
Une station de scorsonère humble (Scorzonera humilis L.) sur le littoral méditerranéen français
Title
A population of viper’s grass (Scorzonera humilis L.) on the French Mediterranean coast
Résumé
Une population de scorsonère humble (Scorzonera humilis L.) a été détectée en basse plaine de l’Aude (commune de Fleury). Les critères morphologiques de l’espèce sont illustrés et comparés à ceux de Scorzonera parviflora Jacq. sous forme de tableau illustré. Le cortège végétal avec scorsonère humble est décrit à travers cinq relevés et son rattachement phytosociologique est discuté. Le possible statut de Scorzonera humilis en tant qu’espèce relique glaciaire est évalué et discuté
Abstract
A population of viper’s grass (Scorzonera humilis L.) has been detected in the Basse plaine de l’Aude (commune of Fleury). The morphological criteria of the species are illustrated and compared with those of Scorzonera parviflora Jacq. in an illustrated table. The plant community with Scorzonera humilis is described through five relevés and its phytosociological classification is explained. The possible status of Scorzonera humilis as a glacial relict species is assessed and discussed.
1. Répartition connue de l’espèce
La scorsonère humble est une astéracée vivace typique de végétations herbacées de stations humides et à aire de répartition eurocaucasienne (Tison, 2014a ; carte 1).
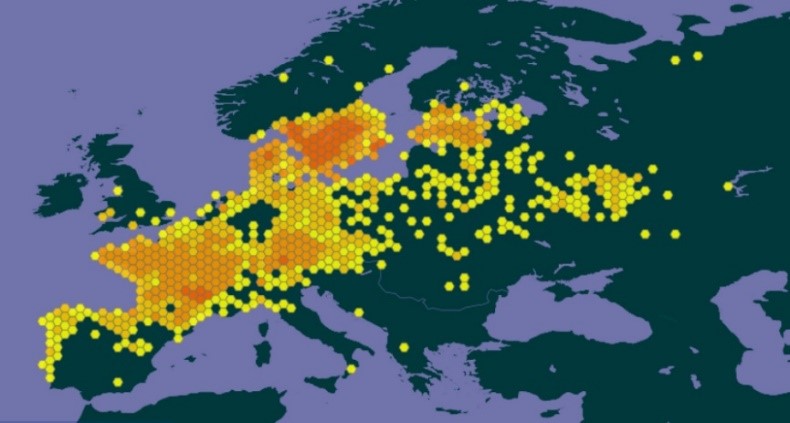
Elle est connue de beaucoup de régions françaises, mais semble très rare dans certains secteurs comme les Hauts-de-France, les alentours toulousains et la région méditerranéenne, notamment (carte 2). En effet, si l’espèce est fréquente sur les reliefs siliceux méridionaux (Cévennes, Montagne Noire, etc. ; voir p. ex. Loret & Barrandon, 1886), les données disponibles pour les plaines méditerranéennes sont très rares (carte 3) et sans doute erronées pour partie.
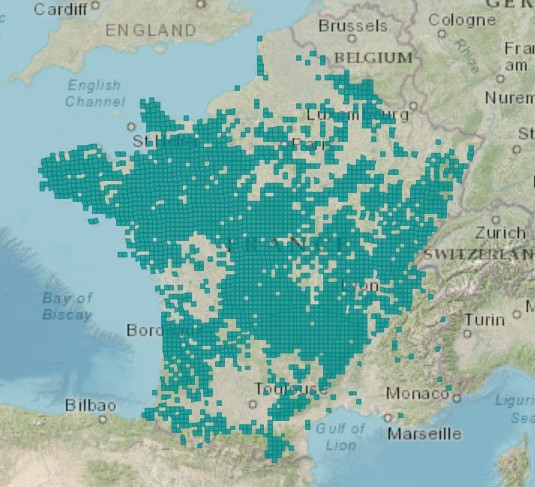
Forsskål et Niebuhr (1775) par exemple citent bien l’espèce de l’Estaque près de Marseille, mais donnent des critères qui ne lui correspondent pas (feuilles sans nervures, aigrettes non plumeuses), il doit de ce fait s’agir d’un autre taxon. Jacquemin (1851) signale Scorzonera humilis pour la ville d’Arles, sans autre précision ; de ce fait la donnée n’est pas vérifiable. Deleuil (1943 : 96) par contre indique une localité précise entre Marseille et Aubagne. Dans son Catalogue des plantes vasculaires des Bouches-du-Rhône, Molinier (s.a.) met en doute les mentions historiques et conclut, sans autre explication : « De toute manière, cette plante n’est pas indigène dans les B.-du-Rh. ». Plus récemment, Pavon et Pires (2020 : 156) se montrent plus prudents en considérant Scorzonera humilis comme « un occasionnel ancien non revu à moins qu’il n’ait été mentionné par erreur […] ».
Le point près de Montpellier indiqué par la carte 2 correspond à une observation récente transmise par PlantNet. La donnée se situe en pleine garrigue sur le massif de la Gardiole, elle nous paraît hautement improbable.
Les flores anciennes donnent quelques indications. Notamment, Grenier et Godron (1850 : 307) écrivent « se retrouve dans la région méditerranéenne (Narbonne !), Delort ». Rouy (1908 : 14) est moins précis en décrivant la répartition de la scorsonère humble ainsi : « Prairies dans presque toute la France, plus rare dans le midi ». De façon plutôt surprenante, l’abbé Coste (1936) rompt avec ses prédécesseurs et inclut Scorzonera parviflora comme simple variante à S. humilis. De ce fait, ses indications de répartition mélangent les deux taxons et perdent leur utilité dans le présent contexte.
Il existe toutefois au moins une localité historique avérée car documentée sous forme d’échantillons d’herbier : c’est en mai et juin 1888 que l’abbé Pons a récolté la scorsonère humble dans des « prés humides à Narbonne (Aude) ». Au moins deux spécimens issus de ces récoltes sont conservés à l’herbier de l’université de Montpellier (MPU817805, MPU817826). Dans son Catalogue de la flore des Corbières, Gautier (1912 : 186) précise les localités avec Scorzonera humilis près de Narbonne : « Fresquet, Moulin du Gua, Livière ». Le Moulin de Gua se situe au centre-ville, les deux autres lieux-dits se trouvent au nord-ouest de Narbonne. Soulignons donc qu’il s’agit là de localités non littorales. Il est également important de signaler que Gautier (1912) mentionne aussi bien Scorzonera humilis que S. parviflora. Il distinguait donc ces deux taxons, tout comme son collègue contemporain Timbal-Lagrave (1886).
Reste à noter que ni la thèse de Roi (1937), sur les espèces eurasiatiques continentales en région méditerranéenne, ni l’Atlas de la flore patrimoniale de l’Aude (Plassart et al., 2016) ne mentionnent Scorzonera humilis ou S. parviflora.
