
Un printemps botanique et phytosociologique au pays du Matin calme (fin mars et avril 2023)
Title
A botanical and phytosociological spring in the Land of the Morning Calm (late March and April 2023)
Résumé
Suite à un voyage en Corée du Sud, l’auteur présente et illustre ici plusieurs habitats : habitats forestiers, habitats de lisière, habitats dunaires, habitats de prés salés, bas-marais, habitats anthropiques.
Abstract
Following a trip to South Korea, the author presents and illustrates several habitats here: forest habitats, edge habitats, dune habitats, salt meadow habitats, low marshes, anthropogenic habitats.
De fin mars à début mai 2023, un voyage en Corée du Sud, plus poétiquement appelé pays du Matin calme, a permis de se familiariser avec la flore et la végétation vernales de ce pays d’Asie orientale proche de la Chine et du Japon. On propose ici un compte rendu illustré des observations réalisées à cette occasion, élargi sur la base de la bibliographie disponible.
Pour la détermination des espèces rencontrées, on s’est appuyé sur la flore de Chine online (www.efloras.org), sur la flore du Japon (Ohwi, 1965), ainsi que sur la checklist de Chang et al. (2014). La nomenclature des taxons cités suit Catalogue of Life (www.catalogueoflife.org).
La carte 1 donne la localisation des principales étapes de ce périple.
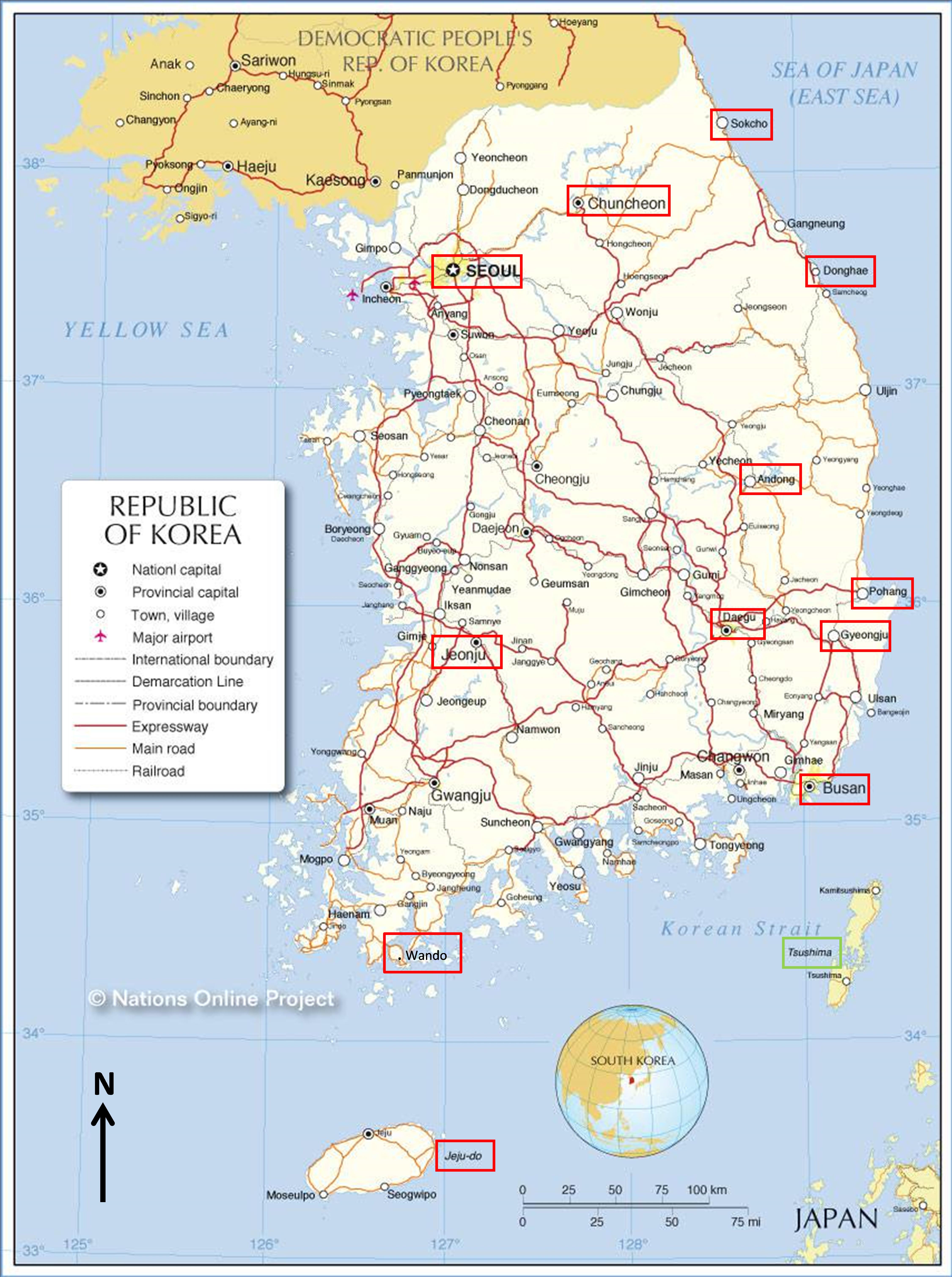
1. Généralités sur la Corée du Sud
À l’une des extrémités orientales de l’Eurasie, la Corée du Sud s’étend sur environ 100 000 km², 500 km du nord au sud et, au plus étroit, 216 km d’est en ouest. C’est un pays surtout granitique, localement calcaire et karstique ; l’île de Jeju-do est volcanique, avec tunnels de lave et volcans éteints (photo 1).

Le climat est tempéré, froid au nord, plus chaud au sud. Les températures moyennes oscillent entre – 5 °C en janvier et + 31 °C en août à Séoul (comme à Pyongyang, en Corée du Nord), entre + 8 °C et + 30 °C à Jeju-si (île de Jeju). La pluviométrie est marquée par la mousson estivale, pouvant atteindre 300 mm mensuels.
D’un point de vue phytogéographique, selon Takhtajan (1986), ce pays appartient à l’empire holarctique, à la région est-asiatique et au domaine nippo-coréen, notamment caractérisé par Sciadopitys verticillata (spontané au Japon et assez fréquemment planté en Corée ; photo 2), Magnolia sieboldii subsp. s. (photo 3), des Corylopsis (photo 4), Pterocarya rhoifolia (photo 5), Erythronium japonicum (photo 53), Abies koreana (photo 43), Pinus densiflora, Castanea crenata (photo 14), Buxus microphylla, Stephanandra incisa (photo 19), Acer palmatum (photo 92)… À cette liste, on pourrait ajouter la remarquable Ophioglossaceae Mankyua chejuense décrite postérieurement à la synthèse phytogéographique de Takhtajan et considérée jusqu’à présent comme endémique de Jeju-do (Sun et al., 2001).




2. La végétation forestière
La forêt occupe des surfaces importantes en Corée du Sud, colonisant quasiment tous les reliefs délaissés par les rizières, le maraîchage et l’urbanisme. On peut étendre à ce pays une observation déjà faite au Japon à propos des prairies : « au Japon, du moins dans la moitié nord d’Honshū, la prairie n’existe pas ; on observe plutôt des forêts, qui s’étendent jusqu’aux limites des villes, et des rizières » (de Foucault, 2016 : 7). En Corée, la continuité forestière est parfois interrompue par des clairières accueillant des tumulus et des stèles (photo 6) : la forêt est ponctuellement défrichée et le terrain plus ou moins aplani pour cet aménagement.
Comme leurs homologues européens, ces forêts présentent une stratification en strate arborescente, strate arbustive, strate herbacée vivace, strate muscinale et sont susceptibles d’être abordées selon le paradigme synusial (Gillet et al., 1991 ; de Foucault, 2020). Toutefois des recherches menées selon le paradigme classique ont donné lieu à des publications antérieures (surtout J.-W. Kim et al., 1996 ; Černý et al., 2014 ; Kolbek et al., 2003 pour la Corée du Nord ; bien d’autres seront utilisées dans la suite).

2.1. La végétation arborescente des îles Ulleng-do
Le tableau 1 décrit la végétation arborescente des îles Ulleng-do (ou encore écrit Ulreung-do) et Tok-do (ou Dog-do) situées en mer du Japon (J.-W. Kim et al., 1996). Deux associations peuvent être distinguées :
- l’une à Fagus multinervis (essence endémique d’Ulleng-do), Acer okamotoanum… (colonnes 1 à 4), l’Aceri okamotoani-Fagetum multinervis ass. nov. hoc loco, recouvrant l’« Hepatico maximae-Fagetum multinervis», typus nominis hoc loco : composante arborescente du relevé 11 du tableau 3 in-W. Kim et al. (1996, Report Surv. Nat. Environment Korea 10 : 151-153) ; elle présente une variation à Ulmus laciniata (colonne 2) et, apparemment, une variation à Taxus cuspidata de transition vers l’association suivante (colonne 4) ;
- l’autre à Pinus parviflora, Taxus cuspidata (photo 7), Tsuga sieboldii (colonnes 5 et 6), le Taxo cuspidatae-Pinetum parviflorae ass. nov. hoc loco, recouvrant le « Tsugo sieboldii-Pinetum parviflorae», typus nominis hoc loco : composante arborescente du relevé 2 du tableau 5 in-W. Kim et al. (1996, Report Surv. Nat. Environment Korea 10 : 157-158).

