. Bot. France 115 : 230).
Synonymes :
– pelouse à Avena sempervirens et Koeleria alpicola Archiloque, Borel & Lavagne 1971 (Colloq. Intern. Mil. Nat. supraforestiers Mont. Bass. Occ. Médit. : 213) (synonyme syntaxonomique inclus) ;
– pelouse à Avena sempervirens Archiloque, Borel & Devaux 1974 (Coll. Bull. Carte Vég. Provence Alpes Sud 1 : 118) (synonyme syntaxonomique inclus) ;
– pelouse à Avena sempervirens et Lavandula angustifolia Lejoly 1975 (Phytosociologie écologique en moyenne montagne méditerranéenne… 2 : 316) (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Helictotrichenion sempervirentis, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : non lectotypifiable, l’auteur appuie sa description sur une liste synthétique.
Physionomie : pelouse écorchée marquée par Helictotrichon sempervirens.
Combinaison caractéristique d’espèces : Helictotrichon sempervirens, H. setaceum, Campanula spicata, Arenaria cinerea, Lilium pomponium, Hypericum hyssopifolium, Pedicularis gyroflexa, Astragalus sempervirens, Plantago maritima subsp. serpentina, Linum suffruticosum subsp. appressum, Lavandula angustifolia, Galium obliquum, Teucrium lucidum, Cyanus gr. triumfettii, Globularia cordifolia, Dianthus scaber, Astragalus monspessulanus, Trinia glauca, Anthyllis montana.
Synécologie : pelouse calcicole mésoxérophile à xérophile, thermophile, montagnarde, établie sur des éboulis en voie de fixation ou sur des versants rocailleux.
Variations : variante géographique de Provence orientale marquée par Arenaria cinerea, Teucrium lucidum, Lilium pomponium.
Synchorologie :
– territoire d’observation : Préalpes du Sud depuis les Préalpes de Grasse jusqu’au Vercors.
Axes à développer :
– variante géographique à étudier dans le Gapençais, le Dévoluy et le Vercors notamment ;
– néotypification.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Archiloque A. et al., 1971 ; Archiloque A. et al., 1974 ; Barbero M., 1968, 1972 ; Lejoly J., 1975, Noble V., 2008 ; Poirion L., 1960 ; Van Es J. et al., 2014.
FICHE N° 27-30
Association : Centaureo triumfettii – Helictotrichetum sempervirentis Barbero 1968 (Bull. Soc. Bot. France 115 : 230).
Synonyme : pelouse à Avena sempervirens et Gregoria vitaliana Lejoly 1975 (Phytosociologie écologique en moyenne montagne méditerranéenne… 2 : 466) (synonyme syntaxonomique inclus provisoirement).
Unités supérieures : Eu-Helictotrichenion sempervirentis, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 4 (neotypus nominis hoc loco) du tab. 26 in Barbero (1972, Études phytosociologiques et écologiques comparées des végétations orophiles… 2 : 114) ; les données de Barbero (1968) ne contenant qu’une colonne synthétique, elles ne permettent pas d’y désigner un lectotype.
Physionomie : pelouse écorchée marquée par Helictotrichon sempervirens et/ou H. parlatorei.
Combinaison caractéristique d’espèces : Helictotrichon sempervirens, H. parlatorei, H. sedenense, H. setaceum, Cyanus triumfettii s. l., Galium obliquum, Teucrium montanum, Astragalus monspessulanus, A. sempervirens, Minuartia rostrata, M. verna, M. capillacea, Arenaria multicaulis, Linaria supina, Globularia cordifolia, Carex halleriana, Knautia collina, Ononis striata, Leucopoa dimorpha, Nepeta nepetella, Bunium bulbocastanum, Dianthus furcatus, D. pavonius, Scutellaria alpina, Anthyllis montana, Oxytropis neglecta.
Synécologie : pelouse calcicole mésoxérophile à xérophile, thermophile, centrée sur l’étage subalpin et établie sur des éboulis en voie de fixation ou des pentes rocailleuses d’adret.
Variations : -.
Synchorologie :
– territoire d’observation : Alpes sous influences ligures, atteint vers l’ouest le secteur d’Allos dans les Alpes-de-Haute-Provence.
Axes à développer :
– étude de la variabilité sur le gradient est-ouest du territoire d’observation ;
– poursuivre les travaux sur les pelouses à Helictotrichon sempervirens de l’étage subalpin des Alpes de Haute-Provence ou des Hautes-Alpes et préciser les liens avec l’association à Avena sempervirens et Gregoria vitaliana décrite par Lejoly (1975) dans la région d’Entrevaux (04), syntaxon inclus ici à titre provisoire.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1968, 1972 ; Lejoly J., 1975 ; Van Es J. et al., 2014.
FICHE N° 27-31
Association : Hieracio monregalensis – Avenetum montanae Barbero 1972 nom. corr. Van Es hoc loco.
Synonyme : Hieracio eriophylli – Avenetum montanae Barbero 1972 (Études phytosociologiques et écologiques comparées des végétations orophiles… : 211), Hieracium villosum var. eriophyllum Fr. n’est pas une combinaison valide et correspond dans sa conception à H. erioleucum Zahn, lui-même synonyme actuel de H. monregalense Burnat & Gremli.
Unités supérieures : Eu-Helictotrichenion sempervirentis, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 35 (lectotypus nominis hoc loco) du tab. 22 in Barbero (1972, Études phytosociologiques et écologiques comparées des végétations orophiles… 2 : 110).
Physionomie : pelouse écorchée dominée par Helictotrichon sedenense, Sesleria caerulea et Festuca laevigata, riche en Hieracium à indument tomenteux ou duveteux.
Combinaison caractéristique d’espèces : Helictotrichon sedenense, Hieracium monregalense, H. tomentosum, Astragalus sempervirens, A. depressus, A. australis, Campanula stenocodon, Plantago maritima subsp. serpentina, Pedicularis gyroflexa, Gentiana ligustica, Onobrychis montana, Hedysarum boutignyanum, Asperula aristata subsp. oreophila, Knautia collina, Dianthus furcatus, Iberis sempervirens, Trinia glauca, Festuca laevigata, Bupleurum ranunculoides, Carex sempervirens, Sesleria caerulea, Arenaria multicaulis, Silene acaulis, Saxifraga moschata, S. caesia, Oxytropis campestris.
Synécologie : pelouse calcicole mésoxérophile à xérophile thermophile de l’étage subalpin établie sur des rendzines au contact d’affleurements rocheux ou de pentes rocailleuses sur versants ensoleillés.
Variations :
– leontopodietosum alpini Barbero 1972 (= typicum), sous-association de l’étage subalpin supérieur et alpin inférieur annonçant le Seslerio caeruleae – Helictotrichetum sedenensis, différenciée par Leontopodium nivale subsp. alpinum, Oxytropis neglecta[7], Silene acaulis, Petrocallis pyrenaica, Dryas octopetala, Androsace vitaliana ;
– rhamnetosum alpinae Barbero 1972, sous-association plus thermophile et moins alticole annonçant le Centaureo triumfettii – Helictotrichetum sempervirentis Barbero 1968, différenciée par Helictochloa pratensis, Teucrium montanum, Astragalus monspessulanus, Rhamnus alpina.
Synchorologie :
– territoire d’observation : Alpes du Sud, principalement sous influences ligures.
Axes à développer : préciser la répartition et les liens floristico-écologiques avec le Seslerio caeruleae – Helictotrichetum sedenensis.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Van Es J. et al., 2014.
FICHE N° 27-32
Association : Festucetum dimorphae Barbero 1968 (Bull. Soc. Bot. France 115 : 232).
Synonymes : -.
Unités supérieures : Helictotrichenion sempervirentis Corriol, Sanz & J.-M. Royer, Avenion sempervirentis Barbero 1968, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : non lectotypifiable, l’auteur appuie sa description sur une liste synthétique.
Physionomie : pelouse écorchée marquée par les touffes de Leucopoa dimorpha.
Combinaison caractéristique d’espèces : Leucopoa dimorpha (= Festuca dimorpha), Nepeta nepetella, Valeriana montana, Linaria supina, Helictotrichon sempervirens, H. setaceum, H. parlatorei, Dianthus furcatus, Bupleurum ranunculoides, Hieracium tomentosum, Thymus serpyllum agg., Carduus defloratus subsp. carlinifolius, Astragalus sempervirens, Globularia cordifolia, Iberis sempervirens, Tolpis staticifolia, Rumex scutatus, Galeopsis angustifolia.
Synécologie : pelouse calcicole pionnière établie sur des éboulis en voie de fixation sur des versants chauds et ensoleillés aux étages montagnard et subalpin.
Variations : -.
Synchorologie :
– territoire d’observation : Alpes sous influences ligures, atteint vers l’ouest le secteur du col de la Cayolle dans les Alpes-de-Haute-Provence.
Axes à développer : lecto- ou néotypification.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1968, 1972 ; Van Es J. et al., 2014.
FICHE N° 27-33
Association : Gentiano occidentalis – Androsacetum villosae Corriol, F. Prud’homme & Fallour-Rubio ass. nov. hoc loco.
Synonymes : -.
Unités supérieures : Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 816 (typus nominis hoc loco) du tab. 4 hoc loco retranscrit ci-dessous :
F. Prud’homme & B. Durand, le 21.VII.2014, cabanes d’Ance, station de ski d’Arette la Pierre-Saint-Martin (Pyrénées-Atlantiques), à 1 686 m, en exposition nord, sur calcaire du Crétacé supérieur, surface 25 m², sur dalle faiblement déclive, recouvrement 60 %, 33 taxons
Asperula hirta 2, Koeleria vallesiana 2, Helianthemum oelandicum subsp. incanum 2, Thymus polytrichus 2, Valeriana montana 2, Globularia repens 2, Agrostis rupestris 2, Androsace villosa 1, Teucrium pyrenaicum 1, Arenaria multicaulis 1, Gentiana occidentalis +, Festuca gautieri subsp. scoparia +, Carex ornithopoda +, Alchemilla alpigena +, Thesium pyrenaicum +, Clinopodium alpinum +, Carduus defloratus +, Scabiosa cinerea +, Erinus alpinus +, Draba aizoides +, Thymelaea tinctoria subsp. nivalis +, Arenaria grandiflora +, Aquilegia pyrenaica +, Trisetum baregense +, Carex halleriana +, Galium pumilum +, Arabis serpyllifolia +, Hypericum nummularium +, Silene acaulis +, Hepatica nobilis +, Juniperus communis +, Phyteuma hemisphaericum +, Hieracium sp. +.
Physionomie : pelouse ouverte ou en guirlande, à floraison riche et colorée, à richesse spécifique élevée (le plus souvent entre 21-36 espèces pour une médiane de 27), généralement sans espèce fortement dominante ; photo 10.
Combinaison caractéristique d’espèces : Helianthemum oelandicum, Koeleria vallesiana, Androsace villosa, Teucrium pyrenaicum, Thymus polytrichus, Gentiana occidentalis, Anthyllis vulneraria subsp. boscii, Sesleria caerulea, Asperula hirta, Sideritis hyssopifolia, Arenaria grandiflora, Carex sempervirens subsp. sempervirens…
Synécologie : pelouse subalpine xérophile, basiphile, sur des sols peu épais (rendzines), issus de roche mère calcaire, à déneigement assez précoce, occupant des vires de paroi, des dalles et fentes de lapiaz et des éboulis calcaires fixés, principalement entre 1 620 et 1 950 m d’altitude ; ces pelouses sont en contact avec des végétations des parois calcaires de l’Asperulo hirtae – Potentilletum alchimilloidis Chouard 1949.
Variations :
– typicum, différencié par Helictotrichon sedenense, Carduus defloratus, Festuca gautieri subsp. scoparia, Thymelaea tinctoria subsp. nivalis, Euphrasia salisburgensis, Hypericum nummularium, Aster alpinus, Potentilla alchimilloides, Hieracium mixtum, Eryngium bourgatii, Thesium pyrenaicum, Arenaria multicaulis, Alchemilla alpigena, Allium lusitanicum, Gypsophila repens, Geranium cinereum…, plus alticole et psychrophile ;
– trinietosum glaucae Corriol & Dupouey subass. nov. hoc loco, différencié par Ononis striata, Teucrium chamaedrys, Trinia glauca, Asperula cynanchica, Globularia cordifolia, Helictochloa pratensis (cf. subsp. iberica) plus thermophile, sur versant fortement exposé au soleil, en dessous de 1 800 m, typus nominis hoc loco : rel. 111 du tab. 4 in Dupouey 1981 (Contribution à l’étude phytosociologique du massif des Eaux-Chaudes… inédit, = rel. 499 du tab. 4 hoc loco) retranscrit ici
versant sud de la crête de Cézy, Laruns (Pyrénées-Atlantiques), à 1 650 m, en exposition sud, sur calcaire gréseux du Santonien, surface 30 m², pente 20°, recouvrement 75 %, 30 taxons
Festuca ovina aggr. 4, Sesleria caerulea 3, Teucrium pyrenaicum 2, Koeleria vallesiana subsp. abbreviata 2, Carex sempervirens subsp. sempervirens 2, C. caryophyllea 2, Thymus polytrichus 2, Androsace villosa 1, Ononis striata 1, Trinia glauca 1, Helianthemum oelandicum subsp. incanum 1, Brimeura amethystina 1, Tractema umbellata 1, Galium pumilum aggr. 1, Orchis mascula 1, Gentiana occidentalis var. aragonensis +, Sideritis hyssopifolia +, Teucrium chamaedrys +, Helianthemum nummularium subsp. tomentosum +, Gentiana verna +, Minuartia verna +, Biscutella brevifolia +, Erinus alpinus +, Rhamnus pumila +, Carduus defloratus +, Saxifraga paniculata +, Scabiosa columbaria +, Pilosella officinarum +, Erica vagans +, Arabis sp. (cité A. stricta) +.
Synchorologie :
– territoire d’observation : versant atlantique des Pyrénées centro-occidentales, depuis le massif de la Pierre-Saint-Martin à l’ouest jusqu’à la vallée des Gaves à l’est ;
– sous-associations ou variantes géographiques : la subass. trinietosum glaucae n’est connue pour le moment que du massif de Cézy (vallée d’Ossau ; Dupouey, 1981).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Dupouey J.-L., 1981.

FICHE N° 27-34
Association : Seslerio caeruleae – Globularietum nudicaulis Nègre 1968 (Portugaliae Acta Biol., sér. B, 9 : 215).
Synonymes :
– groupement à Bartsia spicata et Scabiosa cinerea Gruber 1978 (La végétation des Pyrénées ariègeoises et catalanes occidentales : 134) nom. inval. (art. 3c) (synonyme syntaxonomique plausible) ;
– Scrophulario hoppii – Koelerietum alpicolae Nègre 1970 (Portugaliae Acta Biol., sér. B, 11 (1-2) : 118) (synonyme syntaxonomique inclus ?).
Note : l’unique relevé fourni par Nègre (1970) pour son Scrophulario hoppii – Koelerietum alpicolae est classé par notre analyse avec les deux relevés de son Seslerio – Globularietum nudicaulis, l’unique relevé de Gruber (1978, 1979) de son groupement à Bartsia spicata et Scabiosa cinerea, ainsi que deux relevés attribués par Carreras et al. (1993 : rel. 13 du tab. 50 et rel. 3 du tab. 51) au Festucetum scopariae Braun-Blanq. 1948. Il serait nécessaire de regrouper des relevés supplémentaires pour statuer plus clairement sur ces syntaxons.
Unités supérieures : Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 23 (lectotypus nominis hoc loco) in Nègre (1968, Portugaliae Acta Biol., sér. B, 9 : 216).
Physionomie : pelouse rocailleuse, rase, ouverte, sans espèce nettement dominante, riche en chaméphytes rampantes.
Combinaison caractéristique d’espèces : Gypsophila repens, Teucrium pyrenaicum, Astragalus monspessulanus, Erinus alpinus, Helianthemum nummularium (subsp. tomentosum ?), H. oelandicum, Asperula cynanchica (subsp. pyrenaica ?), Bromopsis erecta, Aster alpinus, Oxytropis campestris, Sideritis hyssopifolia, Hippocrepis comosa, Arctostaphylos uva-ursi, Biscutella cf. flexuosa, Sesleria caerulea.
Synécologie : pelouse subalpine xérophile, des crêtes calcaires ou calschisteuses, sur sol mince (rendzine), bien exposée au soleil.
Variations : un groupe de huit relevés assez pauvres, d’origines bibliographiques diverses, s’inscrit à la base de notre CAH de l’alliance, subordonné à la présente association avec laquelle ils partagent Teucrium pyrenaicum, Hippocrepis comosa, Helianthemum oelandicum, H. nummularium, Erinus alpinus… ; il pourrait s’agir de relevés appauvris et il est prématuré de leur attribuer un statut taxonomique ; ce groupe contient notamment le lectotype obligatoire du Scabioso pyrenaicae – Caricetum sempervirentis Tüxen & Oberd. 1958 décrit de la chaîne Cantabrique (Tüxen & Oberdorfer, 1958 : 164).
Synchorologie :
– territoire d’observation : décrite au versant nord des Pyrénées centrales (Luchonais ; Nègre, 1968), mais aussi relevés attribués de Gruber (1978, 1979) et Nègre (1970), ainsi que du versant sud (Carreras et al., 1993).
Axes à développer : relevés supplémentaires à réaliser dans les Pyrénées centrales, variations, chorologie et synonymie à préciser.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Carreras J. et al., 1993 ; Gruber M., 1978, 1979 ; Nègre R., 1968, 1970 ; Tüxen R. & Oberdorfer E., 1958.
FICHE N° 27-35
Association : Gypsophilo repentis – Festucetum scopariae Corriol ass. nov. hoc loco.
Note : classé à proximité, mais distinct du Scutellario alpinae – Festucetum scopariae prov., comme pour ce dernier, sont regroupés par notre analyse des relevés (seize au total), attribués par leurs auteurs à différents syntaxons. Notre analyse rattache nettement ce groupe à la sous-alliance du Saponarienion caespitosae, à proximité de l’Oxytropido neglectae – Festucetum scopariae, donc en position assez éloignée du Festucetum scopariae, malgré une origine géographique globalement plus orientale des relevés.
Synonyme : Festucetum scopariae astragaletosum catalaunici Baudière & P. Küpfer 1968 (synonyme syntaxonomique inclus max parte [rel. 5, 6, 7, 8, 9]).
Unités supérieures : Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 335 (typus nominis hoc loco) du tab. 5 hoc loco retranscrit ici :
F. Laigneau, le 5.VII.2011, en contrebas du Soum d’Arrail, vallée de Campbielh, Gèdre (Hautes-Pyrénées), à 2 227 m, en exposition sud, sur roche mère calcaire, surface 30 m², en forte pente (entre 27 et 45 °), recouvrement 70 %, 48 taxons
Helianthemum nummularium s. l. 3, Teucrium pyrenaicum (incl. *guarensis) 3, Helictotrichon sedenense 2, Koeleria vallesiana s. l. (incl. *humilis) 2, Clinopodium alpinum 2, Sesleria caerulea 2, Arenaria grandiflora 2, Festuca gautieri (*scoparia) 1, Paronychia serpyllifolia 1, Anthyllis vulneraria (*boscii) 1, Gypsophila repens 1, Poa alpina 1, Carex ornithopoda (incl. *elongata) 1, Androsace villosa 1, Asperula hirta 1, Alchemilla alpigena 1, Medicago suffruticosa *suffruticosa 1, Minuartia verna +, Festuca nigrescens (incl. *microphylla) +, F. trichophylla *asperifolia +, Saxifraga moschata +, S. intricata +, Arabis ciliata +, Thymus serpyllum agr. (incl. nervosus) +, Helianthemum oelandicum s. l. +, H. apenninum +, Cerastium alpinum +, Hypericum nummularium +, Kernera saxatilis +, Rhinanthus pumilus +, Trifolium pratense (incl. *nivale) +, T. thalii +, Carex rupestris +, Oxytropis neglecta +, Vicia pyrenaica +, Bupleurum angulosum +, Globularia nudicaulis (incl. G. gracilis) +, G. fuxensis +, Astragalus monspessulanus +, Euphrasia alpina +, Erigeron uniflorus +, Briza media 1, Cirsium acaulon 1, Luzula nutans 1, Galium papillosum +.
Physionomie : pelouse drue, ouverte, vert jaunâtre, à richesse spécifique moyennement à assez élevée (le plus souvent entre 21-43 espèces pour une médiane de 31), dominée par Festuca gautieri subsp. scoparia, parfois codominée par Helictotrichon sedenense, à floraison diversifiée et colorée, à optimum en début d’été ; photo 11.
Combinaison caractéristique d’espèces : Festuca gautieri subsp. scoparia, Sideritis hyssopifolia, Helictotrichon sedenense, Minuartia verna, Koeleria vallesiana subsp. humilis, Gypsophila repens, Clinopodium alpinum, Hippocrepis comosa, Helianthemum nummularium, Anthyllis vulneraria subsp. boscii, Gentiana verna, Arenaria multicaulis.
Synécologie : pelouse subalpine xérophile, basiphile, bien exposée, à déneigement précoce, soumise aux phénomènes périglaciaires, principalement entre 2 200 et 2 410 m d’altitude.
Variations :
– typicum, différencié par Sesleria caerulea, Paronychia serpyllifolia, Festuca nigrescens, Poa alpina, Alchemilla alpigena, Saxifraga moschata, Arabis ciliata, Carex ornithopoda…, pelouse rocailleuse mature du subalpin supérieur, riche en espèces ;
– astragaletosum catalaunici (Baudière & P. Küpfer 1968) Corriol subass. nov. et comb. nov. hoc loco [basionyme : Festucetum scopariae astragaletosum catalaunici Baudière & P. Küpfer 1968 (Bull. Soc. Neuchâtel. Sci. Nat. 91 : 78)], différencié par Astragalus sempervirens subsp. catalaunicus, Bupleurum ranunculoides subsp. gramineum, Seseli montanum subsp. nanum, Euphrasia salisburgensis, Oxytropis halleri, Campanula cochlearifolia, Crepis pygmaea, groupement pionnier sur éboulis calcaires plus ou moins stabilisés ; typus nominis hoc loco : rel. 7 d’un tab. sans n° in Baudière & Küpfer (1968) = rel. 1055 du tab. 5 hoc loco.
Synchorologie :
– territoire d’observation : Pyrénées orientales et centrales.
