
Bilan en 2022 et 2023 des sous-populations de l’endémique corse Allium corsicum (Amaryllidaceae)
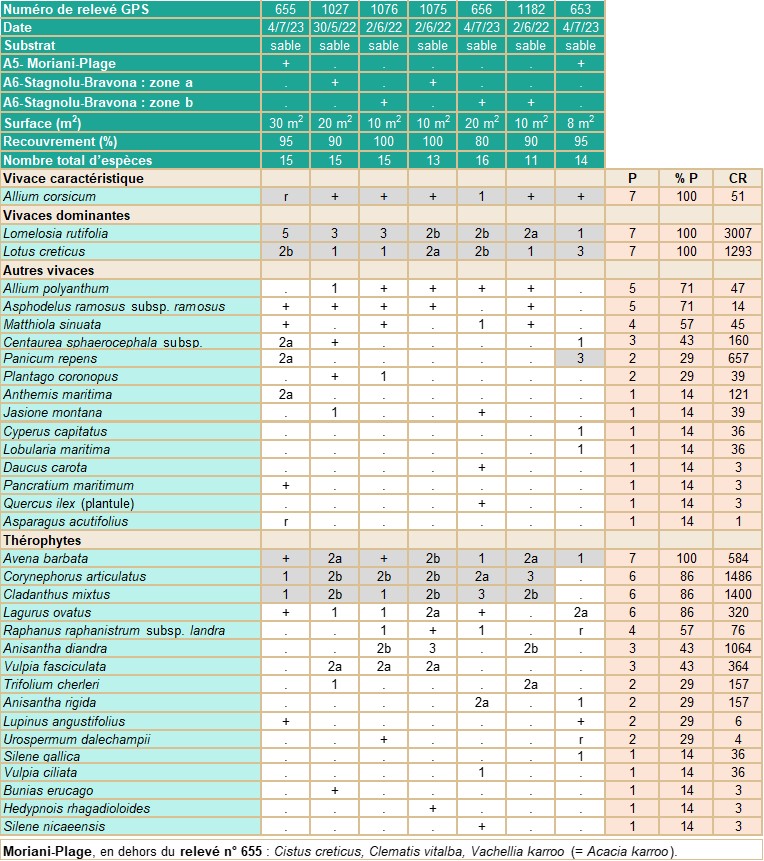
- Carnets botaniques n°180 - Tableau 1. Groupement à Lomelosia rutifolia présentant Allium corsicum.
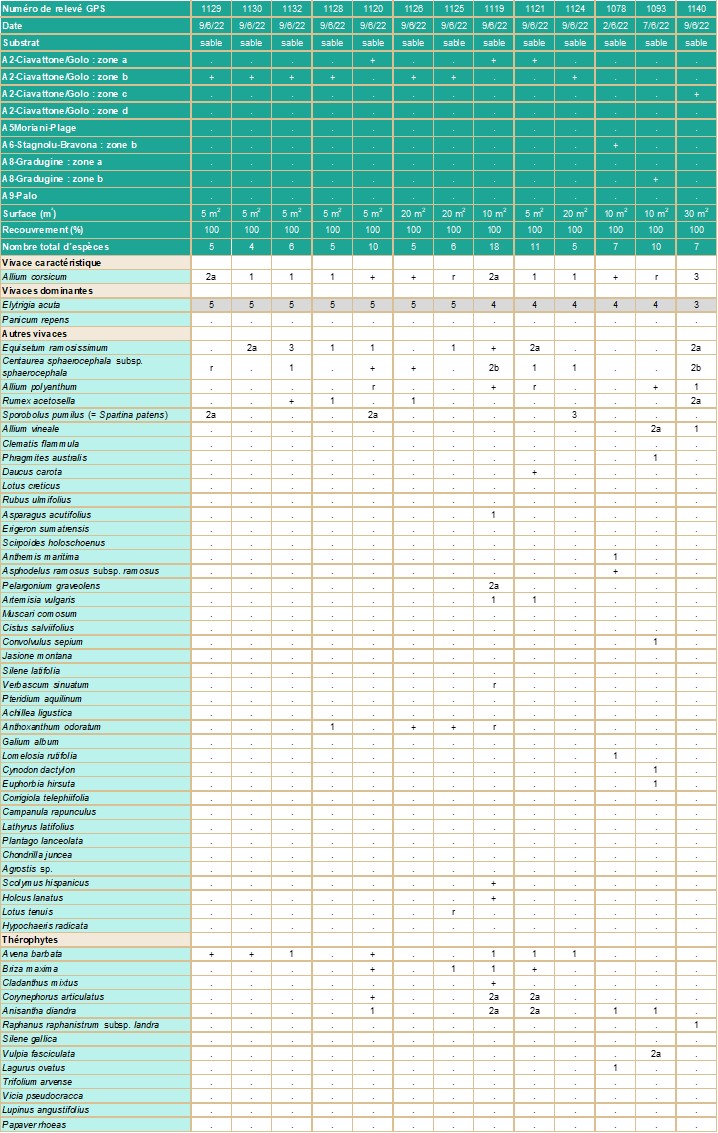
- Carnets botaniques n°180 - Tableau 2. Groupement à Elytrigia acuta présentant Allium corsicum (cf. annexe 1).
- Carnets botaniques n°180 - Tableau 3. Groupement des pelouses thérophytiques présentant Allium corsicum (cf. annexe 2).
- Carnets botaniques n°180 - Tableau 4. Aspects règlementaire et foncier des 11 sites où Allium corsicum a été signalé
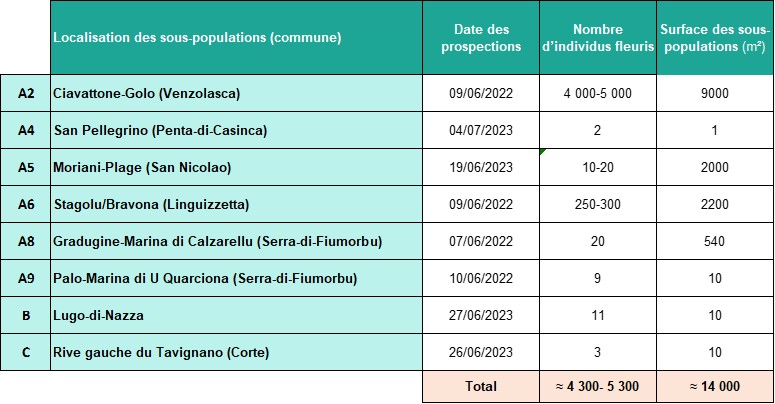
- Carnets botaniques n°180 - Tableau 5. Synthèse du nombre d’individus estimé et des surfaces occupées par A. corsicum en 2022-2023
Title
The corsican endemic species Allium corsicum (Amaryllidaceae) in Corsica: status in 2022 and 2023
Résumé
Allium corsicum est un taxon endémique strict de la Corse, dont l’enjeu de conservation est considéré comme « très fort ». Un bilan stationnel, effectué en 2022 et 2023, montre qu’il ne comprend que sept sous-populations : cinq dans la Plaine orientale, une en semi-montagne à Lugo-di-Nazza et une en bordure du fleuve Tavignano, près de Corte. Le nombre total d’individus est estimé à près de 5 000, sur une superficie d’environ 1,4 ha.
Abstract
Allium corsicum is a strict endemic taxon of Corsica, whose its conservation issue is considered “very strong”. A stationary assessment, carried out in 2022 and 2023, shows that it only includes seven subpopulations: five in the Eastern Plain, one in the semi-mountains at Lugo-di-Nazza and one on the edge of the Tavignano river, near Corte. The total number of individuals is estimated at nearly 5,000, over an area of approximately 1.4 ha.
Introduction
L’ail de Corse (Allium corsicum Jauzein, J.-M. Tison, Deschâtres & H. Couderc, 2002), taxon endémique corse, est considéré comme rare. Par suite de sa description récente (2002), il ne bénéficie d’aucun statut règlementaire. Il est cependant inscrit sur la liste des espèces déterminantes de l’inventaire ZNIEFF (zones naturelles d’intérêt écologique, faunistique et floristique) pour la Corse. En ce qui concerne les évaluations UICN, il est considéré comme :
– CR (« en danger critique d’extinction »), dans la Liste rouge mondiale des espèces menacées ainsi que dans la Liste rouge européenne (évaluation 2011),
– VU (« vulnérable »), dans la Liste rouge des espèces menacées en France (UICN France et al., 2018) ainsi que dans la Liste rouge régionale de la flore vasculaire de Corse (Delage & Hugot, 2015) [d’après le critère D2] (Note 1).
En 2021, le Conservatoire botanique national de Corse (CBNC), dans le cadre de la rédaction de sa stratégie de conservation pour la flore corse (Piazza et al., 2021), a réalisé une hiérarchisation des taxons selon trois critères : la rareté biogéographique, la rareté locale et les menaces (vulnérabilité de l’habitat et artificialisation). L’application de ces critères à l’ensemble des taxons de la flore corse (hors taxons exotiques) a permis d’attribuer une note à chacun d’entre eux, allant de 15 pour les taxons à enjeu de conservation le plus fort, à 3 pour les taxons à enjeu de conservation le plus faible. L’ail de Corse a obtenu, en l’état des connaissances en 2021, la note de vulnérabilité la plus forte, soit 15. C’est donc, selon cette méthodologie, le taxon dont l’enjeu de conservation est le plus fort sur l’île.
La stratégie de conservation ayant été réalisée avec des données de localisation du taxon recueillies entre 2000 et 2021, il a été décidé de faire un bilan stationnel le plus précis possible pour réactualiser et compléter les données existantes (Note 2). En effet, la répartition de ce taxon, relativement discret, peut avoir été sous-estimée et, depuis sa description (Jauzein et al., 2002), il n’y a eu que très peu de nouvelles prospections et quasiment aucun dénombrement de ses individus n’a été effectué. Aussi, nous avons visité toutes les stations déjà connues et, en plus, nous avons recherché des sites potentiellement favorables à sa présence. Un rapport détaillé a été rédigé (Piazza & Paradis, 2023). Cet article résume les résultats de nos prospections.
1. Présentation d’Allium corsicum (Amaryllidaceae)
1.1. Taxonomie et iconographie (Jauzein et al., 2002)
Au début des années 1980, les premiers échantillons récoltés par Deschâtres avaient été classés provisoirement comme Allium gr. pallens, « bien que certains caractères du taxon corse soient éloignés de ce qu’on appelle en Méditerranée occidentale A. pallens L. ou A. coppoleri Tineo, ces deux noms étant considérés ou non comme synonymes selon les auteurs » (Jauzein et al., 2002). Ces échantillons ne correspondant à aucun taxon du groupe paniculatum s. l. (A. paniculatum L., A. savii Parl., A. pallens L. subsp. pallens, A. pallens subsp. tenuiflorum (Ten.) Stearn, A. oleraceum L., A. longispathum F. Delaroche), Jauzein et al. (2002) ont décrit un nouveau taxon sous le nom d’Allium corsicum Jauzein, J.-M. Tison, Deschâtres & H. Couderc, 2002. Cet ail appartient au sous-genre Codonoprasum (Rchb.) Zahar., section Codonoprasum Rchb.
1.2. Caryologie
Des comptages chromosomiques ont été réalisés sur des méristèmes racinaires provenant d’individus de Serra-di-Fiumorbo et de San Nicolao (Jauzein et al., 2002). Ils montrent une garniture chromosomique de 24 chromosomes. Le nombre de base de la section Codonoprasum est admis comme étant constant :
x = 8. Aussi, le caryotype d’A. corsicum est interprété comme triploïde par Jauzein et al. (2002).
Par ailleurs, ces auteurs soulignent que : 1/ les chromosomes sont de grande taille, ce qui est la règle dans le genre Allium ; 2/ le degré de symétrie est élevé, la totalité des chromosomes étant de type m, ce qui est fréquent dans ce qui est nommé le « groupe paniculatum » ; 3/ le nombre 2n = 24 est rare dans le « groupe paniculatum ».
1.3. Description morphologique (Jauzein et al., 2002) (planches 1 à 3)
Géophyte bulbeux de 25 à 60 cm de hauteur ; bulbe brièvement ovoïde à subsphérique d’environ 1,7-2 cm de large et 1,5-2 cm de hauteur, à tunique externe grisâtre.