2.2. La végétation de Corée péninsulaire
La végétation arborescente
Kolbek et al. (2003), puis Černý et al. (2014) apportent des données bien plus conséquentes sur des forêts de la péninsule coréenne. La végétation arborescente décrite par ces travaux est synthétisée dans les tableaux 2 à 4.
Tout d’abord, le tableau 2 décrit des forêts d’affinités boréales caractérisées par Larix gmelinii var. olgensis, parmi lesquelles ont peut isoler :
- une association à gmelinii var. olgensis, Abies nephrolepis, Picea jezoensis, P. koraiensis (colonnes 4 à 9, et peut-être 2 et 3 plus fragmentaires), Abieti nephrolepidis-Laricetum olgensis ass. nov. hoc loco, recouvrant les « Rhododendro aurei-Laricetum olgensis», « Goodyero repentis-Piceetum jezoensis » et « Carici peiktusani-Abietetum nephrolepidis » de Kolbek et al. (2003, tableaux 8.1, 8.2 et 8.3), typus nominis hoc loco : composante arborescente (E3) du relevé 30 du tableau 8.1 in Kolbek et al. (2003, Geobotany 28 : 307-310), avec des variations à Sorbus aucuparia subsp. pohuashanensis (colonne 8) et à Betula pendula subsp. mandshurica (colonne 9) ;
- une association à gmelinii var. olgensis, Betula pendula subsp. mandshurica (colonnes 10 à 16), Betulo mandshuricae-Laricetum olgensis ass. nov. hoc loco, recouvrant les « Ledo procumbentis-Laricetum olgensis» et « Polysticho retroso-paleacei-Rhododendretum dahurici » de Kolbek et al. (2003, tableaux 8.4 et 8.5), typus nominis hoc loco : composante arborescente (E3) du relevé 21 du tableau 8.4 in Kolbek et al. (2003, Geobotany 28 : 316-317), avec une variation à Abies nephrolepis (colonne 10).
Cette végétation est évidemment homologue des taïgas européennes de la classe des Pino cembrae-Piceetea abietis Julve ex B. Foucault 2018. On peut rapprocher ces deux associations dans une alliance originale, l’Abieti nephrolepidis-Laricion olgensis all. nov. hoc loco, surtout caractérisée par Larix gmelinii var. olgensis, Abies nephrolepis, Picea jezoensis, P. koraiensis, Betula pendula subsp. mandshurica, typus nominis hoc loco : Abieti nephrolepidis-Laricetum olgensis nov.
Le tableau 3 synthétise des groupements forestiers arborescents possédant en commun Carpinus laxiflora (photo 8), Quercus mongolica (photo 9), Q. serrata, Q. acutissima (photo 10), Acer pseudosieboldianum (photo 11), A. pictum subsp. mono (photo 12), Magnolia sieboldii subsp. s. (photo 3), Fraxinus mandshurica… Deux coupures majeures passent entre les colonnes 12 et 13, puis 14 et 15.





L’ensemble 1 à 12 est différencié par Prunus leveilleana, la liane Actinidia arguta (photo 13), Kalopanax septemlobus… ; on peut au moins y distinguer le Querco mongolicae-Pinetum densiflorae ass. nov. hoc loco (colonnes 6 à 12), typus nominis hoc loco : composante arborescente (E3) du relevé 26 du tableau 8.12 in Kolbek et al. (2003, Geobotany 28 : 339-343), avec des variations à Fraxinus rhynchophylla… (colonne 6), à Quercus dentata… (colonne 7) et à Castanea crenata (colonne 12 ; photo 14). Cette association peut être le typus nominis hoc loco du Pruno leveilleanae-Quercion mongolicae all. nov. hoc loco, alliance caractérisée par Prunus leveilleana, Actinidia arguta, Carpinus cordata, Pinus koraiensis, Acer tatarica subsp. ginnala (photo 15), Kalopanax septemlobus. Non loin de cette alliance pourrait se placer une association encore indéfinie à Quercus variabilis, Fraxinus rhynchophylla et surtout Platycladus orientalis (= Thuja orientalis ; Cho & Bae, 2001).



L’ensemble 13 et 14 correspond à un ensemble caractérisé par Abies koreana, Betula ermanii subsp. e., Taxus cuspidata (photo 7) et Prunus maximowiczii. On ne peut définir une association originale pour la colonne 13 car les données utilisées (Černý et al., 2014) sont synthétiques. En revanche, on peut définir la colonne 14 comme Betulo chinensis-Abietetum koreanae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du premier relevé du tableau 1 in C.-S. Lee & Cho (1993, Korean J. Ecol. 16 (1) : 80-81).
Les colonnes 15 à 21 correspondent à un autre ensemble caractérisé par Toxicodendron trichocarpum, Vitis amurensis, Abies holophylla, Stewartia coreana, Meliosma myriantha, Cephalotaxus coreana…, sans doute aussi à valeur d’alliance, mais qui ne peut être validement définie.
Le large syntaxon décrit par les colonnes 1 à 48 possède la valeur d’un ordre original, les Carpino laxiflorae-Quercetalia mongolicae ord. nov. hoc loco, caractérisé par Carpinus laxiflora, Quercus mongolica, Q. serrata, Q. acutissima, Acer pseudosieboldianum, A. pictum subsp. mono, Magnolia sieboldii subsp. s., Fraxinus mandshurica, typus nominis hoc loco : Pruno leveilleanae-Quercion mongolicae nov.
Dans ce tableau 3, il reste l’ensemble 49 à 52, qui décrit des forêts subtropicales à essences sempervirentes (photo 16), dont Camellia japonica (photo 17), Castanopsis cuspidata, Machilus thunbergii (photo 18), M. japonica, Cinnamomum japonicum, Neolitsea sericea, Pinus thunbergii (photo 40), Celtis sinensis. Ce syntaxon ne pouvant être validement défini, il faut tenter de s’orienter vers d’autres sources pour préciser leur composition, lesquelles sont synthétisées dans le tableau 4. De telles forêts sont aussi évoquées par O.-J. Lee (1981).



Dans l’ensemble des colonnes 1 à 8 de ce tableau 4, on peut au moins mettre en avant le Castanopsio sieboldii-Camellietum japonicae ass. nov. hoc loco (colonne 1), typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 12 du tableau 1 in C.-S. Kim & Jang [1989, Korean J. Ecol. 12 (3)], et le Camellio japonicae-Pinetum thunbergii ass. nov. hoc loco (colonne 4), typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 11 du tableau 2 in C.-S. Kim & Jang [1989, Korean J. Ecol. 12 (3)]. Cet ensemble possède la valeur d’une alliance, le Castanopsio sieboldii-Camellion japonicae all. nov. hoc loco, caractérisée par Castanopsis sieboldii, Celtis sinensis, Machilus japonica, typus nominis hoc loco : le Castanopsio sieboldii-Camellietum japonicae nov.
L’ensemble formé par les colonnes 10 à 13 possède aussi la valeur d’une alliance, mais les données souces, synthétiques, ne permettent pas une validation définitive.
La colonne 9, à Torreya nucifera, ne peut être rattachée à aucune de ces deux alliances, peut-être à cause du nombre réduit de relevés.
Avec ces deux unités, l’ensemble du tableau 4 définit l’ordre des Machilo thunbergii-Camellietalia japonicae ord. nov. hoc loco, ordre caractérisé par Camellia japonica, Carpinus coreana et les Lauraceae Machilus thunbergii, Cinnamomum japonicum, Neolitsea sericea, typus nominis hoc loco : le Castanopsio sieboldii-Camellion japonicae nov., et la classe des MACHILO THUNBERGII-CAMELLIETEA JAPONICAE classis nov. hoc loco (dont les taxons caractéristiques coïncident avec ceux de l’ordre), typus nominis hoc loco : les Machilo thunbergii-Camellietalia japonicae nov.
Le gradient de végétation menant des taïgas de l’Abieti nephrolepidis-Laricion olgensis aux chênaies-charmaies des Carpino laxiflorae-Quercetalia mongolicae et aux forêts subtropicales des Machilo thunbergii–Camellietea japonicae est tout à fait homologue à celui que l’on observe en Europe avec les taïgas des Pino cembrae-Piceetea abietis, les forêts caducifoliées des Fraxino excelsioris-Quercetea roboris Gillet ex B. Foucault, Gillet & Julve in B. Foucault 2020 et les forêts sempervirentes des Pino halepensis-Quercetea ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020. Du point de vue des structures formelles systématiques (de Foucault, 1987), dans les deux cas les taïgas relèvent des S(Rosaceae-Pinaceae) et les chênaies-charmaies des S(Betulaceae-Fagaceae) ; comme on l’a vu, les forêts de Camellia combinent surtout des Lauraceae (Neolitsea, Cinnamomum, Machilus), des Fagaceae (Castanopsis), des Theaceae (Camellia), rappelant ainsi les laurisylves des S(Aquifoliaceae-Lauraceae).
La végétation arbustive intraforestière
Le tableau 5 synthétise la diversité de la végétation arbustive intraforestière déduite de plusieurs des données déjà citées. Une coupure majeure passe entre les colonnes 16 et 17.
L’ensemble constitué par les colonnes 1 à 16 rassemble des fourrés boréo-asiatiques associés aux boisements de l’Abieti nephrolepidis-Laricion olgensis défini plus haut ; plusieurs essences caractéristiques de cette alliance y apparaissent sous des formes juvéniles. Les colonnes 1 et 2 correspondent à un fourré à Rhododendron aureum qu’on peut définir sous le nom de Larici olgensis-Rhododendretum aurei ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 8 du tableau 8.1 in Kolbek et al. (2003, Geobotany 28 : 307-310) ; les colonnes 3 et 4 pourraient y être rattachées en tant que variations significatives. La colonne 7 peut être distinguée sous le nom de Roso davuricae-Loniceretum caeruleae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 3 du tableau 8.3 in Kolbek et al. (2003, Geobotany 28 : 313-315). La colonne 13 peut être isolée sous le nom de Rhododendro parvifolii-Loniceretum caeruleae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 21 du tableau 8.4 in Kolbek et al. (2003, Geobotany 28 : 316-317). Les colonnes 14 et 15 décrivent globalement le Roso davuricae-Rhododendretum daurici ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 18 du tableau 8.5 in Kolbek et al. (2003, Geobotany 28 : 318-319).
Globalement, ce premier ensemble possède la valeur d’une alliance, le Roso davuricae-Lonicerion caeruleae all. nov. hoc loco, caractérisée par Rosa davurica, Lonicera caerulea subsp. c., Malus baccata, Dasiphora fruticosa, Clematis ochotensis et des formes juvéniles de diverses essences de l’Abieti nephrolepidis-Laricion olgensis, typus nominis hoc loco : Roso davuricae-Loniceretum caeruleae nov. Manifestement ce syntaxon boréo-est-asiatique est vicariant des alliances euro-subarctiques et subapines des Betulo pubescentis-Alnetea alnobetulae Rejmánek in Huml et al. 1979, dont Lonicera caerulea subsp. c. est aussi une bonne caractéristique (de Foucault, 2012).
Au sein du grand ensemble formé par les colonnes 17 à 35, deux coupures secondaires passent entre les colonnes 25 et 26 d’un côté, 32 et 33 d’un autre.
Dans l’ensemble 17 à 25, le syntaxon défini par la colonne 24 est un des plus significatifs : Deutzio grandiflorae-Rhododendretum mucronulati ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 17 du tableau 8.12 in Kolbek et al. (2003, Geobotany 28 : 339-343). Un second est celui défini par la colonne 21 : Rhododendro mucronulati-Juniperetum rigidae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive et arbrisselée (E2) du relevé 14 du tableau 8.10 in Kolbek et al. (2003, Geobotany 28 : 331-334). Plus largement, cet ensemble correspond à une alliance originale, le Lindero obtusilobae-Rhododendrion mucronulati all. nov. hoc loco, caractérisée ou différenciée des alliances voisines par Lindera obtusiloba, Stephanandra incisa (photo 19), Lespedeza juncea, Symplocos paniculata, Corylus heterophylla, Rhamnus davuricus, Euonymus oxyphyllus, E. alatus (photo 20), Kalopanax septemlobus, Vitis amurensis, Vaccinium hirtum subsp. koreanum, Actinidia polygama, Viburnum carlesii, Ligustrum obtusifolium (photo 21), typus nominis hoc loco : Rhododendro mucronulati-Juniperetum rigidae nov.