Axes à développer :
– déterminisme, chorologie et variabilité à préciser ;
– le cas échéant, validations et transferts des noms de syntaxons.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Baudière A. & Küpfer Ph., 1968.

FICHE N° 27-36
Association : Serratulo nudicaulis – Asperuletum pyrenaicae P. Monts. & Villar 1987 (Lazaroa 7 : 10).
Synonyme : Saponario caespitosae – Festucetum scopariae oxytropidetosum pyrenaicae Rivas Mart., Báscones, T.E. Díaz, Fern.-Gonz. & Loidi 1991 (Itinera Geobot. 5 : 338) (synonyme syntaxonomique).
Unités supérieures : Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 1 du tab. 1 in Montserrat & Villar (1987 : 15) désigné par les auteurs (= rel. 629 du tab. 6 hoc loco).
Physionomie : pelouse écorchée, à recouvrement fréquemment < 60 %, drue, vert-jaunâtre par les touffes de Festuca gautieri subsp. scoparia, à floraison variée et forte richesse spécifique (le plus souvent entre 25-48 espèces pour une médiane de 30), à optimum en début d’été.
Combinaison caractéristique d’espèces : Sideritis hyssopifolia, Festuca gautieri subsp. scoparia, F. ovina aggr. (F. altopyrenaica), Koeleria vallesiana, Helictotrichon sedenense, Pilosella lactucella, Galium pumilum (subsp. marchandii), Paronychia serpyllifolia, Androsace villosa, Anthyllis vulneraria (subsp. boscii), Trifolium thalii, Asperula cynanchica subsp. pyrenaica, Viola rupestris, Antennaria dioica, Lotus corniculatus subsp. alpinus, Gentiana verna, Oxytropis neglecta, Carduus defloratus subsp. carlinifolius, Potentilla crantzii, Arenaria grandiflora, Astragalus sempervirens subsp. catalaunicus, Vicia pyrenaica.
Synécologie : pelouse subalpine, xérophile, basiphile, sur sol peu profond, fortement exposée au vent et au soleil, à déneigement précoce et phénomènes périglaciaires intenses, principalement entre 1 950 et 2 140 m d’altitude.
Variations :
– typicum, différencié par Galium pyrenaicum, Arenaria purpurascens, Silene acaulis, Serratula nudicaulis, Jurinea humilis, négativement par Androsace vitaliana, Astragalus sempervirens subsp. catalaunicus, Carlina acaulis, à répartition exclusivement ibérique dans les Pyrénées centro-occidentales ;
– scorzoneretosum aristatae prov., différencié par Cirsium acaulon, Carthamus mitissimus, Scorzonera aristata, Bupleurum ranunculoides, Carex caryophyllea, Medicago suffruticosa, Leontodon hispidus, Gentiana occidentalis, Juniperus communis, Clinopodium alpinum, Anemone alpina, à déterminisme à préciser, d’altitude un peu plus basse (dès 1 600 m) ; noter que cette variation contient le relevé que Montserrat & Villar (1975 : 211) avaient désigné comme typus de l’alliance du Thymelaeion nivalis, désignation invalide pour une alliance ;
– polygaletosum alpinae prov., différencié par Arenaria multicaulis, Polygala alpina, Scorzoneroides pyrenaica, à déterminisme à préciser.
Voir aussi la note dans la fiche F 27-37 de l’Oxytropido neglectae – Festucetum scopariae.
Synchorologie :
– territoire d’observation : Pyrénées centro-occidentales, surtout sur le versant méridional, pénètre localement sur le versant français de la vallée d’Aspe à la vallée des Gaves à la faveur de situations exposées ;
– sous-associations ou variantes géographiques : la subass. typicum n’est pas signalée et improbable en France où c’est la subass. polygaletosum alpinae qui est probablement la plus représentée.
Axes à développer : déterminisme écologique respectif des sous-associations et distribution des sous-associations sur le versant français, validations éventuelles des noms provisoires.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Montserrat P. & Villar L., 1975, 1987 ; Rivas-Martínez S. et al., 1991.
FICHE N° 27-37
Association : Oxytropido neglectae – Festucetum scopariae Rivas Mart., Báscones, T.E. Díaz, Fern.-Gonz. & Loidi 1991 corr. Rivas Mart. et al. 2002.
Synonyme : Oxytropido pyrenaicae – Festucetum scopariae Rivas Mart., Báscones, T.E. Díaz, Fern.-Gonz. & Loidi 1991(Itinera Geobot. 5 : 332).
Unités supérieures : Saponarienion caespitosae (P. Monts. & Villar 1987) Rivas Mart., T.E. Díaz, F. Prieto, Loidi & Penas 1991, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 4 du tab. 52 in Rivas-Martínez et al. (1991 : 332) désigné par les auteurs (= rel. 749 du tab. 7 hoc loco).
Physionomie : pelouse jaunâtre, drue et plus ou moins en guirlande du fait des phénomènes périglaciaires (solifluxion), fréquemment dominée par Festuca gautieri subsp. scoparia, parfois Koeleria vallesiana ou Helictotrichon sedenense, à richesse spécifique assez élevée (le plus souvent entre 17-29 espèces pour une médiane de 23), à floraison souvent assez vive et variée, à optimum en début d’été.
Combinaison caractéristique d’espèces : Festuca gautieri subsp. scoparia, Paronychia serpyllifolia, Koeleria vallesiana (subsp. humilis), Helictotrichon sedenense, Androsace villosa, Galium pyrenaicum, Oxytropis neglecta, Anthyllis vulneraria subsp. alpestris s.l., Seseli montanum subsp. nanum. La combinaison caractéristique présente des similitudes avec celle du Serratulo nudicaulis – Asperuletum pyrenaicae P. Monts. & Villar 1987, tout en étant nettement appauvrie par rapport à cette association.
Synécologie : pelouse subalpine psychroxérophile, basiphile, de substrats rocailleux de roche mère carbonatée, de pente et durée d’enneigement modérées, soumise à des phénomènes périglaciaires d’engel-dégel, principalement entre 1 940 et 2 360 m d’altitude.
Variations :
– typicum, différencié par Pilosella lactucella, Poa molinerii, Erigeron alpinus, Festuca pyrenaica, Potentilla tabernaemontani, qui est plus proche de la subass. ranunculetosum heterocarpi, avec laquelle elle partage aussi Ranunculus parnassifolius subsp. heterocarpus, que d’un groupement modal ;
– ranunculetosum heterocarpi Benito, Aldezábal & Arbella in Benito 2006, typifié par le rel. 1 du tab. 46 in Benito (2006, Vegetación del parque nacional de Ordesa y Monte Perdido… : 151, = rel. 185 du tab. 7 hoc loco), différencié par Crepis pygmaea, Ranunculus parnassifolius subsp. heterocarpus, Carduus carlinoides, Sesleria caerulea, Saxifraga oppositifolia, faisant la transition avec les végétations d’éboulis calcaires (Thlaspietea rotundifolii), avec deux variantes, une (typique) plus froide, plus alticole ou d’exposition fraîche à Festuca pyrenaica, F. glacialis, Veronica nummularia, faisant la transition vers le Festucetum glaciali-pyrenaicae Rivas Mart. 1977, et une mieux exposée et moins alticole, à Brassica repanda, Sideritis hyssopifolia… ;
– erodietosum glandulosi Benito, Aldezábal & Arbella in Benito 2006, typifié par le rel. 1 du tab. 45 désigné par Benito (2006, Vegetación del parque nacional de Ordesa y Monte Perdido… : 151, = rel. 173 du tab. 7 hoc loco), différencié par Erodium glandulosum, Astragalus sempervirens subsp. catalaunicus, Ononis cristata, Brassica repanda, Onobrychis pyrenaica, Potentilla alchimilloides, Echium vulgare, Poterium sanguisorba, Jurinea humilis et, en commun avec d’autres subass., Thymelaea tinctoria subsp. nivalis et Sideritis hyssopifolia, la moins psychrophile et la moins alticole (entre 1 850 et 2 070 m), faisant la transition vers des groupements plus xérophiles des Ononidetalia striatae tels que le Teucrio pyrenaici – Astragaletum catalaunici Carrillo & Ninot 1990 ; noter aussi que ces trois dernières subass. sont différenciées négativement par Oxytropis neglecta et Silene acaulis ;
– thymelaeetosum nivalis prov., différencié positivement par Campanula cf. lanceolata, Gentiana verna, Taraxacum pyrenaicum, Medicago suffruticosa, Trifolium thalii et, en commun avec d’autres subass., Thymelaea tinctoria subsp. nivalis, Astragalus sempervirens subsp. catalaunicus, Sideritis hyssopifolia, Androsace vitaliana, Thymus polytrichus, et négativement par Oxytropis neglecta et Seseli montanum subsp. nanum, de déterminisme à préciser, notamment par rapport à la subass. astragaletosum gypsophili, correspondant à une partie des relevés du Thymelaeion nivalis de Montserrat & Villar (1975) ;
– astragaletosum gypsophili (Loidi & Biurrun 1998) subass. nov. et stat. nov. hoc. loco [basionyme : Astragalo teresiani – Thymelaeetum nivalis Rivas Mart., Báscones, T.E. Díaz, Fern.-Gonz. & Loidi ex Loidi & Biurrun 1998 (Lazaroa 19 : 162), typus nominis hoc loco : rel. 2 du tab. 54 in Rivas-Martínez et al. (1991), désigné pour le basionyme par Loidi & Biurrun 1998 (Lazaroa 19 : 162, = rel. 768 du tab. 7 hoc loco), différencié positivement par Astragalus monspessulanus subsp. gypsophilus, Helictochloa pratensis subsp. iberica, Carex caryophyllea et, en commun avec d’autres subass., Thymelaea tinctoria subsp. nivalis, Thymus polytrichus, à altitude plus modérée (base du subalpin) ;
– bupleuretosum graminei prov., différencié par Bupleurum ranunculoides var. gramineum, Carex humilis, Anthyllis montana, Ononis striata, Campanula cochlearifolia, Rhamnus pumila, Sempervivum montanum, Teucrium pyrenaicum subsp. guarensis, Androsace vitaliana, Helianthemum nummularium, Saponaria caespitosa, Thymelaea tinctoria subsp. nivalis, sur sol mince, rocheux, regroupant deux relevés de Benito (2006, tab. 47 : rel. 5 et 6), attribués par leur auteur à l’Oxytopido pyrenaicae – Festucetum scopariae saponarietosum caespitosae, syntaxon que notre analyse exclut de l’Oxytropido neglectae – Festucetum scopariae (voir note ci-dessous) ;
– caricetosum rupestris Arbella, Aldezábal & Benito in Benito 2006, typifié par le rel. 4 du tab. 44 désigné par Benito (2006, Vegetación del parque nacional de Ordesa y Monte Perdido… : 150, = rel. 163 du tab. 7 hoc loco), différencié par Saxifraga paniculata, Agrostis alpina, Minuartia verna, Leontopodium nivale subsp. alpinum, Arenaria multicaulis, Oxytropis foucaudii et, en commun avec d’autres subass., Carex rupestris et Globularia repens, faisant la transition vers les pelouses cryophiles des crêtes ventées (Oxytropido – Elynion Braun-Blanq. 1950) ;
– arenarietosum purpurascentis prov., différencié par Arenaria purpurascens et, en commun avec la subass. caricetosum rupestris, Carex rupestris, proche de la précédente, mais plus alticole (altitude principalement entre 2 110 et 2 450 m), dont il se différencie en outre par Androsace villosa, Galium pyrenaicum, Erigeron alpinus, Saxifraga oppositifolia ;
– helianthemetosum oelandici prov., faiblement différencié par Helianthemum oelandicum, Carex ornithopoda, Gypsophila repens, regroupant des relevés attribués par leurs auteurs à des syntaxons hétéroclites (Oxytropido pyrenaicae – Festucetum scopariae, Astragalo teresiani – Thymelaeetum nivalis, Serratulo nudicaulis – Asperuletum pyrenaicae, Saponario caespitosae – Festucetum scopariae oxytropidetosum pyrenaicae), de déterminisme à préciser, pourrait correspondre au groupement modal (altitude principalement entre 1 920 et 2 250 m).
Note complémentaire : deux autres sous-associations sont décrites, saponarietosum caespitosae Benito & Aldezábal in Benito 2006 et trifolietosum montani Aldezábal & Benito in Benito 2006 nom. inval. (art. 16) ; leurs relevés types respectifs, de même que la plupart des relevés attribués par leur auteur à ces sytaxons ainsi que deux relevés attribués par leur auteur à la subass. erodietosum glandulosi (rel. 11 et 12 du tab. 45) et trois relevés attribués à la subass. caricetosum rupestris (rel. 7, 8, 9 du tab. 44), sont exclus par notre analyse de l’Oxytropido neglectae – Festucetum scopariae et rapprochés du Serratulo nudicaulis – Asperuletum pyrenaicae P. Monts. & Villar 1987 ; cet ensemble est différencié par Plantago monosperma, Festuca nigrescens, Geranium cinereum, Agrostis capillaris, Thymelaea tinctoria subsp. nivalis, Erinus alpinus, Scorzoneroides pyrenaica, Trifolium pratense, Plantago alpina et des taxons communs au Serratulo nudicaulis – Asperuletum pyrenaicae (voir partie droite du tableau 6).
Synchorologie :
– territoire d’observation : Pyrénées centrales, du massif d’Ordessa-mont Perdu jusqu’au pic d’Orhy, surtout sur le versant méridional, mais pénétrant sur le versant français dans des secteurs bien exposés ;
– sous-associations ou variantes géographiques : les subass. plus thermophiles sont soit très rares, soit absentes sur le versant français.
Axes à développer :
– reconnaissance et distribution des sous-associations sur le versant français ;
– le cas échéant, validation des noms provisoires ;
– identification syntaxonomique des relevés de Benito exclus de l’Oxytropido – Festucetum par notre analyse.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Benito L., 2006 ; Loidi J. & Biurrun I., 1998 ; Montserrat P. & Villar L., 1975, 1987 ; Rivas-Martínez S. et al., 1991.
FICHE N° 27-38
Association : Festuco scopariae – Seslerietum caeruleae Susplugas 1942 (Commun. Stat. Int. Géobot. Médit. Montpellier 29 : 138 et tab. 9).
Synonymes : -.
Unités supérieures : Eu-Festucenion scopariae, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 6 (lectotypus nominis hoc loco) du tab. 9 in Susplugas (1942, Commun. Stat. Int. Géobot. Médit. Montpellier 29 : 139).
Physionomie : pelouse ouverte, d’éboulis calcaires fixés, dominée par Sesleria caerulea et/ou Festuca gautieri subsp. scoparia, à floraison discrète bien qu’assez variée, à richesse spécifique peu élevée (15 à 20 espèces).
Combinaison caractéristique d’espèces : Sesleria caerulea, Plantago media, Festuca gautieri subsp. scoparia, Helictotrichon sedenense, Koeleria vallesiana, Thymus praecox aggr., Anthyllis vulneraria (subsp. ?), Saxifraga oppositifolia, Carex caryophyllea, Euphrasia salisburgensis, Gentiana verna, Globularia repens, Cirsium acaulon.
Synécologie : pelouse montagnarde à subalpine xérophile, psychrophile, des substrats caillouteux et éboulis fixés, calcaires, principalement entre 1 250 et 1 700 m d’altitude.
Variations : -.
Synchorologie :
– territoire d’observation : décrite du Haut-Vallespir dans les Pyrénées orientales (Susplugas, 1942).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Susplugas J., 1942.
FICHE N° 27-39
Association : Oxytropido amethysteae – Caricetum humilis Carrillo & Soriano in Carrillo & Vigo 1997 (Memòria del Mapa de vegetació de Catalunya 1/50.000 : 73).
Synonymes : -.
Unités supérieures : Eu-Festucenion scopariae, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. en note 3 in Carrillo & Vigo 1997 (Memòria del Mapa de vegetació de Catalunya 1/50.000 : 73 ; = rel. 892 du tab. 8 hoc loco) ; attention, il est possible que les mentions pyrénéennes d’Oxytropis amethystea correspondent en réalité à O. neglecta (voir Laínz & Sáez, 1998).
Physionomie : pelouse drue, ouverte, rocailleuse, riche en chaméphytes rampantes, à floraison multicolore, souvent dominée par Carex humilis, Festuca gautieri subsp. scoparia ou Sesleria caerulea, plus rarement par Dryas octopetala, moyennement diversifiée en espèces (médiane à 18 espèces), mais variable selon les variantes, à optimum en début d’été.
Combinaison caractéristique d’espèces : Thymus nervosus, Helictotrichon sedenense, Arenaria grandiflora, Galium pyrenaicum, Helianthemum oelandicum subsp. alpestre, Festuca gautieri subsp. scoparia, Iberis saxatilis, Anthyllis vulneraria subsp. vulnerarioides, Koeleria vallesiana s. l., Viola rupestris, Carex humilis, Oxytropis amethystea (ou O. neglecta ?).
Synécologie : pelouse subalpine xérophile, ventée, des crêtes et hauts de versants, vite déneigées, soumises à de forts contrastes de température et d’intenses phénomènes périglaciaires d’engel-dégel (cryoturbation), sur sols peu évolués, minces, sur roche mère carbonatée, principalement entre 2 200 et 2 530 m d’altitude.
Variations :
– typicum [syn. : caricetosum humilis in Carrillo et al. 2000], différencié positivement par Euphrasia salisburgensis, Agrostis alpina (en commun avec la subass. caricetosum rosae), Carex sempervirens subsp. sempervirens et négativement par Festuca gautieri subsp. scoparia ;
– caricetosum rosae (Carrillo & Carreras in Carrillo, Carreras, Font, Ninot, I. Soriano & Vigo 2000) Corriol subass. nov. et stat. nov. hoc loco [basionyme : Elyno – Oxytropidetum halleri iberidetosum saxatilis Carrillo & Carreras in Carrillo, Carreras, Font, Ninot, I. Soriano & Vigo (Bull. Soc. Linn. Provence 51 : 110), typifié par le rel. 5 (typus nominis hoc loco) du tab. 6, = rel. 946 du tab. 8 hoc loco)], différencié positivement par Carex curvula subsp. rosae, C. capillaris, C. myosuroides, Agrostis alpina, Potentilla nivalis, P. crantzii et négativement par Koeleria vallesiana et Carex humilis, le plus souvent au dessus de 2 500 m d’altitude, en position plus froide, souvent de crête, faisant la transition vers l’Oxytropido – Elynion ;
– helictotrichetosum sedenensis (Carreras & Carrillo in Carrillo & Vigo 1997) Carrillo, Carreras, Font, Ninot, I. Soriano & Vigo 2000 (Bull. Soc. Linn. Provence 51 : 103i, typifié par le rel. 10 du tab. 3, = rel. 906 du tab. 8 hoc loco) (≡ Festucetum scopariae helictotrichetosum sedenensis Carreras & Carrillo in Carreras & Vigo 1997), différencié positivement par Jasione crispa, Petrocallis pyrenaica, Potentilla nivalis et négativement par Carex humilis, avec une variante typique à Saxifraga oppositifolia et Ranunculus parnassifolius subsp. heterocarpus, appauvrie en Oxytropis cf. amethystea, et une variante à Oxytropis cf. amethystea ; cette sous-association plus ouverte (< 50% de recouvrement) et plus pauvre en espèces, généralement entre 12 et 20 par relevé, vient sur des versants rocailleux ou ébouleux instables en plus forte pente ;
– asteretosum alpini Corriol subass. nov. hoc loco, typus nominis hoc loco : rel. 3 du tab. 55 in Soriano 2001 (Acta Bot. Barcinon. 47 : 320, = rel. 1081 du tab. 8 hoc loco), différencié positivement par Aster alpinus, Plantago monosperma, Daphne cneorum, Lotus alpinus, Antennaria dioica, Avenula pratensis (subsp. iberica), Oxytropis campestris, et négativement par Iberis saxatilis, incluant une variante à Dryas octopetala et Saxifraga oppositifolia, dans la frange altitudinale basse (2 200-2 300 m), de richesse spécifique plus élevée (généralement entre 17 et 26 espèces par relevé), de physionomie plus fermée (> 70% de recouvrement), sur pente faible et avec quelques espèces tolérant les humus bruts ; on peut supposer que cette variante se développe sur sol plus stable et plus humifère.
Synchorologie :
– territoire d’observation : décrit principalement des sierras calcaires des Pyrénées orientales espagnoles (hautes vallées du Sègre et du Llobrégat, Sierra del Cadi et alentours) ; dans la mesure où l’Oxytropis éponyme de cette association serait en réalité O. neglecta, celle-ci serait à rechercher en France dans les Pyrénées orientales, la combinaison caractéristique y étant plausible ;
– sous-associations ou variantes géographiques : -.
Axes à développer :
– réévaluation taxonomique de l’Oxytropis éponyme ;
– rechercher l’association au versant français des Pyrénées ;
– préciser le déterminisme écologique des différentes variantes.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Braun-Blanquet J., 1948 ; Carrillo E. & Ninot J., 1992 ; Carrillo E. & Vigo J., 1997 ; Carrillo E. et al., 2000 ; Gruber M., 1979a, b ; Ninot J.-M., 1996 ; Soriano I., 1998, 2001 ; Vigo J., 1972 ; Vigo J. & Massales R.M., 1996.
FICHE N° 27-40
Association : Scutellario alpinae – Festucetum scopariae prov. (tab. 9).
Note : ce syntaxon correspond à un groupe de 44 relevés, attribués par leurs auteurs à des syntaxons hétéroclites, dont beaucoup au Festucetum scopariae ou à des sous-associations rattachées au Festucetum scopariae (astragaletosum catalaunici Baudière & P. Küpfer 1968, iberidetosum saxatilis Gruber 1979, cirsietosum glabri Carrillo & Ninot 1992, var. à Ononis striata Vigo & Massales 1996, var. à Festuca yvesii Vigo & Massales 1996, plantaginetosum mediae (Vigo 1972) Carrillo, Carreras, Font, Ninot, Soriano & Vigo 2000, deschampsietosum flexuosae Soriano 2001, seslerietosum Soriano 2001). Notre analyse statistique des relevés pyrénéens rapproche ce groupe du Gypsophilo repentis – Festucetum scopariae au sein du Saponarienion. Toutefois, le tri du tableau synthétique général montre qu’il s’insère mieux dans le Festucenion scopariae dont il possède plusieurs taxons différentiels (Anthyllis vulneraria subsp. vulnerarioides, Jasione crispa, Festuca yvesii, Dianthus hyssopifolius). Ce rattachement est également cohérent avec l’origine orientale des relevés.