2. Les données initiales en basse plaine de l’Aude
La basse plaine de l’Aude est un site Natura 2000 désigné au titre des deux drectives Oiseaux et Habitats et animé par la communauté de communes (La Domitienne, 2024). C’est dans le cadre de la cartographie initiale des habitats naturels et habitats d’espèces d’intérêt communautaire qu’une première station de Scorzonera humilis a été détectée en mai 2005, à environ 3 km de distance de la mer. Au vu de l’abondance de guimauve officinale (Althaea officinalis) dans cette station, celle-ci a été rattachée à l’habitat 6430 « ourlets à Guimauve » (voir fiche-habitat correspondante ; Klesczewski & Rufray, 2007 : 46). Comme toutes les données naturalistes produites par le CEN Occitanie, ce pointage a été enregistré dans la base de données interne (SICEN) et exporté vers le SINP (Système d’information de l’inventaire du patrimoine naturel) Occitanie. Les données exportées vers le SINP font l’objet d’une procédure de validation à l’échelle régionale par le Conservatoire botanique national méditerranéen (CBNMed) en tant qu’animateur du pôle « flore – fonge – habitats ». Dans ce cadre et au vu de la singularité de la donnée de Scorzonera humilis sur le littoral audois, le CBNMed nous a contactés pour consultation d’éléments permettant une telle validation (photos, spécimen d’herbier). Ne disposant pas de tels éléments, nous avons décidé de retourner sur la station début juin 2023. Les scorsonères en question ont été retrouvées, sur exactement le même point qu’en 2006, les tiges étaient déjà sèches et les fruits envolés. Toutefois, les feuilles, parfois très grandes et à limbe large de plus de 25 mm, ainsi que les tiges sèches, épaisses et souvent ramifiées, ont été prélevées, scannées et envoyées aux experts pour avis. Au vu de la station littorale et des tiges ramifiées données par certaines clés (p. ex. Tison, 2014a) comme caractéristiques de Scorzonera parviflora, les avis des experts ont penché pour cette dernière. C’est pourquoi nous avons décidé de revisiter la station au printemps suivant, dans l’objectif de rendre la détermination certaine, à l’aide des capitules en fleurs et en fruits.
3. La population observée
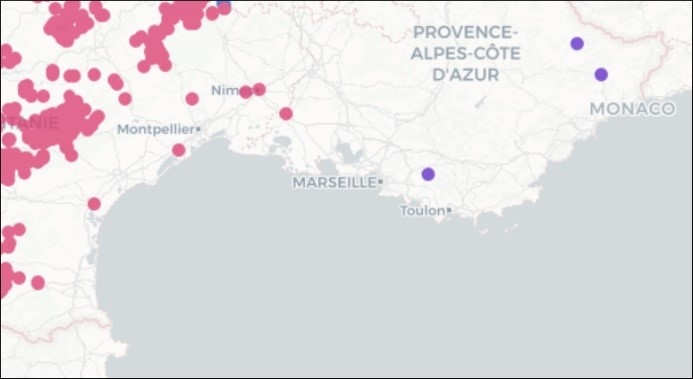
Lors de notre premier passage en 2024, le 8 avril, l’espèce était en pleine floraison, ce qui a permis de confirmer la détermination (photo 1). Ce constat nous permet de cartographier l’espèce dans son contexte régional. La carte 4 montre la distance notable avec les stations récentes les plus proches (plus de 60 km). Les stations historiques en plaine méditerranéenne comme celles de Narbonne n’ont pas été revues depuis plus de soixante-dix ans.

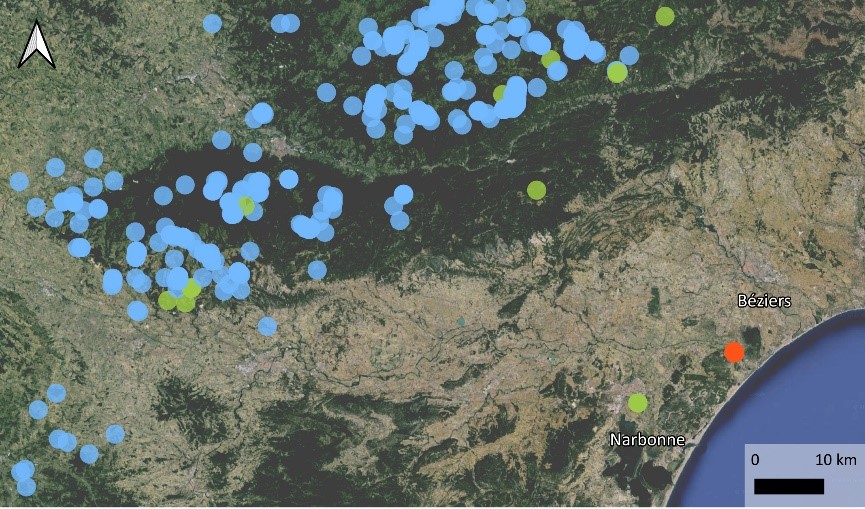
La population de Fleury s’étend sur à peine deux hectares, sous forme de deux petites taches isolées d’à peine dix individus côté nord, et la station principale de plus de deux cents pieds côté sud (carte 5). Les milieux équivalents alentours ont été prospectés, notamment vers le sud jusqu’à l’unique cladiaie de la basse plaine de l’Aude qui indique des conditions stationnelles similaires, également liées à des résurgences d’eaux karstiques (Klesczewski & Rufray, 2007).

4. Critères de détermination
Certains critères reconnus par tous les ouvrages de référence comme la longueur des ligules et la pilosité des tiges et capitules nous ont permis de lever le doute au sujet de la détermination de Scorzonera humilis à Fleury. Il nous semble que d’autres éléments morphologiques cités de façon moins concordante méritent d’être analysés plus en détail.
4.1. Couleur des akènes
Les flores françaises récentes utilisent ce critère dans la clé de détermination. La Flore Méditerranéenne (Tison, 2014b) par exemple donne pour S. humilis les « akènes mûrs (bien formés) noirâtres » et ceux de S. parviflora « roussâtres ». Or, si les akènes de S. parviflora sont toujours d’un brun clair (photo 3), ceux de S. humilis varient (même tout à fait mûrs) de brun clair à brun foncé (photo 2). Sur cette base, ce critère nous paraît relativement peu pertinent pour la distinction des deux taxons.