Scape florifère (tige) arrondie ; feuilles glabres et planes, de 2-4 mm de largeur.
Spathe bivalve à parties inégales, la plus longue pouvant atteindre 4,5 cm et dépasser un peu l’inflorescence.
Inflorescence : ombelle multiflore, lâche, retombante de 4-7 cm de diamètre, à fleurs généralement nombreuses (jusqu’à 75), à pédicelles peu inégaux, de 20-25 mm.
Fleur : périgone long d’environ 5-6 mm, campanulé, blanc nacré lavé de rose, à stries brun pourpré, tépales rétus, étamines dépassant les tépales de 0,5 à 1 mm, ovaire de 3,5 à 4 m, fusiforme et subtronqué à l’apex.
Fruit : capsule subglobuleuse, de 4,5 à 5,5 mm.
Graines très rares, de couleur noire, de 3 mm de long et 2-2,5 mm de large.



1.4. Phénologie et cycle de vie
L’ail de Corse fleurit de la mi-mai à la fin juin, ce qui est très précoce pour un membre de la section Codonoprasum. Les scapes florifères restent visibles jusqu’à la fin de l’été. Dans leur publication, Jauzein et al. (2002) notent que la floraison est « sporadique à l’état sauvage : hormis pour la station de San Nicolao, les pieds fleuris ont toujours été observés isolés ou en petit nombre, et rarement deux années consécutives au même endroit, alors que la fouille du sable environnant révèle toujours de nombreux bulbes ».
Néanmoins, nos prospections nous ont permis d’observer, le 9 juin 2022, à proximité d’une petite station connue, une population de plusieurs milliers d’individus en pleine floraison, localisée entre la rive droite de l’embouchure du Golo et la rive gauche de l’embouchure du Ciavattone, dans des prairies plus ou moins humides (planche 4, A et B). L’année suivante, les pieds fleuris étaient toujours très nombreux sur la station, mais pas aux mêmes endroits qu’en 2022, ce qui confirme les observations de Jauzein et al. (2002).
Les premières feuilles apparaissent en fin d’été (début septembre ; planche 4, C) et disparaissent en fin d’hiver (fin mars). La plante n’est alors plus visible jusqu’à l’apparition des scapes florifères en mai-juin. Jauzein et al. (2002) soulignent que : « lors de printemps secs, la plante disparaît encore plus tôt et ne fleurit pas : ainsi, la station si riche de San Nicolao, où les pieds fleuris se comptaient localement par plus de dix au mètre carré en juin 1998, ne montrait aucun organe aérien de cet ail à la mi-avril 2000 ».

1.5. Importance de la multiplication végétative (planche 2, E ; planche 3, B à F ; planche 4, C et E)
Allium corsicum est un taxon principalement apomictique, se propageant surtout par caïeux souterrains. La multiplication sexuée est rare. Bien qu’elle soit visitée par de nombreux insectes, l’espèce fructifie très peu. Ainsi, seules deux graines ont été trouvées dans un échantillon de cinq ombelles (soit près de quatre cents fleurs) prélevé en 2022, en rive droite du Golo.
In situ, Jauzein et al. (2022) ne l’ont jamais vu en fruits et soulignent qu’« en culture, ses capsules, pour la plupart vides ou monospermes, ont été observées à raison de 1 pour 5 ombelles environ dans les meilleures années. Le taux de germination est en revanche élevé : 80% sur 5 graines. Allium corsicum possède un pollen avorté à 60-80%, mais ce point n’explique pas forcément sa stérilité ». Ainsi, la multiplication végétative semble nettement privilégiée chez ce taxon. D’après ces mêmes auteurs, les colonies d’A. corsicum ont un aspect hivernal très gazonnant. Nos propres observations de terrain le confirment. Pour ces auteurs, il est « avéré que cette densité contribue à diminuer la floraison car, en culture, celle-ci est très améliorée par la dissémination des bulbes ».
Jauzein et al. (2002) ont observé que « chaque bulbe-mère adulte, outre son propre caïeu sessile de renouvellement, produit 4 à 12 caïeux pédicellés en cours d’hiver. Chacun de ces caïeux se forme sur un stolon axillaire feuillé et enraciné, donc partiellement autonome. Les plus longs de ces stolons peuvent dépasser 5 cm (planche 2, E ; planche 4, E). Les premières innovations apparaissent en même temps que les premières feuilles à l’automne, sous forme de faisceaux axillaires conférant à la pousse un aspect caractéristique en gerbe. Cette phase de multiplication végétative intervient au début du cycle annuel ; dès janvier-février, les stolons disparaissent et les jeunes plantes poursuivent leur développement indépendamment du bulbe-mère. Cette précocité explique apparemment la capacité qu’a la plante de disparaître très tôt en cas de printemps sec, sans que sa mise en réserve en soit gravement affectée ».
1.6. Chorologie (planche 5)
Allium corsicum est un taxon endémique strict de Corse. Il est présent dans la Plaine orientale (planche 5 : A1 à A9), à Lugo-di-Nazza (planche 5 : B) et au bord du Tavignano, non loin de Corte (planche 5 : C) (Jauzein et al., 2002 ; Delage et al., 2018). Jauzein et al. (2002) l’ont signalé dans les localités suivantes :
- Biguglia : plage près du déversoir de l’étang, VI.1987, Jauzein s.n. (Hb. privé) ;
- Venzolasca : plage près du village de Cap Sud, 10.VI.1980, Deschâtres s.n. (Hb. privé) ;
- secteur Plaine orientale : Padulella, 500 m au sud de Moriani-Plage, 42° 22’ N, 9° 32’ E, alt. 3 m, arrière-plage, sables fixés, assez abondant, 14.VI.1990, Jeanmonod & M.-A. Thiébaud (G 55140) ;
- San Nicolao : cordon dunaire à 100 m au sud de Moriani-Plage, 02.VI.1998, -M. Tison s.n. (Hb. privé) ;
- Aleria : plage de Padulone, 11.VI.1981, Deschâtres s.n. (Hb. privé) ;
- Serra-di-Fiumorbo : plage de Gradugine, ex cultis, VI.1997, -M. Tison s.n. (G, P, MPU, Hb. privé).
En outre, Jauzein et al. (2002) notent que « la plante a été également observée par Dutartre (comm. pers.) à Anghione, plage de Pinarello, en 1981 ; ce botaniste n’en a pas conservé d’exsiccatum, mais l’a mise en culture au Jardin Botanique de Lyon, où la souche a disparu et qu’une mention de Wilde-Duyfjes (1976) sous « AIlium paniculatum L. » : « Bastia, sables, Requien s.n. (FI) » pourrait se rapporter à ce taxon, compte tenu de sa localité et de son écologie ».
De nouvelles mentions ont été publiées par Delage et al. (2018) :
- commune de Venzolasca, embouchure du Golo, à l’E de Terragliolo, au bord de la piste allant à la plage, à environ 200 m de la mer, arrière-dune, environ 1 m d’altitude, quelques dizaines de plantes fleuries, 31.V.2016, Delage, pointage GPS ;
- commune de Linguizzetta : NW de la Marine de Bravona, route d’accès à la plage au sud de l’étang de Stagnolu, à environ 1 km de la plage, un peu à l’est du lieu-dit Testaccia, pelouse en bord de route, en lisière de cistaie, une vingtaine de plantes fleuries, 6.VI.2010, Delage (Hb. CBNC), pointage GPS, photos ;
- commune de Linguizzetta : N de la Marine de Bravona, au sud de la lagune de Stagnolu, dépression arrière-dunaire à environ 150 m au sud de l’embouchure de l’étang, quelques dizaines de plantes fleuries, 6.VI.2015, Delage (Hb. CBNC), pointage GPS, photos ;
- Corte : rive gauche du Tavignano, à 3 km en aval de la ville (42° 18’ N, 9° 10’ E), sables et galets, 21.V.1992, Deschâtres s.n. (G), dét. C. Habashi, 2014 ;
- Lugo-di Nazza (42° 05’ N, 9°18’ E), rocailles, serpentine, 17.VI.1981, Deschâtres s.n. (G), dét. C. Habashi, 2014.
En outre, Delage a également observé le taxon sur la commune de Penta di Casinca, à environ 560 m au N de la plage de San Pellegrino, dans une pelouse en bordure d’une dépression humide (base de données du CBNC).
Ces mentions complètent la répartition de ce taxon, qui de très rare (RR : moins de cinq stations) au début des années 2000 est passé au statut de rare (R : entre cinq et dix stations).

1.7. Écologie, syntaxonomie et habitats
Les premières observations d’A. corsicum ont fait supposer qu’il était inféodé aux milieux dunaires ou proches des dunes. Ainsi, Jauzein et al. (2002) écrivent « A. corsicum a toujours été observé dans des sables littoraux. Ses compagnes les plus fréquentes sont Vulpia fasciculata (Forssk.) Fritsch, Corynephorus articulatus aggr., Elytrigia juncea (L.) Nevski, Silene nicaeensis Ali., Anthemis maritima L., Centaurea sphaerocephala L., Ononis variegata L., Pycnocomon rutifolium (Vahl) Hoffmanns. & Link. Il est donc lié à la classe des Ammophiletea… Évitant l’influence maritime directe, il apparaît soit sur le versant interne des dunes s’il y en a, soit, si le terrain est plat, à une certaine distance de la mer (80-100 m). Il peut ensuite s’étendre vers l’intérieur aussi loin que vont les sables mobiles, fleurissant apparemment mieux dans les dépressions où se maintient une certaine humidité hivernale. Selon toute apparence, il est réfractaire à tout biotope fermé et à l’eau stagnante ».
Ultérieurement, on s’est aperçu qu’il n’était pas uniquement présent dans les habitats dunaires. Ainsi, les plantes récoltées par Deschâtres et conservées dans l’herbier de Genève ont été prélevées d’une part, le long du Tavignano, en aval de Corte, sur une terrasse alluviale, à 350 m d’altitude, et d’autre part près de Lugo-di-Nazza, à 450 m d’altitude, dans un éboulis de serpentinite (CBNC, 2020). De même, Delage et al. (2018), en se basant sur leurs propres observations et sur la littérature, constatent que « si l’une des populations (Stagnolu/Bravona) se localise, comme ordinairement pour l’espèce, dans une dépression arrière-dunaire, trois autres (San Pellegrino, Tavignano, Lugo-di-Nazza) poussent dans des écologies tout à fait inhabituelles, dans un milieu anthropisé (bord de route en lisière de cistaie), sur des rocailles ou sur des galets et sables, en dehors de l’influence maritime et pas sur sables mobiles ».
Nos prospections de 2022-2023 confirment toutes ces observations : l’ail de Corse vit dans plusieurs types d’habitats. Ceux-ci sont parfois difficiles à caractériser car ils correspondent souvent à des zones de transition écologique (écotones) entre plusieurs écosystèmes. Néanmoins, on trouve l’ail, a minima, dans les habitats décrits ci-dessous, soit présents seuls, soit le plus souvent en mosaïque les uns avec les autres. Dans ce qui suit, la nomenclature syntaxonomique est, en général, celle du Prodrome des végétations de France (Bardat et al., 2004, abrégé en PVF 2004) et celle de Reymann et al. (2016). La nomenclature des habitats suit CORINE Biotopes (Devillers et al., 1991 ; ENGREF, 1997), EUNIS (CBNC, 2016) et, dans quelques cas, le Manuel d’interprétation des habitats de l’Union européenne (Anonyme, 1999 ; Anonymous, 2007).