Synonymes :
– Festucetum scopariae astragaletosum catalaunici Baudière & P. Küpfer 1968 (synonyme syntaxonomique pro parte [rel. 1 et 2]) ;
– Festucetum scopariae cirsietosum glabri Carrillo & Ninot 1992 (Flora i vegetació de les Valls d’Espot I de Boí, II : 74) (synonyme syntaxonomique inclus) ;
– Festucetum scopariae adonidetosum pyrenaicae Vigo & Massales 1996 (El poblament vegetal de Vall de Ribes. Mapa de vegetacio 1:50 000 : 110) (synonyme syntaxonomique inclus) ;
– Festucetum scopariae deschampsietosum flexuosae Soriano 2001 (Acta Bot. Barcinon. 47 : 111) (synonyme syntaxonomique inclus) ;
Note : ce dernier nom a été typifié par erreur par son auteur par un relevé qu’il attribue lui-même à une autre subass. (astragaletosum catalaunici) et qui ne contient pas le taxon éponyme Avenella flexuosa (Soriano, 2001 : 111 et tab. 54 p. 318).
Unités supérieures : Eu-Festucenion scopariae, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : non typifié, nom. prov.
Physionomie : pelouse drue, ouverte, jaunâtre, à richesse spécifique moyennement à assez élevée (le plus souvent entre 17-32 espèces pour une médiane de 25), à floraison diversifié et colorée, à optimum en début d’été.
Combinaison caractéristique d’espèces : Festuca gautieri subsp. scoparia, Sideritis hyssopifolia, Galium pumilum s. l., Lotus alpinus, Euphorbia cyparissias, Arenaria grandiflora, Paronychia serpyllifolia, Scutellaria alpina, Astragalus sempervirens subsp. catalaunicus, Helictotrichon sedenense, Carduus defloratus subsp. carlinifolius.
Synécologie : pelouse subalpine xérophile, basiphile, bien exposée, à déneigement précoce, soumise aux phénomènes périglaciaires, principalement entre 1 850 et 2 400 m d’altitude pour une médiane de 2 100 m.
Variations :
– ononidetosum cristatae prov. (incl. Festucetum scopariae deschampsietosum flexuosae Soriano 2001 et Festucetum scopariae var. à Ononis striata Vigo & Massales 1996 nom. inval.), différencié par Ononis cristata, Dianthus hyssopifolius, Bupleurum ranunculoides subsp. gramineus, Iberis sempervirens, Sesleria caerulea, Androsace villosa, Clinopodium alpinum, Potentilla tabernaemontani, Ononis striata, Vicia pyrenaica, Anthyllis vulneraria subsp. vulnerarioides, Helianthemum oelandicum, bien caractérisé ; pourrait être considéré comme la subass. typicum ;
– adonidetosum pyrenaicae prov. (Festucetum scopariae adonidetosum pyrenaicae Vigo & Massales 1996), différencié positivement par Androsace vitaliana, Adonis pyrenaica, Carduus carlinoides, Erysimum duraei subsp. pyrenacium et négativement par Helictotrichon sedenense, Carduus defloratus subsp. carlinifolius, nettement plus alticole, au dessus de 2 400 m ;
– medicaginetosum suffruticosae prov. (incl. Festucetum scopariae cirsietosum glabri Carrillo & Ninot 1992), différencié surtout par Medicago suffruticosa, paraissant peu homotone.
Synchorologie :
– territoire d’observation : Pyrénées orientales et Pyrénées centrales sur le versant méridional.
Axes à développer :
– déterminisme, chorologie et variabilité à préciser ;
– le cas échéant, validations et transferts des noms de syntaxons.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Baudière A. & Küpfer Ph., 1968 ; Carreras J. et al., 1993 ; Carrillo E. & Ninot J., 1992 ; Carrillo E. et al., 2000 ; Gruber M., 1979b ; Soriano I., 2001 ; Vigo J. & Massales R.M., 1996.
FICHE N° 27-41
Association : Festucetum scopariae Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 151).
Synonyme : Seslerio caeruleae – Festucetum scopariae Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 151) nom. inval. (art. 3b).
Unités supérieures : Eu-Festucenion scopariae, Festucion scopariae Braun-Blanq. 1948, Astragaletalia sempervirentis Barbero 1968.
Type nomenclatural : rel. 12 du tab.19 in Carrillo et al. (2000), lectotypus désigné par ces auteurs p. 98.
Physionomie : pelouse drue et même piquante, ouverte, laissant apparaître le sol caillouteux, dominée par des guirlandes serrées ou des puissantes touffes vert jaunâtre de Festuca gautieri subsp. scoparia et d’Helictotrichon sedenense, généralement riche en espèces (le plus souvent entre 25 et 50 pour une médiane de 33), à floraison multicolore, à optimum en début d’été ; les types biologiques très largement co-dominants sont les chaméphytes naines et les hémicryptophytes.
Combinaison caractéristique d’espèces : Plantago monosperma, Sideritis hyssopifolia, Helianthemum oelandicum (subsp. alpestre), Koeleria vallesiana subsp. humilis, Ononis cristata, Daphne cneorum, Vicia pyrenaica, Anthyllis vulneraria subsp. vulnerarioides, Draba aizoides, Aster alpinus, Festuca gautieri subsp. scoparia, Helictotrichon sedenense, Thymus nervosus, Minuartia verna, Gentiana verna, Arenaria grandiflora, Galium pyrenaicum, Sesleria caerulea.
Synécologie : pelouse subalpine mésoxérophile, des versants bien exposés, vite déneigés, sur sol caillouteux, peu humifère, mais pouvant atteindre quelques dizaines de centimètres de profondeur, issu de roche mère carbonatée, notamment par suite de la fixation d’anciens éboulis calcaires, soumise à des phénonènes périglaciaires d’engel-dégel, particulièrement aux intersaisons, principalement entre 2 040 et 2 450 m d’altitude.
Variations :
– typicum, différencié positivement par Helictochloa pratensis subsp. iberica, Cirsium acaulon, Carlina acaulis, Bupleurum ranunculoides var. gramineum, Carex humilis, Festuca ovina aggr., Trifolium pratense, Taraxacum pyrenaicum, Euphrobia cyparissias, Paronychia serpyllifolia…, avec deux variantes, une typique à Androsace villosa, Onosma tricerosperma subsp. fastigiata[8], Phyteuma orbiculare, Arabis corymbiflora, Oxytropis campestris, Draba aizoides, Carduus defloratus, Colchicum bulbocodium, Trinia glauca, et une appauvrie, diférenciée essentiellement négativement (positivement par Campanula rotundifolia aggr.) ;
– iberidetosum saxatilis Gruber ex Corriol subass. nov. hoc loco, typus nominis hoc loco : rel. 8 du tab. 1 in Gruber (1979, Bull. Soc. Bot. France 126 : 246-247, = rel. 611 du tab. 10 hoc loco), différencié par Iberis saxatilis, Eryngium bourgatii, Clinopodium alpinum, Carduus defloratus subsp. carlinifolius, Oxytropis cf. amethystea[9], Carex sempervirens, Biscutella laevigata aggr., Anthyllis montana, Teucrium pyrenaicum…, plus thermophile, localisé au Cadi ;
– trifolietosum thalii Gruber ex Corriol subass. nov. hoc loco, typus nominis hoc loco : rel. 11 du tab. 1 in Gruber (1979, Bull. Soc. Bot. France 126 : 246-247, = rel. 615 du tab. 10 hoc loco), différencié positivement par Silene acaulis, Trifolium thalii, Saxifraga oppositifolia, Festuca nigrescens et négativement par l’absence d’Anthyllis vulneraria subsp. vulnerarioides, en climat plus humide, avec trois variantes, une typique à Helianthemum nummularium, Polygala alpina, Iberis sempervirens, Taraxacum pyrenaicum et sans Sesleria caerulea, une variante à Jasione crispa, Bupleurum ranunculoides, Erigeron alpinus, plus pauvre en espèces (autour de 25), et une variante à Primula elatior subsp. intricata, Polygala calcarea, Cirsium acaulon, Plantago media, Ranunculus bulbosus et Sesleria caerulea ;
– oxytropidetosum halleri Corriol subass. nov. hoc loco, typus nominis hoc loco : rel. 4 du tab. 19 in Braun-Blanquet (1948, Mon. Est. Pir. Inst. Esp. Edafol. Fisiol. Veg. 9 : 152, = rel. 774 du tab. 10 hoc loco), différencié positivement par Oxytropis halleri, Helianthemum nummularium, Sedum atratum, Alyssum montanum aggr.[10], Astragalus australis, Herniaria alpina, Iberis sempervirens, Poa molinerii… et négativement par l’absence d’Helianthemum oelandicum, Vicia pyrenaica, Carex caryophyllea, Potentilla crantzii, Sesleria caerulea, Lotus corniculatus, plus cryophile, entre 2 400 et 2 700 m d’altitude, faisant la transition avec les pelouses alpines de l’Oxytropido – Elynion ;
– astragaletosum monspessulani prov., regroupant deux relevés intitulés par Braun-Blanquet (1948 : 160) « sous-ass. à Festuca ovina ssp. indigesta[11], différenciée positivement par Astragalus monspessulanus, Allium sphaerocephalon, Helianthemum nummularium et négativement par l’absence d’Helictotrichon sedenense, Thymus nervosus, Arenaria grandiflora, moins alticole et plus thermophile.
La subass. seslerietosum caeruleae Soriano 2001 (Acta Bot. Barcinon. 47 : 110 ; rel. 779 à 547 à droite du tab. 10) est considérée par Soriano (2001 : 110) comme un groupement de transition entre le Festucion scopariae Braun-Blanq. 1948 et les pelouses plus mésophiles du Primulion intricata ; elle est exclue du Festucetum scopariae d’après notre analyse statistique, qui la positionne dans tous les dendrogrammes obtenus dans une branche affiliée au Salicion pyrenaicae ; son positionnement dans cette alliance nécessiterait une recombinaison au rang d’association végétale ; toutefois, la combinaison caractéristique d’une telle association serait entièrement incluse dans celle du Festucetum scopariae, sans aucune différentielle positive ; pour cette raison, nous nous abstenons de faire cette recombinaison mais considérons cette subass. comme extrêmement marginale et très appauvrie dans l’alliance du Festucion scopariae ; sa classification serait peut-être à ré-évaluer sur la base d’un plus grand échantillonnage de relevés ; différentielles négatives : Koeleria vallesiana, Draba aizoides, Aster alpinus, Anthyllis vulneraria subsp. vulnerarioides, Ononis cristata, Daphne cneorum, Plantago monosperma ; richesse spécifique plus faible (le plus souvent entre 13 à 25 espèces) ; pelouse subalpine, xérophile, psychrophile, des versants sur sols minces issus de roche mère calcaire, peu ensoleillés, où elle remplace le Festucetum scopariae Braun-Blanq. 1948, principalement entre 1 850 et 2 300 m d’altitude.
Noter qu’un groupe de relevés (nos 1, 2, 3, 6, 8, 9) issus du tableau original du Saponario caespitosae – Festucetum scopariae de Gruber (1979b) est classé dans notre analyse en variation du Festucetum scopariae (transition vers cette association ?).
Notes complémentaires :
– le Festucetum scopariae caricetosum montanae Vigo & Massales 1996 est d’après notre analyse exclu de la classe et devrait être versé dans les Festuco – Brometea ;
– le relevé type du Festucetum scopariae plantaginetosum mediae (Vigo 1972) Carrillo, Carreras, Font, Ninot, Soriano & Vigo 2000 est exclu des Elyno – Seslerietea d’après notre analyse et représente avec douze autres relevés issus de la bibliographie une association végétale inédite appartenant selon nous aux Ononidetalia striatae.
Synchorologie :
– territoire d’observation : en France, uniquement dans le département des Pyrénées-Orientales, en Cerdagne et Conflent ; mieux représenté dans les Pyrénées catalanes sur le versant sud (Sierra de Cadi, Andorre, val de Ribes, val d’Aran, massif d’Aigues-Tortes) (Braun-Blanquet, 1948 ; Vigo, 1972 ; Gruber, 1979a, 1979b ; Carrillo & Ninot, 1992 ; Vigo & Massales, 1996 ; Soriano, 1998 ; Carrillo et al., 2000 ; Soriano, 2001) ;
– sous-associations ou variantes géographiques : la subass. iberidetosum saxatilis n’est signalée qu’en Sierra de Cadi, la subass. astragaletosum monspessulani prov. n’est connue que des environs de Núria ; la subass. seslerietosum caeruleae décrite de Cerdagne (Sierra de Cadí ; Soriano, 2001), à laquelle se rattachent aussi les relevés attribués au Festucetum scopariae 1 et 9 de Braun-Blanquet (1948 : 152), 1 et 8 de Carreras et al. (1993, tab. 50), est présente jusqu’au massif duTurbón (rel. 6 du tab. 1 in Ninot, 1988 : 682) et la haute Ribagorça en Aragon (rel. 1 et 8 du tab. 50 in Carreras et al., 1993 : 311) ; à rechercher ailleurs dans les Pyrénées, sa présence côté français est probable.
Axes à développer :
– préciser le déterminisme des différentes variations mises en évidence ;
– préciser la variabilité et la position syntaxonomique de la subass. seslerietosum caeruleae Soriano 2001.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.434 ; EUNIS : E4.434.
Bibliographie
Braun-Blanquet J., 1948 ; Carreras J. et al., 1993 ;Carrillo E. & Ninot J., 1992 ; Carrillo E. et al., 2000 ; Gruber M., 1979a, b ; Ninot J.-M., 1988 ; Soriano I., 1998, 2001 ; Vigo J., 1972 ; Vigo J. & Massales R. M., 1996
FICHE N° 27-42
Association : Gentiano acaulis – Primuletum intricatae Vigo 1972 (Pirineos 105 : 51).
Note : cette association a une place très marginale au sein des pelouses orophiles basiphiles pyrénéennes, ce qu’avait déjà bien identifié Dupouey (1986) ; elle est mise à l’écart dès la première classification générale (méthode de Ward), au même titre que les associations relevant des Ononidetalia striatae ; son rattachement à une autre classe de végétation ne semble pourtant pas possible, ce qui nous est confirmé par J.-M. Royer (comm. pers.) en ce qui concerne les Festuco – Brometea ; si l’on force son maintien dans les CAH suivantes, elle sort par défaut dans le groupe du Festucion scopariae au sens large ; l’insertion de sa colonne synthétique dans le tableau général trié-ordonné montre effectivement quelque rapport floristique avec le bloc du Festucion scopariae ; l’appauvrissement en taxons méridionaux xérophiles (dû à l’écologie plus mésophile) et le lot important de différentielles acidiphiles rendent pourtant difficile l’assimilation de cette association au Festucion scopariae ; elle s’insère encore plus mal floristiquement dans le Primulion intricatae dans lequel elle a été traditionnellement rangée, ce que toutes nos classifications automatiques montrent ; en conséquence nous choisissons comme solution d’attente de ranger cette association dans un nouveau syntaxon de fin de classe (Gentiano acaulis – Primulion intricatae), positionné ici provisoirement à proximité du Festucion scopariae.
Synonymes : -.
Unités supérieures : Gentiano acaulis – Primulion intricatae Corriol & J.-M. Royer.
Type nomenclatural : rel. 11 (lectotypus nominis hoc loco) du tab. II in Vigo (1972).
Physionomie : pelouse dense, fermée, assez luxuriante, généralement dominée par Festuca nigrescens et Carex montana, à richesse spécifique très élevée (souvent plus de 40 espèces), richement fleurie, à optimum en début d’été.
Combinaison caractéristique d’espèces : Gentiana acaulis, Primula elatior subsp. intricata, Carex montana, C. umbrosa subsp. huetiana, Koeleria macrantha, Trifolium montanum, Festuca nigrescens, Thymus pulegioides, Cirsium acaulon, Gymnadenia nigra, Helictochloa pratensis (subsp. ?), Hypochaeris maculata, Pilosella lactucella, Potentilla crantzii, Antennaria dioica, Eryngium bourgatii, Carduus defloratus subsp. carlinifolius, Anthyllis vulneraria subsp. vulnerarioides, Sesleria caerulea, Coeloglossum viride, Aster alpinus, Myosotis alpestris, Potentilla erecta…
Synécologie : pelouse subalpine mésophile, neutro-acidiclinophile, des sols profonds développés sur roche mère carbonatée, mais décalcifiés en surface, des versants en pente modérée (10-25°), en exposition fraîche, souvent au nord, principalement entre 1 750 et 2 000 m d’altitude.
Variations : on peut constater dans le tableau princeps un faciès à Sesleria caerulea sans doute plus calcicole.
Synchorologie :
– territoire d’observation : Pyrénées orientales.
Axes à développer : position syntaxonomique.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Vigo J., 1972.
FICHE N° 27-43
Association : Geo pyrenaici – Caricetum sempervirentis Chouard 1943[12].
Synonymes :
– Ass. à Carex sempervirens et Geum pyrenaicum Chouard 1943 (Bull. Soc. Bot. France 90 : 25) ;
– Semperviretum pyrenaicum Chouard 1943 (Bull. Soc. Bot. France 90 : 25) nom. inval. (art. 43) ;
– Carici sempervirentis – Geetum pyrenaici sensu Vanden Berghen 1970 (Bull. Centr. Etudes Rech. Sci. Biarritz 8 (2) : 294) (synonyme syntaxonomique inclus) ;
– Veronico gouanii – Salicetum pyrenaicae Nègre 1970 (Portugaliae Acta Biol., sér. B, 11 (1-2) : 32) (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Salicenion pyrenaicae, Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : la description originale de Chouard (1943) n’étant constituée que d’une liste synthétique, il faudrait désigner comme néotype un relevé en provenance de Gavarnie (locus typicus) appartenant à la subass. typicum et comprenant les deux taxons éponymes.
Physionomie : pelouse verdoyante, luxuriante, fermée, formant des cordons plus ou moins étroits sur des vires ou des fentes de parois, ou occupant des surfaces modestes sur des versant de pieds de falaise ou d’éboulis stabilisés, fréquemment dominée par Carex sempervirens subsp. sempervirens, Salix pyrenaica ou Alchemilla alpigena, à richesse spécifique variable, mais pouvant être élevée, à floraison assez vive et multicolore, à optimum estival, superposée à une strate bryophytique fortement développée.
Combinaison caractéristique d’espèces : Carex sempervirens, Salix pyrenaica, Alchemilla alpigena, Betonica alopecuros, Luzula nutans, Helianthemum nummularium (subsp. tomentosum), Sesleria caerulea, Horminum pyrenaicum, Aquilegia pyrenaica, Bistorta vivipara, Veronica ponae, Geum pyrenaicum, Parnassia palustris et Globularia nudicaulis (rare dans la subass. thalictretosum macrocarpi).
Synécologie : pelouse subalpine psychrophile, mésophile à hygroclinophile, chionoclinophile, basiphile, des vires et replats des parois calcaires et pentes caillouteuses en exposition fraîche ou masquée du soleil, sur des sols humifères, principalement entre 1 640 et 2 100 m d’altitude pour une médiane de 1 850 m.
Variations :
– typicum, différencié par Dryas octopetala, Anemone hepatica, Linum catharticum, Ranunculus thora, Tractema umbellata…, à richesse spécifique élevée (le plus souvent entre 21-43 espèces pour une médiane de 33), des Pyrénées centrales sous influence océanique, principalement entre 1 640 et 1 930 m d’altitude ;
– anthyllidetosum boscii Carreras, Carrillo & Ninot 1996 (Anales Jard. Bot. Madrid 54 : 504), typifié par le rel. 5 du tab. 2 des auteurs (= rel. 155 du tab. 11 hoc loco), différencié positivement par Plantago alpina, Carex ornithopoda, Poa alpina, Galium pumilum, Scorzoneroides pyrenaica, Saxifraga umbrosa, Arenaria multicaulis, Anthyllis vulneraria subsp. boscii (aussi bien représenté dans la subass typicum), et négativement par Horminum pyrenaicum, à richesse spécifique élevée (30-42), défini par ses auteurs comme une variation des Pyrénées centrales, plus abrité des influences océaniques (val d’Aran, Bielsa) ;
– salicetosum reticulatae (Nègre 1970) Corriol subass. nov. et stat. nov. hoc. loco [basionyme : Veronico gouanii – Salicetum pyrenaicae Nègre 1970 (Portugaliae Acta Biol., Sér. B, 11 (1-2) : 32)], typus nominis hoc loco : rel.168 du tab. 7 de Nègre (1970) (= rel. 687 du tab. 11 hoc loco), différencié positivement par Salix reticulata, Saxifraga paniculata, Festuca pumila[13], Campanula scheuchzeri et négativement par l’absence d’Horminum pyrenaicum, Geum pyrenaicum, Carduus defloratus, localisé au subalpin supérieur (2 050-2 200 m), plus chionophile, décrit sur la base d’une race moins atlantique, en position plus centrale dans les Pyrénées ;
– thalictretosum macrocarpi Corriol, Fallour-Rubio & F. Prud’homme subass. nov. hoc. loco, différencié par Carduus defloratus, Thalictrum macrocarpum, Hypericum nummularium, Polystichum lonchitis, Geranium sylvaticum, Valeriana montana…, correspondant au moins en partie aux relevés synthétisés par Vanden Berghen (1970) sous le nom de Cariceto sempervirentis – Geetum pyrenaici, d’altitude similaire à celle du type, surtout dans les fentes des parois et lapiaz, à répartition plus occidentale et richesse spécifique un peu moindre (17-33 pour une médiane de 22), typus nominis hoc loco : rel. 859 du tab. 11 hoc loco, retranscrit ici
D. Fallour-Rubio, le 11.VIII.2009, station de ski d’Arette la Pierre-Saint-Martin (Pyrénées-Atlantiques), à 1 700 m, en exposition nord, sur calcaire du Crétacé supérieur, surface 4 m², pente entre 45° et 70°, recouvrement 100 %, 20 taxons
Carex sempervirens 4, Salix pyrenaica 3, Horminum pyrenaicum 2, Geum pyrenaicum 2, Sesleria caerulea 1, Betonica alopecuros 1, Carduus defloratus (incl. C. medius) 1, Centaurea gr. nigra 1, Galium mollugo 1, Thesium pyrenaicum 1, Hypericum nummularium 1, Aquilegia pyrenaica +, Parnassia palustris +, Bartsia alpina +, Ranunculus thora +, Polystichum lonchitis +, Thalictrum macrocarpum +, Anemone alpina s. l. +, Vaccinium myrtillus +, Potentilla alchimilloides +.