Les ensembles 26 à 32 et 33 à 35 sont définis sur des données synthétiques, de sorte qu’on ne peut valider de nouveaux syntaxons. Parmi les taxons cités pour ces deux ensembles, on a au moins rencontré Daphniphyllum macropodum (photo 22), Juniperus rigida (photo 23), Akebia quinata (photo 24), Eurya japonica (photo 25), Stauntonia hexaphylla (photo 26), auxquels on peut ajouter Eurya emarginata (photo 27).






L’ensemble 17 à 32 à la valeur d’un ordre, les Styraco obassiae-Rhododendretalia mucronulati ord. nov. hoc loco, caractérisé par Rhododendron mucronulatum, Rh. schlippenbachii (photo 28), Styrax obassia, Lespedeza maximowiczii, L. bicolor, Cornus controversa, Weigela florida (photo 29), Zanthoxylum schinifolium (photo 30), Rubus crataegifolius, Juniperus rigida, Actinidia arguta, typus nominis hoc loco : Lindero obtusilobae-Rhododendrion mucronulati nov.



Pour préciser la composition des sous-bois arbustifs des forêts de Camellia japonica, on a élaboré le tableau 6, qui indique les formes juvéniles des essences potentiellement arborescentes pour les colonnes 1 à 4, où ces données sont disponibles, au contraire des colonnes 5 à 13 (cellules non remplies). Seules d’ailleurs les colonnes 1 à 4 peuvent donner lieu à une définition valide d’une association, le Smilaco chinae-Ligustretum japonici ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (S) du relevé 25 du tableau 1 in C.-S. Kim & Jang (1989, Korean J. Ecol. 12 (3) : 148).
L’ensemble des colonnes de ce tableau 6 paraît posséder la valeur d’une alliance, le Smilaco chinae-Euryion japonicae all. nov. hoc loco, caractérisée par Smilax china, Eurya japonica (photo 25), Callicarpa japonica, C. mollis, Viburnum erosum, Rhaphiolepis umbellata (photo 31), Ligustrum japonicum et plusieurs formes juvéniles des essences caractérisant les Machilo thunbergii-Camellietea japonicae. On retrouve cette même alliance au Japon (Tanaka & Ito, 1981).

Données sur la végétation hémicrypto-géo-chaméphytique intraforestière
En ce qui concerne la description des strates intraforestières basses, il n’est évidemment pas question d’aller aussi loin que dans les paragraphes précédents, car leur homogénéité floristique est incertaine ; on se contentera de quelques commentaires et d’illustrations de certains des taxons cités.
Pour les forêts boréo-asiatiques de l’Abieti nephrolepidis-Laricion olgensis, les données de Kolbek et al. (2003) montrent le développement d’une végétation riche en Lycopodiaceae (Diphasiastrum complanatum, Phlegmariurus cryptomerianus, Lycopodium clavatum, Dendrolycopodium obscurum, Huperzia miyoshiana, H. selago) et Ericaceae (Phyllodoce caerulea, Rhododendron tomentosum, Vaccinium uliginosum, V. vitis-idaea, Orthilia secunda, Chimaphila japonica, Pyrola japonica, P. dahurica, P. asarifolia subsp. incarnata), plus Salix arctica, Juniperus communis subsp. nana, Linnaea borealis, Calypso bulbosa… ; en terme de structures systématiques, elle peut se rattacher à la S(Lycopodiaceae-Ericaceae).
Pour les forêts des Carpino laxiflorae-Quercetalia mongolicae, on trouve cités les taxons suivants : Hedera rhombea (photo 32), Rubia chinensis (photo 33), Stemmacantha uniflora (photo 34), Leibnitzia anandria (photo 35), Disporum smilacinum (photo 36), des Pseudostellaria (photo 37), Asarum sieboldii (photo 38) ; de cet asaret, on peut rapprocher le spectaculaire Isotrema manshuriense rencontré au parc national de Seoraksan (photo 39).








2.3. La végétation forestière de Jeju-do
La connaissance de la végétation de l’île volcanique méridionale de Jeju-do est due notamment à la publication de Yim et al. (1990), lesquels ont rassemblé une centaine de relevés rattachés à des associations selon la méthode phytosociologique classique.
La végétation arborescente
Le tableau 7 revient sur une définition plus moderne de ces associations forestières.
À basse altitude (colonne 1 ; autour de 150 m), c’est la forêt à Torreya nucifera et Lauraceae, Neolitseo aciculatae-Torreyetum nuciferae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 9 du tableau 9 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 126-128), recouvrant l’« Ardisio japonicae-Torreyetum nuciferae ».
Un peu plus haut (colonne 2 ; 330 à 650 m, exceptionnellement 1 065 m), s’étend la forêt à Pinus thunbergii (photo 40), le Pinetum densifloro-thunbergii ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 6 du tableau 8 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 124-126).

De 1 050 à 1 430 m (colonne 3), les auteurs décrivent une forêt à Acer pseudosieboldianum (photo 11) et Quercus mongolica (photo 9), le Querco mongolicae-Aceretum pseudosieboldiani ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 3 du tableau 3 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 109-110, en remplaçant Quercus grosseserrata par Q. mongolica), recouvrant l’« Hepatico asiaticae–Quercetum grosseserratae ». C’est sans doute cette association que nous avons parcourue au départ de l’ascension vers Hallasan, vers 1 150 m (photo 41). Sur certains arbres, on peut y observer l’espèce hémiparasite Viscum coloratum (photo 42).


De 550 à 1 450 m (colonnes 4 à 6), s’étendent diverses formes d’un unique Carpino laxiflorae-Aceretum pseudosieboldiani ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 3 du tableau 6 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 119-121), recouvrant les « Asaro maculati–Quercetum serratae », « Daphniphyllo macropodi-Carpinetum tschonoskii » et « Saso quelpaertensis-Carpinetum laxiflorae ».
La colonne 7 correspond à une forêt encore indéfinie (seulement cinq relevés) d’une pinède à Pinus densiflora et Acer pseudosieboldianum.
Enfin (colonne 8), vers le sommet du mont Halla (1 520 à 1 830 m), s’étend la sapinière à Abies koreana (photo 43), Betulo ermanii-Abietetum koreanae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T1 + T2) du relevé 3 du tableau 2 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 108), recouvrant le « Saso quelpaertensis-Abietetum koreanae ». On retrouve encore cette végétation dans le tableau 1 de Song & Nakanishi (1985). Selon M.-H. Kim et al. (1999), la régression de cette forêt mène au développement d’une lande et d’un fourré qu’ils ont dénommés respectivement Festuco ovinae-Empetretum nigri (à dénommer plutôt Festuco ovinae-Empetretum asiatici ; il faudrait toutefois revoir la détermination du premier taxon éponyme) et Rhododendretum mucronulati. On en a rapproché (colonnes 9 et 10) des boisements à Abies koreana de la péninsule.