4.2. Ramification
Si la plupart des flores décrivent les deux taxons comme mono- et plus rarement pluricéphales (p. ex. Rouy, 1908 ; Hegi et al., 1929 ; Chater, 1976 ; Adler et al., 1994 ; Bolòs (de) & Vigo i Bonada (1995) ; Wörz, 1996 ; Goujon, 2024), la clé de Flora Gallica (Tison, 2014a) utilise ce critère en donnant Scorzonera humilis avec « tige normalement simple et monocéphale » et S. parviflora avec « tige ramifiée et pluricéphale chez les plantes robustes ». Nos observations nous amènent à considérer ce critère comme non pertinent. D’une part, S. humilis est régulièrement observée avec plusieurs capitules, à tel point que ces variantes rameuses sont décrites depuis le début du xixe siècle (Brotero, 1804 : 329 ; Hoffmansegg & Link, 1820 : 123 ; photo 4) et reconnues – notamment par Flora Iberica (Díaz de la Guardia & Blanca, 2017) – comme var. ramosa Hoffmans. & Link, 1820. Nous avons par ailleurs réalisé un comptage des pieds ramifiés à Fleury : sur 100 pieds sélectionnés au hasard, 42 étaient ramifiés.
D’autre part, les individus robustes et ramifiés de Scorzonera parviflora nous paraissent extrêmement rares, du moins en France. En vingt ans de prospections et de suivis, nous n’en avons rencontré qu’un seul exemple, dans le site des marais du Vigueirat (voir aussi Timbal-Lagrave, 1886).

4.3. Pilosité
Ce critère pourtant très intéressant n’est que rarement évoqué dans les clés de détermination (p. ex. Schubert & Vent, 1990). Il est pertinent puisque Scorzonera humilis est poilue sur ses parties végétatives, depuis la base des pétioles jusqu’aux bractées du capitule, alors que S. parviflora est totalement glabre. La pilosité de S. humilis est évidente jusqu’à sa floraison, mais peut se perdre ensuite (Hegi & Hayek, 1919 ; Adler et al., 1994).
5. Comparaison morphologique illustrée
Les deux taxons sont aisés à distinguer grâce à une série de critères faciles à observer (tableau 1). Les photos de Scorzonera humilis du tableau 1 proviennent toutes de la station à Fleury-d’Aude.







6. Interprétation phytosociologique
6.1. Les données acquises
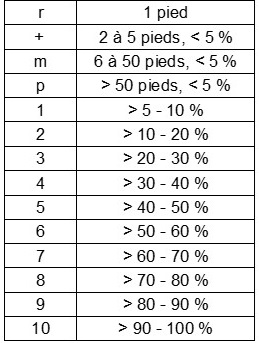
Il nous a semblé intéressant de décrire le contexte phytocénotique de Scorzonera humilis en basse plaine de l’Aude. À cette fin, nous avons réalisé cinq relevés exhaustifs centrés sur différentes sous-populations détectées, sur une surface de 25 m2. L’ensemble de la phytocénose (plantes vasculaires) a été relevé. Les recouvrements des espèces ont été notés sur la base de l’échelle de Londo (in Dierschke, 1994 ; tableau 2).
Les relevés sont détaillés en tableau 3. On observe un cortège singulier, très majoritairement vivace, constitué d’un mélange d’espèces typiques de prairies hygrophiles voire paratourbeuses (Carex distans, Juncus subnodulosus, Scorzonera humilis), de roselières (Althaea officinalis, Apium graveolens, Iris pseudacorus, Phragmites australis) ou encore de prés halophiles (Carex extensa, Juncus maritimus, Sonchus maritimus). N’abritant que 15 à 19 espèces sur 25 mètres carrés, le groupement paraît relativement spécialisé.
6.2. Rattachement phytosociologique du cortège observé
Toute interprétation phytosociologique nécessite une comparaison avec les syntaxons proches déjà connus. Pour le groupement étudié ici, nous avons pu profiter d’une compilation réalisée par Philippe Julve et mise à disposition sous forme de tableau synthétique sur eVeg (Delplanque et al., 2024 ; https://www.e-veg.net/app/5077). Ce travail concerne le Junco subnodulosi-Galietum debilis Braun-Blanq. in Braun-Blanq., Roussine & Nègre 1952, une association de « prés paratourbeux basiphiles, hygrophiles, mésoméditerranéens » (Julve, 1998 ff.). Au-delà du tableau original de l’association, cette compilation prend en compte les syntaxons suivants :
- Gentiano pneumonanthes-Cladietum marisci & Tallon 1950,
- Cirsio monspessulani-Scirpoidetum holoschoeni Braun-Blanq. ex Tchou 1948,
- Loto jordanii-Molinietum caeruleae (Braun-Blanq., Roussine & Nègre 1952) B. Foucault & Géhu 1980 [sub Dorycnio herbacei gracilis-Molinietum caeruleae (Braun-Blanq. 1931) B. Foucault & Géhu 1980],
- Dorycnio gracilis-Schoenetum nigricantis Braun-Blanq. in Braun-Blanq., Roussine & Nègre 1952 [sub Dittrichio viscosae-Schoenetum nigricantis Braun-Blanq. 1924].
Nous avons complété ces données avec la synthèse de nos relevés du tableau 3 et retrié cette compilation dans l’objectif de mettre en évidence les espèces différentielles de chaque syntaxon (tableau 5).
Ce tableau 5 met en évidence la singularité des syntaxons affichés. Avec onze espèces différentielles en commun, les relevés réalisés à Fleury (colonne 2) s’apparentent de façon très nette au Junco subnodulosi-Galietum debilis (colonne 1). Par ailleurs, ces deux colonnes se distinguent par deux groupes d’espèces différentielles. Les relevés de Fleury par exemple sont notamment caractérisés par Scorzonera humilis et Apium graveolens. Il conviendrait alors de considérer le présent groupement comme une variante géographique du Junco subnodulosi-Galietum debilis.
7. Conditions stationnelles
La présence d’espèces typiques de prés halophiles (Carex extensa, Juncus maritimus, Lotus corniculatus subsp. preslii, Plantago crassifolia, Sonchus maritimus) indique une certaine influence saumâtre. Afin de quantifier le niveau de salinité, nous avons réalisé cinq mesures en période de floraison de Scorzonera humilis (mi-avril 2024), à l’aide d’une sonde multiparamètres (Hannah instrument modèle HI98194). Trois de ces mesures ont été prises dans les flaques d’eau apparentes au sein de la station (photo 5), une quatrième dans un petit fossé qui traverse le site, la cinquième dans une mare ou résurgence située à la limite sud des pointages de scorzonères (tableau 4). Il s’avère que les mesures prises dans la nappe affleurante au sein de la station sont situées entre 3,84 et 4,3 g/l, alors que celles du fossé et de la mare/résurgence donnent des valeurs de moitié plus faibles. On pourrait en conclure que la résurgence et le fossé amènent de l’eau karstique quasi douce qui rentre ensuite en contact avec un milieu plus saumâtre.