Présence dans les végétations d’arrière-dune (tableau 1 ; planche 6)
Sur les sites de Stagnolu-Bravona (tableau 1, relevés 653 et 655) et de Moriani-Plage (tableau 1, relevés 656, 1027, 1075, 1076 et 1182), A. corsicum se trouve, pour partie, sur du sable submobile d’arrière-dune, dans les végétations thermophiles, constituées d’une mosaïque entre (a) des espèces vivaces dominées par Lomelosia rutifolia (= Pycnocomon rutifolium) et (b) des thérophytes dominées par Avena barbata, Corynephorus articulatus, Vulpia fasciculata et Cladanthus mixtus. La syntaxonomie des groupements et la codification des habitats sont les suivantes :
a. Euphorbio paraliae-Ammophiletea australis Géhu & Géhu-Franck 1988 corr. Géhu 2004
Crucianelletalia maritimae G. Sissingh 1974
Crucianellion maritimae Rivas Goday & Rivas-Mart. 1958 (PVF 25.0.1.0.3)
Groupement à Lomelosia rutifolia (= Pycnocomon rutifolium).
CORINE biotopes 16.223 (Dunes du Crucianellion maritimae), Natura 2000 (EUR 28) 2210 (Dunes fixées du littoral du Crucianellion maritimae). EUNIS B1.43 (Dunes grises fixées méditerranéo-atlantiques).
b. Helianthemetea guttati (Braun-Blanq. ex Rivas Goday 1958) Rivas Goday & Rivas Mart. 1963
Malcolmietalia ramosissimae Rivas Goday 1958
Laguro ovati-Vulpion fasciculatae Géhu & Biondi 1994
Groupement à Avena barbata, Corynephorus articulatus, Vulpia fasciculata et Cladanthus mixtus.
CORINE biotopes 16.228 (Groupements dunaires à Malcolmia, Malcolmietalia), Natura 2000 (EUR 28) 2230 (Dunes avec pelouses des Malcolmietalia), EUNIS B1.48 (Communautés de thérophytes des sables dunaires profonds du domaine téthyen).

Présence dans les végétations hygrophiles d’arrière-dune (tableau 2 ; planche 7)
A. corsicum se trouve le plus souvent dans des groupements hygrophiles, situés dans les zones basses en arrière des dunes et des cordons littoraux sableux. Ces formations sont généralement dominées par Elytrigia acuta et sont comprises entre une zone à Phragmites australis et/ou Rubus ulmifolius et la végétation plus xérophile des dunes ou des cordons.
En 2022 et 2023, la plus importante sous-population (comportant plusieurs milliers d’individus) a été observée sur un milieu humide, entre les embouchures du Ciavattone et du Golo (tableau 2, relevés 649 et 650, 1119 à 1126, 1128 à 1130, 1132, 1134, 1140). De même, plusieurs sous-populations de Moriani-Plage (tableau 2, relevé 654), de Stagnolu/Bravona (tableau 2, relevé 1078), de Gradugine (tableau 2, relevés 644 et 1093) et de Palo (tableau 2, relevé 635) se localisent aussi dans des milieux humides.
Sur ces sites, A. corsicum se trouve principalement dans le groupement à Elytrigia acuta, mais également dans des zones de transition composées de mosaïques entre des groupements à E. acuta et des pelouses dominées par Corynephorus articulatus, Avena barbata et Cladanthus mixtus et/ou des espèces du Crucianellion maritimae (à dominante de Centaurea sphaerocephala sur le site du Ciavattone/Golo ou de Lomelosia rutifolia ailleurs). En quelques points, l’exotique Panicum repens est abondant. C’est le cas notamment à Gradugine et ponctuellement sur les sites de Ciavattone-Golo et de Moriani Plage.
La syntaxonomie des groupements et la codification des habitats sont les suivantes :
Juncetea maritimi Braun-Blanq. in Braun-Blanq., Roussine & Nègre 1952
Juncetalia maritimi Géhu & Biondi ex Géhu 2004
Plantaginion crassifoliae Braun-Blanq., Roussine & Nègre 1952 (PVF 35.0.1.0.3)
Groupement à Elytrigia acuta
CORINE biotopes 15.51 (Prés salés méditerranéens à Juncus maritimus et Juncus acutus) / 15.53 (Prés méditerranéens halo-psammophiles), Natura 2000 (EUR 28) 1410 (Prés salés méditerranéens – Juncetalia maritimi), EUNIS B1.84 (Pelouses et landes des pannes dunaires) / E3.1 (Prairies humides hautes méditerranéennes).

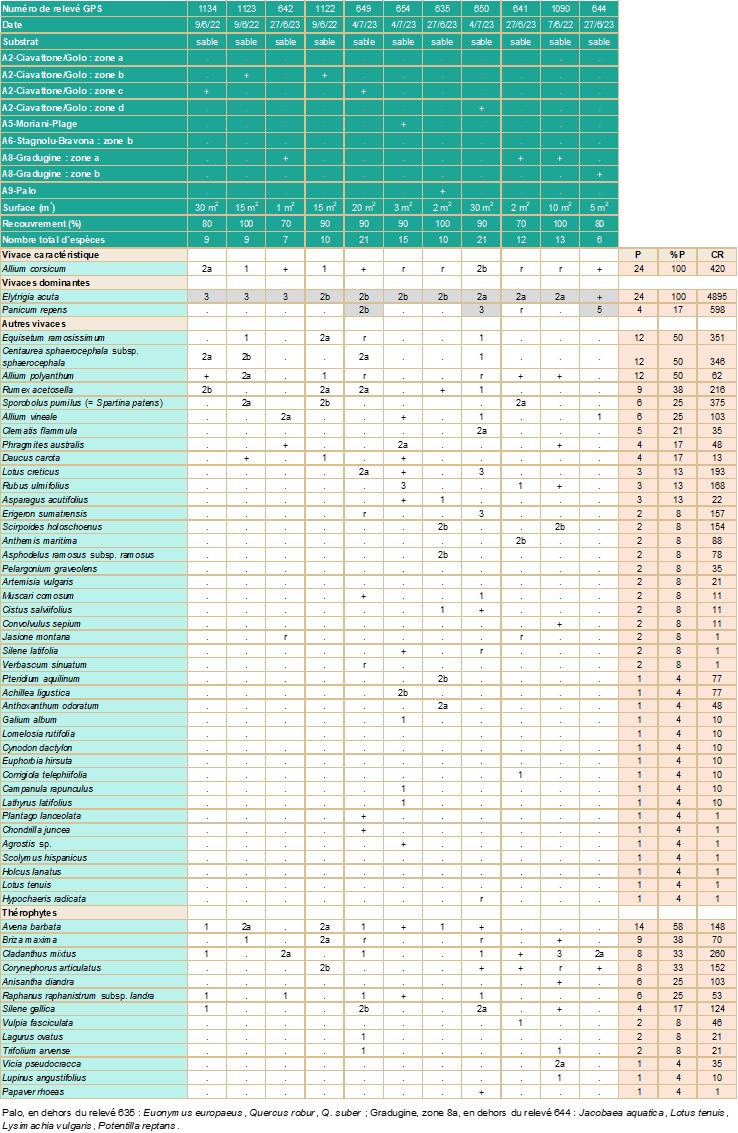
Présence dans les milieux perturbés (tableau 3 ; planche 8)
Allium corsicum est aussi présent dans des milieux perturbés, constitués de pelouses classables dans les Sisymbrietea officinalis (a) ou les Helianthemetea guttati (b), mais parfois difficiles à caractériser. C’est le cas :
- à San Pellegrino (pour partie), où il se trouve à 200 m de la mer, dans une pelouse xérophile à Anthoxanthum odoratum et Agrostis stolonifera dominants (tableau 3, relevé 652) ;
- au bord de la route conduisant au site de Stagnolu/Bravona, à Testaccia, à 1,5 km de la mer, où la végétation est composée de pelouses subnitrophiles dominées par Phalaris aquatica et/ou Avena barbata. Ces pelouses occupent des zones ouvertes dans les formations boisées à Quercus suber et sont aussi présentes en bord de route, en plusieurs endroits, variables d’une année sur l’autre (tableau 3, relevés 657, 657’, 1074, 1077, 1155’, 1187). La présence, par endroits, de Phalaris aquatica et de Schoenus nigricans laisse supposer un certain degré d’humidité du substrat ;
- à Palo, où corsicum occupe une dépression arrière-dunaire, colonisée par une mosaïque complexe, constituée de groupements à Vulpia fasciculata et Corynephorus articulatus dominants, de quelques rares Lomelosia rutifolia et d’autres espèces, parfois abondantes, telles Scirpoides holoschoenus, Pteridium aquilinum et Smilax aspera. Cette formation se situe entre le cordon littoral et la ripisylve (tableau 3, relevés 1162 et 1164) ;
- à Lugo-di-Nazza, où le taxon est présent le long d’un sentier rocailleux et probablement soumis au ruissellement et à l’érosion. Ce milieu peu stable est colonisé par une végétation claire (ayant 50 % de recouvrement), constituée surtout de thérophytes (Avena barbata, Sideritis romana romana, Brachypodium distachyon) (tableau 3, relevé 634) ;
- sur une terrasse du Tavignano, près de Corte, où corsicum est sur un substrat de sables et de galets, régulièrement renouvelé par les crues, au sein de pelouses à Avena barbata, Andryala integrifolia et d’espèces vivaces dont Cynodon dactylon dominant (tableau 3, relevé 618’).


La syntaxonomie des groupements et la codification des habitats sont les suivantes :
a. Sisymbrietea officinalis Gutte & Hilbig 1975
Brometalia rubenti-tectorum Rivas Mart. & Izco 1977
Echio plantaginei-Galactition tomentosae O. Bolòs & Molin. 1969 (PVF 66.0.1.0.1)
Groupement à Avena barbata
CORINE biotopes 87 (Terrains en friches et terrains vagues). EUNIS E1.61 (Communautés méditerranéennes à graminées subnitrophiles).
b. Helianthemetea guttati (Braun-Blanq. ex Rivas Goday 1958) Rivas Goday & Rivas Mart.1963
Helianthemetalia guttati Braun-Blanq. in Braun-Blanq., Molin. & Re. Wagner 1940
Helianthemion guttati Braun-Blanq. in Braun-Blanq., Molin. & Re. Wagner 1940 (PVF 32.0.1.0.2)
Anthoxanthetum ovati Gamisans & Paradis 1992
CORINE biotopes 35.21 (Pelouses siliceuses à annuelles naines) / 35.3 (Pelouses siliceuses méditerranéennes), EUNIS E1.811 (Pelouses siliceuses ouest-méditerranéennes).
2. Bilan stationnel d’Allium corsicum pour la Corse en 2022-2023
2.1. Méthodologie
Allium corsicum est une espèce dont les parties aériennes fleuries ne sont visibles que de la mi-mai à la fin juin environ. Par la suite, les inflorescences fanées sont plus difficiles à repérer. Aussi, la période de prospection optimale est courte. Notre étude sur le terrain a débuté le 30 mai 2022 sur le site de Stagnolu-Bravona et s’est achevée par une recherche tardive du taxon le long du Tavignano, à proximité de Corte, le 2 août 2022. Les prospections se sont poursuivies en 2023, entre le 15 mai et le 21 juillet. D’autres prospections ont également été effectuées durant la période automnale (septembre, octobre), pour compléter les observations et améliorer les connaissances sur la phénologie et la biologie du taxon.
En 2022 et 2023, toutes les stations anciennement signalées ont été recherchées. Quelques sites potentiellement favorables à la présence du taxon ont été également prospectés mais sans succès : cas de la rive nord du Golo-Foce di Tanghiccia et de la rive nord du Fiumorbo.
Localisation au GPS : chaque individu ou groupe d’individus observé a été pointé au GPS de façon à établir une cartographie précise de la répartition de l’espèce sur les sites.
Comptage des pieds : le nombre d’individus a été compté dans chaque sous-population, à l’exception de celle de Ciavattone-Golo. Pour ce site, aux endroits où la densité était trop importante (jusqu’à plusieurs dizaines individus au m2), un nombre moyen de pieds a été calculé en multipliant la surface de la zone de présence par la moyenne du nombre de pieds sur trois placettes de 1 m2.
Relevés phytosociologiques : la description de la végétation se base sur 45 relevés, réalisés suivant la méthode phytosociologique sigmatiste (Géhu & Rivas-Martínez, 1981). Les divers relevés sont groupés dans les tableaux 1 à 3. Les coefficients de recouvrement (CR) ont été calculés en suivant la pratique habituelle (Vanden Berghen, 1982 ; Géhu, 2006) (Note 3). Les espèces ont été déterminées avec les flores de Coste (1906), de Pignatti (1982), de Jeanmonod & Gamisans (2013) et de Tison & de Foucault (2014). La nomenclature suit TaxRef 16 (Gargominy et al., 2022).