L’ensemble formé par les colonnes 3 à 7 possède manifestement la valeur d’une alliance, le Carpino tschonoskii-Acerion pseudosieboldiani all. nov. hoc loco, caractérisée par Acer pseudosieboldianum, Carpinus tschonoskii, C. laxiflora, Prunus sargentii, P. maximowiczii, Taxus cuspidata, Maackia fauriei…, typus nominis hoc loco : Carpino laxiflorae-Aceretum pseudosieboldiani nov., alliance pouvant être incluse dans les Carpino laxiflorae-Quercetalia mongolicae définis en Corée péninsulaire (cf. supra, § 2.2).
Le Betulo ermanii-Abietetum koreanae nov. n’entre pas dans cette alliance et peut être considéré comme le typus nominis hoc loco du Betulo ermanii-Abietion koreanae all. nov. hoc loco, caractérisé par Abies koreana (photo 43), Betula ermanii subsp. e., alliance dans laquelle on peut aussi inclure le Betulo chinensis-Abietetum koreanae (colonne 10) défini en Corée péninsulaire (cf. supra, § 2.2).
On ne peut statuer pour l’instant sur le rattachement des Neolitseo aciculatae-Torreyetum nuciferae et Pinetum densifloro-thunbergii.
La végétation arbustive intraforestière
Le tableau 8 reprend la végétation arbustive intraforestière de Jeju-do.
De basse altitude et liée au Neolitseo-Torreyetum nuciferae, la colonne 1 décrit le Kadsuro japonicae-Orixetum japonicae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (S) du relevé 7 du tableau 9 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 126-128).
Reliée à la précédente, la colonne 2 peut être individualisée sous le nom de Rhoo chinensis-Ligustretum obtusifolii ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (S) du relevé 3 du tableau 8 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 126-128).
Laissant de côté la colonne 3, surtout définie par des formes juvéniles de taxons potentiellement arborescents, les colonnes 4 à 7 semblent se rattacher à un unique Ilici crenatae-Daphniphylletum macropodi ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (S) du relevé 2 du tableau 5 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 116-118).
Enfin, vers le sommet du mont Halla, la colonne 8 décrit le Lonicero maackii-Rhododendretum mucronulati ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (S) du relevé 11 du tableau 2 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 108). Il est assez distinct du Rhododendretum mucronulati décrit par M.-H. Kim et al. (1999) du même massif, synthétisé dans ce même tableau en colonne Rm.
L’ensemble formé par les colonnes 3 à 7 paraît posséder la valeur d’une alliance, le Daphniphyllo macropodi-Styracion japonici all. nov. hoc loco, caractérisée par Styrax japonicus, Daphniphyllum macropodum (photo 44), Ilex crenata, Callicarpa mollis, Cornus kousa (photo 45), C. controversa, Viburnum furcatum, Neoshirakia japonica…, typus nominis hoc loco : l’Ilici crenatae-Daphniphylletum macropodi nov.


La végétation herbacée intraforestière
Le tableau 9 reprend la végétation herbacée intraforestière de Jeju-do. Pour le simplifier, les formes juvéniles des taxons potentiellement arborescents et arbustifs, ainsi que les taxons herbacés de présence inférieure à II n’y sont pas reportés.
La colonne 1 peut être distinguée comme Arisaemo ringentis-Ardisietum japonicae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 1 du tableau 9 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 126-128).
La colonne 2 peut être interprétée comme Cocculo orbiculati-Oplismenetum undulatifolii ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 1 du tableau 8 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 124-125).
Les colonnes 3 à 8 corespondent à des végétations herbacées intraforestières souvent massivement dominées par le petit bambou Sasa palmata (= S. quelpaertensis ; photo 46). Le syntaxon de la colonne 3 est décrit ici sous le nom de Asaro sieboldii-Sasetum palmatae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 3 du tableau 3 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 109-111). La colonne 5 est distinguée sous le nom de Violo albidae-Sasetum palmatae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 1 du tableau 5 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 109-111, en remplaçant Viola dissecta var. chaerophylloides par V. albida) ; la colonne 4 pourrait être être une variation significative de cette dernière association. La colonne 6 est un Codonopsio lanceolatae-Sasetum palmatae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 4 du tableau 6 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 119-121). Laissant de côté la colonne 7 qui synthétise seulement cinq relevés, la colonne 8 peut être isolée comme Maianthemo bifolii-Sasetum palmatae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (H) du relevé 4 du tableau 2 in Yim et al. (1990, Korean J. Ecol. 13 (2) : 108).

L’ensemble des colonnes 1 et 2 possède la valeur d’une alliance, le Phrymo nanae-Dioscoreion japonicae all. nov. hoc loco, notamment caractérisée par Dioscorea japonica, D. quinquelobata, Phryma nana, Isodon inflexus…, typus nominis hoc loco : Arisaemo ringentis-Ardisietum japonicae nov. Il en est de même pour l’ensemble 3 à 6, Hylodesmo oxyphylli-Sasion palmatae all. nov. hoc loco, alliance notamment caractérisée par Codonopsis lanceolata, Hylodesmum podocarpum subsp. oxyphyllum (= Desmodium oxyphyllum), Hedera rhombea (photo 32), Oplismenus undulatifolius, Ophiopogon japonicus, Trachelospermum asiaticum (photo 47), Liriope muscari, Arisaema amurense (photo 48), A. heterophyllum, Viola acuminata, V. rossii, Disporum smilacinum (photo 36), Ainsliaea apiculata, Carex siderosticta subsp. pilosa, typus nominis hoc loco : Asaro sieboldii-Sasetum palmatae nov. La colonne 7 pourrait se rattacher aussi à cette alliance, mais bien en limite.


Lors de nos investigations floristiques, diverses géophytes vernales et d’autres espèces ont été rencontrées, non citées dans les travaux précédemment cités. Leurs communautés sont évidemment notamment vicariantes de celles du Scillion bifoliae d’Europe occidentale (de Foucault, 2020). Les principaux taxons sont Adoxa moschatellina, Allium monanthum (photo 49 ; taxon déterminé avec l’assistance de J.-M. Tison), des Maianthemum (quatre espèces en Corée du Sud ; photo 50) et Polygonatum (onze espèces en Corée du Sud ; photo 51), Amana edulis (photo 52), Erythronium japonicum (photo 53), Gentiana squarrosa (photo 54), Hepatica maxima (photo 55), Nanocnide japonica (photo 56), Petasites japonicus (photo 57), Semiaquilegia adoxoides (photo 58), des Corydalis (vingt-trois espèces en Corée du Sud ; photos 59 et 60).












3. La végétation des lisières
Au voisinage des forêts, quelques lisières ont fait l’objet d’observations floristiques. En premier lieu, on peut évoquer cet ourlet thermophile observé à Donghae, sur la colline boisée proche de Cheongokdonggul, connu pour sa grotte. C’est en effet à ce niveau qu’on a pu observer une communauté à Dictamnus albus (photo 61), Ajuga multiflora (photo 62), Polygala cf. japonica (photo 63), Thalictrum minus subsp. thunbergii (photo 64) qui rappelle un peu le Geranio sanguinei-Dictamnetum albi Wendelberger ex T. Müll. 1962 décrit d’Europe centrale où le second taxon éponyme peut apparaître associé à Thalictrum minus subsp. saxatile.




En ce qui concerne les lisières arbustives, ou manteaux, on peut évoquer le fourré colonisant la falaise littorale de Seongsan Ilchulbong (Jeju-do) qui accueille Elaeagnus umbellata (photo 65), Eurya emarginata (photo 27), Litsea japonica (photo 66), Pittosporum tobira (photo 67), Euonymus japonicus (photo 68), Akebia quinata (photo 24), Hedera rhombea (photo 32), fourré illustré en photo 69.





Dans des lisières arbustives à Seogwipo (sud de Jeju-do), on peut rencontrer le seul poivrier spontané du pays, Piper kadsura (photo 70).