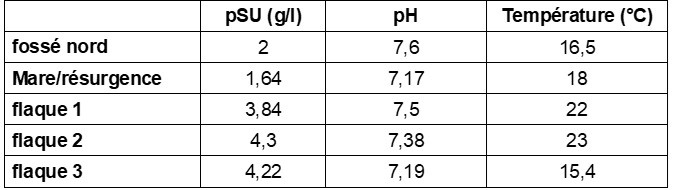
Force est de constater que les valeurs de salinité observées correspondent à des stations dites oligohalines d’après Julve (1998 ff., in Tela Botanica, 2024). La scorsonère humble arrive donc à résister à une faible salinité. Or les valences écologiques élaborées à l’échelle française Julve (1998 ff.) voire européenne (Tichý et al., 2022a, b) attribuent Scorzonera humilis à la catégorie « 0 = non tolérant ». À l’image de nos propositions au sujet de Kickxia commutata (Klesczewski et al., 2023), il nous semble que cette valence « salinité » mériterait d’être ajustée pour Scorzonera humilis.
Les observations de scorsonère humble en milieux proches du littoral sont très rares. Le taxon est historiquement cité dans des prés salés en Moravie (Rép. Tchèque ; Hegi & Hayek, 1919 : 424 ; Horvatić, 1934 : 165, sub S. candollei). Ce groupement est actuellement reconnu comme Scorzonero humilis-Juncetum maritimi Horvatić 1934 (de Foucault, 2021). Toutefois, d’après Z. Dítě (comm. pers.), le seul taxon du genre actuellement connu sur le site serait Scorzonera parviflora. Dans le tableau de synthèse de la classe compilé par Foucault (2021), Scorzonera humilis n’apparaît dans aucun autre syntaxon. De manière générale, le taxon paraît très peu connu en contexte saumâtre.
Les résurgences observées à Fleury, plus ou moins apparentes, paraissent du moins en partie équivalentes à celles décrites de Camargue par Giudicelli et al. (1980 : 271) sous le terme de « laurons » : « Ce sont des puits naturels, ouverts dans les alluvions récentes de la Crau d’où jaillit une eau claire, douce et fraîche qui se répand le plus souvent dans les marais ». Ces auteurs expliquent leur démarche ainsi : « Les botanistes, en particulier Molinier & Tallon (1949), ont relevé dans ces marais du sud de la Crau la présence d’espèces boréoalpines. […] Ils ont émis l’hypothèse que ces espèces représentent des reliques glaciaires qui se seraient maintenues dans les marais grâce à la présence des eaux froides des laurons. Il était intéressant de rechercher, dans le peuplement animal, d’éventuelles originalités faunistiques et, en particulier, l’existence de formes sténothermes d’eau froide qui pourraient être les homologues des végétaux boréoalpins cités plus haut ». En effet, Giudicelli et al. (1980 : 295) ont réussi à mettre en évidence « 28 espèces qui […] sont caractéristiques des laurons et de leur lisière. Elles présentent, pour la plupart, des exigences thermiques et des particularités d’habitat qui sont exceptionnelles dans un écosystème palustre soumis aux conditions du climat méditerranéen ». Les auteurs concluent que « l‘hydroclimat créé dans les laurons par les résurgences de la nappe de Crau confère à ce biotope un caractère d’enclave froide de type septentrional, qui échappe largement aux caractéristiques bioclimatiques de la région méditerranéenne ».
8. La scorsonère humble : une espèce relique glaciaire ?
8.1. Évaluation formalisée du statut potentiel de relique glaciaire
Le cortège végétal observé, singulier et constitué d’espèces vivaces aux exigences stationnelles très particulières, nécessite sans doute une durée considérable pour s’installer. Pour l’avoir observé il y a presque vingt ans déjà, nous pouvons aussi affirmer que ce groupement paraît très stable.
Pour le littoral méditerranéen français, ce type d’assemblage floristique a été interprété par Braun-Blanquet (1932) comme des colonies de « survivants glaciaires » qu’il définit comme « espèces immigrées à la faveur d’un climat assez froid […] qui se distinguent soit par leur distribution géographique et leur aire très disjointe, soit par leur appétences […] synécologiques » (photo 6). D’après cet auteur, les survivants glaciaires peuvent être classés en deux catégories, l’une avec les espèces montagnardes et rupicoles, l’autre regroupant les cortèges liés aux groupements prairiaux et riverains du littoral.

Dickinson (1934 : 129) ajoute une observation intéressante au sujet des espèces survivantes glaciaires : elles « se trouvent dans des stations qui ont échappé à l’influence destructrice de l’homme et où le « climax climatique » de la forêt de chênes verts n’a pas réussi à les étouffer ». Cet argument par rapport aux ligneux dominants et l’accès à la lumière a été repris par la définition récente proposée par Dítě et al. (2018) : « Compte tenu des connaissances actuelles sur l’évolution du paysage d’Europe centrale au cours du Quaternaire, nous définissons les reliques glaciaires comme des espèces au moins partiellement exigeantes en lumière qui étaient régionalement plus communes à n’importe quelle période avant l’expansion des arbres à feuilles caduques ou de résineux au début et au milieu de l’Holocène ».
Si à l’époque de Braun-Blanquet et ses élèves il s’agissait d’hypothèses peu étayées, de nombreux travaux plus récents ont apporté des arguments solides en faveur de ce raisonnement, souvent basés sur des analyses génétiques, sur des espèces aussi bien végétales qu’animales (voir p. ex. Taberlet et al. 1998 ; Guillon, 2012 ; Chiocchio et al., 2024). D’autres travaux ont proposé des éléments méthodologiques innovants comme la formalisation des arguments en faveur d’un statut relictuel des taxons.