On observe une certaine variabilité, avec une partie du tableau à relevés appauvris dans laquelle on peut identifier une variante plus saxicole à Coincya monensis subsp. cheiranthos, Dethawia splendens, Armeria pubinervis, Euphorbia pyrenaica, Buglossoides gastonii…
Synchorologie :
– territoire d’observation : Pyrénées occidentales et centrales, du massif du Pic d’Anie jusqu’au val d’Aran au moins ;
– sous-associations ou variantes géographiques : anthyllidetosum bosci restreinte aux Pyrénées centrales plus abritées, thalictretosum macrocarpi restreinte aux Pyrénées occidentales, jusqu’au val d’Azun.
Axes à développer : néotypification de l’association et du typicum avec un relevé de Gavarnie.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Carreras J. et al., 1996 ; Chouard P., 1943 ; Nègre R., 1970 ; Vanden Berghen C., 1970.
FICHE N° 27-44
Association : Geranio cinerei – Ranunculetum gouanii Gruber 1978 (La végétation des Pyrénées ariègeoises et catalanes occidentales : 127 et tab. 30).
Synonymes : –.
Unités supérieures : Eu-Salicenion pyrenaicae, Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 7 (lectotypus nominis hoc loco) du tab. 3 in Gruber (1979, Ecol. Medit. 4 : 91), identique au rel. 5 du tab. 30 in Gruber (1978, thèse).
Physionomie : pelouse verdoyante, plus ou moins ouverte, parfois caillouteuse, riche en espèces (32 à 45), dominée par les hémicryptophytes, même si des chaméphytes sont présentes, sans espèce dominante, assez richement fleurie, à optimum estival (avec toutefois une phénophase vernale à Primula elatior subsp. intricata).
Combinaison caractéristique d’espèces : Primula elatior subsp. intricata, Geranium cinereum, Trifolium thalii, Potentilla crantzii, Arenaria multicaulis, Clinopodium alpinum, Erigeron alpinum, Lotus alpinus, Alchemilla alpigena, Salix pyrenaica, Dryas octopetala, Trifolium badium, Arabis corymbiflora, Sedum atratum, Bistorta vivipara, Helictotrichon sedenense, Gentianella campestris, Festuca nigrescens,Carex sempervirens, Saxifraga moschata, Sempervivum arachnoideum, Euphrasia alpina...
Synécologie : pelouse mésophile à hygroclinophile, moyennement chionophile, de l’étage subalpin supérieur, des replats humides et des faibles pentes en ombrée, développée sur roche mère carbonatée, mais sol humifère plus ou moins épais, de pH à tendance neutre.
Variations : -.
Synchorologie :
– territoire d’observation : Pyrénées centro-orientales, Ariège, jusqu’au port de Saleix à l’est, massif d’Aygues-Tortes ? (les relevés de Gruber, 1978, 1979, en provenance du val d’Aran sont exclus de l’association par notre analyse) ; semble prendre le relais du Geo pyrenaici – Caricetum sempervirentis à l’est de son aire de répartition.
Axes à développer : chrologie et variabilité à préciser sur la base de nouveaux relevés.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Gruber M., 1978, 1979.
FICHE N° 27-45
Association : Dryado octopetalae – Salicetum pyrenaicae Chouard 1943 (Bull. Soc. Bot. France 90 : 27).
Synonyme : Alchemillo plicatulae – Dryadetum octopetalae Soriano 1998 (J. Bot. Soc. Bot. France 5 : 23) (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Salicenion pyrenaicae, Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : la description originale de Chouard (1943) n’étant constituée que d’une liste synthétique, nous désignons ici un néotype également en provenance de Gavarnie, donc dans le territoire princeps ; neotypus nominis hoc loco : rel. 17 du tab. 48 in Rivas-Martínez et al. (1991, Itinera Geobot. 5 : 319, = rel. 531 du tab. 12 hoc loco).
Physionomie : chaméphytaie verdoyante, fermée, dominée ou co-dominée par Dryas octopetala et Salix pyrenaica, à floraison plutôt discrète hormis les tapis de fleurs blanches du Dryas et les grandes hampes florales de Anemone alpina, moyennement diversifiée en espèces (médiane à 21 espèces), mais variable selon les variantes, à optimum estival à tardi-estival.
Combinaison caractéristique d’espèces : Salix pyrenaica, Dryas octopetala, Sesleria caerulea, Carex sempervirens subsp. sempervirens, Bistorta vivipara, Alchemilla alpigena[14], Helictotrichon sedenense et dans une moindre mesure Silene acaulis, Geranium cinereum.
Synécologie : landine occupant, en fonction des variantes, l’étage subalpin jusqu’à la base de l’étage alpin, psychrophile à cryophile, moyennement à faiblement chionophile, sur des sols minces ou squelettiques, issus de roche mère carbonatée, constituant un climax stationnel.
Variations : malgré une variabilité assez importante constatée pour cette association, nous préférons traiter les différentes variations au rang infra-association, car elle demanderait dans le cas contraire d’éclater le Dryado – Salicetum pyrenaicae en nombreuses associations distinctes plus ou moins bien caractérisées et à déterminisme chorologique peu net. La subass. typicum est basée sur la branche de notre classification qui contient la liste synthétique de Chouard (1943) (et également celui de Vanden Berghen, 1970), en ne tenant compte que des fréquences > II. Ne s’agissant pas d’un relevé élémentaire, la chose est toujours délicate et, si l’on regarde toutes les fréquences du relevé de Chouard, il est probable qu’une partie de la variation de l’association y est comprise ;
– typicum, différencié par Ranunculus thora, Anemone alpina, Gentiana verna, Soldanella alpina, Hypericum nummularium, Primula integrifolia, Aquilegia pyrenaica, modal, à optimum à l’étage subalpin moyen, entre 1 750 et 2 200 m, pour une médiane de 1 850 m, dans les Pyrénées centrales ; photo 12 ;
– leontopodietosum alpini Corriol subass. nov. hoc loco, différencié par Leontopodium nivale subsp. alpinum, Anthyllis vulneraria subsp. boscii, Gentianella campestris, Festuca nigrescens, Erigeron alpinus, Euphrasia minima, Carex myosuroides, Antennaria dioica, Aster alpinus, Thalictrum alpinum, Thesium pyrenaicum, moins chionophile et donc en position plus froide, souvent de crête rocheuse, bien drainée, à richesse spécifique plus élevée (24-34), et apparition de quelques acidiphiles probablement liées à l’accumulation d’humus brut dans ces conditions microclimatiques ; typus nominis hoc loco : rel. 158 du tab. 12 hoc loco retranscrit ici
G. Corriol, le 11.VIII.2003, rive gauche du vallon d’Estaubé, Gèdre (Hautes-Pyrénées), à 1 700 m, en exposition nord-est, sur calcaire du Dévonien, surface 3 m², pente 70 %, recouvrement 70 %, 24 taxons
Dryas octopetala 3, Carex sempervirens 3, Anthyllis vulneraria subsp. boscii 2, Salix pyrenaica 1, Sesleria caerulea 1, Bistorta vivipara 1, Gentiana occidentalis 1, Androsace villosa 1, Festuca nigrescens 1, Briza media 1, Alchemilla alpigena +, Leontopodium nivale subsp. alpinum +, Antennaria dioica +, Gentianella campestris +, Aster alpinus +, Silene acaulis +, Minuartia verna +, Arenaria multicaulis +, Juniperus communis f. prostrée +, Saxifraga paniculata +, Asperula hirta +, Helianthemum nummularium +, Koeleria vallesiana +, Plantago media + ;
– caricetosum rupestris Corriol subass. nov. hoc loco, différencié positivement par Carex rupestris, Helianthemum oelandicum, Anthyllis vulneraria subsp. vulnerarioides, Poa molinerii, Dethawia splendens, Androsace villosa, Oxytropis campestris, Minuartia verna…, le plus alticole et cryo-xérophile, situé à la base de l’étage alpin, principalement entre 2 300 et 2 400 m, à richesse spécifique moyenne (généralement entre 16 et 24 espèces), faisant la transition vers les pelouses calcicoles cryophiles alpines de l’Oxytropido campestris – Caricetum rupestris ; photo 13 ; typus nominis hoc loco : rel. 955 du tab. 12 hoc loco retranscrit ici
Pène Nègre, à la Mongie, Bagnères-de-Bigorre (Hautes-Pyrénées), à 2 370 m en exposition nord-nord-est, sur calcaire du Dévonien, surface 5 m², pente 70 %, recouvrement 95 %, hauteur végétative 5 cm, 26 taxons
Dryas octopetala 3.3, Carex rupestris 3.3, Sesleria caerulea 2.2, Oxytropis neglecta 2.2, Helictotrichon sedenense 1.1, Aquilegia pyrenaica 1.1, Silene acaulis 1.2, Carex myosuroides 1.2, Minuartia verna 1.1, Helianthemum oelandicum var. piloselloides 1.2, Androsace villosa 1.2, Thymus nervosus 1.1, Salix pyrenaica +, Agrostis schleicheri +, Bistorta vivipara +, Sedum atratum +, Festuca gautieri subsp. scoparia +, Arctostaphylos uva-ursi +, Erigeron alpinus +, Juniperus communis f. prostrée +, Poa alpina +, Valeriana apula +, Lotus corniculatus subsp. alpinus +, Paronychia kapela subsp. serpyllifolia +, Festuca glacialis +, Galium pumilum aggr. + ;
– salicetosum reticulatae Carreras, Carrillo & Ninot 1996, typifié par le rel. 6 du tab. 1 in Carreras et al. (1996, Anales Jard. Bot. Madrid 54 : 510, = rel. 962 du tab. 12 hoc loco), différencié par Arctostaphylos alpinus, Salix reticulata, Rhododendron ferrugineum, Bartsia alpina, Euphrasia salisburgensis, Vaccinium uliginosum subsp. microphyllum, Anemone narcissiflora, Carex ornithopoda, Tofieldia calyculata, Parnassia palustris…, en commun avec la subass. typicum, Primula integrifolia, Ranunculus thora, Soldanella alpina et en commun avec la subass. leontopodietosum Festuca nigrescens, Selaginella selaginoides, Anthyllis vulneraria subsp. boscii ; nettement plus riche en espèces (24 à 32 pour les six relevés concernés), en limite d’association, tend à basculer vers le Geo pyrenaici – Caricetum sempervirentis en fonction des classifications automatiques obtenues, maintenu ici pour des raisons structurelles (chaméphytaie) ; est plus chionohygrophile, sur substrat à accumulation d’humus brut sur la rocaille calcaire, faisant la transition avec les landines riches en Ericacées du Salici pyrenaicae – Arctostaphyletum alpini Rivas Mart. et al. 1991 (Loiseleurio-Vaccinion), bien illustrées par les relevés de Fromard (1982).
Reste un lot de relevés appauvris (12-24 espèces pour une médiane de 18), tout au plus différencié positivement par Saxifraga oppositifolia et dans une moindre mesure par Jasione crispa et Pedicularis pyrenaica, auquel appartiennent entre autres la majorité des relevés attribués par Soriano (1998) à l’Alchemillo plicatulae – Dryadetum ; il pourrait s’agir pour partie de relevés fragmentaires et pour partie de groupements non saturés, situés en périphérie de leur optimum climatique comme le suggère Soriano (1998 : 24) pour ses relevés du versant ibérique des Pyrénées orientales, opinion également formulée par Carreras et al. (1996 : 510) ; notons par ailleurs que, dans cet ensemble, le relevé type de l’Alchemillo plicatulae – Dryadetum fait la transition vers le typicum ; pour ces raisons, nous préférons ne pas nommer à part ce groupement mais le considérer comme une simple forme appauvrie de la subass. typicum.
Noter enfin que le relevé type du Carici parviflorae – Salicetum retusae salicetosum pyrenaicae Rivas Mart. et al. 1991 (rel. 323 du tab. 12 hoc loco), qui ne contient ni Carex parviflora ni Salix retusa et dont nous avions obtenu l’exclusion des Salicetea herbaceae dans notre synthèse de cette classe (Corriol & Mikolajczak, 2014 : 32), se classe dans un groupe de relevés appauvris rattaché au Dryado – Salicetum pyrenaicae à proximité de la subass. caricetosum rupestris (dix derniers relevés, nos 102 à 73 à droite du tab. 12 hoc loco), groupe intermédiaire avec l’Oxytropido – Elynion.
Synchorologie :
– territoire d’observation : ensemble de la chaîne pyrénéenne dans les hauts massifs atteignant l’étage subalpin ; mieux développé dans les secteurs sous influence océanique ;
– sous-associations ou variantes géographiques : -.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Benito J. L., 2006 ; Carreras J. et al., 1996 ; Chouard P., 1943 ; Rivas-Martínez S. et al., 1991 ; Soriano I., 1998 ; Vanden Berghen C., 1970.


FICHE N° 27-46
Association : Ranunculo thorae – Seslerietum caeruleae Vigo in Rivas Mart. et al. 2002 (Itinera Geobot. 15 (1) : 196).
Synonymes : Ranunculo thorae –Seslerietum caeruleae Vigo 1979 (Folia Bot. Misc. 1 : 9) nom. inval. (art. 5).
Unités supérieures : Ranunculo thorae – Seslerienion caeruleae Peyre & Font ex Lazare 2017, Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 1 du tab. 1 in Vigo (1979, Folia Bot. Misc. 1 : 9), désigné par Rivas-Martínez et al. (2002 : 196).
Physionomie : pelouse luxuriante, verte et fermée, à allure de pelouse-ourlet, notamment du fait de la présence de Laserpitium nestleri ainsi que de quelques espèces forestières (Anemone hepatica, Convallaria majalis…) et de mégaphorbiaie (Allium victorialis, Pedicularis foliosa, Astrantia major…), à floraison riche, dominée par le blanc et le jaune, à optimum en début d’été ; la richesse spécifique semble moins élevée que pour les autres associations de l’alliance, mais apparaît variable dans le tableau et pose question sur les surfaces relevées (le plus souvent entre 11-28 espèces pour une médiane de 17).
Combinaison caractéristique d’espèces : Sesleria caerulea, Valeriana montana, Festuca gautieri subsp. scoparia, Cruciata glabra, Ranunculus thora, Anemone alpina (subsp. font-queri), A. hepatica, Laserpitium nestleri, Pedicularis foliosa.
Synécologie : pelouse subalpine psychrophile, chionoclinophile, basiphile, des vires et pieds de parois et petits cirques entourés de parois calcaires, masqués du soleil, principalement entre 1 450 et 1 800 m d’altitude pour une médiane de 1 700 m.
Variations :
– typicum, pelouse-ourlet aérohygrophile des vires d’ombrée ;
– avenuletosum pratensis Vigo & Massales 1996 (El poblament vegetal de Vall de Ribes… : 112), typifié par le rel. 1 du tab. 64 des auteurs, p. 302, différencié positivement par Leucanthemum maximum, Helictochloa pratensis (subsp. ?), Koeleria macrantha, Bupleurum ranunculoides, Euphorbia angulata, Briza media, Trifolium montanum, Eryngium bourgatii… et négativement par l’absence de Laserpitium nestleri, Pedicularis foliosa, Veronica urticifolia, moins masqué du soleil et moins hygrophile.
Synchorologie :
– territoire d’observation : sierras pré-pyrénéennes catalanes, depuis le Ripollès jusqu’au Turbón (Carrillo & Ninot, 1992 ; Carrillo et al., 2000 ; Ninot, 1996 ; Soriano, 2001 ; Vigo, 1979 ; Vigo & Massales, 1996) ; en France, occurrence probable au moins dans les Pyrénées orientales.
Axes à développer :
– à rechercher sur le versant français ;
– variations à préciser notamment au regard de la grande variabilité de richesse spécifique constatée parmi les relevés existants.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Carrillo E. & Ninot J. M., 1992 ; Carrillo E. et al., 2000 ; Ninot J.M., 1996 ; Soriano I., 2001 ; Vigo J., 1979 ; Vigo J. & Massales R.M., 1996.
FICHE N° 27-47
Association : Armerio orissonensis – Alchemilletum amphisericeae Lazare 2017 (Botanique 3 : 56).
Synonymes : -.
Unités supérieures : Ranunculo thorae – Seslerienion caeruleae Peyre & Font ex Lazare 2017, Salicion pyrenaicae Vigo in Rivas Mart., T.E. Díaz, Fern.-Gonz., Izco, Loidi, Lousã & Penas 2002, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 4 du tab. 6 in Lazare (2017, Botanique 3 : 57) désigné par l’auteur.
Physionomie : pelouse luxuriante, formant des banquettes verdoyantes et couvrantes sur gradins rocheux, à floraison colorée en début d’été, à richesse spécifique modérée pour l’alliance (entre 12 et 21 espèces).
Combinaison caractéristique d’espèces : Festuca ochroleuca subsp. ochroleuca, Alchemilla amphisericea, Armeria pubinervis subsp. orissonensis, Sesleria caerulea, Scabiosa cinerea, Saxifraga hirsuta subsp. paucicrenata, Anemone hepatica, Carex ornithopoda subsp. ornithopoda.
Synécologie : pelouse montagnarde psychrophile, basiphile, des vires calcaires exposées au nord, masquées du soleil, autour de 1 250 m d’altitude sur son site de description.
Variations : -.
Synchorologie :
– territoire d’observation : uniquement connu sur le site où il a été décrit, au sommet d’Errozaté, en basse Navarre (Pyrénées-Atlantiques) (Lazare, 2017).
Axes à développer : chorologie et variations à préciser en recherchant cette pelouse ailleurs dans le Pays basque espagnol.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Lazare J.-J., 2017.
FICHE N° 27-48
Association : Festuco commutatae – Trifolietum thalii Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 179).
Synonymes :
– Oxytropido foucaudii – Elynetum myosuroidis plantaginetosum mediae Benito 2006 (synonyme inclus) ;
– Oxytropido foucaudii – Elynetum myosuroidis sensu Benito 2006 pro parte (synonyme inclus) ;
– ? ass. à Scabiosa velutina et Crepis blattarioides Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 181) (synonyme inclus ?).
Commentaire : l’unique relevé fourni par Braun-Blanquet (1948 : 181) pour cette dernière association se trouve dans notre CAH à proximité immédiate de tous les relevés du Festuco commutatae – Trifolietum thalii de la circonscription originale ; peut-être n’est-ce qu’une forme ourlifiée de cette association.
Unités supérieures : Eu-Primulenion intricatae, Primulion intricatae Braun-Blanq. ex Vigo 1972, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. D (lectotypus nominis hoc loco) in Braun-Blanquet (1948, Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 179, = rel. 791 du tab. 13 hoc loco).
Physionomie : pelouse généralement fermée, verdoyante, fréquemment pâturée étant donné son appétance, généralement dominée par Festuca nigrescens, souvent co-dominée par Trifolium thalii, Lotus corniculatus ou Plantago alpina, à floraison multicolore, à optimum en début d’été, mais souvent discrète (notamment du fait du pâturage), à richesse spécifique moyenne (le plus souvent entre 18 à 28 espèces pour une médiane de 23).
Combinaison caractéristique d’espèces : Festuca nigrescens, Poa alpina, Lotus coniculatus, Trifolium thalii, Helictotrichon sedenense, Plantago alpina, Potentilla crantzii, Gentiana verna et, dans une moindre mesure, Bistorta vivipara, Ranunculus gouanii…
Synécologie : pelouse de l’étage subalpin supérieur et de la base de l’étage alpin, mésophile, neutrophile à acidiclinophile, développée sur roche mère carbonatée ; les altitudes sur l’ensemble des relevés considérés s’échelonnent de (1 820) 2 050-2 360 (2 450) m, pour une médiane de 2 220 m.
Variations : notre classification regroupe dans une même ramification les différentes sous-associations décrites pour le Festuco – Trifolietum thalii. S’y insèrent également d’autres relevés attribués différemment par leurs auteurs. Il s’y distribue notamment une partie des relevés attribués par Benito (2006) à l’Oxytropido foucaudii – Elynetum. Le tout présente une certaine hétérogénéité. Dans l’état actuel des connaissances, nous considérons le Festuco – Trifolietum dans un sens large et présentant donc une assez grande variabilité. Un traitement plus homogène nécessiterait un démantèlement en plusieurs associations qui nous semble prématuré.