Lors de l’étude sur le terrain, les caractères concernant l’écologie, la biologie et la phénologie du taxon ont également été notés. Les supports topographiques utilisés ont été les cartes au 1 : 25 000 (IGN, 2019) et les photos aériennes (IGN/CdC, 2019). Les aspects règlementaire et foncier des divers sites sont résumés dans le tableau 4.
Sept sites (soit 63,6%) sont classés en ZNIEFF : Banda Bianca, Ciavattone-Golo, San Pellegrino, Stagnolu/Bravona, Gradugine-Marina di Calzarellu, Palo et possiblement la Marina d’Aleria, au niveau de l’embouchure du Tavignano (car la localisation d’A. corsicum sur cette station est imprécise). Trois sites sont inscrits au réseau Natura 2000 : Biguglia, Ciavattone-Golo et possiblement la Marina d’Aleria, au niveau de l’embouchure du Tavignano (car la localisation d’A. corsicum sur cette station est imprécise). Deux sites appartiennent au Conservatoire du littoral : Banda Bianca et Gradugine-Marina di Calzarellu. Quatre sites (soit 45,5%) ne bénéficient d’aucun statut : Marina di Pinarello, Moriani-Plage, Lugo-di-Nazza et Tavignano, près de Corte.

2.2. Sites où Allium corsicum n’a plus été observé
Malgré une prospection attentive, Allium corsicum n’a pas été retrouvé sur trois sites.
Biguglia (planche 5 : A1)
Jauzein a signalé A. corsicum sur « Biguglia : plage près du déversoir de l’étang », en juin 1987 (Jauzein et al., 2002, cf. leur tableau 5 t). En fait, le déversoir de l’étang est situé sur la commune de Furiani et il n’est pas précisé, dans le texte, si la localisation se trouve en rive gauche ou en rive droite de l’étang. Ainsi, le point de ces auteurs a été placé de façon approximative dans la base de données (BDD) du Conservatoire botanique national de Corse. Ultérieurement, il n’y a eu aucune autre observation sur ce secteur.
Le 9 juin 2022, nous avons effectué, sans succès, une recherche en rive droite de l’embouchure de l’étang, au lieu-dit « Banda Bianca » (commune de Furiani). Ce site, classé en site Natura 2000 et en ZNIEFF, appartient depuis les années 2000 au Conservatoire du littoral (tableau 4). Il est aménagé et géré par les agents de la Collectivité de Corse. Le milieu, qui tend à se refermer en l’absence d’impacts, ne semble pas, dans cette partie du site, susceptible d’être favorable à A. corsicum.
À l’avenir, des prospections devraient être étendues en rive gauche de l’embouchure et, plus globalement, sur l’ensemble du lido de l’étang de Biguglia, voire sur tout le pourtour de l’étang, dans les zones présentant Elytrigia acuta.
Marina di Pinarello (planche 5 : A3)
Jauzein et al. (2002) signalent que la plante a été également observée par Dutartre à Anghione, Marina di Pinarello, en 1981. Il n’y a pas, pour cette station, de localisation précise.
Le 2 juin 2022, nous avons prospecté, sans succès, une zone de 1 km de long, située de part et d’autre de la route d’accès à la Marina di Pinarello. Dans sa partie sud, la portion inventoriée est soumise à de nombreux impacts (urbanisation, aménagements divers, fréquentation). Par contre, sa partie nord est plus préservée et abrite, d’ailleurs, d’autres taxons relativement rares comme Thesium humile et Phleum arenarium… Certaines parties du site pourraient être favorables à A. corsicum, qu’il conviendra de rechercher à nouveau.
Marina d’Aleria (planche 5 : A7)
Allium corsicum a été observé par Deschâtres, le 11 juin 1981 (Jauzein et al., 2002 ; BDD du CBNC), sur la plage de la Marina d’Aleria-Padulone (commune d’Aleria), en rive gauche du Tavignano et au nord de la route d’accès à la mer.
Malgré deux passages sur le site (les 7 et 29 juin 2022) et une prospection sur 1,5 km environ au nord de l’embouchure du Tavignano, aucun individu n’a été observé. Des zones potentiellement favorables au taxon semblent présentes sur le site et devront faire l’objet de nouvelles prospections. Une partie du site est classé en Natura 2000 et en ZNIEFF (tableau 4).
2.3. Sites où Allium corsicum a été observé en 2022 et 2023
Ciavattone-Golo (planche 5 : A2 ; planche 9)
La première observation d’A. corsicum en Corse a été faite le 10 juin 1980 (cf. leur tableau 6), sur la commune de Venzolasca, par Deschâtres (Jauzein et al., 2002), qui signale le taxon sur la « plage, près du village de Cap Sud ». La localisation est peu précise et le point a donc été placé de façon approximative dans la BDD du CBNC. Le 31 mai 2016, Delage (Delage et al., 2018), observe « quelques dizaines de plantes fleuries à l’embouchure du Golo, à l’Est de Terragliolo, au bord de la piste allant à la plage, à environ 200 m de la mer » (planche 9 : secteur « a »).
Les 9 et 27 juin 2022, de nouvelles prospections ont été faites entre Mucchiatana et l’embouchure du Golo. Aucun individu n’a été observé entre la rive sud de l’embouchure du Ciavattone et Mucchiatana. Mais ces nouvelles prospections ont permis d’étendre l’aire de présence du taxon à une grande partie des parcelles situées entre l’embouchure du Golo au nord, l’embouchure du Ciavattone au sud et Terragliolo à l’ouest, sur près d’un hectare. Delage est également passé sur ce secteur le 31/06/2016, mais n’a pas observé le taxon à cette date. Deux passages sur le site en 2023 (16 juin et 4 juillet) nous ont permis de revoir le taxon.
Cette station fait partie d’une ZNIEFF et du site Natura 2000 « FR 400572 – Tanghiccia – embouchure du Golo – Ciavattone – Mucchiatana » (tableau 4).
Le secteur a, observé pour la première fois par Delage en 2016 (Delage et al., 2018, tableau 6), se trouve sur la rive droite du Golo, au bord de la piste menant à la mer. Sa superficie est d’environ 50 m2. 55 individus fleuris ont été dénombrés le 9 juin 2022 et 15 individus le 16 juin 2023. La zone à A. corsicum est située à la lisière de la ceinture à Phragmites australis et Rubus ulmifolius qui borde la rive droite du Golo, dans sa partie terminale. Elle se trouve sur du sable, en légère dépression. Sa végétation est dominée par Elytrigia acuta dont le recouvrement est important (tableau 2, relevés 1119 à 1121). Un taxon des dunes fixées, Centaurea sphaerocephala subsp. sphaerocephala, est également abondant par place. La topographie de la zone bordant la piste a été légèrement rehaussée lors de travaux de nivèlement. Ainsi, du côté de la piste, de nombreuses espèces rudérales sont également présentes, ainsi qu’une espèce ornementale, le géranium rosat (Pelargonium graveolens), qui occupe l’extrémité est de la zone et qui est en contact avec A. corsicum, ce qui réduit les possibilités d’extension de celui-ci.
Les secteurs b et c à A. corsicum n’avaient pas été identifiés avant nos prospections de 2022. Ils occupent une parcelle agricole étendue juste à l’ouest d’une piste menant à un hangar, entre l’embouchure du Golo au nord et l’embouchure du Ciavattone au sud. Près de 4 500 individus fleuris ont été dénombrés le 9 juin 2022, sur une superficie d’un peu moins d’un hectare. Il s’agit, en l’état des connaissances, de la plus importante sous-population (en nombre d’individus et en superficie) de cette endémique. Néanmoins, les effectifs semblent fluctuants puisque en 2023, la zone « c » ne comptait que peu d’individus. La zone à A. corsicum est limitée à l’ouest par une ceinture à Phragmites australis et Rubus ulmifolius et s’étend sur des terrains sableux, plus ou moins en dépression. Au niveau des parties les plus hautes s’installent des cistaies à Cistus salviifolius, tandis que les dépressions sont colonisées (1) par des formations à Elytrigia acuta dominant pour les plus humides et (2), pour les autres, par des mosaïques entre un groupement à Elytrigia acuta, un groupement des dunes fixées à Centaurea sphaerocephala subsp. sphaerocephala dominant et un groupement thérophytique des Malcolmietalia.
Le secteur d se trouve du côté est de la piste. Il a été incendié durant l’hiver 2021-2022 (planche 16 : C et D) et aucun pied n’a été observé au printemps 2022. Par contre, en 2023, le taxon y était très abondant (plus d’un millier d’individus). Ce secteur, occupé avant l’incendie par une cistaie à Cistus monspeliensis, est aujourd’hui en cours de recolonisation par des groupements à Elytrigia acuta et à Panicum repens dominant (tableau 2, relevé 650) ou par des groupements rudéraux à Lotus creticus et Agrostis stolonifera dominants et à thérophytes dont Raphanus raphanistrum subsp. landra (tableau 2, relevé 651). Durant l’hiver 2022-2023, une partie de la cistaie située du côté ouest de la piste et comprise entre les secteurs « b » et « c » a également été incendiée sur 200 à 300 m2 (planche 16 : E). Ces brûlages récurrents semblent plutôt favorables au taxon. En effet, chaque année, ils ne touchent qu’une faible surface et ne concernent que les végétations fermées, telles les cistaies, dont le recouvrement est néfaste à A. corsicum, taxon héliophile. Donc ici, en dehors d’une éventuelle fermeture du milieu, aucune menace ne semble peser aujourd’hui sur l’espèce et le mode de gestion par des incendies de faible extension et de faible intensité semble approprié.

San Pellegrino (planche 5 : A4 ; planche 10)
Delage a découvert le taxon sur la commune de Penta-di-Casinca, à environ 560 m au N de la plage de San Pellegrino, dans une pelouse en bordure d’une dépression humide (BDD du CBNC). La station, incluse dans une ZNIEFF (tableau 4), a été très précisément localisée et comptait, le 14/06/2018, une cinquantaine d’individus. Des recherches élargies autour de la station ont été entreprises le 2 juin 2022. Elles n’ont donné aucun résultat. En revanche, l’espèce a été revue en 2023, mais la station ne comptait que deux individus, situés au sein d’une pelouse plus ou moins humide à Anthoxanthum odoratum et Agrostis stolonifera (tableau 3, relevé 652).