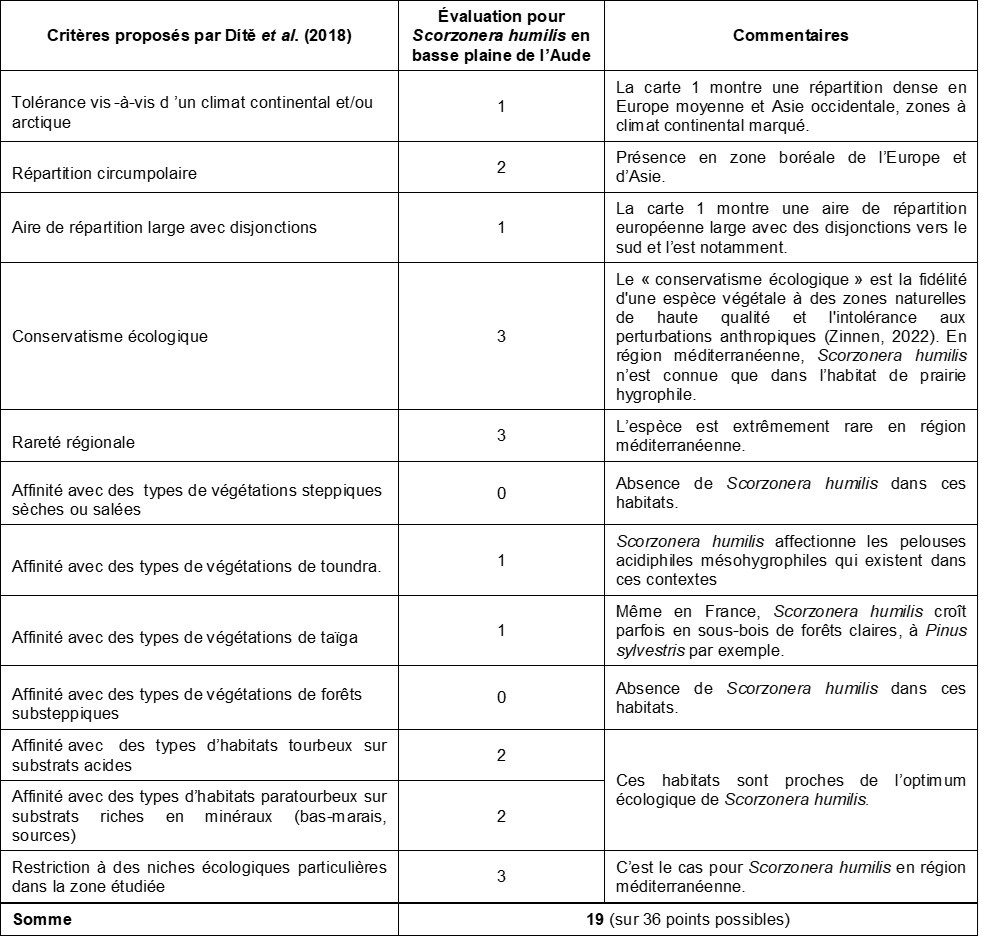
L’approche récente par Dítě et al. (2018) nous a paru particulièrement intéressante. Ces auteurs ont défini une série de douze critères argumentés vis-à-vis desquels les taxons potentiellement relictuels ont été évalués sur une échelle de 0 à 3. Ainsi est calculé un score final qui correspond à la somme des points attribués, avec un score potentiel maximal de 36. Les auteurs expliquent : « Cette somme a été utilisée comme une approximation de la probabilité qu’une espèce donnée soit effectivement une relique glaciaire. Les espèces atteignant la valeur du score final arbitrairement fixée à ≥ 10 ont été considérées comme des reliques à forte probabilité et ont donc été incluses dans les analyses ultérieures et dans la liste des reliques les plus probables ».
Le tableau 6 énumère ces critères et propose une évaluation pour le taxon Scorzonera humilis. La population de S. humilis en basse plaine de l’Aude obtient un score de 19/36, ce qui la range parmi les reliques glaciaires les plus probables d’après les critères de Dítě et al. (2018). Reste à noter que, dans le cadre de cette étude en Carpates occidentales, seules dix espèces, soit 3,5 %, arrivent à 19 points ou plus. Scorzonera humilis et S. parviflora ont d’ailleurs également été évaluées pour cette zone géographique. Ces taxons ont respectivement atteint des scores de 10 et 12 points, et font de ce fait partie de la liste des reliques glaciaires probables dans ce secteur. Sur cette base, il nous paraît évident de considérer Scorzonera humilis en basse plaine de l’Aude comme une relique glaciaire tout à fait probable.
8.2. Discussion
Braun-Blanquet esquisse dès 1932 le débat autour de l’origine des espèces eurasiatiques dans les stations influencées par les sorties d’eau douce sur le littoral méditerranéen (photo 7). Ces cortèges ont-ils survécu dans cet écosystème durant et depuis la dernière glaciation ou bien sont-elles arrivées plus récemment sous forme de semences transportées par les oiseaux, des crues ou encore le vent ?

Braun-Blanquet signale d’abord qu’il est « très difficile pour une espèce d’un autre climat de prendre pied dans la végétation déjà installée, parmi les espèces mieux adaptées […] ». En effet, quiconque ayant déjà tenté de cultiver des plantes sauvages en pleine terre aura sans doute constaté la facilité avec laquelle les espèces naturellement présentes à proximité auront pris le dessus sur le taxon introduit.
Par la suite, l’auteur développe un autre argument évident : les espèces eurasiatiques « se trouvent en effet pour la plupart réunies dans des associations végétales bien définies […] ». De notre point de vue, il est très difficile d’imaginer comment autant d’espèces auraient pu arriver par le hasard dans un même secteur pour former ces cortèges proches de groupements connus ailleurs sur le littoral méditerranéen.
Malgré ces raisonnements intrigants, Médail (2018) considère « cette intrigante question biogéographique » comme non encore définitivement close. Il cite la paléoécologue Hélène Triat (1975) qui avait formulé un avis contraire à celui de Braun-Blanquet, sans toutefois évoquer cet auteur à l’origine de la discussion : « Pour expliquer la présence de la majorité de ces taxons à aire disjointe [dans le golfe de Fos], on est conduit à invoquer plutôt un transport relativement récent de graines et diaspores par des oiseaux, pour lesquels le marais constituait un gîte, une étape et donc une survivance récente ». Quelques années plus tard, Triat-Laval et Reille (1981) décrivent des spectres polliniques en provenance d’un site du Var, à l’étage mésoméditerranéen, datant de la période dite Atlantique (entre -7500 et -3800 années ; https://fr.wikipedia.org/wiki/Atlantique_(palynozone)). Ces zones humides étaient notamment dominées par la fougère des marais (Thelypteris palustris) et des « végétations de type Holoschoenion, Cladietum ». Les auteurs supposent « que les groupements végétaux qui ont participé à l’élaboration de la tourbe à Cypéracées sont de même nature depuis la fin de l’Atlantique » (voir aussi Garnier & Garinot, 2013 : 6).
Dans ce contexte, il nous paraît judicieux de citer les conclusions de Giudicelli et al. (1980 : 296) sur les cortèges faunistiques relevés dans les laurons de Crau : « Néanmoins, la particularité remarquable de cette communauté est la présence d’espèces à la fois sténothermes froides et boréo-alpines. En cela les laurons apparaissent comme des refuges où ont pu survivre et se maintenir des espèces animales et végétales dont l’aréal s’étendait, aux périodes glaciaires, jusqu’à la région méditerranéenne ».
Médail (2018) signale par ailleurs les travaux plus récents d’Andrieu-Ponel et al. (2000) pour conclure que « ces données suggèrent donc l’existence de végétations liées à un climat plus frais et humide que l’actuel, ce qui est compatible avec une flore palustre de mise en place assez ancienne et riche en éléments septentrionaux ».