Mais la grande classe de végétation de lisière arbustive dans le domaine nippo-coréen reste celle des Rosetea multiflorae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 86) qui se divise en deux ordres, les Rosetalia rugosae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 86) pour les fourrés littoraux, qui seront analysés dans le paragraphe consacré à la végétation dunaire (cf. § 4.2), et les Rosetalia multiflorae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 102) pour les fourrés de l’intérieur.
Étant donné que la belle étude de Ohba et al. (1973) porte surtout sur la végétation dunaire, le nom de Rosetalia multiflorae est invalide, n’étant pas soutenu par une association valide. Il semble qu’iI faille plutôt utiliser le nom de Dioscoreo-Puerarietalia lobatae Ohba 1973, incluant pour la Corée les Lonicero japonicae-Puerarion lobatae Jung & J.-W. Kim 1998 (Korean J. Ecol. 21 (6) : 740) et Dioscoreo-Actinidion argutae Jung & J.-W. Kim 1998 (Korean J. Ecol. 21 (6) : 740). Ces noms en Dioscoreo… ne peuvent être précisés car deux Dioscorea participent à cette végétation, D. nipponica et D. japonica, le nom Dioscoreo-Puerarietalia lobatae est valide, mais pas le Dioscoreo-Actinidion argutae, pas plus d’ailleurs que l’association type de cette alliance, le Dioscoreo-Actinidietum argutae Jung & J.-W. Kim 1998 (art. 3g). Il faut donc valider l’association en Dioscoreo nipponicae-Actinidietum argutae Jung & J.-W. Kim ex ass. nov. hoco loco [typus nominis hoc loco : relevé du tableau 9 in Jung & J.-W. Kim (1998, Korean J. Ecol. 21 (6) : 746)] et l’alliance en Dioscoreo nipponicae-Actinidion argutae Jung & J.-W. Kim ex all. nov. hoc loco, typus nominis hoc loco : Dioscoreo nipponicae-Actinidietum argutae Jung & J.-W. Kim ex B. Foucault nov. Le tableau 10 synthétise quelques associations du Dioscoreo nipponicae-Actinidion argutae déduites du tableau 1 in Jung & Kim (1998) pour les syntaxons 1 à 8 (pour les syntaxons 9 à 14, voir infra, § 4.2) : 1 – Mallotetum japonicae Jung & J.-W. Kim 1998, 2 – Clerodendretum trichotomi Jung & J.-W. Kim 1998 (‘… trichotomae’, art. 41b), 3 – Akebietum quinatae Jung & J.-W. Kim 1998, 4 – Lonicero japonicae-Puerarietum lobatae Jung & J.-W. Kim 1998, 5 – Spiraeetum salicifoliae (‘Spiraetum…’, art. 41b), 6 – Dioscoreo nipponicae-Actinidietum argutae Jung & J.-W. Kim ex B. Foucault nov., 7 – Sorbarietum stellipilae Jung & J.-W. Kim 1998, 8 – Tripterygietum wilfordii Jung & J.-W. Kim 1998 nom. mut. hoc loco (initialement Tripterygietum regelii).
Plus au sud (Nansei Islands au sud-ouest du Japon), les Rosetea multiflorae sont relayés par une classe d’affinités plus nettement tropicales, dont deux alliances sont connues, le Clerodendro inermis-Derrion trifoliatae A. Miyaw. et al. 1985 et l’Aristolochio liukiuensis-Clematidion ryukyuensis Murakami 1991 (Murakami, 1991).
4. La végétation des dunes côtières
4.1. La végétation herbacée
La végétation des dunes sableuses circumboréales a fait récemment l’objet d’une synthèse phytosociologique (de Foucault, 2021a) au sein de la classe des Honckenyo peploidis-Leymetea arenarii Tüxen 1966, qui inclut notamment l’ordre boréo-japonais des Honckenyo majoris-Leymetalia mollis Ohba, A. Miyaw. & Tüxen 1973. Avec Leymus mollis (photo 71), Convolvulus soldanella, Carex pumila (photo 72), C. kobomugi (photo 73), Lathyrus japonicus (photo 74), Glehnia littoralis (photo 75), Ixeris repens (photo 76), Ischaemum anthephoroides (photo 77), la végétation du nord de la péninsule coréenne peut également se rattacher à cet ordre. Celle-ci a été préalablement étudiée par Min & Je (2002), qui apportent peu de données phytosociologiques, puis J.-S. Lee et al. (2007), qui publient un tableau synthétique, toutefois difficilement compréhensible, certaines lignes montrant deux taxons, dont certains apparaissant en outre dans d’autres lignes. De toute façon, ces auteurs ne décrivent que des communautés, pas des unités phytosociologiques valides.







De notre côté, le 4 avril, on a parcouru la plage de sable volcanique de Seongsampo Beach Park (N 33° 27’ 20,5’’, E 126° 55’ 38,3’’) où Ixeris repens est associé à Convolvulus soldanella, Lathyrus japonicus, Peucedanum japonicum (photo 78), Vitex rotundifolia (photo 79), Crinum asiaticum var. japonicum (photo 80), une Boraginaceae indéterminée (photo 81). Une telle végétation est perceptible aussi dans le tableau 1 de Nakanishi (1980) qui la décrit sous le nom de Vitici rotundifoliae-Crinetum japonici.




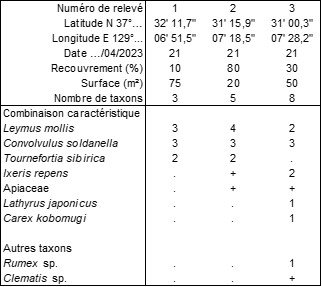
Annexe du tableau 11 (tous les relevés viennent du littoral de Donghae, prov. de Gangwon-do) – rel. 1 : plage au sud du parc Bugog, Haeandoldamsup ; rel. 2 : plage Hanseom Beach côté nord, au bout de Donggal-ro ; rel. 3 : plage Hanseom Beach côté sud.
Le 21 avril, c’est le bord de mer de Donghae qu’on a étudié avec trois relevés assez précis réunis dans le tableau 11. Cette végétation est caractérisée ici par Tournefortia sibirica (syn. : Messerschmidia sibirica ; photo 82) et s’apparente nettement au “Messerschmidia sibirica-Elymus mollis Ges. Okuda, Fujiwara & A. Miyaw. 1970” cité à nouveau mais sans validation par Ohba et al. (1973, Vegetatio 26 (1-3) : 34). Le fait de le retrouver sur certaines côtes coréennes conforte sa valeur d’association : Tournefortio sibiricae-Leymetum mollis Okuda, Fujiwara & A. Miyaw. ex ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 7 in Ohba et al. (1973, Vegetatio 26 (1-3) : 34) (photo 83). La validation de cette association permet aussi de valider définitivement le Messerschmidio sibiricae-Elymion mollis Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 33) nom. inval. (art. 8) sous le nom de Tournefortio sibiricae-Leymion mollis Ohba, A. Miyaw. & Tüxen ex hoc loco, typus nominis hoc loco : Tournefortio sibiricae-Leymetum mollis Okuda, Fujiwara & A. Miyaw. ex B. Foucault hoc loco.


Le 24 avril, enfin, c’est le littoral sableux de Sokcho qui a fait l’objet d’observations floristiques :
sud de Sokcho Beach, N 38° 12’ 06’’, E 128° 35’ 14,9’’, 70 % 30 m², 8 taxons
- Lathyrus japonicus 3, Convolvulus soldanella 3, Ixeris repens 2, Carex pumila 2, Mertensia simplicissima 1 (photo 84), Leymus mollis +, Glehnia littoralis +,
- autre taxon : Rumex acetosella +,
qui peut se rattacher au Mertensio asiaticae-Elymetum mollis (Tatewaki 1931) Ohba, A. Miyaw. & Tüxen 1973 muté ici en Mertensio simplicissimae-Leymetum mollis (Tatewaki 1931) Ohba, A. Miyaw. & Tüxen 1973 nom. mut. hoc loco, initialement décrit des côtes du nord d’Honshū, de Hokkaido et des îles Kouriles (Ohba et al., 1973) et se rattachant au Jacobaeo pseudoarnicae-Leymion mollis Ohba, A. Miyaw. & Tüxen 1973 (de Foucault, 2021a).

L’ordre subboréal des Honckenyo majoris-Leymetalia mollis est relayé vers le sud, sous climat plus chaud, par les Viticetea rotundifoliae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 73) et les Viticetalia rotundifoliae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 73), incluant l’Ischaemo anthephoroidis-Viticion rotundifoliae Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 77) et le Thuareo involutae–Viticion rotundifoliae Ohba in Ohba, A. Miyaw. & Tüxen 1973 (Vegetatio 26 (1-3) : 75). Il semble bien que les Glehnietea littoralis et Glehnietalia littoralis cités en nominis nudi par Tüxen (1966) soient des synonymes invalides de ces syntaxons à Vitex rotundifoliae. Le tableau 12 synthétise les données du tableau 1 de Jung (2000a), avec les syntaxons suivants (Jung, 2000b, apporte aussi des données sur certains de ces syntaxons).
- Linario japonicae-Viticetum rotundifoliae Ohba, A. Miyaw. & Tüxen 1973.
- Imperato cylindricae-Viticetum rotundifoliae (Ochi 1951) Ohba, A. Miyaw. & Tüxen 1973.
- « Roso–Viticetum rotundifoliae», notamment marqué par la présence de Paederia scandens (photo 85).
- 4a. « Vitici rotundifoliae-Juniperetum confertae » ; 4b correspond à la même association, mais selon le tableau 20 in Ohba et al. (1973) ; en séparant la composante arbustive (cf. infra, § 4.2, Pino thunbergii-Juniperetum confertae), l’association herbacée peut être modernisée en Indigofero bungeanae-Viticetum rotundifoliae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 9 du tableau 20 in Ohba et al. (1973, Vegetatio 26 (1-3) : 84).
- « Vitici rotundifoliae-Juniperetum procumbentis».
- Groupement à Vitex rotundifolia.
- Groupement à Vitex rotundifolia-Elaeagnus umbellata.
- Groupement à Lathyrus japonicus-Rosa lucieae.
- Thuareo involutae-Viticetum rotundifoliae A. Miyaw. & Suzuki 1976.
- Wedelietum biflorae A. Miyaw. & Suzuki 1976 muté en Wollastonietum biflorae nom. mut. hoc loco.
- Groupement à Rhaphiolepis umbellata-Juniperus taxifolia.

Il montre que le Roso-Viticion rotundifoliae Nakanishi 1979 (colonnes 3 à 5) n’est guère distinct de l’Ischaemo anthephoroidis-Viticion rotundifoliae (colonnes 1, 2 et 8) et qu’il doit par conséquent y être inclus, alors que le Thuareo involutae–Viticion rotundifoliae s’affranchit des dernières irradiations des Honckenyo-Leymetalia mollis qui persistent encore dans l’Ischaemo-Viticion rotundifoliae ; Thuarea involuta est une poacée vivace de plage plus ou moins tropicale (photo 86).