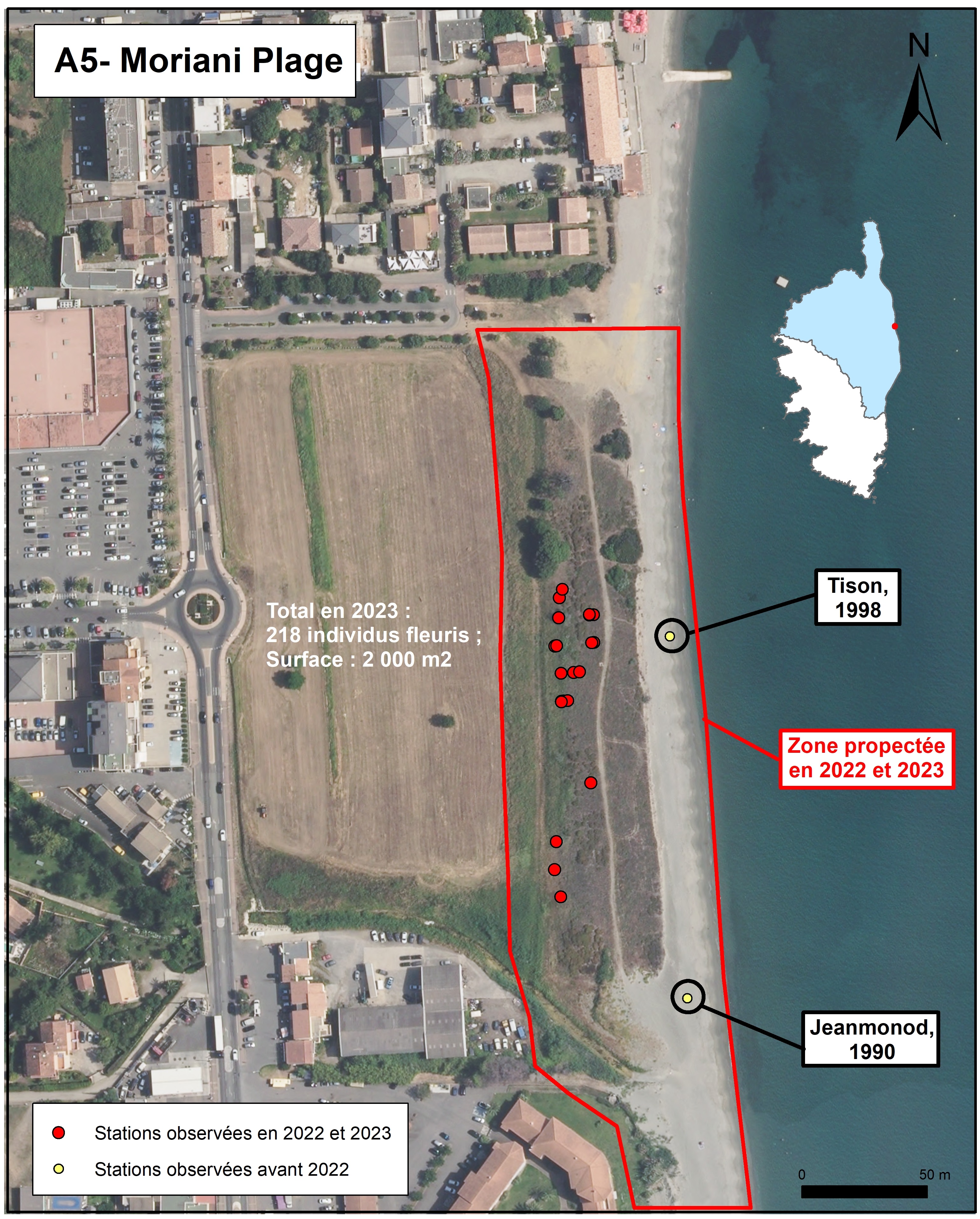
Moriani-Plage (planche 5 : A5 ; planche 11)
Sur le cordon dunaire de Moriani-Plage (commune de San Nicolao), Allium corsicum a d’abord été signalé par Jeanmonod & Thiébaud le 14 juin 1990. Ensuite il a été observé par plusieurs auteurs (Jauzein et al., 2002 ; BDD-CBNC). Tison le revoit ici en 1998. Dans leur publication, Jauzein et al. (2002) signalent que les densités peuvent être importantes (10 pieds par m2). Delage (comm. pers.) indique avoir vu un individu au sud de Padulella, en bordure de la route RT 10 (ex-RN 198), dans une plate-bande, au niveau d’une zone urbanisée.
Un passage sur le site le 26/10/2022 a permis l’observation, sur le sommet de la dune, d’un individu avec un reste de hampe florale et, au pied de ce dernier, d’une douzaine de jeunes plants avec leurs feuilles. Début 2023, plus de deux cents nouveaux individus avec leurs feuilles étaient également présents sur le revers de la dune et le long d’un sentier situé à l’arrière du bourrelet dunaire. Mais, en 2023, seuls dix pieds fleuris ont été observés. Les formations dans lesquelles se trouve A. corsicum sont :
- des groupements denses à Lomelosia rutifolia sur le sommet et sur le revers de la dune, avec comme autres espèces dominantes Panicum repens et Lotus creticus (tableau 2, relevés 653 et 655) ;
- des groupements à Elytrigia acuta et Rubus ulmifolius dominants, le long d’un sentier créé entre l’arrière-dune et un fossé bordant une vaste prairie régulièrement fauchée (tableau 2, relevé 654).
Une menace pour le taxon pourrait être la fermeture du milieu, notamment au niveau du sentier situé à l’arrière du bourrelet dunaire qui, en août 2023, commençait à être envahi par les ronces. Mais, ce dernier semble, pour l’heure, régulièrement ouvert au début du printemps (planche 16 : A et B).
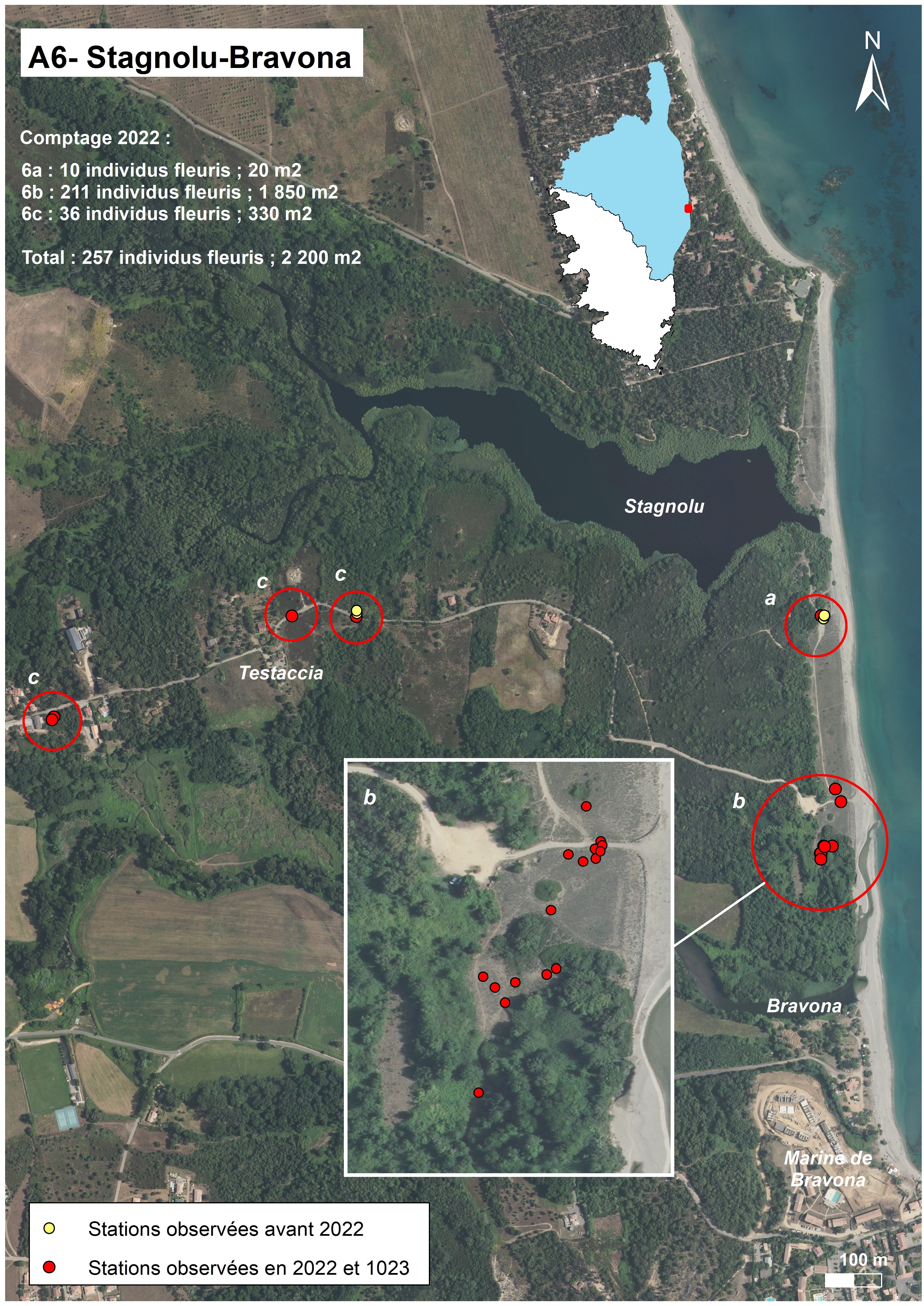
Stagnolu-Bravona (planche 5 : A6 ; planche 12)
Sur le site de Stagnolu-Bravona (commune de Linguizzetta), qui est inclus dans une ZNIEFF (tableau 4), A. corsicum a été découvert par Delage en 2010 puis revu en 2015 :
- observations du 6 juin 2010 : « Linguizzetta : NW de la Marine de Bravona, route d’accès à la plage au sud de l’étang de Stagnolu, à environ 1 km de la plage, un peu à l’est du lieu-dit Testaccia, pelouse en bord de route, en lisière de cistaie, une vingtaine de plantes fleuries » (Delage et al., 2018) (« c ») ;
- observations du 6 juin 2015 : « Linguizzetta : nord de la Marine de Bravona, au sud de la lagune de Stagnolu, dépression arrière-dunaire à environ 150 m au sud de l’embouchure de l’étang, quelques dizaines de plantes fleuries » (Delage et al., 2018) (« a »).
Les localisations, pointées au GPS, sont très précises.
En 2022, nos recherches ont porté sur les portions où le taxon avait déjà été observé (« a » et « c ») mais aussi sur de plus vastes surfaces, ce qui a permis de trouver de nouvelles sous-populations. Sur ce site, A. corsicum a une répartition beaucoup plus étendue que ce qui était connu : ainsi, il est présent au nord de la rive gauche de la Bravona (« b ») et à proximité de la route d’accès à la mer (« c »).
Le secteur a se trouve au sud de la rive droite de l’embouchure temporaire de l’étang de Stagnolu, autour d’une ancienne zone de stationnement aménagée sur la dune fixée. Seuls dix individus y ont été comptés en 2022, en deux points distants de quelques mètres l’un de l’autre, sur une superficie de moins de 20 m2. Les relevés 1027, 1075 (tableau 1) et 1074 (tableau 3) montrent une nette dominance de thérophytes de pelouses xérophiles d’arrière-dune, composées principalement de graminées (Corynephorus articulatus, Vulpia fasciculata, Avena barbata, Anisantha diandra…), de Cladanthus mixtus et/ou de Lomelosia rutifolia. Ce secteur est aujourd’hui fermé à la circulation motorisée.
Le secteur b se trouve au nord de la rive gauche de l’embouchure de la Bravona. Il a également été mis en protection par des ganivelles et des blocs rocheux. En 2022, 211 individus ont été observés aux endroits suivants : sur la dune fixée, au bord de la piste et dans les prairies plus ou moins humides situées à proximité de la ripisylve. Au total, la superficie occupée par A. corsicum est de près de 2 000 m2. En ce qui concerne la végétation, A. corsicum se trouve (1) dans les pelouses xérophiles d’arrière-dune à thérophytes, composées principalement de graminées (Corynephorus articulatus, Anisantha diandra, Avena barbata, …) et de Cladanthus mixtus (tableau 3, relevés 657, 1077 et 1187), (2) dans le groupement à Lomelosia rutifolia (tableau 1, relevés 656, 1076 et 1182), (3) dans un groupement à Elytrigia acuta (tableau 2, relevé 1078). Le taxon est le plus abondant dans les pelouses plus ou moins humides situées à l’arrière de la dune, à proximité de la ripisylve de la Bravona.
Le secteur c, discontinu, se trouve dans des prairies xérophiles, au bord de la route conduisant à la mer, près du lieu-dit Testaccia. 36 individus ont été comptés sur une superficie de près de 330 m2. Les relevés 657’ et 1155-1 (tableau 3) montrent une nette dominante de thérophytes de pelouses xérophiles composées principalement de graminées (Anisantha diandra, Avena barbata…) et d’une autre graminée pérenne (Phalaris aquatica). Néanmoins, la présence de Schoenus nigricans semble indiquer une certaine humidité du substrat par endroits.
Globalement, sur les trois secteurs de présence du taxon, en dehors d’une éventuelle fermeture du milieu, aucune menace ne semble peser aujourd’hui sur l’espèce. Pour rappel, le site a été aménagé par la commune (mise en défens des dunes, canalisation du public et des véhicules).
Gradugine-Marina de Calzarellu (planche 5 : A8 ; planche 13 : a et b)
Le cordon littoral face à l’étang de Gradugine appartient au Cdl et est une ZNIEFF (tableau 4). Des bulbes d’Allium corsicum ont été observés et récoltés par Tison en juin 1997 (Jauzein et al., 2002). Ces bulbes, mis en culture, ont donné des individus, dont un, en juin 1999, a été choisi comme holotype (Jauzein et al., 2002).