Reste à signaler que des travaux très récents et notamment celui de Chiocchio et al. (2024) démontrent que « les refuges glaciaires sont des unités très dynamiques. La plupart des zones refuges identifiées ont été détruites par l’élévation du niveau de la mer après la période glaciaire. Par conséquent, la persistance des espèces au cours du Pléistocène tardif n’a pas été assurée par la stabilité environnementale à long terme, mais par la possibilité de modifier la répartition des espèces le long des gradients altitudinaux, en fonction des changements climatiques et de l’adéquation de l’habitat ». Déjà Triat (1975 : 120) avait évoqué cette hypothèse au sujet des potentielles survivantes glaciaires du golfe de Fos. Elle la réfute sur la base d’un argument non étayé et arrive à la conclusion citée plus haut du transport relativement récent de diaspores par des oiseaux. L’hypothèse d’une telle installation d’espèces étrangères à une végétation actuelle, de type prairie hygrophile, par l’arrivée fortuite de diaspores plus ou moins isolées serait d’ailleurs intéressante à vérifier.
En conclusion, force est de constater que les chercheurs s’accordent sur l’existence postglaciaire de végétations palustres avec éléments aussi bien floristiques que faunistiques à connotation septentrionale. Si débat il y avait, il reposait sur une publication qui annonçait avoir trouvé « la solution du problème de l’origine d’espèces considérées en ce lieu comme des survivantes glaciaires » sans la fournir.
9. Gestion conservatoire de la station de Scorzonera humilis à Fleury
Deux facteurs apparaissent comme essentiels pour le maintien de cette population en état de conservation favorable : l’alimentation en eau douce et l’utilisation pastorale. Comme exposé plus haut, l’alimentation de la station par l’eau douce des résurgences constitue vraisemblablement le facteur clé ayant permis le maintien des prairies à scorsonère humble à Fleury. Il est de ce fait primordial de veiller, dans la mesure du possible, au bon fonctionnement de ces résurgences.
En termes d’utilisation agricole, les parcelles à Scorzonera humilis de Fleury sont actuellement pâturées par un troupeau de bovins Camargue, de début mai à fin juin. La période d’arrivée du troupeau dans le site paraît relativement précoce, mais à ce moment les scorsonères ont déjà fructifié et dispersé leurs akènes. De ce fait, l’espèce n’est pas perturbée dans son cycle de reproduction par l’utilisation pastorale du clos. Le « degré de raclage » relevé début août 2023 dans le clos paraissait moyen et donc adapté aux enjeux présents. Sur cette base, nous pouvons recommander de maintenir l’utilisation pastorale actuelle.
10. Perspectives
Notre étude a permis la mise en évidence du caractère exceptionnel de la station à scorsonère humble de Fleury d’Aude. La maîtrise foncière étant un facteur stratégique essentiel de conservation du patrimoine naturel, il serait important de mettre en place une veille permettant à terme l’acquisition des parcelles concernées par une structure avec mission conservatoire.
Dans ce contexte, il est important de signaler qu’au vu des changements globaux en cours ces parcelles situées à quelques mètres au-dessus du niveau de la mer sont à terme menacées de salinisation. Il apparaît alors essentiel d’anticiper les changements à venir, en incluant à la veille foncière les parcelles attenantes situées plus à l’intérieur des terres. Ces terrains pourraient à terme constituer des zones de repli pour les habitats humides de transition vers les zones plus salées.
À une échelle plus large, par exemple régionale, il nous paraîtrait intéressant de recenser les zones avec concentration d’espèces reliques glaciaires. Un tel projet ferait suite aux travaux de Braun-Blanquet et collègues, mais serait basé sur l’analyse des données désormais disponibles pour l’ensemble du territoire à l’échelle nationale, voire via le Système d’informations de l’inventaire du patrimoine naturel (SINP). Un tel recensement permettrait d’identifier de façon systématique ces zones de présence d’enjeux du patrimoine naturel. Ces « colonies d’espèces survivantes glaciaires » mériteraient d’être considérées comme des zones d’exception même si les espèces reliques qui la constituent comme la scorsonère humble ne bénéficient d’aucun statut patrimonial dans les listes régionales pour les ZNIEFF ni de statut de protection.
Bibliographie
Adler W., Oswald K. & Fischer R., 1994. Exkursionsflora von Österreich. Bestimmungsbuch für alle in Österreich wildwachsenden sowie die wichtigsten kultivierten Gefäßpflanzen (Farnpflanzen und Samenpflanzen) mit Angaben über ihre Ökologie und Verbreitung. Eugen Ulmer, Stuttgart & Wien, 1180 p.
Bolòs, O. (de) & Vigo i Bonada J., 1995. Flora manual dels Països Catalans, III – Pirolàcies – Compostes. Barcino, Barcelona, 1 230 p.
Braun-Blanquet J., 1932. Les survivants des périodes glaciaires dans la végétation méditerranéenne du Bas-Languedoc, leur valeur indicatrice et leur signification pratique. Communication de la Station internationale de géobotanique méditerranéenne et alpine (Montpellier) 16 : 1-10.
Braun-Blanquet J., Roussine N. & Nègre R., 1952. Les groupements végétaux de la France Méditerranéenne. CNRS, Paris, 297 p.
Brotero F.A., 1804. Flora lusitanica, seu plantarum, quae in Lusitania vel sponte crescunt, vel frequentius coluntur, ex florum praesertim sexubus systematice distributarum, synopsis, I. Typographia regia, Lisbonne, 607 p.
Chater A.O., 1976. Scorzonera L. In T.G. Tutin, V.H. Heywood, N.A. Burges, D.M. Moore, D.H. Valentine, S.M. Walters, D.A. Webb, A.O. Chater, R.A. DeFilipps & I.B.K. Richardson (eds.), Flora Europaea, 4 – Plantaginaceae to Compositae (and Rubiaceae), Cambridge University Press, Cambridge, London, New York, Melbourne : 317-322.
Chiocchio A., Maiorano L., Pezzarossa A., Bisconti R. & Canestrelli D., 2024. From the mountains to the sea: Rethinking Mediterranean glacial refugia as dynamic entities. Journal of Biogeography 00, 1-12, https://doi.org/10.1111/jbi.14799.
Coste H., 1937. Flore descriptive et illustrée de la France de la Corse et des contrées limitrophes, second tirage. Librairie des Sciences et des Arts, Paris, 728 p.
Deleuil G., 1943. Contributions à l’étude de la flore provençale ; localités nouvelles de plantes rares ou intéressantes et précisions sur certaines localités déjà connues. Annales de la Faculté des Sciences de Marseille 16 (2) : 87-104.
Delplanque S. et al., 2024. eVeg, une base de données pour les végétations d’Europe, https://www.e-veg.net/accueil, [05/03/2024].