– typicum, différencié par Primula elatior subsp. intricata, Soldanella alpina, Trifolium badium, Alchemilla alpigena, Cerastium arvense, Botrychium lunaria ; dépressions fraîches, légèrement enneigées, sur roche mère carbonatée, à sol riche en matière organique, neutre à légèrement acide, pouvant être enrichi par les déjections des troupeaux ;
– euphrasietosum salisburgensis Corriol subass. nov. hoc. loco, typus nominis hoc loco : rel. 5 du tab. 7 in Klein (1979, Phytocoenologia 5 (2) : 185, sous le pas de Gerbats, Gèdre (Hautes-Pyrénées), à 2 400 m, en exposition sud-ouest, surface 4 m², pente 20°, recouvrement 100 %, = rel. 702 du tab. 13 hoc loco) ; différencié par Erigeron uniflorus subsp. aragonensis, Galium anisophyllon, Euphrasia salisburgensis, Clinopodium alpinum, Paronychia serpyllifolia, Carex pyrenaica, Phleum alpinum s. l, Draba aizoides, Carlina acaulis, Agrostis rupestris, Medicago suffruticosa… ; proche de la subass. typicum avec laquelle elle partage en outre les différentielles Alchemilla alpigena, Cerastium arvense, Botrychium lunaria, Galium anisophyllon, Helianthemum nummularium, mais plus riche en espèces (24-33), à déterminsime à préciser (Klein, 1979 : 147 évoque un caractère plus chionophile en position plus occidentale que le type dans les Pyrénées, avec notamment Carex pyrenaica ou Gnaphalium supinum, mais en même temps une exposition plus ensoleillée favorisant la pénétration d’espèces plus thermophiles comme Medicago suffruticosa) ;
– caricetosum myosuroidis (Benito 2006) prov. [syn. : Oxytropido foucaudii – Elynetum myosuroidis plantaginetosum mediae Benito 2006 (Vegetación del parque nacional de Ordesa … : 158), typifié par le rel. 6 du tab. 54 p. 293, = rel. 293 du tab. 13 de la présente publication), différencié par Plantago media, Carex myosuroides, C. sempervirens, Carduus defloratus subsp. carlinifolius, Sesleria caerulea, Briza media, Trifolium montanum, Androsace villosa, Colchicum montanum, Poterium sanguisorba, Hippocrepis comosa, Oxytropis neglecta, Anemone alpina…, des convexités à déneigement plus précoce, subalpine, à une altitude moindre (relevés entre 1 840 et 2 150 m) ; nous ne validons pas le transfert dans le Festuco – Trifolietum thalii pour ce nom en raison des incertitudes évoquées plus haut autour de la circonscription de cette association ; en revanche, le syntaxon de Benito (2006) est clairement situé en dehors de l’Oxytropido foucaudii – Kobresietum, ce qui n’est pas surprenant étant donné l’écologie indiquée par son auteur, sur convexités peu inclinées à l’étage subalpin, contrairement à l’Oxytropido foucaudii – Kobresietum, association cryophile, chionophobe et alpine des reliefs vite déneigés ;
– thalictretosum alpini Corriol subass. nov. hoc. loco, typus nominis hoc loco : rel. 4 du tab. 49 in Benito (2006, Vegetación del parque nacional de Ordesa … : 284), versant du Tobacor en face de Góriz et S. Custodia (Aragon), le 18.VIII.1993, sol profond sur roche calcaire et dolomie, à 2 360 m, en exposition sud-ouest, surface 25 m², pente 5°, recouvrement 100 % (= rel. 221, tab. 13 hoc loco), différencié par Thalictrum alpinum, Taraxacum pyrenaicum, Campanula scheuchzeri, Colchicum montanum, Agrostis capillaris, Armeria bubanii, Myosotis alpestris, Euphrasia minima, Carex parviflora, Nardus stricta, Potentilla brauniana, Gentiana nivalis, comportant deux sous-groupes, dont l’un est appauvri en ces cinq derniers taxons, regroupant la plupart des relevés attribués par Benito (2006) au Festuco – Trifolietum thalii typicum ; possiblement plus chionophile que la subass. geranietosum cinerei dont elle partage l’aire de répartition ;
– geranietosum cinerei Benito 2006 (Vegetación del parque nacional de Ordesa … : 155), typifié par le rel. 8 du tab. 50 de Benito, 2006, p. 286, = rel. 240 du tab. 13 hoc loco), différencié positivement par Geranium cinereum, Astragalus alpinus, Primula integrifolia, ainsi que, en commun avec la subass. thalictretosum alpini, Scorzoneroides pyrenaica et dans une moindre mesure Thalictrum alpinum, groupement mésophile, propre aux massifs calcaires des Pyrénées centrales ;
– silenetosum acaulis prov., différenciée par Silene acaulis, Agrostis alpina, Daphne cneorum, Minuartia verna et, en commun avec la subass. geranietosum cinerei, Primula integrifolia et, avec la subass. arenarietosum moehringioidis, Pilosella lactucella, Plantago monosperma, et négativement par Plantago alpina, qui correspond à un groupe de relevés attribués à des syntaxons hétéroclites, y compris au Festuco – Trifolietum thalii, et essentiellement dans les Pyrénées catalanes ; semble correspondre au pendant de l’arenarietosum moehringioidis dans la partie orientale des Pyrénées, mais il est possible que ce groupe de relevés soit hétérogène ;
– arenarietosum moehringioidis Benito 2006 (Vegetación del parque nacional de Ordesa … : 155), typifié par le rel. 4 du tab. 51 de Benito, 2006, p. 288, = rel. 253 du tab. 13 hoc loco), différencié positivement par Festuca gautieri subsp. scoparia, Luzula spicata, Arenaria purpurascens, Leucanthemopsis alpina, Ranunculus parnassifolius subsp. heterocarpus, Trifolium alpinum, Seseli montanum, Pilosella lactucella, Antennaria dioica, Plantago monosperma, Androsace vitaliana, Galium pyrenaicum et négativement par l’absence de Bistorta vivipara, Plantago alpina, Gentiana verna, plus ouvert, des sols plus rocailleux et érodés.
Synchorologie :
– territoire d’observation : sur une grand partie de la chaîne pyrénéenne, depuis la Cerdagne à l’est jusque à l’ouest des Hautes-Pyrénées au moins ; au sud, cité jusqu’au Turbón ;
– sous-associations ou variantes géographiques : typicum connu des Pyrénées orientales aux Pyrénées centrales ; euphrasietosum salisburgensis connu dans les Pyrénées centrales (de la vallée d’Aure à Gèdre) ; arenarietosum moehringioidis : Pyrénées centrales (massif de Gavarnie – Mont-Perdu) ; silenetosum acaulis : surtout Pyrénées catalanes, jusqu’au Turbón en Espagne (toutefois également un relevé en provenance des Pyrénées-Atlantiques est attribué à ce groupe) ; geranietosum cinerei : Pyrénées centrales (massif de Gavarnie – Mont-Perdu) ; thalictretosum alpini : Pyrénées centrales (massif de Gavarnie – Mont-Perdu) ; caricetosum myosuroidis : Pyrénées centrales (massif de Gavarnie – Mont-Perdu), jusqu’au massif d’Aiguestortes au moins dans les Pyrénées catalanes (relevés 3 et 4 du tab. 42 du Festuco – Trifolietum thalii de Carrillo & Ninot, 1992).
Axes à développer : circonscription et variabilité à préciser.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4142 ; EUNIS : E4.414.
Bibliographie
Benito J.L., 2006 ; Braun-Blanquet J., 1948 ; Carreras J. et al., 1993 ; Carrillo E. & Ninot J.M., 1992 ; Gruber M., 1978, 1979 ; Klein J.-C., 1979 ; Ninot J.M., 1988.
FICHE N° 27-49
Association : Primulo intricatae – Horminetum pyrenaici Lazare & Mauric 1986 (Actes du Colloq. internat. de botanique pyrénéenne, la Cabanasse : 416).
Synonyme : Helictotricho sedenensis – Bellardiochloetum violaceae Lazare & Mauric 1986 (Actes du Colloq. internat. de botanique pyrénéenne, la Cabanasse : 414) (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Primulenion intricatae, Primulion intricatae Braun-Blanq. ex Vigo 1972, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 7 du tab. 2 in Lazare & Mauric (1986, Actes du Colloq. internat. de botanique pyrénéenne, la Cabanasse : 417, = rel. 582 du tab. 14 hoc loco) désigné par les auteurs.
Physionomie : pelouse généralement fermée, verdoyante, marquée à son optimum par la floraison violette de l’Horminelle, plus terne dans sa subass. bellardiochloetosum (qui en constitue un faciès dominé par Bellardiochloa variegata), à richesse spécifique moyenne (principalement entre 18-30 taxons pour une médiane de 23), à optimum en début d’été, mais à phénophase vernale, à la fonte des neiges, constellée des corolles jaune soufre de Primula elatior accompagnées de celles plus discrètes, violacées, des Soldanelles ; photo 14.
Combinaison caractéristique d’espèces : Horminum pyrenaicum, Alchemilla alpigena, Primula elatior subsp. intricata, Festuca nigrescens[15], Trifolium thalii, Soldanella alpina, Ranunculus gouanii, Armeria pubinervis, Luzula nutans, Trifolium pratense (var. ?), Poa alpina.
Synécologie : pelouse subalpine aérohygrophile, psychrophile, sur sols issus de roche mère calcaire, pouvant être superficiellement enrichis en matière organique, principalement entre 1 650 et 2 200 m d’altitude.
Variations :
– typicum, différencié par Carduus carlinoides, Taraxacum panalpinum cf.[16], Conopodium majus et, en commun avec la subass. suivante qui en est floristiquement très proche, Vicia pyrenaica, Cerastium arvense, Silene vulgaris, Pedicularis sylvatica, Phleum alpinum, Myosotis alpestris, Arabis serpyllifolia ; étage subalpin moyen à supérieur, dans les couloirs frais à enneigement prolongé, à horizon humifère restant longtemps imbibé ;
– bellardiochloetosum variegatae (Lazare & Mauric 1986) Corriol subass. nov. et stat. nov. hoc loco [basionyme : Helictotricho sedenensis – Bellardiochloetum violaceae Lazare & Mauric 1986 (Actes du Colloq. internat. de botanique pyrénéenne, la Cabanasse : 414 et tab. 1 p. 415), typus nominis hoc loco : rel. 3 du tab. 1 in Lazare & Mauric (1986, = rel. 573 du tab. 14 hoc loco)] , différencié par Bellardiochloa variegata, Scabiosa columbaria, Arenaria multicaulis, à proximité de la subass. précédente sur des croupes ou convexités exposées au vent, plus rapidement déneigées et aux sols plus filtrants ;
– primuletosum integrifoliae Corriol subass. nov. hoc. loco, différencié par Primula integrifolia, Selaginella selaginoides, Carex sempervirens subsp. sempervirens, Scorzoneroides pyrenaica, Carex ornithopoda, Salix reticulata, S. pyrenaica, Bistorta vivipara, plus chionophile, typus nominis hoc loco : rel. 42 du tab. 3 in Dupouey (1981, Contribution à l’étude phytosociologique du massif des Eaux-Chaudes, inédit ; = rel. 521 du tab. 14 hoc loco), retranscrit ici :
vallon à l’ouest de la crête d’Arcizette, Laruns (Pyrénées-Atlantiques), à 2 115 m, en exposition nord-ouest, sur calcaire gréseux du Santonien, surface 6 m², pente 30°, recouvrement 75 %, 25 taxons
Horminum pyrenaicum 2, Festuca cf. nigrescens 2, Trifolium thalii 2, Primula elatior *intricata 1, Ranunculus montanus aggr. (incl. gouanii) 1, Geum montanum 1, Carex macrostylon 1, Plantago alpina 1, Armeria pubinervis 1, Poa alpina 1, Primula integrifolia 1, Galium pyrenaicum 1, Alchemilla alpigena +, Helictotrichon sedenense +, Soldanella alpina +, Pedicularis pyrenaica +, Selaginella selaginoides +, Carex sempervirens +, C. ornithopoda (incl. *elongata) +, Gentiana verna +, Thymus polytrichus +, Sagina procumbens +, Scorzoneroides pyrenaica +, Salix reticulata +, Taraxacum cf. panalpinum x ;
– plantaginetosum mediae Corriol subass. nov. hoc. loco, différencié par Plantago media, Carduus defloratus, Achillea millefolium, Carex caryophyllea, Helleborus viridis subsp. occidentalis, sur la frange altitudinale inférieure (sommet de l’étage montagnard, base du subalpin), typus nominis : rel. 9 du tab. 2 in Lazare & Mauric (1986, Actes du Colloq. internat. de botanique pyrénéenne, la Cabanasse : 417, = rel. 584 du tab. 14 hoc loco).
Reste enfin un groupe de relevés (rel. 519 à 740 à droite du tab. 14) peu différenciés, peut-être fragmentaires, mais rattachables à cette association, issus pour la plupart de Rivas-Martínez et al. (1991 : 323).
Synchorologie :
– territoire d’observation : hautes vallées d’Ossau et d’Aspe, massif du Pic d’Anie et du Pic d’Aspe (Lazare & Mauric, 1986 ; Rivas-Martínez et al., 1991), massif du Cézy (Dupouey, 1981) ; probablement représenté ailleurs dans les hautes vallées navarraises et la partie occidentale de l’Aragon sur le versant espagnol ; constitue un vicariant du Festuco commutatae – Trifolietum thalii Braun-Blanq. 1948 dans Pyrénées occidentales.
Axes à développer :
– chorologie et variabilité de la subass. bellardiochloetosum décrite sur seulement cinq relevés et sur un tout petit territoire en haute vallée d’Ossau, non signalée depuis ;
– limite orientale à préciser pour l’association à l’est de la vallée d’Ossau.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Dupouey J.-L., 1981 ; Lazare J.-J. & Mauric A., 1986 ; Rivas-Martínez S. et al., 1991.

FICHE N° 27-50
Association : Alchemillo plicatulae – Adonidetum pyrenaicae Braun-Blanq. ex Gruber 1978 (La végétation des Pyrénées ariègeoises et catalanes occidentales : 131 et tab. 32).
Synonymes :
– gpt. à Adonis pyrenaica et Primula intricata Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 182) nom. inval. (art. 3c) ;
– Primulo intricatae – Adonidetum pyrenaicae Braun-Blanq. ex Carrillo & Ninot 1992 (Flora I vegetació de les Valls d’Espot I de Boí II : 76) nom. superfl.
Unités supérieures : Eu-Primulenion intricatae, Primulion intricatae Braun-Blanq. ex Vigo 1972, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. (lectotypus nominis obligatoire) in Braun-Blanquet (1948, Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 182-183).
Physionomie : végétation luxuriante et fermée, à physionomie d’ourlet, dominée par Adonis pyrenaica, à floraison spectaculaire, jaune vif, à optimum en début d’été, à richesse spécifique peu élevée (environ 10 à 20 espèces), occupant de petite combes caillouteuses.
Combinaison caractéristique d’espèces : Adonis pyrenaica, Alchemilla alpigena, Vicia pyrenaica, Trifolium thalii, Myosotis alpestris, Alchemilla fissa, Poa alpina, Taraxacum pyranaicum, Urtica dioica, Primula elatior subsp. intricata, Polystichum lonchitis.
Synécologie : végétation subalpine calcicole, chionoclinophile, des couloirs frais peu déclives et combes masquées, caillouteuses ou ébouleuses, entre 2 000 et 2 300 m d’altitude.
Variations : -.
Synchorologie :
– territoire d’observation : Pyrénées orientales (vallée d’Eyne, Sierra del Cadi), jusqu’au massif du Turbón au versant sud dans les Pyrénées centrales (Braun-Blanquet, 1948 ; Gruber, 1978, 1979 ; Carrillo & Ninot, 1992).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4142 ; EUNIS : E4.414.
Bibliographie
Braun-Blanquet J., 1948 ; Carrillo E. & Ninot J. M., 1992 ; Gruber M., 1978, 1979.
FICHE N° 27-51
Association : Anthyllido vulnerarioidis – Plantaginetum alpinae Corriol ass. nov. hoc loco.
Synonymes : -.
Unités supérieures : Eu-Primulenion intricatae, Primulion intricatae Braun-Blanq. ex Vigo 1972, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural, typus nominis hoc loco : rel. 88b du tab. 7 in Labroue (1976, Étude écologique des sols alpins du pic du Midi de Bigorre : 219, = rel. 417 du tab. 15 hoc loco), retranscrit ici
pic du Midi de Bigorre (Hautes-Pyrénées), 2 650 m, en exposition nord-est, surface 10 m², pente 45°, recouvrement 90 %, 23 taxons
Potentilla crantzii 2, Plantago alpina 2, Trifolium thalii 2, Festuca nigrescens 2, Anthyllis vulneraria *vulnerarioides 1, Lotus corniculatus *alpinus 1, Alchemilla alpigena 1, Primula elatior *intricata 1, Carex curvula *rosae 1, Geum montanum 1, Poa alpina (incl. *brevifolia) +, Gentiana verna +, Astragalus alpinus +, Taraxacum ‘pyrenaicum’ +, Helianthemum nummularium s. l. +, Medicago suffruticosa *suffruticosa +, Gnaphalium supinum +, Trisetum baregense +, Potentilla brauniana +, Primula integrifolia +, Myosotis alpestris +, Draba aizoides +.
Physionomie : pelouse rase, relativement fermée, bien verte et richement fleurie, souvent dominée ou co-dominée par Plantago alpina, Potentilla crantzii ou Helianthemum nummularium, à richesse spécifique moyenne (principalement entre 18-28 taxons pour une médiane de 23), à optimum estival.
Combinaison caractéristique d’espèces : Poa alpina, Anthyllis vulneraria subsp. vulnerarioides, Potentilla crantzii, Carex curvula subsp. rosae, Draba aizoides, Lotus corniculatus subsp. alpinus, Gentiana verna, Myosotis alpestris, Plantago alpina, Taraxacum cf. panalpinum[17] et dans une moindre mesure Astragalus alpinus.
Synécologie : pelouse alpine mésophile, à tendance chionophile, sur sol caillouteux mais relativement profond et disposant d’une certaine réserve en eau, à horizon superficiel plus ou moins enrichi en matière organique, sur matériaux riches en bases, principalement entre 2 450 et 2 800 m d’altitude.
Variations :
– typicum, différencié positivement par Geum montanum, Potentilla brauniana, Trifolium thalii, Carex pyrenaica, C. parviflora, Nardus stricta, Ranunculus gouanii, Trifolium pratense, négativement par Oxytropis neglecta, Erigeron uniflorus, Helictotrichon sedenense, Arenaria multicaulis ; bas de versants à sol épaissi, en exposition fraîche, à enneigement prolongé, à horizon superficiel acidifié enrichi en matière organique ;
– scabiosetosum cinereae Corriol subass. nov. hoc loco, différencié positivement par Scabiosa cinerea, Carex sempervirens, et négativement par Primula integrifolia, Minuartia sedoides, Festuca glacialis, dans la frange altitudinale inférieure de l’association, en dessous de 2 650 m ; typus nominis : rel. 24b du tab. 4 in Labroue (1976, Étude écologique des sols alpins du pic du Midi de Bigorre : 216, = rel. 391 du tab. 15 hoc loco), retranscrit ici
pic du Midi de Bigorre (Hautes-Pyrénées), à 2 600 m, en exposition nord-est, surface 5 m², pente 35°, recouvrement 80 %, 28 taxons
Helianthemum nummularium s. l. 4, Plantago alpina 2, Festuca nigrescens 2, Poa alpina (incl. *brevifolia) 1, Anthyllis vulneraria *vulnerarioides 1, Helictotrichon sedenense 1, Oxytropis neglecta 1, Scabiosa cinerea 1, Carex curvula *rosae 1, Thymus nervosus 1, Sempervivum tectorum 1, Saxifraga moschata 1, Draba aizoides +, Medicago suffruticosa *suffruticosa +, Lotus corniculatus *alpinus +, Androsace vitaliana s. l. +, Cerastium arvense (*strictum) +, Primula elatior *intricata +, Gnaphalium supinum +, Trisetum baregense +, Gentiana verna +, Bartsia alpina +, Myosotis alpestris +, Arenaria multicaulis +, Alchemilla alpigena +, Erigeron uniflorus +, Potentilla crantzii + ;
en plus de la variante typique, on note une variante pionnière appauvrie sur éboulis fixés, différenciée par Cerastium alpinum, Carduus carlinoides, Vicia pyrenaica, Veronica fruticans (cf. subsp. cantabrica)… ;
– silenetosum acaulis Corriol subass. nov. hoc loco, différencié positivement par Silene acaulis, Hornungia alpina, Pedicularis pyrenaica, Linaria alpina, Veronica nummularia, Saxifraga moschata, Festuca pyrenaica et négativement par l’absence d’Helianthemum nummularium, Medicago suffruticosa, Festuca nigrescens, Trisetum baregense, Primula elatior subsp. intricata, Gnaphalium supinum, Alchemilla alpigena, plus cryophile, dans la frange altitudinale supérieure de l’association, au dessus de 2 600 m (transition vers l’Oxytropido – Elynion), typus nominis hoc loco : rel. 60 du tab. 6 in Labroue (1976, Étude écologique des sols alpins du pic du Midi de Bigorre : 218, = rel. 410 du tab. 15 hoc loco), retranscrit ici
pic du Midi de Bigorre (Hautes-Pyrénées), à 2 780 m, en exposition sud, surface 25 m², pente 32°, recouvrement 80 %, 28 taxons
Potentilla crantzii 3, Helictotrichon sedenense 2, Festuca glacialis 2, Lotus corniculatus *alpinus 2, Taraxacum ‘pyrenaicum’ 2, Anthyllis vulneraria *vulnerarioides 1, Minuartia sedoides 1, Saxifraga moschata 1, Carex curvula *rosae 1, Poa alpina (incl. *brevifolia) 1, Plantago alpina 1, Gentiana verna +, Geum montanum +, Arenaria multicaulis +, Silene acaulis +, Hornungia alpina +, Myosotis alpestris +, Erigeron uniflorus +, Primula integrifolia +, Minuartia verna +, Pedicularis pyrenaica +, Armeria alpina +, Leucanthemopsis alpina +, Scorzoneroides pyrenaica +, Veronica nummularia +, Linaria alpina s. l. +, Draba aizoides +.
Synchorologie : pic du Midi de Bigorre (Labroue, 1976) ; probablement bien représentée sur les hauts massifs des Pyrénées centrales.
Axes à développer :
– chorologie et variabilité sur son aire de répartition ;
– vérification de l’identification du Taraxacum (Labroue cite T. pyrenaicum) et de la fétuque rouge (Labroue cite « une sous-espèce de Festuca rubra ».
Correspondances : HIC : 6170 ; CORINE biotopes : 36.4112 ; EUNIS : E4.4112.
Bibliographie
Labroue L., 1976.
FICHE N° 27-52
Association : Festuco violaceae – Trifolietum thalii Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2): 244).
Synonymes : -.
Unités supérieures: Festuco violaceae – Trifolienion thalii Van Es & Sanz, Primulion intricatae Braun-Blanq. ex Vigo 1972, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 2 (lectotypus nominis hoc loco) du tab. 9 in Braun-Blanq. & Jenny (1926, Denksch. Schweiz. Naturf. Ges. 63 (2) : 245).
Physionomie : pelouse fermée à Festuca violacea riche en dicotyélédones à floraisons colorées ; photo 15.
Combinaison caractéristique d’espèces : Festuca violacea (s. l), Sesleria caerulea, Trifolium thalii, T. badium, Linum alpinum, Crepis aurea, Astragalus alpinus, Trollius europaeus, Oxytropis campestris, Luzula spicata, Pedicularis verticillata, P. rostratospicata subsp. helvetica, Gentiana verna, Scabiosa lucida, Phyteuma orbiculare, Aster alpinus, Onobrychis montana, Lathyrus occidentalis, Anemone narcissiflora, Poa alpina, Phleum rhaeticum, Potentilla crantzii, Viola calcarata.