En juin 1999, Paradis et Piazza (in Paradis et al., 2002) ont aussi observé l’ail de Corse sur le cordon de Gradugine.
En 2017, Delage a pointé, au GPS, 2 sous-populations distantes l’une de l’autre d’environ 450 m : une au nord-est avec moins de 10 individus et une au sud-ouest avec plusieurs dizaines d’individus.
En juin 2022, deux petites sous-populations ont été observées sur ce cordon, mais avec assez peu d’individus : 4, pour la station « a » et 15 pour la station « b ». La superficie occupée par le taxon est un peu supérieure à 500 m2. En 2023, seuls 10 individus fleuris ont été comptés à la fin du mois de juin. Mais des passages en fin d’automne et début d’hiver ont permis d’observer de très nombreux individus en feuilles.
Ces deux sous-populations se trouvent dans des zones de transition, entre la roselière à Phragmites australis et le bas du cordon sableux. Ces zones sont colonisées par une mosaïque entre une pelouse d’arrière-dune (à Cladanthus mixtus, Vulpia fasciculata, Corynephorus articulatus…), une formation à Scirpoides holoschoenus et des repousses, après incendie, de Panicum repens (tableau 3, relevé 1097 et tableau 2, relevés 641, 642, 1090 et 1093).
D’un point de vue géographique, les secteurs potentiellement favorables à A. corsicum se trouvent dans la partie centrale du cordon de Gradugine, qui est relativement épargnée par les impacts anthropiques (surféquentation, nettoyage de plage…), qui sont très importants au nord-est, à Calzarellu. Par contre, ces secteurs sont régulièrement parcourus par des incendies destinés à limiter l’extension des roselières. Si leur fréquence et leur intensité ne sont pas trop importantes, ces feux, en réduisant la concurrence végétale, peuvent être bénéfiques au taxon.
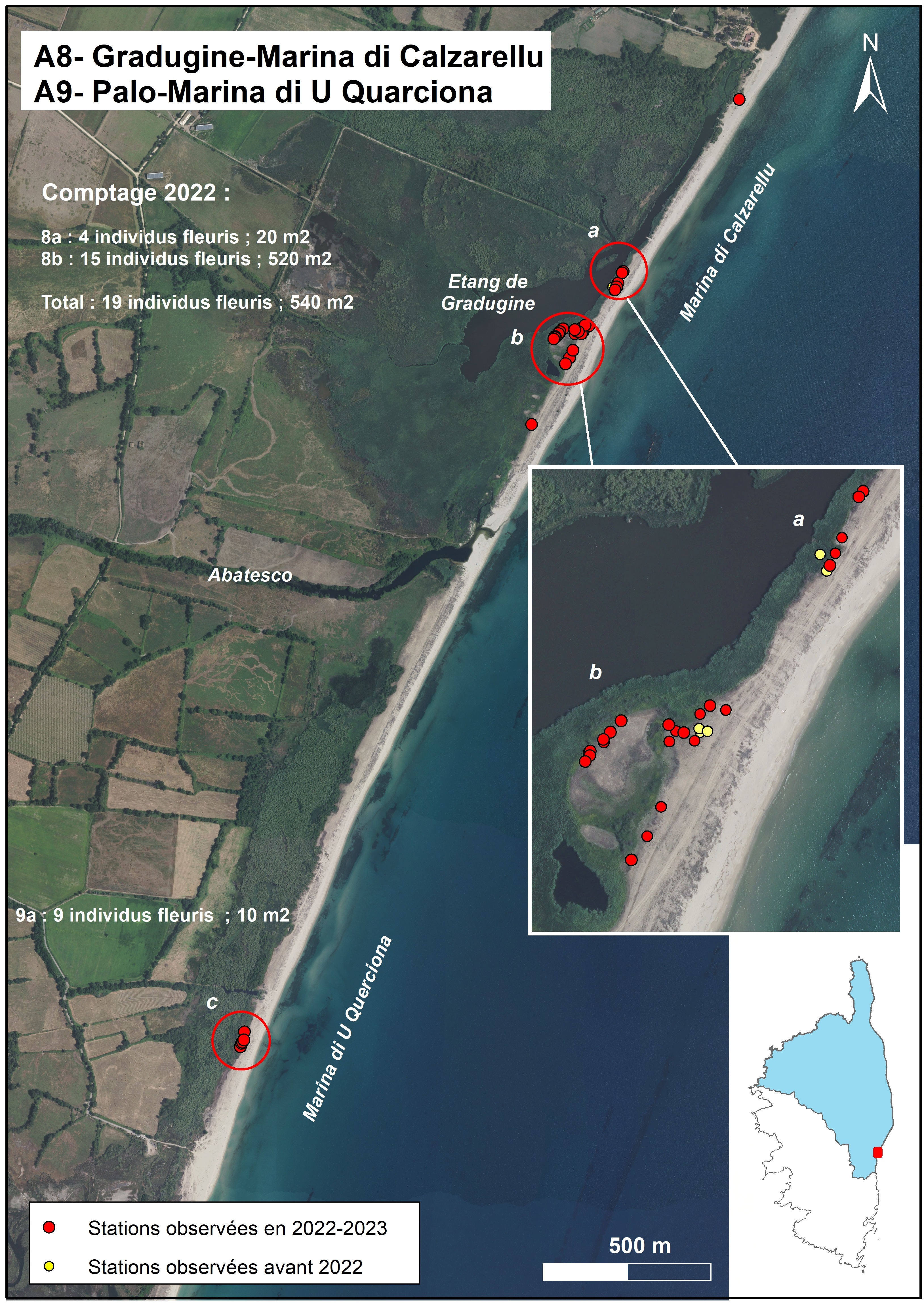
Palo-Marina di U Quarciona (planche 5 : A9 ; planche 13 : c)
Jusqu’à présent A. corsicum n’avait pas été signalé sur le cordon littoral de la Marina di u Querciona, qui est compris entre les embouchures de l’Abatescu et l’embouchure de l’étang de Palo. En juin 2022, la prospection de ce cordon a permis de découvrir une dizaine d’individus à environ 350 m au nord de l’entrée du site (face à l’extrémité est de la RD 745).
Cette petite sous-population se localise sur la partie arrière du cordon dunaire, dans une zone en légère dépression, située à l’est de la ripisylve peuplant une très ancienne lagune.
A. corsicum se trouve dans une végétation composée d’une mosaïqueentre une pelouse d’arrière-dune à Corynephorus articulatus, Vulpia fasciculata, Anthoxanthum odoratum… et un groupement à Elytrigia acuta, Scirpoides holoschoenus et/ou Pteridium aquilinum (tableau 3, relevés 636, 1132 et 1163 et tableau 2, relevé 635).
Cette station d’A. corsicum fait partie d’une ZNIEFF et est en bordure de terrains appartenant au Cdl (tableau 4). Elle ne semble pas subir d’impact particulier.
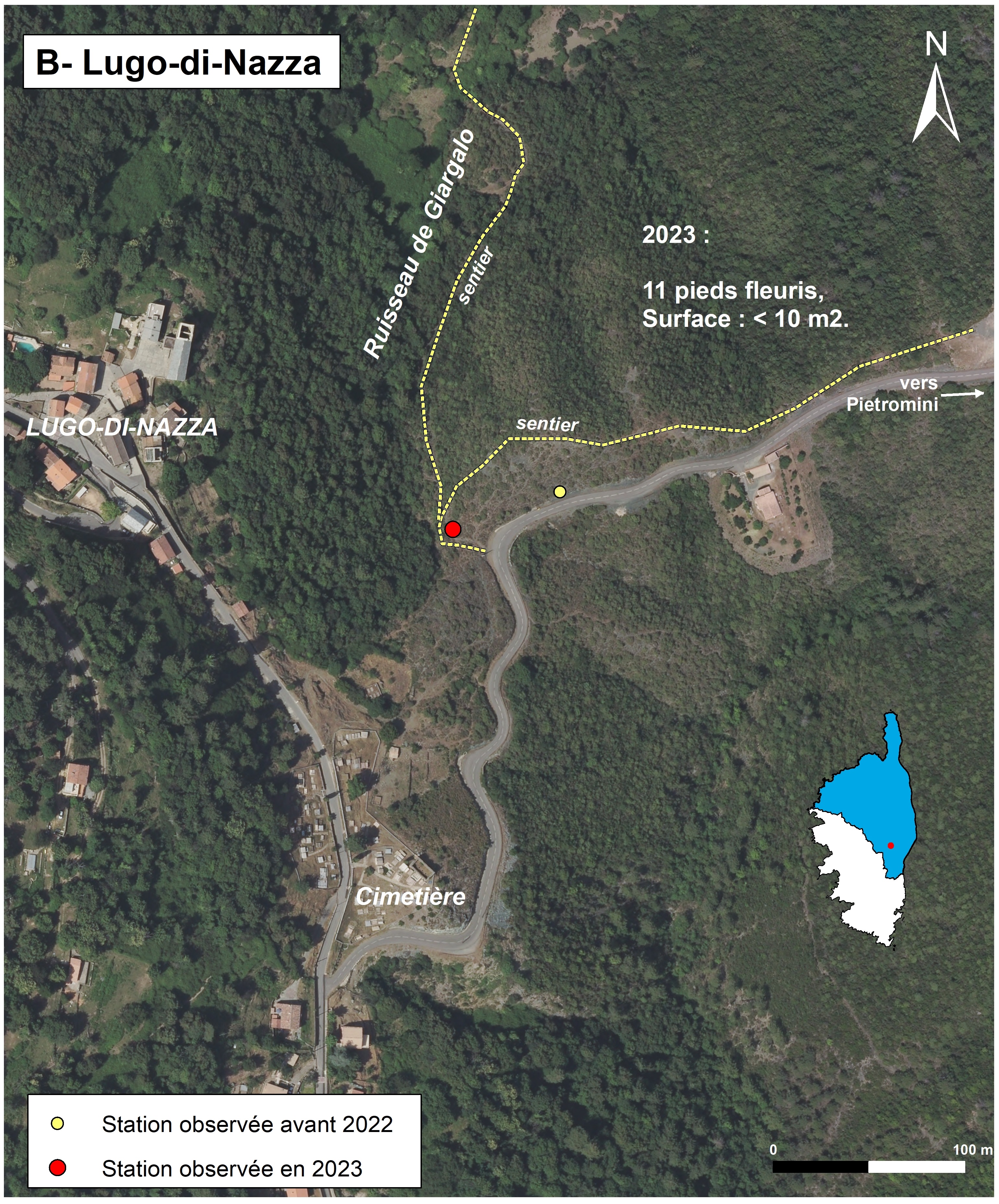
Lugo-di-Nazza (planche 5 : B ; planche 14)
Deschâtres a signalé le taxon à Lugo-di-Nazza sur des rocailles et serpentines, dans un secteur relativement éloigné de l’influence de la mer (Delage et al., 2018). La localisation exacte n’a pas été précisée. Dans la BDD du CBNC, deux points ont été localisés, de façon approximative, au nord et au sud du village de Lugo-di-Nazza.
Des recherches ont été entreprises dans le secteur nord, en juin 2022, sans résultats. Mais le 26 juin 2023, la prospection du secteur sud a permis de retrouver la station, au départ (du côté de Lugo-di-Nazza) du chemin qui longe la rive droite du ruisseau de Giargolo et qui fait une boucle entre Lugo-di-Nazza et Pietromini. La sous-population, qui ne compte qu’une dizaine d’individus sur quelques mètres linéaires, se trouve sur et au bord d’un sentier. Le substrat meuble est très peu épais et de petits rochers affleurent çà et là. La végétation est claire (50% de recouvrement) et constituée de thérophytes, telles Brachypodium distachyon (2a), Sideritis romana subsp. romana (2a), Vulpia ciliata (1) et Avena barbata (1). En dehors du sentier, la végétation est constituée de chaméphytes et de phanérophytes, avec comme espèces dominantes Juniperus oxycedrus subsp. oxycedrus, Cistus monspeliensis, Euphorbia spinosa, Genista corsica et Teucrium marum (tableau 3, relevé 634). Cette station ne bénéficie d’aucun statut de protection. Les menaces semblent faibles, car le site est relativement isolé. Le piétinement lié à la fréquentation du sentier et le ravinement ne semblent pas mettre en danger la station. Néanmoins, la sous-population ne compte qu’un faible nombre d’individus et sera donc à surveiller.