Díaz de la Guardia C. & Blanca G., 2017. Scorzonera L. In S. Talavera, A. Buira, A. Quintanar, M.Á. García, M. Talavera, P. Fernández Piedra & C. Aedo (eds.), Flora Iberica – Plantas vasculares de la Península Ibérica e Islas Baleares, XVI (II) – Compositae (partim), Real Jardín Botánico, CSIC, Madrid : 773-788.
Dickinson O., 1934. Les espèces survivantes tertiaires du Bas-Languedoc. Thèse Faculté des Sciences de Montpellier (mention sciences). Impr. Lion et fils, Toulouse, 157 p.
Dierschke H., 1994. Pflanzensoziologie, Grundlagen und Methoden. UTB, Stuttgart, 683 p.
Dítě D., Hájek M., Svitková I., Košuthová A., Šoltés R. & Kliment J., 2018. Glacial-relict symptoms in the Western Carpathian flora. Folia Geobotanica 53 (3) : 277-300, https://www.jstor.org/stable/48713523.
Forsskål P. & Niebuhr C., 1775. Florula Littoris Galliae ad Estac prope Massiliam. In Flora Aegyptiaco-Arabica : sive descriptiones plantarum quas per Aegyptum inferiorem et Arabiam felicem detexit, illustravit Petrus Forskål. Möller, Haunia [Copenhague] : [I] – CXXVI, https://doi.org/10.5962/bhl.title.41.
Foucault B. (de), 2021. Contribution à une synthèse des Juncetea maritimi. Carnets botaniques 68 : 1-30 + 22 tableaux, https://doi.org/10.34971/nbat-mh09.
Garnier G. & Garinot F., 2013. Les cladiaies, un paradis pour les araignées. CEN Rhône-Alpes, Vourles, 19 p.
Gautier G., 1912. Catalogue de la flore des Corbières, mis en ordre par L. Marty. Société d’études scientifiques de l’Aude, Carcassonne, 347 p.
GBIF Secretariat, 2024. Scorzonera humilis L. in GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei [02/03/2024].
Giudicelli J., Moubayed Z. & Tourenq J.N., 1980. Un biotope hydrobiologique remarquable : les laurons de la Crau (Bouches-du-Rhône, France) ; la communauté animale et ses relations avec le peuplement des biotopes aquatiques voisins. Annales de Limnologie, International Journal of Limnology 16 (3) : 271-297, https://doi.org/10.1051/limn/1980002.
Goujon P., 2024. Scorsonère des prés (Scorzonera humilis L.), http://flore-en-ligne.fr/herbier/Scorsonere_des_pres.html.
Grenier C. & Godron D.-A., 1850. Flore de France ou description des plantes qui croissent naturellement en France et en Corse, II. Sainte-Agathe Aîné, Besançon, 760 p.
Guillon M., 2012. De la physiologie à la répartition : adaptations climatiques et sensibilité thermique chez une relique glaciaire. Sciences de l’environnement. Faculté des Sciences fondamentales et appliquées, Université de Poitiers, HAL Id: tel-00996071, https://theses.hal.science/tel-00996071.
Hegi G. & von Hayek A., 1919. Illustrierte Flora von Mitteleuropa. Mit besonderer Berücksichtigung von Oesterreich, Deutschland und der Schweiz. Zum Gebrauche in den Schulen und zum Selbstunterricht, VI. A. Pichler’s Witwe & Sohn, Wien : 1-544.
Hegi G., Beger H., Gams H., von Hayek A. & Zahn H., 1929. Illustrierte Flora von Mitteleuropa. Mit besonderer Berücksichtigung von Oesterreich, Deutschland und der Schweiz. Zum Gebrauche in den Schulen und zum Selbstunterricht. VI (2), Dicotyledones (V). Sympetalae (Schluss der Compositae). A. Pichler’s Witwe & Sohn, Wien : 549-1386.
Hoffmansegg J.C. & Link H.F., 1820. Flore portugaise ou description de toutes les plantes qui croissent naturellement en Portugal avec figures coloriées, cinq planches de terminologie et une carte, II. Imprimerie des auteurs, Berlin, 504 p. & 109 gravures, https://bibdigital.rjb.csic.es/idurl/1/9784.
Horvatić S., 1934. Flora i vegetacija otoka Paga. Prirodoslovna istraživanja kraljevine Jugoslavije 19 : 116-372.
Jacquemin L., 1851. Essai de statistique sur la ville d’Arles et son territoire. Répertoire des travaux de la Société de statistique de Marseille 14 : 109-312, https://gallica.bnf.fr/ark:/12148/bpt6k57751223/f111.item.
Julve Ph., 1998 ff. Baseveg, répertoire synonymique des groupements végétaux de France, version du 11 juin 2023, http://perso.wanadoo.fr/philippe.julve/catminat.htm.
Julve Ph., 1998 ff. Baseflor, index botanique, écologique et chorologique de la flore de France, version du 27 avril 2021, http://perso.wanadoo.fr/philippe.julve/catminat.htm.
Klesczewski M., Rossi S., Coulon F., Dubois A. & Pagano N., 2023. L’étang de l’Or, bastion héraultais pour la kickxie variable (Kickxia commutata). Carnets botaniques 159 : 1-10, https://doi.org/10.34971/N8ZH-X193.
Klesczewski M. & Rufray X., 2007. Document d’objectifs du site Natura 2000 « Basse Plaine de l’Aude » valant Plan de gestion ; cartes, fiches et annexes à l’état des lieux, SIC FR 9110108 et ZPS FR 9101435. https://www.occitanie.developpement-durable.gouv.fr/docob-de-la-basse-plaine-de-l-aude-a1125.html.
La Domitienne, 2024. https://www.ladomitienne.com/services/espaces-naturels/basse-plaine-de-laude/ [27/05/2024].
Loret H. & Barrandon A., 1886. Flore de Montpellier ou Analyse descriptive des plantes vasculaires de l´Hérault, 2e édition revue et corrigée par Henri Loret. Masson, Paris, 663 p.
Médail F., 2018. Mise en place de la flore : approche biogéographique. In M. Pires & D. Pavon (coord.), La flore remarquable des Bouches-du-Rhône, plantes, milieux naturels et paysages, Biotope, Mèze : 22-51. https://amu.hal.science/hal-01888954/document.
MNHN & OFB [eds], 2003-2024. Fiche de Scorzonera humilis L., 1753. Inventaire national du patrimoine naturel (INPN), https://inpn.mnhn.fr/espece/cd_nom/121960 [02/03/2024].
Molinier R., s. a. Catalogue des plantes vasculaires des Bouches-du-Rhône (ouvrage publié à titre posthume avec la participation de Paul Martin). Imprimerie municipale de Marseille, 375 p.