Synécologie : pelouse mésophile mésotrophile des étages subalpins supérieur et alpin, neutrobasiphile à acidiclinophile, installée sur faibles pentes ou replats assez longuement enneigés, sur des sols profonds et relativement frais.
Variations : nombreuses, selon la durée d’enneigement, le pH du sol ou encore l’usage pastoral :
– variante plus chionophile à Plantago serpentina distinguée par Guinochet (1938), formant un tapis plus ras ;
– variante acidiclinophile, différenciée par des espèces du Campanulo barbatae – Potentillion aureae, notamment Potentilla aurea ;
– variante fortement pâturée, différenciée par l’abondance de Poa alpina, Ranunculus sartorianus et Phleum rhaeticum.
Synchorologie :
– territoire d’observation : toutes les grandes Alpes, plus rare dans les Préalpes du Nord.
Axes à développer :
– classement phytosociologique difficile, initialement classée dans le Seslerion caeruleae par Braun-Blanquet, cette pelouse a ensuite été classée dans le Caricion ferrugineae (Mikolajczak et al., 2014) ou dans le Poion alpinae (Prunier et al., 2019) au sein des Arrhenatheretea elatioris ; la dominance des espèces des Oxytropido – Elynetalia nous incite à la classer dans cette alliance ;
– poursuite de l’étude de la variabilité.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Braun-Blanquet J. & Jenny H., 1926 ; Guinochet M., 1938 ; Mikolajczak A., 2018 ; Mikolajczak A. et al., 2014 ; Villaret J.-C., 2007.

FICHE N° 27-53
Association : Salici pyrenaicae – Bistortetum viviparae Corriol ass. nov. hoc loco.
Synonyme : Carici parviflorae – Salicetum retusae salicetosum pyrenaicae sensu Carrillo & Ninot 1992 pro max. parte (Flora I vegetació de les Valls d’Espot I de Boí II : 249, rel. 4, 5, 6, 7).
Unités supérieures : Eu-Oxytropido – Elynenion myosuroidis, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 5 (typus nominis hoc loco) du tab. 1 in Mustin & Baudière (1986, Veröff. Geobot. Inst. E.T.H. Stiftung Rübel Zürich 87 : 294, = rel. 353 du tab. 16 hoc loco).
Physionomie : microphorbiaie verdoyante, fermée ou un peu ouverte, riche en chaméphytes rampantes, à physionomie marquée par les feuilles ovales ou arrondies des saules, Bistorta vivipara, Soldanella, Bartsia, Alchemilla, plus ou moins riche en espèces (le plus souvent entre 13 et 26 pour une médiane de 18), à floraison multicolore (Saxifraga, Gentiana, Soldanella, Silene acaulis, Myosotis, Oxytropis), fortement marquée par Dryas octopetala dans la subass. dryadetosum, à optimum estival à tardi-estival ; photo 16.
Combinaison caractéristique d’espèces : Bistorta vivipara, Helictotrichon sedenense, Poa alpina, Gentiana verna, Salix reticulata, S. retusa, S. pyrenaica, Bartsia alpina, Alchemilla alpigena, Silene acaulis, Saxifraga aizoides et, dans une moindre mesure, Festuca glacialis, Oxytropis neglecta et Carex parviflora.
Synécologie : pelouse de l’étage subalpin supérieur chionophile des combes à neige et bas de versants exposés au nord ou masqués du soleil, des sols souvent caillouteux, issus de roche mère carbonatée, mais pouvant être légèrement décalcifiés en surface, à horizon superficiel soumis à des phénomènes de solifluxion, principalement localisée entre 2 000 et 2 300 m d’altitude, pour une médiane de 2 140.
Variations :
– typicum, différencié par Dryas octopetala, Carex curvula subsp. rosae, C. myosuroides, C. ericetorum Myosotis alpestris, Saxifraga oppositifolia, Anthyllis vulneraria, Oxytropis campestris, O. halleri, Astragalus alpinus…, sur zones convexes et perméables, à tendance cryophile, plus typique de l’alliance de l’Oxytropido – Elynion, avec une variante à Iberis sempervirens, Luzula nutans, Pedicularis pyrenaica… sur humus brut.
– soldanelletosum alpinae Corriol subass. nov hoc loco, différencié positivement par Hornungia alpina, Soldanella alpina, Taraxacum pyrenaicum/panalpinum, Arenaria purpurascens, Linaria alpina, appauvri et en limite d’alliance, typus nominis hoc loco : rel. 5 du tab. 56 in Carrillo & Ninot (1992, Flora I vegetació de les Valls d’Espot I de Boí II : 249, = rel. 324 du tab. 16 hoc loco), avec deux variantes, une typique faisant la transition vers le Salicion pyrenaicae et une plus chionophile, enrichie en Gnaphalium supinum, Ranunculus alpestris, Galium pyrenaicum, Saxifraga praetermissa, Veronica alpina…, faisant la transition avec les végétations de combes à neige alpines du Primulo integrifoliae – Salicetum reticulatae Corriol 2014 (Salicetea herbaceae).
Synchorologie :
– territoire d’observation : toute la chaîne pyrénéenne (Braun-Blanquet, 1948, sub « Salicetum retusae-reticulatae » p.p (rel. 9) ; Dupouey, 1981, sub « combes à neiges » ; Mustin, 1983 ; Mustin & Baudière, 1986, sub « groupement à saules dominants, pelouse à Elyne, landine à Dryas et hémicryptophytes » ; Carrillo & Ninot, 1992, sub « Carici parviflorae – Salicetum retusae salicetosum pyrenaicae et Elyno – Oxytropidetum dryadetosum » p.p.) ;
– sous-associations ou variantes géographiques : Oxytropis halleri ne différencie la subass. typicum que dans la partie orientale des Pyrénées (race locale de la subass.).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.422 ; EUNIS : E4.422.
Bibliographie
Braun-Blanquet J., 1948, Carrillo E. & Ninot J., 1992 ; Dupouey J.-L., 1981 ; Mustin L., 1983 ; Mustin L. & Baudière A., 1986.

FICHE N° 27-54
Association : Oxytropido foucaudii – Kobresietum myosuroidis Chouard 1943 nom. inv. et nom mut. Rivas Mart. et al. 2011 (Itinera Geobot. 18 (1) : 260).
Synonymes :
– Elyno myosuroidis – Oxytropidetum lazicae Chouard 1943 (Bull. Soc. Bot. France 90 : 3) ;
– Carici rosae – Elynetum myosuroidis Rivas Mart. 1987 (syn. inclus) (Memoria del mapa de series de vegetacion de España : 162) ;
– ? Agrostio alpinae – Caricetum rupestris Lazare 2017 (Botanique 3 : 54).
Commentaire : cette dernière association, relevée sur de petits patchs de végétation fortement dominés par Carex rupestris, à la base de l’étage alpin, est très appauvrie en taxons des Oxytropido – Elynetalia (voir col. 60e, tab. 2c) et montre une faible richesse spécifique (9-16) ; sa description dans un secteur réduit autour de Gavarnie correspond au territoire de description de l’Oxytropido foucaudii – Kobresietum par Chouard (1943). Très peu caractérisée, elle pourrait être interprétée comme une forme d’épuisement de l’Oxytropido foucaudii – Kobresietum à la frange inférieure de distribution altitudinale, dans des situations exiguës à déterminisme édaphique plus accusé (sols plus squelettiques ?). Pour comparaison, nous avons positionné son relevé type au sein de notre tableau de synthèse de l’Oxytropido foucaudii – Kobresieteum (tableau 17, col. A).
Unités supérieures : Eu-Oxytropido – Elynenion myosuroidis, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural :: rel. 19 (neotypus nominis hoc loco) du tab. 6 in Klein (1979, Phytocoenologia 5 (2) : 174, = rel. 19 du tab. 17 hoc loco).
Physionomie : pelouse écorchée, ouverte, drue, riche en Cyperaceae, souvent dominée ou co-dominée par Carex myosuroides, C. curvula subsp. rosae, parfois Dryas octopetala, à richesse spécifique élevée (principalement entre 20-40 taxons pour une médiane de 27), à optimum en début d’été, à floraison diversifiée (blanc, bleu-violet, jaune, rose), mais pouvant être assez discrète en fonction des variantes et faciès.
Combinaison caractéristique d’espèces : Silene acaulis, Carex curvula subsp. rosae, C. myosuroides, C. rupestris, Primula integrifolia, Gentiana verna, Thalictrum alpinum, Helictotrichon sedenense, Saxifraga moschata, S. oppositifolia, Festuca glacialis, Poa molinerii, Thymus nervosus, Minuartia verna, Arenaria purpurascens, A. multicaulis, Draba aizoides.
Synécologie : pelouse alpine cryophile, xérophile, des reliefs ventés, vites déneigés, à sol peu développé, sur roche mère riche en bases, généralement carbonatée, principalement entre 2 340 et 2 900 m d’altitude, pour une médiane de 2 600 m.
Variations :
– typicum, différencié positivement par Oxytropis foucaudii, Dryas octopetala, Aster alpinus, Agrostis alpina… et, d’après la colonne synthétique originale de Chouard, Artemisia eriantha (mais dans les relevés attribués, cette espèce demeure rare) ;
– antennarietosum carpaticae (Rivas Mart. 1987) Corriol subass. nov. et stat. nov. hoc. loco [basionyme : Carici rosae – Elynetum myosuroidis Rivas Mart. 1987 (Memoria del mapa de series de vegetacion de España 1 : 400.000 : 162)], différencié positivement par Antennaria carpatica (fréquence très élevée), Plantago monosperma et, en commun avec la subass. euphrasietosum minimae, Carex parviflora, Oxytropis neglecta, Agrostis rupestris, Erigeron alpinus, appauvri en espèces (généralement entre 20 et 30 pour une médiane de 23), typus nominis hoc loco : rel. non numéroté in Rivas Mart. 1987 (Memoria del mapa de series de vegetacion de España 1 : 400.000 : 162, = rel. 66 du tab. 17 hoc loco) ; déterminisme à préciser ;
– euphrasietosum minimae Corriol subass. nov. hoc. loco, différencié par Lotus corniculatus subsp. alpinus, Euphrasia minima, Festuca nigrescens, Plantago alpina, Salix herbacea, S. retusa, Botrychium lunaria, Trifolium thalii, Taraxacum pyrenaicum, Gentianella campestris, Campanula scheuchzeri, Carex ornithopoda, Hornungia alpina, plus riche en espèces (27-45), typus nominis hoc loco : rel. 11 du tab. 53 in Benito (2006, Vegetación del parque nacional de Ordesa … : 291, = rel. 279 du tab. 17 hoc loco) ; la présence des saules et de différentielles acidiphiles suggère des sols plus riches en matière organique, présentant une acidification superficielle et permettant un meilleure rétention en eau.
Synchorologie :
– territoire d’observation : Pyrénées centrales principalement, ainsi que les plus hauts massifs calcaires des Pyrénées occidentales.
Axes à développer : déterminisme des subass. typicum et antennarietosum carpaticae.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.422 ; EUNIS : E4.422.
Bibliographie
Benito J.L., 2006 ; Chouard P., 1943 ; Lazare J.-J., 2017 ; Rivas-Martínez S., 1987 ; Rivas-Martínez S. et al., 1991.
FICHE N° 27-55
Association : Oxytropido campestris – Caricetum rupestris Corriol ass. nov. hoc loco.
Synonymes : -.
Unités supérieures : Eu-Oxytropido – Elynenion myosuroidis Corriol & Sanz, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 82 (typus nominis hoc loco) du tab. 8 in Labroue (1976, Étude écologique des sols alpins du pic du Midi de Bigorre : 220, = rel. 371 du tab. 18 hoc loco), retranscrit ici
pic du Midi de Bigorre (Hautes-Pyrénées), à 2 720 m, en exposition nord-est, surface 10 m², recouvrement 80 %, 33 taxons
Carex myosuroides 3, Agrostis alpina 2, Carex curvula *rosae 2, C. rupestris 1, Arenaria multicaulis 1, Helictotrichon sedenense 1, Dryas octopetala 1, Comastoma tenellum 1, Antennaria carpatica 1, Minuartia verna 1, Oxytropis campestris (incl. *azurea) 1, Aster alpinus +, Gentiana verna +, Thymus nervosus +, Sesleria caerulea +, Anthyllis vulneraria *vulnerarioides +, Helianthemum oelandicum s. l. +, Bistorta vivipara +, Carex sempervirens +, C. ornithopoda (incl. *elongata) +, Bartsia alpina +, Saxifraga paniculata +, Galium pyrenaicum +, Potentilla crantzii +, Anemone vernalis +, Primula integrifolia +, Gentianella campestris +, Salix retusa +, Hieracium lawsonii +, Poa alpina +, Erigeron uniflorus +, Minuartia sedoides +, Bupleurum ranunculoides (incl. *gramineum) +, Draba aizoides +.
Physionomie : pelouse écorchée, ouverte, drue, riche en Cyperaceae, dominée ou co-dominée par Carex rupestris, C. curvula subsp. rosae, Helictotrichon sedenense, Oxytropis campestris, parfois Carex myosuroides ou Dryas octopetala dans la subass. typicum, à richesse spécifique moyennement élevée (principalement entre 18-35 taxons pour une médiane de 23), à optimum en début d’été, à floraison riche et diversifiée (blanc, bleu, jaune, rose, violet) ; photo 17.
Combinaison caractéristique d’espèces : Oxytropis campestris, Carex rupestris, C. curvula subsp. rosae, Draba aizoides, Helictotrichon sedenense, Gentiana verna[18], Thymus nervosus, Arenaria multicaulis, Silene acaulis, Galium pyrenaicum, Agrostis alpina, Minuartia verna, Daphne cneorum et, dans une moindre mesure, Aster alpinus, Polygala alpina, Oxytropis neglecta.
Synécologie : pelouse de l’étage alpin moyen et supérieur cryophile, xérophile, des arêtes rocailleuses ventées, vite déneigées, d’exposition variée mais fréquemment au sud, délitées par la gélifraction, à sol peu développé, sur roche mère riche en bases, mais pouvant être siliceuse mêlée d’intercalations carbonatées, principalement entre 2 440 et 2 800 m d’altitude, pour une médiane de 2 650 m.
Variations :
– typicum, différencié par Carex myosuroides, Dryas octopetala, Saxifraga oppositifolia, Sesleria caerulea, Helianthemum oelandicum s. l., Anthyllis vulneraria subsp. vulnerarioides, Bupleurum ranunculoides (subsp. gramineum) ;
– potentilletosum nivalis Corriol subass. nov. hoc. loco, différencié par Potentilla nivalis, Androsace villosa, Petrocallis pyrenaica, Erigeron uniflorus, Festuca glacialis, Veronica fruticans (subsp. cantabrica ?), Minuartia sedoides, à richesse spécifique un peu moindre (principalement entre 17-25 taxons), à l’étage alpin supérieur (altitudes principalement entre 2 650 et 2 800 m pour une médiane de 2 750 m), typus nominis nominis : rel. 23 du tab. 18 retranscrit ici
G. Corriol, le 28.VI.2008, crête de Caoubère, Bagnères-de-Bigorre (Hautes-Pyrénées, France), à 2 540 m, en exposition ouest, sur bancs gréso-pélitiques carbonatés du Dévonien inférieur, surface 10 m², pente 50°, recouvrement 50 %, hauteur végétative 1-5 cm, 31 taxons
Androsace villosa 2.2, Thymus nervosus 2.2, Helictotrichon sedenense 2.2, Festuca niphobia 2.2, Poa alpina 2.2, Carex rupestris 1.1, Potentilla nivalis 1.1, Petrocallis pyrenaica 1.2, Draba aizoides 1.1, Arenaria multicaulis 1.1, Potentilla crantzii subsp. latestipula 1.1, Silene acaulis 1.2, Gentiana verna 1.1, Saxifraga moschata subsp. moschata 1.1, Globularia repens 1.2, Androsace carnea subsp. laggeri 1.1, Oxytropis campestris +, O. neglecta +, Minuartia verna +, Sedum atratum +, Armeria alpina +, Paronychia serpyllifolia +, Saxifraga intricata +, S. paniculata +.2, Sempervivum montanum +, Bupleurum ranunculoides var. nanum, Coincya monensis subsp. cheiranthos +, Polygala alpestris +, Galium cf. pyrenaicum +, Poa cf. minor +, Juniperus communis subsp. nana +.
Synchorologie : large répartition pyrénéenne dans les hauts massifs des Pyrénées centrales et orientales.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.422 ; EUNIS : E4.422.
Bibliographie
Labroue L., 1976.

FICHE N° 27-56
Association : Oxytropido halleri – Kobresietum myosuroidis (Braun-Blanq. 1948) P. Küpfer 1974 (Boissiera 23 : 242) nom. inv. et nom. mut. Rivas Mart. et al. 2011 (Itinera Geobot. 18 (1) : 260).
Synonymes :
– Elyno myosuroidis – Oxytropidetum foucaudii Braun-Blanq. 1948 nom. illeg. (art. 34, non Chouard 1943), par ailleurs basé sur une erreur d’identification de l’Oxytropis éponyme ;
– Elyno myosuroidis – Oxytropidetum dryadetosum Braun-Blanq. 1948 (Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 164) (syn. inclus).
Unités supérieures : Eu-Oxytropido – Elynenion myosuroidis, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 10 du tab. 20 in Braun-Blanquet (1948, Mon. Est. Pir. Inst. Edaf. Ecol. Fisiol. Veg. 9 : 164).
Physionomie : pelouse écorchée, bien qu’assez recouvrante (60-95 %), drue, riche en Cyperaceae et Poaceae, jaunâtre, dominée par Carex myosuroides, plus rarement par Dryas octopetala, secondairement, par Oxytropis halleri, Helictrotrichon sedenense et Bistorta vivipara, à richesse spécifique élevée (principalement entre 26-43 taxons pour une médiane de 37), à optimum en début d’été, à floraison diversifiée (violet, blanc, bleu, jaune, rose).
Combinaison caractéristique d’espèces : Carex myosuroides, C. curvula subsp. rosae, Oxytropis halleri, Poa molinerii, Thymus nervosus, Arenaria grandiflora, Draba siliquosa, D. aizoides, Plantago monosperma, Thalictrum alpinum, Astragalus australis, Pedicularis pyrenaica, Cerastium alpinum, Minuartia sedoides, Silene acaulis, Erigeron uniflorus subsp. aragonensis, Sedum atratum, Antennaria carpatica.
Synécologie : pelouse alpine cryophile, xérophile, des reliefs ventés, vites déneigés, à sol peu développé, sur roche mère riche en bases, généralement carbonatée, principalement entre 2 300 et 2 650 m d’altitude, pour une médiane de 2 480 m.
Variations : la subass. dryadetosum indiquée par Braun-Blanquet (1948 : 175) s’insère parfaitement dans le tableau princeps de l’Oxytropido halleri – Kobresietum qui ne peut être scindé sur la base de la présence de l’espèce éponyme ; le relevé correspondant n’est qu’un faciès plus riche en Dryas octopetala.
Synchorologie :
– territoire d’observation : décrit d’un petit secteur des Pyrénées orientales restreint à quelques massifs calcaires du pourtour de la Cerdagne et du Capcir, et en Andorre (Braun-Blanquet, 1948) et non rapporté ultérieurement dans la bibliographie ; l’aire d’Oxytropis halleri s’étend jusqu’au val d’Aran à l’ouest.
Axes à développer : chorologie générale et limite occidentale d’aire de répartition à préciser.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.422 ; EUNIS : E4.422.
Bibliographie :
Braun-Blanquet J., 1948.
FICHE N° 27-57
Association : Elyno myosuroidis – Salicetum retusae Nègre 1968 (Portugaliae Acta Biol., Sér. B, 9 : 217).
Synonymes :
– Elyno – Oxytropidetum dryadetosum sensu Carrillo & Ninot (1992), Carreras et al. (1993), Soriano (1998, 2001), non Braun-Blanq. 1948 = Oxytropido halleri – Kobresietum myosuroidis (Braun-Blanq. 1948) Rivas Mart. et al. 2011 ;
– Elyno – Oxytropidetum var. à Festuca airoides sensu Vigo & Masalles (1996) ;
– Elyno – Salicetum retusae Nègre 1968 (synonyme syntaxonomique inclus).
Unités supérieures : Eu-Oxytropido – Elynenion myosuroidis, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. in Nègre (1968, Portugaliae Acta Biol., Sér. B, 9 : 217, = rel. 93 du tab. 19 hoc loco).
Physionomie : chaméphytaie relativement fermée et dense, assez verdoyante, dominée ou co-dominée par Dryas octopetala et Carex myosuroides, secondairement par Primula integrifolia et Helictotrichon sedenense, à richesse spécifique assez élevée (principalement entre 19-35 taxons pour une médiane de 28), à floraison mulitcolore (rose, blanc, bleu, jaune), commençant peu après le départ de la neige, à optimum en tout début d’été.
Combinaison caractéristique d’espèces : Bistorta vivipara, Lotus corniculatus subsp. alpinus, Helictotrichon sedenense, Dryas octopetala, Carex myosuroides, Primula integrifolia, Thymus nervosus, Agrostis alpina, Silene aucaulis, Thalictrum alpinum, Oxytropis campestris.
Synécologie : pelouse de la base de l’étage alpin et du sommet de l’étage subalpin, psychrophile à cryophile, mésoxérophile, des reliefs accusés, en conséquence assez vites déneigés, en exposition fraîche, sur sols peu épais issus de roche mère riche en bases, souvent carbonatée, principalement entre 2 090 et 2 450 m d’altitude, pour une médiane de 2 250 m.
Variations :
– typicum, plus pauvre en espèces, faiblement différencié par Antennaria carpatica, Selaginella selaginoides, Euphrasia minima, Vaccinium uliginosum subsp. microphyllum, Festuca nigrescens, Salix reticulata, Carex sempervirens, Botrychium lunaria, suggérant des sols plus riches en matière organique et à meilleure rétention d’eau que ceux de la subass. plantaginetosum monospermae ;
– plantaginetosum monospermae Corriol subass. nov. hoc. loco, différencié par Plantago monosperma, Antennaria dioica, Daphne cneorum, Festuca ovina subsp. molinieri, Helianthemum italicum subsp. alpestre, Sesleria caerulea, Carex ericetorum var. approximata, qui correspond à la situation centrale de l’association calcicole sur sols minces, typus nominis hoc loco : rel. 9 du tab. 49 in Carreras et al. (1993, Acta Bot. Barcinon. 42 : 308, = rel. 543 du tab. 19 hoc loco).