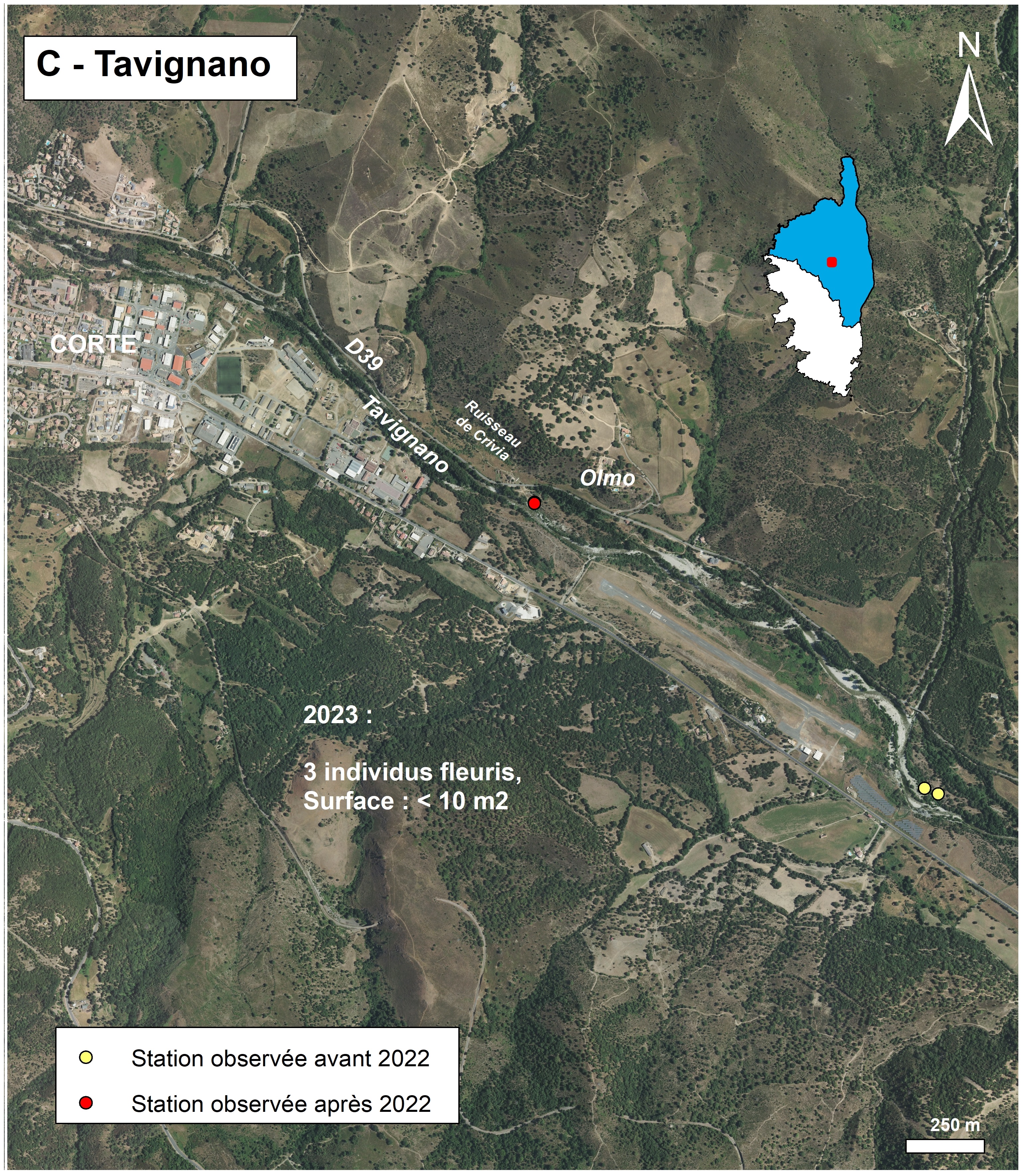
Rive gauche du Tavignano, près de Corte (planche 5 : C ; planche 15)
Deschâtres a signalé le taxon à Corte, en rive gauche du Tavignano, à 3 km en aval de la ville (Delage et al., 2018). Là encore, le taxon est très éloigné de l’influence de la mer. Il se trouve, à proximité du fleuve, sur sables et galets. La localisation exacte n’a pas été précisée. Dans la BDD-CBNC, le point a été localisé, de façon approximative, au lieu-dit Alzeta, à environ 800 m à l’ouest du pont de Papineschi (situé sur la RD 214), à quelques dizaines de mètres du champ photovoltaïque (situé lui en rive droite du Tavignano) et à l’extrémité sud-est de la piste de l’aérodrome de Corte.
Des recherches tardives ont été entreprises le 2 août 2022 sur les bancs de sables et de galets situés aux alentours du point de la BDD-CBN, sans résultats. Le 27 juin 2023, de nouvelles recherches menées plus en amont ont permis de trouver trois individus sur substrat sableux, à environ 15 m du Tavignano et 20 m de la RD 39, près du lieu-dit Olmo. Le cortège floristique correspond à des pelouses à Avena barbata, Chondrilla juncea et Cynodon dactylon dominants. Le recouvrement est d’environ 80% (tableau 3, relevé 618’). Ce site ne possède aucun statut de protection. Bien que la station se trouve à proximité d’un sentier, elle ne semble pas menacée par les activités humaines. En revanche, la présence d’espèces exotiques envahissantes (Ailanthus altissima et Cercis siliquastrum) sera à surveiller.
3. Menaces et actions réalisées
3.1. Menaces (tableau 5 ; planche 16)
Les effectifs d’A. corsicum sont faibles (environ 5 000 individus) et les surfaces occupées sont réduites à un peu plus d’un hectare (tableau 5). Néanmoins, peu de menaces potentielles ou actives ont été constatées sur les sites. La succession naturelle de la végétation risquant de provoquer la fermeture du milieu semble constituer un frein à l’apparition des scapes florifères, sans que cela ne soit forcément néfaste au taxon qui peut survivre sous forme de bulbes et bulbilles dans le sol, du moins pendant un certain temps, grâce à leurs réserves.
Cette évolution de la végétation concerne les sous-populations de Ciavattone-Golo, du cordon de Gradugine, du hameau de Testaccia (Linguizzetta), de Lugo-di-Nazza et de la bordure du Tavignano près de Corte.

Ciavattone-Golo
Cette sous-population, la plus importante actuellement connue, est située dans des pelouses plus ou moins humides, aujourd’hui colonisées, pour partie, par Cistus salviifolius et Rubus ulmifolius, deux espèces ligneuses pouvant les recouvrir. Néanmoins, cette crainte est à nuancer car :
- il s’agit de parcelles agricoles qui sont probablement pâturées ;
- la microtopographie des parcelles, et notamment l’existence de dépressions plus humides, ne permet pas, partout, l’installation des cistaies ;
- les terrains, situés entre les deux cours d’eau (Ciavattone et Golo), sont périodiquement « rajeunis » par les crues, ce qui remanie le substrat par l’apport ou le retrait de sédiments et éclaircit la végétation ;
- enfin, en 2022, l’extrémité orientale de la parcelle a été incendiée et, en 2023, une autre parcelle l’a également été (planche 16 : C à E). Par ailleurs, des traces d’incendies plus anciens sont encore visibles du côté sud, ce qui, là aussi, limite la concurrence avec les ligneux, mais peut, en revanche, avoir un impact négatif sur les cailleux présents dans les zones superficielles du sol.
Hameau de Testaccia (Linguizzetta)
Les pelouses présentant A. corsicum, localisées dans les clairières au sein des boisements de Quercus suber, sont susceptibles, en l’absence d’entretien, de se refermer sur le long terme. Il en va de même pour les sous-populations situées en bord de route. Des travaux d’entretien des bords de route, réalisés avant la mi-mai, pourraient favoriser le taxon.
Cordon de Gradugine
Un incendie des roselières à la fin de 2021 ou au début de 2022 a provoqué l’ouverture du milieu sur la rive est de l’étang de Gradugine, ce qui pourrait être favorable à l’expansion d’A. corsicum (planche 16 : F).
Lugo-di-Nazza et bords du Tavignano
Les très petites sous-populations isolées de Lugo-di-Nazza et des bords du Tavignano près de Corte seront à surveiller avec attention, du fait de leur très faible nombre d’individus.
3.2. Actions réalisées
Deux stations littorales appartiennent au Conservatoire du littoral (Biguglia-Banda Bianca et cordon de Gradugine). Ces sites sont aménagés et gérés. La station de Palo est en limite des terrains du Conservatoire et est assez peu soumise aux impacts.
Après la découverte de la grande extension de la station de Ciavattone-Golo en 2022, un signalement a été fait, par le CBNC, à l’animateur du site Natura 2000 « FR 9400572 Tanghiccia, embouchure du Golo, Ciavattone, Mucchiatana ». Ainsi, le taxon sera pris en compte et pourra faire l’objet, si besoin, de fiches « action » dans le document d’objectifs du site. Par ailleurs, ce site est couvert par un arrêté préfectoral de protection de biotope depuis le 17 mars 2017 (arrêté : DDTM2B/SEBF/BIODIVERSITE/N°208/2017), qui règlemente l’usage des lieux.
Le cordon littoral compris entre l’étang de Stagnolu et l’embouchure de la Bravona a fait l’objet, suite à une étude réalisée par le CBNC à la demande de la commune en 2012, d’aménagements (pose de ganivelles et de blocs rocheux, panneaux d’information) pour protéger la dune et empêcher la circulation de véhicules sur le site.
Enfin, le site de Moriani-Plage est géré. Un platelage nord-sud a été installé sur le sommet de la dune pour la protéger du piétinement et le sentier situé à l’arrière du bourrelet dunaire semble, pour l’instant, régulièrement ouvert au début du printemps, ce qui limite le développement des ronces et donc la fermeture du milieu (planche 16 : A et B).
Conclusions (planche 17)
Ce travail a été l’occasion d’étudier plus en détail les populations d’Allium corsicum. Il a permis d’affiner la caractérisation des habitats et de faire des comptages sur l’ensemble des sous-populations observées. En 2022-2023, la population a été estimée à environ 5 000 individus, occupant une surface de près de 1,4 ha.
Les prospections de 2022-2023 ont également permis : (1) d’étendre, de façon importante, le périmètre de la station de l’embouchure du Golo et de la station de Stagnolu-Bravona, (2) de découvrir une petite population au nord de l’étang de Palo (commune de Serra-di-Fiumorbo) et (3) de retrouver les localisations anciennement découvertes par Deschâtres à Lugo-di-Nazza et en bordure du Tavignano, qui n’avaient plus été vues depuis les années 1980 pour la première et les années 1990 pour la seconde. Par contre, malgré les recherches effectuées, les stations de Biguglia, Marina di Pinarello et Marina d’Aleria-Padulone n’ont pas été revues.
Dans la BDD du CBNC, le taxon est signalé dans onze mailles de 5 km x 5 km, entre les années 1980 et 2021 (planche 17 : A). Entre 2000 et 2021, intervalle de temps retenu dans le cadre du travail du CBNC sur la stratégie de conservation de la flore de Corse, on ne dénombrait que quatre mailles de présence (planche 17 : B), ce qui a contribué au classement du taxon dans la catégorie d’enjeu de conservation « très fort ». Le travail réalisé en 2022-2023 permet de situer le taxon dans sept mailles (planche 17 : C). En conséquence, son score passe de 15 à 14, ce qui entraîne sa rétrogradation de la première à la quatrième place, dans la hiérarchisation des taxons. Mais il reste classé dans la catégorie des taxons à enjeu « très fort ».