4.2. La végétation arbustive
La végétation arbustive des fonds de dune reliée aux Honckenyo-Leymetalia mollis se rattache aux Rosetalia rugosae et aux Rosetea multiflorae et est caractérisée surtout par Rosa rugosa (photo 87). Plusieurs associations sont synthétisées dans les colonnes 9 à 14 du tableau 10, respectivement Viti coignetiae-Rosetum rugosae Ohba, A. Miyaw. & Tüxen 1973, Roso rugosae-Maletum mandshuricae Ohba, A. Miyaw. & Tüxen 1973, « Potentillo fragarioidis-Rosetum rugosae » Ohba, A. Miyaw. & Tüxen 1973 prov., Roso rugosae-Juniperetum confertae Ohba, A. Miyaw. & Tüxen 1973, Elaeagno umbellatae-Rosetum rugosae Ohba, A. Miyaw. & Tüxen 1973 et un groupement indéterminé à R. rugosa. Le troisième syntaxon de cette liste, d’ailleurs décrit à titre provisoire et sur la base de seulement quatre relevés, n’est pas conforme au paradigme suivi ici. On retient le Roso rugosae-Juniperetum confertae Ohba, A. Miyaw. & Tüxen 1973 comme lectotypus nominis hoc loco du Rosion rugosae.

La présence d’arbustes dans la végétation des Viticetea rotundifoliae incite aussi à séparer des associations biologiquement plus homogènes, mais qu’il n’est pas toujours possible de valider, car les données sont souvent synthétiques. Ces données sont reprises dans la partie inférieure du tableau 12 à partir du tableau 1 de Jung (2000a). Dans le « Vitici rotundifoliae-Juniperetum confertae », avec l’assistance des données apportées par Ohba et al. (1973), en colonne 4b, on peut cependant distinguer le Pino thunbergii-Juniperetum confertae ass. nov. hoc loco, caractérisé par les lianes Cocculus orbiculatus et Clematis terniflora en plus des deux taxons éponymes, typus nominis hoc loco : composante arbustive du relevé 7 du tableau 20 in Ohba et al. (1973, Vegetatio 26 (1-3) : 84). En ce qui concerne la colonne 5, on n’a pas eu accès à des données détaillées pour la Corée, on peut cependant trouver dans Itow et al. (1986, tableaux 9 et 10) deux fourrés à Juniperus procumbens de l’île ouest-japonaise de Tsushima (carte 1, rectangle vert) très proche de la Corée du Sud sud-orientale (colonnes 12 et 13 de notre tableau 12), dont on peut dénommer le plus diversifié (colonne 13) sous le nom de Trachelospermo asiatici-Juniperetum procumbentis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 6 du tableau 10 in Itow et al. (1986, Vegetation of Kuroshima Island, Tsushima… : 21). On peut estimer que l’ensemble des colonnes 11 à 13 possède la valeur d’une alliance de fourrés nippo-coréens notamment caractérisée par Juniperus procumbens, Rhaphiolepis umbellata (photo 31), Paederia foetida (photo 88), Rosa lucieae, Euonymus japonicus (photo 68), Ligustrum japonicum, le Rhaphiolepido umbellatae-Juniperion procumbentis all. nov. hoc loco, typus nominis hoc loco : Trachelospermo asiatici-Juniperetum procumbentis nov.

5. La végétation des prés salés
Durant ce voyage, nous avons laissé de côté la végétation halophile colonisant les vases salées, mais on peut s’appuyer sur des publications antérieures pour en présenter quelques aspects.
En ce qui concerne la végétation thérophytique tout d’abord, la végétation décrite du Japon par Umezu (1964) puis Miyawaki et Ohba (1969) a été incluse au sein de la large classe des Thero-Suaedetea splendentis Rivas Mart. 1972 dans une synthèse récente (de Foucault & Guitton, 2021), laquelle a donné lieu à la définition d’une alliance originale, l’Atriplici gmelinii-Suaedion japonicae B. Foucault & Guitton 2021, caractérisée surtout par Suaeda japonica, Atriplex gmelinii, Limonium tetragonum (indiqué comme bisannuel par Ohwi, 1965) et incluant les Suaedetum japonicae A. Miyaw. & Ohba 1969, Atriplici gmelinii-Suaedetum maritimae Umezu ex A. Miyaw. & Ohba 1969 et Limonietum tetragoni Umezu ex A. Miyaw. & Ohba 1969. D’après les données apportées par C.-S. Kim & Byung (1988, tableau 2), Yang (1999), puis Min & Je (2002), on peut étendre l’alliance et les deux premières associations à la Corée du Sud.
Malgré la présence ponctuelle de Tripolium pannonicum et Triglochin maritima, la végétation halophile vivace ne semble pas devoir se rattacher aux Juncetea maritimi Tüxen & Oberd. 1958 (de Foucault, 2021b). Du sud-ouest de la péninsule coréenne, C.-S. Kim et Byung (1988) définissent les Zoysietalia sinicae et Zoysion sinicae, mais ces noms sont invalides, car aucun type n’est désigné pour l’alliance, ce qui rend l’ordre lui-même invalide. On les valide ici : Zoysion sinicae C.-S. Kim & Byung ex B. Foucault, C.-S. Kim & Byung all. nov. hoc loco, caractérisée au moins par Zoysia sinica et Artemisia scoparia (tableau 13 : colonnes 4 à 6), typus nominis hoc loco : Artemisietum scopariae C.-S. Kim & Byung ex B. Foucault, C.-S. Kim & Byung ass. nov. hoc loco [syn. : Artemisietum scopariae C.-S. Kim & Byung 1988 (Korean J. Ecol. 11 (4) : 182) nom. inval.] dont on désigne aussi ici le typus nominis hoc loco : relevé 1-55 du tableau 4 in C.-S. Kim & Byung (1988, Korean J. Ecol. 11 (4) : 181) ; Zoysietalia sinicae C.-S. Kim & Byung ex B. Foucault, C.-S. Kim & Byung ord. nov. hoc loco, caractérisée au moins par Zoysia sinica et Artemisia scoparia (tableau 13 : colonnes 1 à 6), typus nominis hoc loco : Zoysion sinicae C.-S. Kim & Byung ex B. Foucault, C.-S. Kim & Byung nov. On reconnaît cette végétation aussi chez Min & Je (2002). Les colonnes 1 à 3 se distinguent par une présence significative d’Hololeion maximowiczii. Enfin l’ensemble des colonnes 7 à 13 de ce même tableau paraît ne pas relever des unités précédentes. Les treize syntaxons rassemblés dans le tableau 13 sont pour une partie non nommés ; seuls sont à signaler le Zoysietum sinicae Ohba, A. Miyaw. & Tüxen 1973 en colonne 4, l’Artemisietum scopariae C.-S. Kim & Byung ex B. Foucault, C.-S. Kim & Byung nov. en colonnes 5 et 6, le Triglochinetum maritimae C.-S. Kim & Byung 1988 nom. inval. en colonne 9, fort peu caractérisé, le Caricetum scabrifoliae A. Miyaw. & Ohba 1969 en colonne 10 et le Phaceluretum angustifolii C.-S. Kim & Byung 1988 nom. inval. en colonne 12.
6. La végétation des bas-marais
Nous n’avons pas eu l’occasion non plus de parcourir des bas-marais, mais, là aussi, on peut s’appuyer sur des publications antérieures pour en présenter quelques aspects.
Ainsi J.-W. Kim et J.-H. Kim (2003) présentent quelques résultats d’une étude sur un marais du sud-est du pays et décrivent surtout le Platanthero-Molinietum japonicae, mais sans désigner de type ; on peut valider cette association en Platanthero mandarinorum-Moliniopsietum japonicae J.-W. Kim & J.-H. Kim ex B. Foucault, J.-W. Kim & J.-H. Kim ass. nov. hoc loco, typus nominis hoc loco : relevé 16 du tableau 1 in J.-W. Kim & J.-H. Kim (2003, Korean J. Ecol. 26 (5) : 283). À côté de cette association, d’autres syntaxons à Moliniopsis japonica sont rapportés par les mêmes auteurs, de sorte qu’on peut définir une alliance de bas-marais oligotrophiles coréens, le Miscantho sinensis-Moliniopsion japonicae all. nov. hoc loco, caractérisée par Moliniopsis japonica (vicariant de Molinia caerulea), Miscanthus sinensis, Parnassia palustris, Hololeion maximowiczii, Euphorbia pekinensis, Carex dickinsii, typus nominis hoc loco : Platanthero mandarinorum-Moliniopsietum japonicae J.-W. Kim & J.-H. Kim ex B. Foucault, J.-W. Kim & J.-H. Kim nov. Une végétation à Moliniopsis japonica est aussi rapportée par J.-W. Kim & Han (2005).
J.-W. Kim et J.-H. Kim (2003) décrivent aussi des végétations arborescentes, l’une réduite à Alnus japonica (relevés 27 et 28 de leur tableau 1) et l’autre plus riche en taxons (relevés 35 à 39) : Quercus serrata V, Pinus densiflora IV, Quercus dentata IV, Q. mongolica IV, Q. acutissima III, Prunus padus III, Platycarya strobilacea III, Lindera glauca III, qui peut se placer dans les Carpino laxiflorae-Quercetalia mongolicae définis plus haut.
7. La végétation liée aux activités humaines
7.1. Un voile thérophytique à Draba nemorosa
Assez souvent en plaine, notamment lors de randonnées en ville, on peut remarquer une végétation thérophytique formant un voile (selon l’expression de Géhu & Géhu-Franck, 1985) en mosaïque avec la végétation vivace plus ou moins anthropophile, voire même artificielle, caractérisée par Draba nemorosa (photo 89), Trigonotis peduncularis (photo 91) et Capsella bursa-pastoris. Par leur port, les Trigonotis rappellent beaucoup les Myosotis d’Europe, mais les nucules sont tétraédriques et non ovoïdes, d’où le nom du genre. On peut parfois y observer aussi Androsace umbellata (photo 90). Le tableau 14 rapporte huit relevés (mais on aurait pu en ramener bien d’autres) de cette végétation originale que l’on peut dénommer Trigonotido peduncularis-Drabetum nemorosae ass. nov. hoc loco, typus nominis hoc loco : relevé 2 du tableau 14 hoc loco (photo 92).