Synchorologie :
– territoire d’observation : hauts massifs de l’ensemble de la chaîne pyrénéenne.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.422 ; EUNIS : E4.422.
Bibliographie :
Benito J. 2006 ; Carrillo E. & Ninot J.M., 1992 ; Carrillo E et al., 2000 ; Carreras J. et al.,1993 ; Dupouey J.-L., 1981 ; Mustin L., 1983 ; Nègre R., 1968 ; Soriano I., 1998, 2001 ; Vigo J. & Masalles R.M., 1996.
FICHE N° 27-58
Association : Carici atratae – Kobresietum myosuroidis Braun-Blanq. 1913 (Denkschr. Schweiz. Nat. Ges. 48 : 101).
Synonymes :
– Elynetum Braun-Blanq. 1913 p.p. ;
– Elynetum Braun-Blanq. & H. Jenny 1926 ;
– Elynetum myosuroidis Braun-Blanq.(1926) 1969 ;
– Cetrario – Elynetum Gensac 1979.
L‘Elynetum Rübel 1911, basé sur un relevé très pauvre et difficilement interprétable, est considéré comme nomen ambiguum (art. 37). Il est impossible de l‘identifier précisément à la présente association, à la suivante ou encore à d’autres groupements à Carex myosuroides.
Unités supérieures: Festuco pumilae – Elynenion myosuroidis Corriol, Sanz & Van Es, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 6 (lectotypus nominis hoc loco) du tab. in Braun-Blanquet (1913, Denkschr. Schweiz. Nat. Ges. 48 : 101).
Physionomie : pelouse plus ou moins fermée dominée par Carex myosuroides ; photo 18.
Combinaison caractéristique d’espèces : Carex myosuroides, C. atrata, C. parviflora, Euphrasia minima, Antennaria carpatica subsp. helvetica, Agrostis alpina, Pachypleurum mutellinoides, Helictochloa versicolor subsp. versicolor, Minuartia sedoides, Phyteuma globularifolium subsp. pedemontanum, Androsace obtusifolia, Salix serpyllifolia, Oxytropis campestris, Leontopodium nivale subsp. alpinum, Silene acaulis, Festuca pumila, Erigeron uniflorus.
Synécologie : pelouse primaire de l’étage alpin, cryophile, liée aux crêtes et hauts de versant exposés au vent et au gel une grande partie de l’année, installée sur des substrats carbonatés humifères.
Variations : -.
Synchorologie :
– territoire d’observation : toutes les Alpes.
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Bensettiti F. et al., 2002 ; Braun-Blanquet J., 1913 ; Baun-Blanquet J., & Jenny H., 1926 ; Delpech R., 2003 ; Gensac P., 1979 ; Guinochet M., 1938 ; Mikolajczak A. et al., 2014 ; Quantin A. & Nétien G., 1940 ; Rivas-Martínez S. & Géhu J.-M., 1978 ; Rübel E., 1911 ; Van Es J. et al., 2014 ; Villaret J.-C., 2007.

FICHE N° 27-59
Association : Kobresio myosuroidis – Caricetum rosae Erschbamer 1992 (Phytocoenologia 21 : 99).
Synonymes :
– Elynetum Friedel (1956) ;
– groupement à Sesleria varia et Carex rosae (in Gensac, 1977).
Unités supérieures : Festuco pumilae – Elynenion myosuroidis Corriol, Sanz & Van Es, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 86 du tab 1 in Erschbamer (1992, Phytocoenologia 21 : 96) désigné par l‘auteur.
Physionomie : pelouse plus ou moins fermée dominée par Carex curvula subsp. rosae et C. myosuroides ; photo 19.
Combinaison caractéristique d’espèces : Carex curvula subsp. rosae, C. myosuroides, C. parviflora, Oxytropis campestris, O. jacquini, O. helvetica, Pachypleurum mutellinoides, Gentiana orbicularis, G. brachyphylla, Saussurea alpina, Sesleria caerulea, Helianthemum oelandicum, H. nummularium, Thymus serpyllum agg., Antennaria carpatica subsp. helvetica, Bartsia alpina, Astragalus australis, Minuartia verna, Sedum atratum, Festuca pumila, Dryas octopetala, Potentilla crantzii, Leontopodium nivale subsp. alpinum.
Synécologie : pelouse primaire de l’étage alpin, mésocryophile à cryophile, des hauts de versant plus ou moins ensoleillés mais exposés au vent et au gel, roches tendres carbonatées ou schistes lustrés, sols assez peu humifères à pH toujours proche de la neutralité, pouvant être régulièrement perturbés par l’érosion.
Variations : -.
Synchorologie :
– territoire d’observation : Alpes internes.
Axes à développer : -.
Correspondances :
HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie : Erschbamer B., 1992 ; Friedel J., 1956 ; Gensac P., 1979 ; Mikolajczak A. et al., 2014.

FICHE N° 27-60
Association : Festucetum pumilae Gams 1927 (Beitr. Geobot. Landesaufn. 15 : 619).
Synonymes :
– Festucetum pumilae Braun-Blanq. 1913 ;
– Seslerio – Caricetum festucetosum pumilae Braun-Blanq. & H. Jenny 1926.
Unités supérieures: Festuco pumilae – Elynenion myosuroidis Corriol, Sanz & Van Es, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 2 (lectotypus nominis hoc loco) du tab. in Gams (1926 : 617).
Physionomie : pelouse plus ou moins ouverte à Festuca pumila.
Combinaison caractéristique d’espèces : Festuca pumila, Gentiana clusii, Astragalus alpinus, Carex rupestris, C. myosuroides, C. capillaris, C. ornithopoda subsp. ornithopodioides, Dryas octopetala, Arenaria multicaulis, Silene acaulis, Saxifraga oppositifolia, Polygala alpina, Gentiana nivalis, Veronica aphylla, Oxytropis campestris, Pedicularis verticillata, Agrostis alpina, Minuartia verna, Aster alpinus, Galium anisophyllon, Helianthemum oelandicum, Salix retusa, (Festuca alpina).
Synécologie : pelouse plus ou moins ouverte pionnière, établie sur des croupes, buttes rocailleuses, corniches ou vires de parois aux sols squelettiques, bien exposés au vent et au gel, occupant parfois de petites surfaces, préférentiellement sur calcaires durs aux étages subalpin supérieur et alpin.
Variations : -.
Synchorologie :
– territoire d’observation : massifs calcaires des Alpes internes et intermédiaires (Mikolajczak et al., 2014), Préalpes du Nord (Gams, 1927) ; forme appauvrie et plus thermophile sur les crêtes du Jura (Béguin, 1972).
Axes à développer : confirmer le rattachement phytosociologique pour ce syntaxon intermédiaire entre les deux sous-alliances.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Béguin C., 1972 ; Braun-Blanquet J., 1913 ; Gams H., 1927 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019 ; Villaret J.-C., 2007.
FICHE N° 27-61
Association : Caricetum firmae Braun-Blanq. in Braun-Blanq. & H. Jenny 1926 (Denksch. Schweiz. Naturf. Ges. 63 (2) : 227).
Synonyme : Elynetum sensu Barbero 1972 (synonyme syntaxonomique inclus provisoirement).
Unités supérieures : Caricenion firmae (Gams 1936) Theurillat in Theurillat, Aeschimann, P. Küpfer & Spichiger 1995, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : rel. 11 (lectotypus nominis hoc loco) du tab. 7 in Braun-Blanquet & Jenny (1926 : 229).
Physionomie : pelouse plus ou moins ouverte à Carex firma ; photo 20.
Combinaison caractéristique d’espèces : Carex firma, C. ornithopoda subsp. ornithopodioides, C. rupestris, Carex mucronata, Saxifraga caesia, Bistorta vivipara, Dryas octopetala, Sesleria caerulea, Soldanella alpina, Chamorchis alpina, Sedum atratum, Festuca pumila, Sesleria caerulea, Salix reticulata, S. serpyllifolia, Crepis jacquini, Gentiana tergoulensis.
Note : Crepis jacquini et Gentiana tergoulensis sont absentes de France.
Synécologie : pelouse pionnière plus ou moins ouverte établie sur des vires et des versants peu pentus sur calcaires durs ou éboulis stabilisés dans des situations fortement soumises au froid et au gel, fréquemment déneigées l’hiver, sols généralement peu humifères (absence des taxons acidiphiles cryophiles), à l’étage alpin.
Variations : -.
Synchorologie :
– territoire d’observation : surtout Alpes centrales ; syntaxon en limite d’aire occidentale (donc appauvri) dans les Préalpes des Bornes (Charpin, 1968), le Haut-Giffre, le Briançonnais (Mikolajczak et al., 2014), fragmentaire en Haute-Roya (Barbero, 1972), où Carex firma est historiquement signalé mais non confirmé. Présence à étudier en Vanoise.
Axes à développer : étudier plus précisemment l’Elynetum sensu Barbero 1972 qui, marqué par la présence de Carex mucronata, pourrait être comparé à d’autres groupements contenant cette espèce dans les Alpes françaises (Vercors, Mont-Cenis, Haute-Ubaye, Haute-Roya). Étudier les groupements de Vanoise, très proches par leur situation écologique et une bonne partie de leur cortège floristique (abence de C. firma).
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie
Barbero M., 1972 ; Braun-Blanquet J. & Jenny H., 1926 ; Charpin A., 1968 ; Mikolajczak A. et al., 2014 ; Prunier P. et al., 2019.

FICHE N° 27-62
Association : Dryadetum octopetalae Rübel 1911 (Pflanzengeschichte und Pflanzengeographie 47: 135) nom. inval. (art. 7).
Synonymes : -.
Unités supérieures : Caricenion firmae (Gams 1936) Theurillat in Theurillat, Aeschimann, P. Küpfer & Spichiger 1995, Oxytropido – Elynion myosuroidis Braun-Blanq. 1950, Oxytropido – Elynetalia myosuroidis Oberd. ex Albrecht 1969.
Type nomenclatural : l‘unique relevé sans coefficient donné par Rübel (1911 : 135) ne peut être retenu pour typification.
Physionomie : landine ouverte pionnière à Dryas octopetala. Cette espèce pouvant constituer des landines dans d’autres situations écologiques, il conviendra de vérifier la combinaison caractéristique de taxons ; photo 21.
Combinaison caractéristique d’espèces : Campanula cochlearifolia, Bistorta vivipara, Salix retusa, S. serpyllifolia, Festuca pumila, Sedum atratum, Carex myosuroides, Gentiana verna, Aster alpinus, Helianthemum oelandicum, Saxifraga paniculata, S. oppositifolia, S. aizoides.
Synécologie : landine pionnière ouverte assez pauvre en espèces, développée sur des éboulis stabilisés et moraines en situation plus longuement enneigée que l’association précédente, mésocryophile à chionoclinophile, dans les expositions froides préférentiellement. Association typiquement de transition entre le Caricenion firmae et l’Arabidion caeruleae.
Variations : -.
Synchorologie :
– territoire d’observation : massifs calcaires des Alpes internes (Prunier et al., 2019 ; obs récentes CBNA en Vanoise et Queyras).
Axes à développer : -.
Correspondances : HIC : 6170 ; CORINE biotopes : 36.41 ; EUNIS : E4.4.
Bibliographie : Rübel E., 1911 ; Prunier P. et al., 2019.

[1] Archiloque et al. (1971 : 220) font référence à une proposition antérieure de 1966, référence non citée.
[2] Caractéristique dans les Pyrénées, plutôt différentiel dans les Alpes.
[3] Dans la mesure où il s’agit bien de ce taxon, étant donné que sa présence dans les Pyrénées reste discutée (voir Laínz & Sáez, 1998).
[4] La subass. horminetosum est tellement proche du typicum que leurs relevés types respectifs sortent sur la même ramification terminale de notre CAH.
[5] C’est généralement T. pyrenaicum (ou l’équivalent T. dissectum ss. auct.) qui est cité dans la bibliographie. L’écologie de T. panalpinum nous semble toutefois plus compatible. À étudier plus précisément.
[6] L’alchémille désignée par Lacoste (1965) sous le nom de A. conjuncta correspond en réalité plus probablement à A. alpigena.
[7] Oxytropis helvetica Scheele était originellement cité mais très probablement par confusion avec O. neglecta Ten., le seul des deux taxons présent à l’est de la Tinée.
[8] Inclus subsp. alpicola (Vayr.) O. Bolòs & Vigo.
[9] Ou Oxytropis neglecta, voir note sous l’Oxytropido amethysteae – Caricetum humilis.
[10] Cité sous Alyssum diffusum mais, d’après Zozomová-Lihová et al. (2014), seuls A. montanum sensu stricto et A. cacuminum Španiel et al. sont présents dans la dition. Peut-être est-ce de ce dernier plus orophile dont il s’agit ici ?
[11] Selon Vigo & Massales (1996 : 300), il s’agirait de Festuca yvesii.
[12] L’ordre des taxons éponymes retenu ici répond à l’exigence de l’article 10b du code international de nomenclature phytosociologique.
[13] Identification à vérifier, car écologie inhabituelle pour Festuca pumila.
[14] Chouard (1943) cite Alchemilla alpina dans la colonne princeps, mais nous pensons qu’il s’agit en réalité de A. alpigena comme l’indique l’ensemble des relevés que nous avons réunis. A. alpina est une espèce silicicole acidiphile.
[15] La description orginale donne « Festuca gr. rubra », mais les relevés attribués contiennent F. nigrescens, ce qui est vraisemblable.
[16] La description originale donne « Taraxacum alpinum », taxon non signalé dans les Pyrénées par Tison & de Foucault (2014).
[17] C’est Taraxacum pyrenaicum qui est cité dans les relevés.
[18] On trouve dans ces pelouses des écomorphoses de Gentiana verna qui ont parfois été confondues avec G. orbicularis ou G. brachyphylla.
Bibliographie
Allier C., 1971. La végétation des terrasses quaternaires du bassin moyen du Buech. Annales littéraires de l’Université de Besançon 21 : 81-108.
Allier C. & Ritter J., 1971. L’Androsaco–Gentianetum du Vercors méridional et les groupements vicariants des Baronnies. Annales littéraires de l’Université de Besançon 125, Actes du colloque sur la flore et la végétation des chaînes alpine et jurassienne : 271-292.
Archiloque A., Borel L. & Lavagne A., 1970. Feuille de la Javie (XXXIV-40) au 1/50 000. Documents pour la carte de la végétation des Alpes, 37 p.
Archiloque A., Borel L. & Lavagne A., 1971. La notion d’étage pseudo-alpin dans les Préalpes françaises méridionales. Colloque interdisciplinaire sur les milieux naturels supra-forestiers des montagnes du bassin occidental de la Méditerranée : 201-232.
Archiloque A., Borel L. & Devaux J.-P., 1974. Carte des séries de végétation, feuille d’Entrevaux (XXXV-41) au 1/50000. Bulletin pour la carte de la végétation de Provence et des Alpes du Sud 1 : 1-129.
Aubert S. & Luquet A., 1930. Études phytogéographiques sur la chaîne jurassienne. Recherches sur les associations végétales du Mont-Tendre. Revue de géographie alpine 18 (3) : 491-536.
Bannes-Puygiron G. (de), 1933. Le Valentinois méridional, esquisse phytosociologique. Recueil des travaux de l’Institut botanique et Annales de l’Université de Montpellier et du Languedoc méditerranéen-Roussillon 19 : 1-200 (Communications de la SIGMA 19).
Barbero M., 1968. À propos des pelouses écorchées des Alpes maritimes et ligures. Bulletin de la Société botanique de France 115 : 219-244.
Barbero M., 1972. Études phytosociologiques et écologiques comparées des végétations orophiles alpine, subalpine et mésogéenne des Alpes maritimes et ligures. Thèse de doctorat d’État, Université de Provence (Marseille), 418 p., pl. + fig.
Barbero M. & Bonin G., 1969. Signification biogéographique et phytosociologique des pelouses écorchées des massifs méditerranéens nord-occidentaux, des Apennins et des Balkans septentrionaux (Festuco-Seslerietea). Bulletin de la Société botanique de France 116 : 227-243.
Barbero M., Bonin G. & Quézel P., 1971. Signification bioclimatique des pelouses écorchées surt les montagnes du pourtour méditerranéen, leurs relations avec les forêts d’altitude. Bulletin de la Société botanique de France 118 : 17-56.
Barbero M., Bonin G. & Quézel P., 1975. Les pelouses écorchées des montagnes circum-méditerranéennes. Phytocoenologia 1 (4) : 427-459.
Barbero M. & Quézel P., 1975. Végétation culminale du mont Ventoux. Sa signification dans une interprétation phytogéographique des Préalpes méridionales. Ecologia Mediterranea 1 : 3-33.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Patrimoines naturels (publications scientifiques du Muséum, Paris) 61 : 1-171.
Baudière A. & Küpfer Ph., 1968. Sur les peuplements d’astragales épineux de la partie orientale de la chaîne pyrénéenne. Bulletin de la Société des sciences naturelles de Neuchâtel 91 : 75-85.
Béguin C., 1972. Contribution à l’étude phytosociologique et écologique du haut-Jura. Matériaux pour le levé géobotanique de la Suisse 54 : 1-190.
Benito Alonso J.L., 2006. Vegetación del parque nacional de Ordesa y Monte Perdido (Sobrarbe, Pirineo central aragonés). Publicaciones del Consejo de Protección de la Naturaleza de Aragón, 421 p. + carte.
Berset J., 1969. Pâturages, prairies et marais montagnards et subalpins des Préalpes fribourgeoises. Bulletin de la Société fribourgeoise des sciences naturelles 58 : 1-55.
Biondi E., Blasi C., Allegrezza M., Anzellotti I., Azzella M. M., Carli E., Casavecchia S., Copiz R., Del Vico E., Facioni L., Galdenzi D., Gasparri R., Lasen C., Pesaresi S., Poldini L., Sburlino G., Taffetani F., Vagge I., Zitti S. & Zivkovic L., 2014. Plant communities of Italy: The Vegetation Prodrome. Plant Biosystems 148 (4) : 728-814.
Bioret F. & Royer J.-M., 2009. Présentation du projet de déclinaison du Prodrome des végétations de France. Journal de botanique de la Société botanique de France 48 : 47-48.
Bioret F. & Gaudillat V. & Royer J.-M., 2013. The Prodrome of French vegetation: a national synsystem for phytosociological knowledge and management issues. Plant Sociology 50 : 17-21.
Boissier J.-M., 2005. Guide d’interprétation des habitats naturels du massif des Bauges. Conservatoire botanique national alpin, 148 p.
Bonin G., 1969. À propos de la valeur phytosociologique des pelouses écorchées de l’Apennin. Annales de la Faculté des sciences de Marseille 42 : 139-144.
Braun-Blanquet J., 1913. Die Vegetationsverhältnisse der Schneestufe in den Rätisch-Lepontinischen Alpen N. Denkschriften der schweizerischen Naturforschenden Gesellschaft 48 : 1-347.
Braun-Blanquet J., 1931. Recherches phytogéographiques sur le massif du Gross Glockner (Hohe Tauern). Communication de la Station internationale géobotanique méditeranéenne et alpine 13 : 5-65.
Braun-Blanquet J., 1948. La végétation alpine des Pyrénées-Orientales. Monografía de la Estación de Estudios Pirenaicos y del Instituto de Edafología, Ecología e Fisiología Vegetal 9 : 1-306.
Braun-Blanquet J., 1950. Übersicht der Pflanzengesellschaften Rätiens (IV). Vegetatio 2 (1) : 20-37.
Braun-Blanquet J., 1954. La végétation alpine et nivale des Alpes françaises. Communications de la SIGMA 125 : 26-96.
Braun-Blanquet J. & Jenny H., 1926. Vegetations-Entwicklung und Bodenbildung in der alpinen Stufe ders Zentralalpen (Klimaxgebiet des Caricion curvulae). Denkschriften der schweizerischen Naturforschenden Gesellschaft 63 (2) : 184-349.
Carreras J., Carrillo E. & Ninot J. M., 1996. Las comunidades con Salix pyrenaica en el Valle de Arán. Anales del Jardin Botánico de Madrid 54 : 504-511.
Carreras J., Carrillo E., Masalles R.M., Ninot J.M. & Vigo J., 1993. El poblament vegetal de les valls de Barravés i de Castanesa. Acta Botanica Barcinonensia 42 : 3-392.
Carrillo E. & Ninot J.M., 1990. Noves comunitats pratenses deis Pirineus centrals. Folia Botanica Miscellanea 7 : 99-114.
Carrillo E. & Ninot J.M., 1992. Flora I vegetació de les Valls d’Espot I de Boí, II. Ed. Institut d’Estudis Catalans, Barcelona, 351 p.
Carrillo E. & Vigo J., 1997. Mapa de vegetació de Catalunya 1 : 50.000. Memòria del full 254 (35-11) Gósol. Institut Cartogràfic de Catalunya i Institut d’Estudis Catalans, Barcelona.
Carrillo E., Carreras J., Font X., Ninot J.M., Soriano I. & Vigo J., 2000. La vegetació de les serres prepirininenques compreses entre els rius Segre i Llobregat, 4 – Pastures alpines i subalpines. Bulletin de la Société linnéenne de Provence 51 : 95-120.
Chaix G., 1954. Étude phytogéographique des vallées supérieures de la Romanche et de la Guisane aux abords du col du Lautaret. D.E.S. de l’Université de Marseille.
Charpin A., 1968. Le Carex firma Host et le Caricetum firmae Br.-Bl. dans les Préalpes des Bornes (Haute-Savoie). Candollea 23 (1) : 121-130.
Chouard P., 1943. Le peuplement végétal des Pyrénées centrales, 1 – Les montagnes calcaires de la vallée de Gavarnie (suite 1). Bulletin de la Société botanique de France 90 : 1-4.
Chouard P., 1943. Le peuplement végétal des Pyrénées centrales, 1 – Les montagnes calcaires de la vallée de Gavarnie (suite 2). Bulletin de la Société botanique de France 90 : 25-29.
Corriol G. & Mikolajczak A., 2014. Contribution au Prodrome des végétations de France : les Salicetea herbaceae Braun-Blanq. 1948. Journal de botanique de la Société botanique de France 68 : 15-49.
Corriol G. & Mikolajczak A., 2017. Contribution au Prodrome des végétations de France : les Caricetea curvulae Braun-Blanq. 1948 nom. conserv. propos. Journal de botanique de la Société botanique de France 77 : 57-876.