Molinier R. & Tallon G., 1949-50. La végétation de la Crau (Basse-Provence). Revue générale de botanique 56 : 525-540, 57 : 48-61, 97-127, 177-192, 235-251, 300-315.
Pavon D. & Pires M., 2020. Flore des Bouches-du-Rhône. Naturalia, Turriers, 351 p.
Plassart C., Barreau D. & Andrieu F. (coord.), 2016. Atlas de la flore patrimoniale de l’Aude. Biotope, Mèze, 432 p.
Roi J., 1937. Les espèces eurasiatiques continentales et les espèces boréo-alpines dans la région méditerranéenne occidentale. Thèse Faculté des sciences de Montpellier (mention sciences). Imprimerie Fournié, Toulouse, 216 p.
Rouy G., 1908. Flore de France ou description des plantes qui croissent spontanément en France, en Corse et en Alsace-Lorraine, X. Société des sciences naturelles de la Charente-inférieure, 404 p.
Schubert R. & Vent W. (éds.), 1990. Exkursionsflora von Deutschland, 4, Kritischer Band. 8. Aufl. Volk und Wissen, Berlin, 811 p.
Svenning J. C., Normand S. & Kageyama M. 2008. Glacial refugia of temperate trees in Europe: insights from species distribution modelling. Journal of Ecology 96 :1117-1127, https://doi.org/10.1111/j.1365-2745.2008.01422.x.
Taberlet P., Fumagalli L., Wust-Saucy A.G. & Cosson J.-F. 1998. Comparative phylogeography and postglacial colonization routes in Europe. Molecular Ecology 7 :453-464.
Tela Botanica, 2024. eFlore, L’encyclopédie botanique collaborative, fiche Scorzonera humilis, https://www.tela-botanica.org/bdtfx-nn-61914-synthese [18/05/2024].
Tichý L., Axmanová I., Dengler J., Guarino R., Jansen F., Midolo G., Nobis M.P., Van Meerbeek K., Aćić S., Attorre F., Bergmeier E., Biurrun I., Bonari G., Bruelheide H., Campos J.A., Čarni A., Chiarucci A., Ćuk M., Ćušterevska R., Didukh Y., Dítě D., Dítě Z., Dziuba T., Fanelli G., Fernández-Pascual E., Garbolino E., Gavilán R.G., Gégout J.-C., Graf U., Güler B., Hájek M., Hennekens S.M., Jandt U., Jašková A., Jiménez-Alfaro B., Julve P., Kambach S., Karger D.N., Karrer G., Kavgacı A., Knollová I., Kuzemko A., Küzmič F., Landucci F., Lengyel A., Lenoir J., Marcenò C., Moeslund J.E., Novák P., Pérez-Haase A., Peterka T., Pielech R., Pignatti A., Rašomavičius V., Rūsiņa S., Saatkamp A., Šilc U., Škvorc Ž., Theurillat J.-P., Wohlgemuth T. & Chytrý M., 2022a. Ellenberg-type indicator values for European vascular plant species. Journal of Vegetation Science 34, e13168, https://doi.org/10.1111/jvs.13168.
Tichý L., Axmanová I., Dengler J., Guarino R., Jansen F., Midolo G., Nobis M.P., Van Meerbeek K., Aćić S., Attorre F., Bergmeier E., Biurrun I., Bonari G., Bruelheide H., Campos J.A., Čarni A., Chiarucci A., Ćuk M., Ćušterevska R.,… Chytrý M., 2022b. Ellenberg-type indicator values for European vascular plant species, https://doi.org/10.5281/zenodo.7427088.
Timbal-Lagrave Éd., 1886. Essai monographique sur les espèces du genre Scorzonera L. de la Flore française. Mémoires de l’Académie des sciences, inscriptions et belles-lettres de Toulouse, 8e série, VIII : 379-394, https://gallica.bnf.fr/ark:/12148/bpt6k57202762/f383.item.zoom.
Tison J.-M., 2014a. Scorzonera L. In J.-M. Tison & B. de Foucault (coords.), Flora Gallica, Flore de France, Biotope, Mèze : 474-477.
Tison J.-M., 2014b. Scorzonera L. In J.-M. Tison, Ph. Jauzein & H. Michaud, Flore de la France méditerranéenne continentale, Naturalia Publications, Turriers : 1738-1741.
Triat H., 1975. L’analyse pollinique de la tourbière de Fos-sur-Mer (B. du Rh.). Ecologia Mediterranea 1 : 109-121. https://doi.org/10.3406/ecmed.1975.914 ; https://www.persee.fr/doc/ecmed_0153-8756_1975_num_1_1_914.
Triat-Laval H., 1978. Contribution pollenanalytique à l’histoire tardi-et postglaciaire de la végétation de la basse vallée du Rhône. Thèse de doctorat ès sciences, Université Aix-Marseille III, 343 p., 34 fig. 5 tab., 29 diag. h.t.
Triat-Laval H. & Reille M., 1981. Analyse pollinique d’une séquence tourbeuse de Provence orientale (Seillons-Source d’Argens, Var, France). Ecologia Mediterranea 7 (2) : 31-37, https://doi.org/10.3406/ecmed.1981.993 ; https://www.persee.fr/doc/ecmed_0153-8756_1981_num_7_2_993.
Wörz A., 1996. Scorzonera L., 1753. In O. Sebald, S. Seybold, G. Philippi & A. Wörz (eds.), Die Farn- und Blütenpflanzen Baden-Württembergs, 6 – Spezieller Teil (Spermatophyta, Unterklasse Asteridae) – Valerianaceae bis Asteraceae, Eugen Ulmer, Stuttgart : 324-331.
Zinnen J., 2022. The concept of ecological conservatism and the characteristics of conservative plant species. Dissertation University of Illinois Urbana-Champaign, 221 p., https://www.ideals.illinois.edu/items/125373.
Remerciements
Merci beaucoup aux personnes suivantes : Bruno de Foucault (relecture du manuscrit et corrections syntaxonomiques), Arthur de France (Syndicat mixte de la Camargue gardoise ; envoi de mesures et de photos de Scorzonera parviflora), Rémi Jullian (référent littoral au CEN Occitanie ; soutien logistique), Aymeric Brissaud (chargé de projets territorial Ouest-Hérault, CEN Occitanie ; accompagnement prospections de terrain), Jean-Marc Tison (aide à la détermination de Taraxacum), Zuzana Dítě (Plant Science and Biodiversity Center, Slovak Academy of Sciences ; envoi d’informations et de bibliographie). Plusieurs journées de terrain en vue de cette publication ont été réalisées grâce au projet européen LIFE Biodiv’Paysanne (https://life-biodivpaysanne.fr/), dans le cadre de la rédaction du plan de gestion multi-sites de la basse plaine de l’Aude.