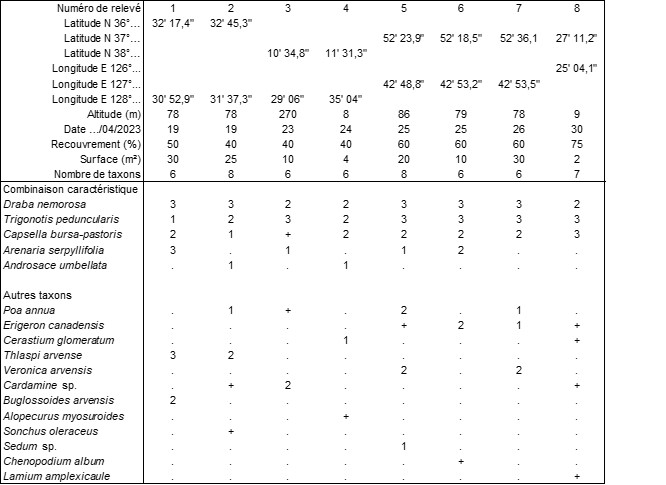
Annexe du tableau 14 – rel. 1 : Hahoe Village (prov. de Gyeongsangbuk-do), bord de la rivière ; rel. 2 : id. 1, près du parking du musée des masques ; rel. 3 : Sokcho (prov. de Gangwon-do), Seoraksan National Park, près du Sinheungsa Temple ; rel. 4 : id. 3, vers Ekseupo Convenience Store ; rel. 5 : Chuncheon (prov. de Gangwon-do), parc Gongjicheon, nord Jongaggang-won ; rel. 6 : id. rel. 5, partie sud du parc, au-dessus de la rivière ; rel. 7 : id. rel. 5, vers Gongji-ro 482beon-gil ; rel. 8 : Incheon, bord de rizière non loin de l’hôtel Ora.
7.2. La végétation commensale des rizières
La végétation commensale des cultures de riz a fait aussi l’objet d’une synthèse récente (de Foucault, 2022). Dans l’une des alliances, le Ludwigio prostratae-Echinochloion hispidulae B. Foucault 2022, il faut ajouter l’Eclipto prostratae-Cardaminetum flexuosae ass. nov. hoc loco (Song, 1997, tableau 1 : relevés 49 à 84, sub Sagittario pygmaeae-Monochorietum plantagineae ; colonne LEh5 dans notre tableau 15), typus nominis hoc loco : composante thérophytique du relevé 64 du tableau 1 in Song (1997, Korean J. Ecol. 20 (3) : 194).
Toujpurs dans le tableau 15, les colonnes Aa1 à Aa3 synthétisent trois syntaxons se rattachant à l’Alopecurion amurensis A. Miyaw. & Okuda 1972 (Vegetatio 24 (4-6) : 247) non considéré dans la synthèse de 2022 et typifié par l’Alopecuro amurensis–Ranunculetum scelerati A. Miyaw. & Okuda 1972 (Vegetatio 24 (4-6) : 246). Ces trois associations sont l’Hemistepto lyratae-Capselletum bursae-pastoris J.-W. Kim & Nam 1998 (J.-W. Kim & Nam, 1998, tableau 3 ; Aa1) muté en Saussureo lyratae-Capselletum bursae-pastoris nom. mut. hoc loco, le Stellario undulatae-Alopecuretum amurensis J.-W. Kim & Nam 1998 (J.-W. Kim & Nam, 1998, tableau 2 ; Aa2), muté en Stellario alsines-Alopecuretum aequalis nom. mut. hoc loco, et l’Alopecuro–Ranunculetum scelerati A. Miyaw. & Okuda 1972 (Miyawaki & Okuda, 1972, tableau 6 ; Aa3). D’après le même tableau 15, cette alliance ne peut se placer dans les Cypero difformis-Echinochloetalia oryzoidis, il est nécessaire de définir un ordre nouveau : les Murdannio keisak-Alopecuretalia aequalis ord. nov. hoc loco, typus nominis hoc loco : Alopecurion amurensis A. Miyaw. & Okuda 1972 (Vegetatio 24 (4-6) : 247) muté ici en Alopecurion aequalis, dont les taxons caractéristiques coïncident avec ceux qui caractérisent l’alliance, dont Murdannia keizak, Alopecurus aequalis (incl. subsp. amurensis), Cardamine flexuosa, Rorippa palustris, R. cantoniensis, Saussurea lyrata. Sur les trois associations citées pour l’Alopecurion aequalis, voir aussi S.-Y. Kim et al. (2019).
Il faut ajouter que, si l’on ne tient pas compte de la graminée cultivée Oryza sativa comme on devrait le faire en phytosociologie moderne, ces deux ordres devraient relever de deux classes distinctes.
8. Conclusion
Malgré la date assez peu favorable à une observation optimale de la flore, ce voyage a donc été une belle occasion de découvrir les forêts coréennes et leurs lisières, la végétation littorale et anthropique. Une abondante bibliographie a par ailleurs permis d’élargir ce compte rendu aux prés salés, bas-marais et rizières. De nombreux syntaxons de rang divers ont été validés ou décrits à cette occasion.
Dans ces milieux naturels, on peut observer plusieurs taxons cultivés ou naturalisés en Europe, dont Abies koreana (photo 43), Acer palmatum (photo 93), A. tatarica subsp. ginnala (photo 15), Akebia quinata (photo 24), Aucuba japonica (à Beomeosa ; photo 94), Camellia japonica (photo 17), Cyrtomium falcatum (photo 95), Euonymus alatus (photo 20), Fatsia japonica (photo 96), Forsythia sp. (trois espèces en Corée du Sud ; photo 97), Kerria japonica (photo 98), Lonicera japonica, Ophiopogon japonicus, Pittosporum tobira (photo 67), Trachelospermum asiaticum (photo 47), Weigela florida (photo 29), Zelkova serrata (photo 99)… C’est assez dire le grand intérêt horticole qu’ont trouvé les premiers voyageurs botanistes en parcourant l’Extrême-Orient à partir du xviie siècle…







Toutes les plantes recontrées lors de ce périple ne sont évidemment pas reprises ici. Pour achever ce compte rendu, les photos 100 à 110 pourront encore faire rêver certains de nos lecteurs.