Cette étude ne constitue qu’un premier travail, qui devra être poursuivi sur plusieurs années pour permettre d’améliorer les connaissances sur la biologie et l’écologie du taxon, sur la dynamique de ses sous-populations et sur les menaces. De plus, les stations retrouvées dans l’intérieur de l’île (à Lugo-di-Nazza et au bord du Tavignano à Corte) montrent que l’aire de distribution de l’espèce n’est pas limitée au littoral et est plus étendue que ce qui était supposé. L’effort de prospection devra être renforcé pour rechercher de nouvelles stations.
À la suite de ce bilan stationnel et dans la mesure où aucune menace importante ne semble peser actuellement sur les sous-populations retrouvées en 2022-2023, il n’est pas préconisé, pour l’heure, de mettre en œuvre un plan d’action. Nous préconisons cependant (1) un suivi sur plusieurs années pour préciser la chorologie du taxon, étudier la dynamique de ses sous-populations et améliorer la caractérisation de ses habitats, (2) une poursuite de la recherche des stations non revues en 2022-2023.
Enfin on pourrait envisager l’inscription d’Allium corsicum sur la liste des espèces règlementées.

Note 1. Le critère D2 est utilisé uniquement pour la catégorie VU (espèce vulnérable). Il correspond à une « zone d’occupation restreinte ou à un nombre de localités limité et susceptibles d’être affectées à l’avenir par une menace vraisemblable pouvant très vite conduire le taxon vers les catégories EX (espèce disparue) ou CR (espèce en danger critique d’extinction) » (UICN France, FCBN, AFB & MNHN, 2018).
Note 2. Bilan stationnel (site inventory) d’un taxon, des taxons ou d’une communauté végétale : méthode d’inventaires spécifiques de terrain pour toutes les stations avérées et historiques, ou pour un nombre représentatif de stations (si n > 20), avec relevé des informations précises sur l’état des populations ou des communautés végétales (Le Berre et al., 2019). Cet état des lieux sert également à évaluer l’état de conservation d’une espèce ou d’une communauté végétale (RESEDA-FLORE, 2019).
Note 3. Les coefficients de recouvrement (CR) ont été légèrement modifiés pour les coefficients 2a et 2b, c’est-à-dire en donnant les valeurs suivantes (quantités moyennes) aux coefficients d’abondance-dominance : 5 = 87,5 ; 4 = 62,5 ; 3 = 37,5 ; 2b = 18, 5 ; 2a = 8,5 ; 1 = 2, 5 ; + = 0,2 ; r = 0,1. Ainsi, dans chaque relevé, la quantité moyenne des coefficients d’abondance-dominance a été additionnée, pour chacun des taxons, et la somme obtenue a été divisée par le nombre total des relevés du tableau, puis multipliée par 100 pour obtenir le CR.
Annexe 1
Les espèces ne figurant que dans un seul relevé ont été retirées du tableau 2. Elles sont listées ci-dessous.
relevé 635 : 1/ vivaces : Anthoxanthum odoratum (2a), Pteridium aquilinum (2b), en dehors du relevé : Euonymus europaeus, Quercus robur, Q. suber ; relevé 641 : 1/ vivace : Corrigiola telephiifolia (1) ; en dehors du relevé 644 : Jacobaea aquatica, Lotus tenuis, Lysimachia vulgaris, Potentilla reptans ; relevé 649 : 1/ vivaces : Chondrilla juncea (+), Plantago lanceolata (+) ; relevé 650 : 1/ vivace : Hypochaeris radicata (r), 2/ thérophyte : Papaver rhoeas (+) ; relevé 654 :1/ vivaces : Achillea ligustica (2a), Agrostis sp. (+), Campanula rapunculus (1), 2/ Galium album (1), Lathyrus latifolius (1) ; relevé 1090 : 1/ thérophytes : Lupinus angustifolius (1), Vicia pseudocracca (2a) ; relevé 1093 : 1/ vivaces : Cynodon dactylon (1), Euphorbia hirsuta (1) ; relevé 1119 : 1/ vivaces : Holcus lanatus (+), Scolymus hispanicus (r) ; relevé 1125 : 1/ vivace : Lotus tenuis (r) ; relevé 1178 : 1/ vivace : Lomelosia rutifolia (1).
Annexe 2
Les espèces ne figurant que dans un seul relevé ont été retirées du tableau 3. Elles sont listées ci-dessous.
relevé 1074 : 1/ thérophyte : Lolium rigidum (1), 2/ vivaces : Asparagus acutifolius (+), Centaurea sphaerocephala subsp. sphaerocephala (1), Chondrilla juncea (+), Plantago coronopus (1) ; relevé 1077 : vivace : Anthemis maritima (2b) ; relevé 651 : vivaces : Panicum repens (1), Setaria parviflora (1), Sporobolus pumilus (= Spartina patens, 2a) ; relevé 652 : 1/ thérophyte : Lotus angustissimus (1), 2/ vivaces : Oenanthe lachenalii (r), Rubus ulmifolius (1), en dehors du relevé : Alnus glutinosa, Crataegus monogyna, Quercus pubescens, Salix atrocinerea ; relevé 657 : thérophyte : Echium plantagineum (r) ; relevé 1155 : 1/ vivaces : Foeniculum vulgare (2a), Quercus suber (r) ; relevé 657′ : 1/ thérophytes : Knautia integrifolia (1), Trifolium stellatum (1), Urospermum dalechampii (+), 2/ vivaces : Artemisia vulgaris (2a), Asparagus acutifolius (1), Foeniculum vulgare (+), Prunus spinosa (+), Schoenus nigricans (2a), en dehors du relevé : Fraxinus ornus subsp. ornus, Pyrus spinosa, Quercus suber ; relevé 1097 : vivace : Lotus tenuis (+) ; relevé 1163 : 1/ thérophyte : Lupinus angustifolius (+), 2/ vivace: Matthiola sinuata (r) ; relevé 636 : vivaces : Euonymus europaeus (2b), Smilax aspera (3) ; relevé 634 : 1/ thérophytes : Brachypodium distachyon (2a), Hedypnois rhagadioloides (+), Gastridium ventricosum (+), Linum trigynum (+), Phedimus stellatus (1), Plantago bellardii (r), Polycarpon tetraphyllum subsp. tetraphyllum (1), Rumex bucephalophorus subsp. gallicus (1), Tuberaria guttata (+), Plantago bellardii (r), 2/ vivaces : Asplenium trichomanes (+), Brachypodium retusum (+), Dactylis glomerata (r), Euphorbia spinosa (r), Holcus lanatus (+), Petrorhagia saxifraga subsp. gasparrinii (+), Poterium sanguisorba (+), Sedum album (1), en dehors du relevé : Cistus monspeliensis, Genista corsica, Juniperus oxycedrus subsp. oxycedrus, Teucrium marum.
Bibliographie
Anonyme, 1999. Natura 2000, Manuel d’interprétation des habitats de l’Union européenne, EUR 15/2. Commission européenne, DG Environnement, Protection de la nature, zones côtières et tourisme, 127 p.
Anonymous, 2007. Interpretation Manual of European Union Habitats, EUR 27. European Commission, DG Environment – Nature and Biodiversity, 142 p.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Patrimoines naturels (Publications scientifiques du MNHN) 61 : 1-171.
CBNC (Conservatoire botanique national de Corse), 2016. EUNIS – Liste pour la Corse. Système d’information européen sur la nature. Classification des habitats. Office de l’environnement de la Corse-CBNC, Corte, 32 p.
CBNC (Conservatoire botanique national de Corse), 2020. Atlas biogéographique de la flore de Corse. Albiana-Office de l’environnement de la Corse, Ajaccio, 607 p.
Coste H., 1900-1906. Flore descriptive et illustrée de la France, de la Corse et des contrées limitrophes. Albert Blanchard, Paris, 3 volumes.
Delage A. & Hugot L., 2015. Liste rouge régionale de la flore vasculaire de Corse. Conservatoire botanique national de Corse-Office de l’environnement de la Corse, Corte, 72 p.
Delage A., Jeanmonod D. & Habashi C., 2018. Allium corsicum Jauzein et al. In D. Jeanmonod (ed.), Notes et contributions à la flore de Corse XXVI, Candollea 73 (1) : 77-78.
Devillers P., Devillers-Terschuren J., Ledant J.-P. et collab., 1991. CORINE biotopes manual. Habitats of the European Community. Data specifications, part 2. EUR 12587/3 EN. European Commission, Luxembourg, 300 p.
ENGREF, 1997. Nomenclature CORINE biotopes. Types d’habitats français (travail réalisé par M. Bissardon & L. Guibal, sous la direction de J.-C. Rameau). Muséum national d’histoire naturelle, Paris, 217 p.
Gargominy O., Tercerie S., Regnier C., Ramage T., Dupont P., Daszkiewicz P. & Poncet L., 2022. TAXREF, référentiel taxonomique pour la France : méthodologie, mise en œuvre et diffusion. Rapport PatriNat (OFB-CNRS-MNHN), Muséum national d’Histoire naturelle, Paris, 47 p.
Géhu J.-M., 2006. Dictionnaire de sociologie et synécologie végétales. J. Cramer, Berlin-Stuttgart, 899 p.
Géhu J.-M. & Rivas-Martínez S., 1981. Notions fondamentales de phytosociologie. Berichte der Internat. Symposium d. Internat. Vereinigung f. Vegetationskunde : 5-33.
IGN, 2019. Cartes à 1 : 25 000, TOP 25 (4250 OT-Corte, Monte Cinto ; 4253 ET-Aiguilles de Bavella, Solenzara ; 4348 OT-Bastia ; 4349 OT-Vescovato ; 4351 OT-Cervione, PNR de Corse ; 4352 OT-Aleria, Ghisonaccia). Institut national de l‘information géographique et forestière.
IGN/CdC, 2019. Photographies aériennes, mission Corse. Institut national de l‘information géographique et forestière.
Jauzein Ph., Tison J.-M., Deschâtres R. & Couderc H., 2002. Allium corsicum Jauzein, J.-M. Tison, Deschâtres & H. Couderc spec. nova, espèce méconnue de la flore corse. In D. Jeanmonod & A. Schlüssel (ed.), Candollea 56 (2) : 301-314.
Jeanmonod D. & Gamisans J., 2013. Flora Corsica, 2e éd. Bulletin de la Société botanique du Centre-Ouest, n. s., n° sp. 39 : 1-1074.
Le Berre M., Pires M. & Diadema K., 2019. Typification des actions – Que faire pour quelle espèce en région Provence-Alpes-Côte d’Azur ? Rapport inédit, CBNMéd, région PACA, 17 p. + ann.
Paradis G., Piazza C. & Ravetto S., 2002. Paysage végétal du site de Canna-Gradugine (cordon littoral, marais, prairies). Bulletin de la Société des sciences historiques et naturalistes de Corse 698-701 : 43-166.
Piazza C., Spinosi P., Quiquerez I., Delage A. & Hugot L., 2021. Stratégie de conservation de la flore vasculaire de Corse, 1re partie – Hiérarchisation des taxons et détermination des enjeux de conservation. Conservatoire botanique national de Corse-Office de l’environnement de la Corse, 102 p.
Piazza C., Spinosi P., Delage A. & Hugot L., 2022. Stratégie de conservation de la flore vasculaire de Corse, 2e partie – Typification des actions, priorisation et choix des projets. Conservatoire botanique national de Corse-Office de l’environnement de la Corse : 104 p.
Pignatti S., 1982. Flora d’Italia. Edagricole, 3 volumes.
RESEDA-FLORE (coord.), 2019. Glossaire pour la conservation de la flore méditerranéenne. Rapport inédit, CBNMéd, CBNA, CBNMC, CEFE-CNRS, IMBE, ISEM, 11 p.
Reymann J., Panaïotis Ch. & Bioret F. (coord.), 2016. Prodrome des végétations de Corse. Documents phytosociologiques, série 3, 4 : 1-175.
Tison J.-M. & de Foucault B. (coords), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.
UICN France, FCBN, AFB & MNHN, 2018. La Liste rouge des espèces menacées en France, chapitre Flore vasculaire de France métropolitaine. Paris, 31 p.
Vanden Berghen C., 1982. Initiation à l’étude de la végétation. Jardin botanique national de Belgique, Meise, 263 p.