Dalmas J.-P., 1972. Études phytosociologique et écologique de l’étage alpin des Alpes sud-occidentales françaises et plus particulièrement de la région Vars-Escreins (H.A). Thèse de doctorat d’État, Université de Provence, 173 p.
Daniëls F.J.A., 1982. Vegetation of the Angmagssalik District, Southeast Greenland, IV – Shrub, dwarf shrub and terricolous lichens. Meddelelser om Grønland, Bioscience 10 : 5-81.
Delarze, R., Gonseth Y & Galland P. ,1998. Guide des milieux naturels de Suisse. Delachaux & Niestlé, Lausanne, 415 p.
Delpech R., 2003. Typologie floristico-écologique des peuplements de prairies et pelouses du Parc national de la Vanoise (Savoie). Journal de botanique de la Société botanique de France 22 : 1-88.
Dupouey J.L., 1981. Contribution à l’étude phytosociologique du massif des Eaux-Chaudes (Pyrénées-Atlantiques). DEA d’Écologie végétale, Université Paris-Sud centre d’Orsay & Université Bordeaux I, Centre d’écologie montagnarde de Gabas, 60 p.
Dupouey J.L., 1986. Essai de synthèse sur les groupements végétaux des pelouses calcicoles pyrénéennes. Actes du colloque international de botanique pyrénéenne, La Cabanasse (Pyrénées-Orientales) : 399-412.
Erschbamer B., 1992. Zwei neue Gesellschaften mit Krummseggen (Carex curvula ssp. rosae, Carex curvula ssp. curvula) aus den Alpen, ein Beitrag zur Klärung eines alten ökologischen Rätsels. Phytocoenologia 21 : 91-116.
Farràs A., Masalles R.M., Velasco E. & Vigo J., 1981. Sobre la flora i la vegetació de la Serra de Cadí. Butlletí de la Institució Catalana d’Història Natural 46 : 131-145.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques. Thèse de doctorat d’État, Université de Rouen, 675 p.
Foucault B. (de), 2012. Contribution au Prodrome des végétations de France : les Nardetea strictae Rivas Goday in Rivas Goday & Rivas Mart. 1963. Journal de botanique de la Société botanique de France 59 : 241-344.
Foucaut L., & Bethouse J.-E, 2006. À propos d’un nouvel habitat trouvé dans les Préalpes de Castellane : les couloirs herbeux à Carex autro-alpina Becherer. Courrier scientifique du Parc naturel régional du Verdon 1 : 22-48.
Friedel J., 1956. Die alpine Vegetation des obersten Mölltales (Hohe Tauern). Erläuterung zur Vegetationskarte der Umgebung der Pasterze (Grossglockner). Wissenschaftliche Alpenvereinshefte 16 : 1-153.
Fromard F., 1982. À propos de quelques stations nouvelles ou méconnues de l’Arctostaphylos alpina (L.) Sprengel dans les Pyrénées. Bulletin de la Société botanique de France, Lettres botaniques 129 (2) : 167-175.
Gams H., 1927. Von den Follatères zur Dent de Morcles. Vegetationsmonographie aus dem Wallis. Beiträge zur Geobotanischen Landesaufnahme 15 : 1-760.
Gams H., 1936. Beiträge zur pflanzengeographischen Karte Österreichs, I – Die Vegetation des Grossglocknergebietes. Abhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 16 (2) : 1-79.
Gaultier C., 1989. Relations entre pelouses eurosibériennes (Festuco-Brometea Br.-Bl. et Tx. 43) et groupements méditerranéens (Ononido-Rosmarineta Br.-Bl. 47). Étude régionale (Diois) et synthèse sur le pourtour méditerranéen nord-occidental, II. Thèse de doctorat d’État, Université de Paris-sud, Orsay, I : 230 p. et II : 119 p. + 54 tab. et fig.
Gensac P., 1979. Les pelouses supraforestières du massif de la Vanoise. Travaux scientifiques du Parc national de la Vanoise 9 : 11-243.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Gontard P., 1955. Contribution à l’étude géobotanique du mont Ventoux. Thèse de doctorat d’État, Université de Montpellier, Faculté des sciences, 4 vol.
Gruber M., 1978. La végétation des Pyrénées ariègeoises et catalanes occidentales. Thèse de doctorat d’État, Université d’Aix-Marseille, 305 p. + 1 vol. fig. & tabl.
Gruber M., 1979a. Les pelouses calcicoles orophiles et nordiques des étages subalpin de type médio-européen et alpin en Ariège et en Pyrénées catalanes occidentales. Ecologia Mediterranea 4 : 75-94.
Gruber M., 1979b. Les pelouses écorchées calcicoles de l’étage subalpin en Ariège et en Pyrénées catalanes occidentales. Bulletin de la Société botanique de France 126 : 241-255.
Guinochet M., 1938. Étude sur la végétation de l’étage alpin dans le bassin supérieur de la Tinée (Alpes-Maritimes). Communication de la Station internationale géobotanique méditeranéenne et alpine, Montpellier, 59 : 1-458.
Guinochet M., 1975. Sur quelques associations de pelouses des Préalpes de Grasse. Anales del Instituto Botánico A.J. Cavanilles 32 (2) : 1291-1314.
Herrera Gallastegui M., Loidi, J. & Fernández Prieto J.A., 1991. Vegetación de las montañas calizas vasco cantábricas: Comunidades culminícolas. Lazaroa 12 : 345–353.
Illa E., Carrillo E. & Ninot J.M., 2006. Patterns of plant traits in Pyrenean apine vegetation. Flora 201 : 528-546.
Jiménez-Alfaro B., Alonso Felpete J.I., Bueno Sánchez A. & Fernández Prieto J.A., 2014. Alpine plant communities in the Picos de Europa calcareous massif (Northern Spain). Lazaroa 35 : 67-105.
Julve Ph., 1988. Réflexions sur la structure et la dynamique des lisières forestières. Conséquences sur le synsystème. Colloques phytosociologiques XIV : 55-79.
Julve Ph., 2017 ff. (1998 ff). baseveg. World vegetation database [09 février 2017]. Programme Catminat. http//www.tela-botanica.org.
Klein J.-C., 1979. Application de l’analyse factorielle des correspondances à l’étude phytosociologique de l’étage alpin des Pyrénées centrales. Phytocoenologia 5 (2) : 125-188.
Koroleva N.E., 2015. Survey of Dryas octopetala – dominated plant communities in the European and North-West-Siberian Arctic. Botanica Pacifica 4 (1) : 1-20.
Küpfer Ph., 1974. Recherches sur les liens de parenté entre la flore orophile des Alpes et celle des Pyrénées. Boissiera 23 : 1-322 + 10 pl.
Labroue L., 1976. Étude écologique des sols alpins du pic du Midi de Bigorre. Thèse de doctorat d’État, Université Paul-Sabatier, Toulouse, 230 p.
Lacoste A., 1964. Premières observations sur les associations subalpines des Alpes maritimes : étude phytosociologique des pelouses sèches basophiles. Bulletin de la Société botanique de France 111 : 61-69.
Lacoste A., 1965. Étude phytosociologiques des forêts de mélèzes dans les Alpes maritimes : leurs relations avec les pelouses mésophiles subalpines et les rhodoraies. Revue générale de botanique 72 : 603-614.
Laínz M. & Sáez L., 1998. Más aún acerca de Oxytropis DC. (Leguminosae). Anales del Jardin Botánico de Madrid 56 (2) : 407-408.
Lancioni A., Facchi J. & Taffetani F., 2011. Syntaxonomical analysis of Kobresio myosuroidis-Seslerietea caerulea and Carici rupestris-Kobresietea bellardii classes in central southern Apennines. Fitosociologia 48 (1) : 3-21.
Lazare J.-J., 2017. Syntaxons pyrénéens nouveaux des Littorelletea Br.-Bl. & Tüxen ex Westhoff, Dijk & Passchier 1946, Thlaspietea rotundifolii Br.-Bl. 1948, Carici rupestris-Kobresietea bellardii Ohba 1974, Elyno myosuroidis-Seslerietea caeruleae Br.-Bl. 1948, Mulgedio alpine-Aconitetea variegatae Hadač & Klika in Klika & Hadač 1944 et Loiseleurio procumbentis-Vaccinietea microphylli Eggler ex Schubert 1960. Botanique 3 : 39-69.
Lazare J.-J. & Mauric A., 1986. L’Helictotricho-Bellardiochloetum violaceae et le Primulo-Horminetum pyrenaici, associations orophiles nouvelles des Pyrénées occidentales. Actes du colloque international de botanique pyrénéenne, la Cabanasse : 413-420.
Lejoly J., 1975. Phytosociologie écologique en moyenne montagne méditerranéenne. Groupes écologiques, associations stationnelles et séries de végétation dans une séquence bioclimatique méditerranéo-alpine de la région d’Entrevaux-Peyresq (Alpes de Haute-Provence, France). Thèse de doctorat d’État, Université libre de Bruxelles, 2 tomes, 594 p.
Lippert W., 1966. Die Pflanzengesellschaften des NSG Berchtesgaden. Berichte der Bayerischen Botanischen Gesellschaft zur Erforschung der heimischen Flora 39 : 67-122.
Lippmaa T., 1933. Aperçu général sur la végétation autochtone du Lautaret (Hautes Alpes). Acta Instituti et Horti Botanici Tartuensis (Dorpatensis) 3 : 1-104.
Loidi J., 1982. Datos sobre la vegetación de Guipúzcoa (País Vasco). Lazaroa 4 : 63-90.
Loidi J. & Biurrun I., 1998. Notas nomenclaturales sobre la vegetación del norte de la Península Ibérica, VII. Lazaroa 19 : 161-165.
Lüdi W., 1921. Die Pflanzengesellschaften des Lauterbrunnentales und ihre Sukzession. Beiträge zur geobotanischen Landesaufnahme der Schweiz 9, 364 p.
Mikolajczak A., 2018. Cartographie des habitats naturels de la montagne du Saut. Conservatoire botanique national alpin, 85 p.
Mikolajczak A., Abdulhak D., Garraud L., Legland T., Pache G., Van Es J. & Villaret J.-C., 2014. Fiches descriptives des habitats naturels et semi-naturels du territoire d’agrément du CBNA. Conservatoire botanique national alpin, 580 p.
Misset C., 2017. Observations sur les Nardetea strictae Rivas Goday in Rivas Goday & Rivas-Mart. 1963 et des pelouses calcicoles acidoclines dans la Drôme et l’Isère. Psychodrômia, Bulletin de la Société botanique de la Drôme 2 : 48-93.
Montserrat P. & Villar L., 1975. Les communautés à Festuca scoparia dans la moitié occidentale des Pyrénées (notes préliminaires). Documents phytosociologiques 9-14 : 207-218.
Montserrat P. & Villar L., 1987. Las comunidades de Saponaria caespitosa en el Pirineo. Lazaroa 7 : 9-24.
Moor M., 1957. Die kartographische Darstellung der Vegetation des Creux du Van-Gebietes (Jura des Kantons Neuenburg). Matériaux pour le levé géobotanique de la Suisse 37, 114 p.+annexes
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Y.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M. & Tichý L., 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 19 (1) : 3-264.
Mustin L., 1983. Contribution à l’étude de la végétation des milieux supraforestiers pyrénéens : le vallon de Laurenti (Ariège). Thèse de doctorat d’État, Université Paul-Sabatier, Toulouse, 271 p.
Mustin L. & Baudière A., 1986. Les groupements à Dryas octopetala L. du vallon de Laurenti (Ariège), analyse structurale et approche édaphique. Veröffentlichungen des Geobotanischen Institutes der Eidg. Technischen Hochschule, Stiftung Rübel in Zürich 87 : 291-308.
Nègre R., 1968. La végétation du bassin de l’One (Pyrénées centrales). Portugaliae Acta Biologica, Série B, 9 : 196-290.
Nègre R., 1970. La végétation du bassin de l’One (Pyrénées centrales), trosième note : les landes. Portugaliae Acta Biologica, Série B, 11 (1-2) : 51-167.
Ninot J.M., 1988. Sobre la vegetació pratense supraforestal del massís del Turbó (Prepirineu aragonés). Monografias del Instituto de Estudios Pirenaicos 4 : 677-685.
Ninot J.M., 1996. Estudio fitocenológico del macizo del Turbón (Prepirineo central), II – Pastos. Lucas Mallada 8 : 121-182.
Noble V., 2008. Typologie des groupements végétaux des Alpes d’Azur (Préalpes de Grasse – Alpes-Maritimes). Projet pour un futur parc naturel régional. Conservatoire botanique national méditerranéen de Porquerolles, 85 p.
Oberdorfer, E., 1957. Süddeutsche Pflanzengesellschaften. Pflanzensoziologie 10 :1-567.
Ohba T., 1974. Vergleichende Studien über die alpine Vegetations Japans, 1 – Carici rupestris-Kobresietea bellardii. Phytocoenologia 1 : 339-401.
Peyre G. & Font X., 2011. Syntaxonomic revision and floristic characterization of the phytosociological alliances corresponding to subalpine and alpine grasslands of the Pyrenees and Cantabrian Mountains (classes Caricetea curvulae, Carici-Kobresietea, and Festuco-Seslerietea). Plant Biosystems 145, suppl.1 : 220-232.
Poirion L., 1960. Avena sempervirens Vill. dans les Préalpes de la Côte d’Azur. Bulletin de la Société botanique de France 107 (4-6) : 200-202.
Prunier P., Greulich F., Béguin C., Boissezon A., Delarze R., Hegg O., Klötlzi F., Pantke R., Steffen J., Steiger P. & Vittoz P., 2019. Phytosuisse : un référentiel pour les associations végétales de Suisse, v. 4, http://www.infoflora.ch/fr/milieux/phytosuisse/ ISSN 2624-6171.
Quantin A. & Nétien G., 1940. Les associations végétales de l’étage alpin des Alpes de l’Oisans. Bulletin de la Société botanique de France 87 (1-3) : 27-47.
Redžić S., 2003. The syntaxonomy and syngenesis of the Elyno-Seslerietea Br.-Bl. 1948 in the Balkan peninsula. Annali di Botanica (nuova serie) 3 : 53-74.
Richard J.-L, 1977. La végétation du Vanil Noir et du vallon des Mortheys. Bulletin de la Société fribourgeoise des sciences naturelles 66 : 1−52.
Richard J.-L, 1984. Quelques associations végétales xérophiles du Val de Binn (Haut-Valais, Suisse). Botanica Helvetica 94 : 161-176.
Richard J.-L., 1985. Pelouses xérophiles alpines des environs de Zermatt (Valais suisse). Botanica Helvetica 92 (2) : 193-211.
Ritter J., 1972. Les groupements végétaux des étages subalpin et alpin du Vercors méridional, essai d’interprétation statistique. Vegetatio 24 : 313-403, 25 : 357-365.
Rivas-Martínez S. (coord.), 1987. Memoria del mapa de series de vegetacion de España 1 : 400.000. ICONA, Serie Técnica, Madrid, 268 p. + 30 cartes.
Rivas-Martínez S. & Géhu J.-M., 1978. Observations syntaxonomiques sur quelques végétations du Valais Suisse. Documents phytosociologiques, NS, III : 371-423.
Rivas-Martínez S., Díaz T.E., Prieto J.A., Loidi J. & Penas A., 1984. Los Picos de Europa: la vegetación de la alta montaña cantábrica. Ed. Leonesas, León, 295 p.
Rivas-Martínez S., Báscones J.C., Díaz T.E., Fernández-González F. & Loidi J., 1991a. Vegetación del Pireneo occidental y Navarra. Itinera Geobotanica 5 : 5-456.
Rivas-Martínez S., Díaz T.E., Fernández Prieto J.A., Loidi J. & Penas A., 1991b. Festuco hystricis-Ononidetea striatae y Rosmarinetea officinalis clases de vegetación independientes. Itinera Geobotanica 5 : 505-516.
Rivas-Martínez S. & Costa M., 1998. Datos sobre la vegetación y el bioclima del valle de Arán. Acta Botanica Barcinonensia 45 : 473-499.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Loidi J., Lousã M. & Penas A., 2001. Syntaxonomical checklist of vascular plant communituies of Spain and Portugal to association level. Itinera Geobotanica 14 : 5-341.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal, addenda to the syntaxonomical checklist of 2001. Itinera Geobotanica 15 : 5-922.
Rivas-Martínez S. & coautores, 2011. Mapa de series, geoseries y geopermaseries de vegetación de España, II. Itinera Geobotanica 18 (1) : 5-424.
Royer J.-M., 1991. Synthèse eurosibérienne, phytosociologique et phytogéographique de la classe des Festuco-Brometea. Dissertationes Botanicæ 178 : 1-296.
Rübel E., 1911. Pflanzengeographische Monographie des Berninagebietes. Botanische Jahrbücher fur Systematik, Pflanzengeschichte und Pflanzengeographie 47 : 1-616
Shimwell D.W., 1968. The phytosociology of calcareous grasslands in the British Isles. Durham theses, Durham University, 266 p.
Sillinger P., 1932. Festucetum carpaticae v Nízkých Tatrách ve srovnání s analogickou asociací v jiných částech oblasti západokarpatské. Rozpravy České Akademie Císaře Františka Josefa pro Vědy, Slovesnost a Umění, 2, vědy mathematické, přírodní, 41 (1931)/16 :1-21.
Soriano I., 1998. Las matorrales rastreros de Dryas octopetala en la vertiente sur del Pirineo oriental; aproximación sintaxonómica y ecológica Journal de botanique de la Société botanique de France 5 : 23-30.
Soriano I., 2001. La vegetació de la serra de Moixeró i el massís de la Tosa d’Alp (Pirineus orientals). Acta Botanica Barcinonensia 47 : 5-400.
Susplugas J., 1942. Le sol et la végétation dans le Haut-Vallespir (Pyrénées-Orientales). Communication de la Station internationale géobotanique méditeranéenne et alpine, Montpellier, 29 : 1-225.
Svitková I. & Šibík J., 2013. An expert-based classification of high-altitude arctic-alpine vegetation of the class Carici rupestris-Kobresietea Ohba 1974: achievements and obstacles. Plant Biosystems 147 (2) : 315-327.
Theurillat J.-P., Aeschimann D., Küpfer Ph. & Spichiger R., 1995. The higher vegetation units of the Alps. Colloques phytosociologiques XIV : 55-79.
Thiele K., 1978. Vegetationskundkliche und pflanzenökologische Untersuchungen im Wimbachgries. Aus den Naturschutzegebieten Bayerns 1 : 1-73.
Tibor S. & Tamás P., 2012. New aspects of the alpine vegetation of Parâng mountains (south Carpathians). Journal of Plant Development 19 : 99-129.
Tichý L., 2002. Juice, software for vegetation classification. Journal of Vegetation Science 13 : 451-453.
Tison J.-M. & de Foucault B. (coord.), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Tüxen R. & Oberdorfer E., 1958. Eurosibirische Phanerogamengesellschaften Spanien. Veröffentlichengen des Geobotanischen Institutes Rübel in Zürich 32 : 1-328.
Valls A., 2003. Revisió sintaxonòmica dels prats oromediterranis de l’ordre Ononidetalia striatae Br.-Bl. 1947. Acta Botanica Barcinonensia 48 : 67-198.
Vanden Berghen C., 1970. La végétation des falaises calcaires des Pyrénées occidentales (France). Bulletin du Centre d’études et de recherches ccientifiques de Biarritz 8 (2) : 291-303.
Van Es J., 2003. Étude de la végétation du site Natura 2000 « Le Mercantour ». Zone centrale du Parc national du Mercantour (département des Alpes-de-Haute-Provence). Typologie phyto-écologique des habitats. Conservatoire botanique national alpin, 239 p.
Vigo J. & Masalles R.M., 1996. El poblament vegetal de Vall de Ribes. Les comunitats vegetals i el paisatge. Mapa de vegetacio 1:50 000. Institut Cartografic de Catalunya, 468 p.
Vigo J., 1972. Notes sur les pelouses subalpines des Prépyrénées orientales. Pirineos 105 : 47-59.
Vigo J., 1979. El Ranunculo (thorae) – Seslerietum, una comunitat Pirinenca de peu de cingle. Folia Botanica Miscellanea 1 : 7-12.
Villaret J.-C., 2007. Guide des milieux ou habitats naturels et semi-naturels du département de l’Isère. Conservatoire botanique national alpin, 1049 p.
Walker D.A. et al., 2017. Circumpolar arctic vegetation classification. Phytocoenologia, DOI: 10.1127/phyto/2017/0192.
Zozomová-Lihová J., Marhold K. & Španiel S., 2014. Taxonomy and evolutionary history of Alyssum montanum (Brassicaceae) and related taxa in southwestern Europe and Morocco: diversification driven by polyploidy, geographic and ecological isolation. Taxon 63 (3) : 562-591.
Remerciements
Le premier auteur remercie Delphine Fallour-Rubio (ONF), François Prud’homme et Françoise Laigneau (CBNPMP) pour leurs relevés, Cécile Vignau (CBNPMP) pour son aide dans les recherches bilbiographiques, Vincent Boullet pour son éclairage sur une question de nomenclature et Luis Villar (IPE) pour ses indications d’interprétation taxonomique dans ses relevés originaux antérieurement publiés. Le premier auteur est redevable à Laurence Corriol d’une aide bénévole considérable dans la constitution du tableau de 1 112 relevés élémentaires pour les Pyrénées sans laquelle ce travail n’aurait pu aboutir.
Les autres auteurs remercient Jean-Charles Villaret et David Paulin (CBNA) pour leurs éclairages sur certaines associations des Alpes du Nord, ainsi que le personnel de la documentation du CBNA, en particulier Candice Winter et Marie-Hélène Marie pour leur précieuse aide dans l’obtention des références bibliographiques.
Les trois auteurs remercient Jean-Marie Royer pour les fructueux échanges, notamment sur la délimitation de certaines unités entre les classes des Festuco – Brometea et Elyno – Seslerietea. Emmanuel Catteau, Alexis Mikolajczak, Claude Misset et Virgile Noble pour leur relecture, corrections et compléments au manuscrit. Bruno de Foucault, enfin, est remercié pour avoir accepté, en tant que membre du conseil scientifique du CBNPMP, de réaliser une relecture exhaustive du manuscrit avant soumission.