Bibliographie
Černý T., Kopecký M., Petřik P., Song J.-S., Šrůtek M., Valachovič M., Altman J. & Doležal J., 2014. Classification of Korean forests: patterns along geographic and environmental gradients. Applied Vegetation Science, https://doi.org/10.1111/avsc.12124.
Chang C.-S., Kim H. & Chang K.-S., 2014. Provisional checklist of vascular plants fo he Korea peninsula flora (KPF). Designpost, 660 p.
Cho H.-J. & Bae K.-H., 2001. Species composition and structure of the Oriental Arbor-Vitae (Thuja orientalis L.) forest in Daegu, Southeastern Korea. Korean Journal of Ecology 24 (4) : 205-211.
Foucault B. (de), 1987. Nouvelles recherches sur les structures systématiques végétales : caractérisation, ordination, signification. Phytocoenologia 15 (2) : 159-199.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Betulo carpaticae-Alnetea viridis Rejmánek in Huml, Lepš, Prach & Rejmánek 1979. Journal de botanique de la Société botanique de France 60 : 47-68.
Foucault B. (de), 2016. Contribution au prodrome des végétations de France : les Arrhenatheretea elatioris Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952. Documents phytosociologiques, 3e série, 3 : 1-219.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021a. Contribution à une synthèse des classes des Honckenyo peploidis-Leymetea arenarii et des Euphorbio paraliae-Ammophiletea arundinaceae. Carnets botaniques 64 : 1-22 + 16 tableaux, https://doi.org/10.34971/4ff3-vd67.
Foucault B. (de), 2021b. Contribution à une synthèse des Juncetea maritimi. Carnets botaniques 68 : 1-30 + 22 tableaux, https://doi.org/10.34971/nbat-mh09.
Foucault B. (de), 2022. Compléments aux synthèses des Juncetea bufonii et Crypsietea aculeatae ; essai sur les Oryzetea sativae. Carnets botaniques 107 : 1-10 + 5 tableaux, https://doi.org/10.34971/vgyx-k502.
Foucault B. (de) & Guitton H., 2021. Une déclinaison moderne des Thero-Suaedetea splendentis. Carnets botaniques 76 : 1-9 + 2 tableaux, https://doi.org/10.34971/3XK3-N640.
Géhu J.-M. & Géhu-Franck J., 1985. Les voiles nitrophiles annuels des dunes armoricaines anthropisées. Colloques phytosociologiques XII, Les végétations nitrophiles et anthropogènes : 1-21.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Itow S., Nakanishi H., Kawasato H. & Chichibu Y., 1986. Vegetation of Kuroshima Island, Tsushima, western Japan. Provided by Nagasaki university’s Academic OUTPUT site, https://core.ac.uk/reader/483769092 (en japonais).
Jung Y.-K., 2000a. The Viticetea rotundifoliae in South Korea and Japan. Korean Journal of Ecology 23 (5) : 383-389 (en coréen).
Jung Y.-K., 2000b. Rosetalia rugosae and Viticetea rotundifoliae in South Korea. Vegetation Science 17 : 39-51.
Jung Y.-K. & Kim J.-W., 1998. Syntaxonomy of mantle communities in South Korea. Korean Journal of Ecology 21 (6) : 739-750 (en coréen).
Jung Y.-K. & Kim W., 2001. Syntaxonomical reconsideration on the Rosetalia rugosae. Korean Journal of Ecology 24 (5) : 267-271 (en coréen).
Kim C.-H., Kang S.-H. & Kil B.-S., 1991. The vegetation of Mt. Choksang. Korean Journal of Ecology 14 (2) : 137-148 (en coréen).
Kim C.-S. & Byung S.-I., 1988. Studies on the vegetation of the salt marsh in the southwestern coast of Korea. Korean Journal of Ecology 11 (4) : 175-192 (en coréen).
Kim C.-S. & Jang Y.-S., 1989. Phytosociological study on the vegetation od Daehŭksan Island. Korean Journal of Ecology 12 (3) : 145-160 (en coréen).
Kim C.-S. & Oh Y.-S., 1990. Phytosociological study on the evergreen broad-leaved forests in Dadohae National Marine Park, the vegetation of Kŭmo Archipelago. Korean Journal of Ecology 13 (4) : 343-359 (en coréen).
Kim I.-T., Park T.-H. & Choi J.-E., 2005. An ecological study on the vegetation of Bijin and Yongcho Islets. Korean Journal of Ecology 28 (4) : 223-230 (en coréen).
Kim J.-U. & Yim Y.-J., 1986. Classification of forest vegetation of Seonunsan area, Southwestern Korea. Korean Journal of Ecology 9 (4) : 209-223 (en coréen).
Kim J.-U. & Yim Y.-J., 1993. Classification and multidimensional analysis of plant communities in Mt. Moak Provincial Park, Korea. Korean Journal of Ecology 16 (1) : 1-15 (en coréen).
Kim J.-W., Song S.-D. & Kim S.-J., 1996. A syntaxonomical study on the vegetation of Ulreung-do and Tok-do, Korea. Report of the Survey of Natural Environment in Korea 10 : 137-202 (en coréen).
Kim J.-W. & Nam H.-K., 1998. Syntaxonomical and synecological characteristics of rice field vegetation. Korean Journal of Ecology 21 (3) : 203-215 (en coréen).
Kim J.-W. & Kim J.-H., 2003. Vegetation of Moojechi moor in Ulsan: syntaxonomy and syndynamics. Korean Journal of Ecology 26 (5) : 281-287 (en coréen).
Kim J.-W. & Han S.-U., 2005. Moor vegetation of Mt. Shinbul in Yangsan. Korean Journal of Ecology 28 (2) : 85-92 (en coréen).
Kim M.-H., Yoshikawa M. & Hukusima T., 1999. Studies on the floristic composition and succession of the shrub communities at the summit of M. Halla, Cheju Island, Korea. Korean Journal of Ecology 22 (6) : 325-335.
Kim S.-Y., Kim M.-S., Ryu Y.-M. & An S.-L., 2019. A phytosociological study of spring-type rice field vegetation in Angye Plains, South Korea. Journal of Asia-Pacific Biodiversity 12 : 661-667.
Kolbek J., Jarolímek I. & Valachovič M., 2003. Forest vegetation of the Northern Korean Peninsula. Geobotany 28, Forest Vegetation of Notheast Asia : 263-361, https://doi.org/10.1007/978-94-017-0143-3_8.
Kwak S.-H., Chom H.-Y., Kim C.-H. & Kil B.-S., 1991. The vegetation of Pyŏnsan Peninsula National Park, Buan. Korean Journal of Ecology 14 (2) : 181-194 (en coréen).
Lee C.-S. & Cho H.-J., 1993. Structure and dynamics of Abies koreana Wilson community in Mt. Gaya. Korean Journal of Ecology 16 (1) : 75-91 (en coréen).
Lee H.-J., Kim H.-S. & Cho E.-B., 1991. Phytosociological study of the forest vegetation in Mt. Kŏmdan. Korean Journal of Ecology 14 (3) : 273-303 (en coréen).
Lee H.-J., Choung H.-L., Byun D.-W. & Kim C.-H., 1993. Analysis of the forest vegetation of Mt. Ilwol. Korean Journal of Ecology 16 (3) : 239-259 (en coréen).
Lee H.-J., Kim J.-H., Kang J.-G., Chun Y.-M. & Byung H.-B., 1995a. Forest vegetation and soil environment on Mt. Mohu. Korean Journal of Ecology 18 (3) : 367-383 (en coréen).
Lee H.-J., Choung H.-L. & Bae B.-H., 1995b. Syntaxonomy and analysis of interspecific association on the foret vegetation of Mt. Ch’ŏngnyang. Korean Journal of Ecology 18 (1) : 121-136 (en coréen).
Lee J.-S., Kim H.-K. & Song J.-S., 2005. A phytosociological study of the Quercus spp. Forests in the lower montane zone o middle and northern parts, Chungbuk Province, Korea. Korean Journal of Ecology 28 (4) : 207-214 (en coréen).
Lee J.-S., Ihm B.-S., Cho D.-S. & Kim J.-W., 2007.Coastal dune vegetation of South Korea. Journal of Ecology and Field Biology 30 (2) : 135-142.
Lee O.-J., 1981. A study on the vegetation of the Saeng-il Island. Korean Journal of Ecology 4 (3-4) : 80-92.
Min B.-M. & Je J.-G., 2002. Typical coastal vegetation of Korea. Ocean and Polar Research 24 (1) : 79-86.
Miyawaki A. & Ohba T., 1969. Studien über Strand-Salzwiesengesellschaften auf Honshu, Shikoku and Kyushu (Japan). Science Reports of the Yokohama National University, sect. II, 15 : 1-23.
Miyawaki A. & Okuda S., 1972. Pflanzensoziologische Untersuchungen über die Auen-Vegetation des Flusses Tama bei Tokyo, mit einer vergleichenden Betrachtung über die Vegetation des Flusses Tone. Vegetatio 24 (4-6) : 229-311.
Murakami Y., 1991. Mantle communities in the Nansei Islands, southwestern Japan. Japanese Journal of Ecology 41 : 191-208.
Nakanishi H., 1980. Ecological and phytosociological studies on Crinum asiaticum var. japonicum community. Japanese Journal of Ecology 30 : 251-257.
Ohba T., Miyawaki A. & Tüxen R., 1973. Pflanzengesellschaften der japanischen Dünen-Küsten. Vegetatio 26 (1-3) : 3-143.
Ohwi J., 1965. Flora of Japan. Smithsonian Institution, Washington, 1 067 p.
Song J.-S., 1997. A phytosociological study on the weed communities in the cultivated and abandoned fields of Korea. Korean Journal of Ecology 20 (3) : 191-200 (en coréen).
Song J.-S. & Nakanishi S., 1985. Phytosociological study of the subalpine forests on Mt. Halla of Cheju Island, Korea. Japanese Journal of Ecology 35 : 317-328.
Song J.-S., Song S.-D., Park J.-H., Seo B.-B., Chung H.-S., Roh K.-S. & Kim I.-S., 1995. A phytosociological study of Quercus mongolica forest of Mt. Sobaek by ordination and classification techniques. Korean Journal of Ecology 18 (1) : 63-87 (en coréen).
Sun B.-Y., Kim M. H., Kim C. H. & Park C.-W., 2001. Mankyua (Ophioglossaceae): a new fern genus from Cheju Island, Korea. Taxon 50 : 1019-1024.
Takhtajan A., 1986. Floristic regions of the world. University of California Press, Berkeley, Los Angeles & London, 522 p.
Tanaka T. & Ito K., 1981. Phytosociological studies of the communities with Ophiopogon japonicus var. caespitosus. Japanese Journal of Ecology 31 : 1-11.
Tüxen R., 1966. Über nitrophile Elymus-Gesellschaften an nordeuropäischen, nordjapanischen und nordamerikanischen Küsten. Annales Botanici Fennici 3 (3) : 358-367.
Umezu Y., 1964. Über die Salzwasserpflanzengesellschaften in der Nähe von Yukuasi, Nordkyûsyû, Japan. Japanese Journal of Ecology 14 : 153-160.
Yang H.S., 1999. A syntaxonomical study o the vegetation of ruined salt field in Chonnam Province. Korean Journal of Ecology 22 (5) : 265-270 (en coréen).
Yim Y.J., Kim J.U., Lee N.J., Kim Y.B. & Paek K.S., 1990. Phytosociological classification of plant communities of Mt. Halla National Park, Korea. Korean Journal of Ecology 13 (2) : 101-130 (en coréen).
Remerciements
Ils s’adressent chaleureusement à Chin Sung Chang, botaniste au T.B. Lee Herbarium (College of Agriculture and Life Sciences, Seoul National University), qui a apporté sa contribution à ce compte rendu en donnant les noms de plusieurs des taxons cités ici.


