
Nouvelles perspectives phytosociologiques sur les pinèdes des Erico-Pinetea Horvat 1959
Title
New phytosociological perspectives on the pine forests of Erico-Pinetea Horvat 1959
Résumé
Cette monographie phytosociologique est consacrée aux pinèdes à Erica carnea revues selon le paradigme synusial ; elle a permis de reconnaître une cinquantaine d’associations originales replacées dans neuf alliances, six plutôt basiphiles, les trois autres acidiphiles venant s’ajouter au classique Ericion carneae dans les Kalmio-Vaccinietea microphylli.
Abstract
This phytosociological monograph is devoted to Erica carnea pine forests reviewed according to the synusial paradigm; it has enabled the recognition of some fifty original associations linked to nine alliances, six of which are mainly basiphilic and three acidiphilic, in addition to the classic Ericion carneae within the Kalmio-Vaccinietea microphylli.
1. Introduction
La classe des Erico-Pinetea sylvestris rassemble les pinèdes (à Pinus sylvestris, P. uncinata, P. nigra subsp. n.) thermoxérophiles des étages montagnards à subalpins d’Europe centrale développées sur des sols calcaires plus ou moins évolués, éventuellement acidifiés. Cette classe a été sortie des Vaccinio-Piceetea abietis Braun-Blanq. in Braun-Blanq. G. Sissingh & J. Vlieger (1939) par Horvat (1959, Biološki Glasnik 12 : 25). Elle sera l’objet du présent article à la lumière du paradigme synusial. On n’y inclura pas les données de Vanden Berghen (1963) sur les pinèdes des Causses, qui n’en relèvent pas, ni celles de Thévenin & Royer (2001, Pyrolo chloranthae-Pinetum), qui sont problématiques dans la mesure où la strate arborescente associant Pinus sylvestris et P. nigra subsp. n. résulte de plantations initiales entreprises au xxe siècle et donc à caractère plutôt artificiel (de Foucault, 2020 : 7). Globalement, cette végétation est considérée comme d’intérêt communautaire (inscription à l’annexe I de la directive Habitats-Faune-Flore de 1992) en tant que partie des « Forêts à Pinus uncinata » (code Corine-biotopes 42.42, code Eur 27 9430, prioritaire sur substrat gypseux ou calcaire ; Rameau, 2001).
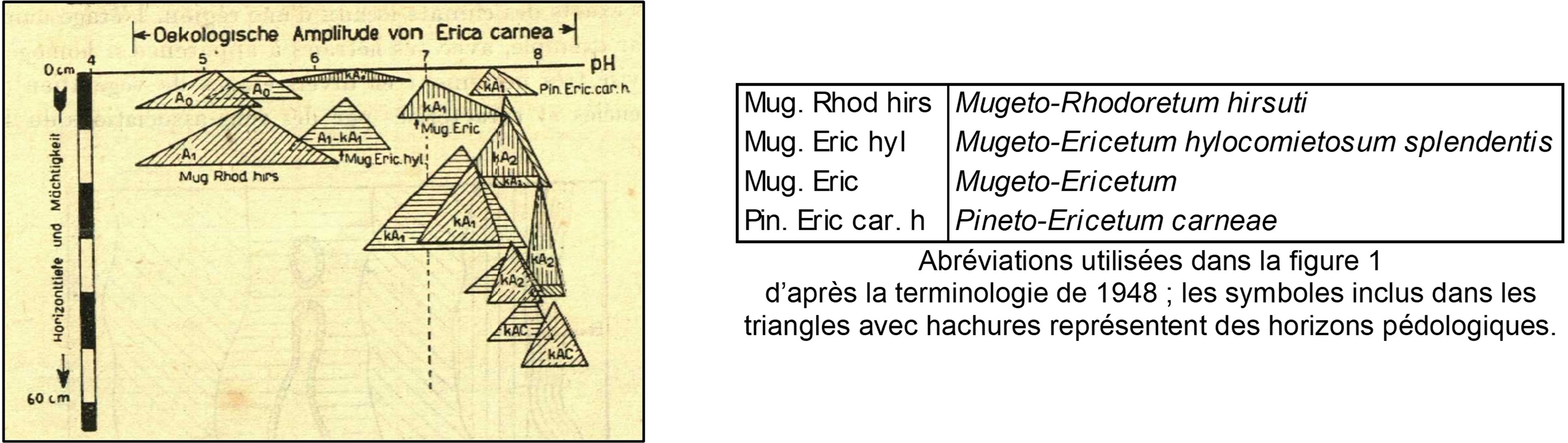
D’une manière générale, la végétation arborescente est le plus souvent caractérisée par des pins spontanés, Pinus sylvestris, mais aussi P. nigra subsp. n., parfois Picea abies, alors que la végétation basse du sous-bois mêle des cryptophytes (hémicryptophytes et géophytes) et des chaméphytes, surtout Erica carnea [syn. : E. herbacea ; photo 1] et Polygala chamaebuxus (Chamaebuxus alpestris dans POWO, référentiel nomenclatural suivi dans cet article malgré parfois son retard sur la taxonomie et la nomenclature actuelles ; photo 2). On peut d’ores et déjà préciser qu’E. carnea a une large amplitude édaphique (figure 1) ; ce taxon ne peut donc être utilisé seul pour caractériser cette végétation.
On commencera par détailler les données phytosociologiques rassemblées par Bartoli (1966) en Haute-Maurienne, avant d’étendre la synthèse vers l’Europe centrale.


2. La végétation de hêtraies et pinèdes de Haute-Maurienne
On doit effectivement à Bartoli (1966) une belle étude des boisements de Haute-Maurienne, selon le paradigme phytosociologique classique. Il n’a pas toujours été facile de s’y retrouver, car la nomenclature phytosociologique du texte ne correspond pas toujours aux titres des tableaux annexes, en outre ceux-ci ne sont pas appelés par leur numéro dans le texte.
La partie arborescente a été analysée précédemment par le présent auteur (2018) qui y a mis en évidence l’Abieti albae-Fagetum sylvaticae Julve ex B. Foucault 2018, le Pinetum uncinato-sylvestris B. Foucault 2018, le Piceo abietis-Pinetum uncinatae B. Foucault 2018, l’Abieti albae-Piceetum abietis Julve ex B. Foucault 2018.
Le tableau 1 en synthétise la végétation arbustive intraforestière. Les colonnes 1 à 12 relèvent de l’Amelanchiero ovalis-Buxion sempervirentis O. Bolòs & Romo 1989 dans sa sous-alliance Cotoneastro integerrimi-Amelanchierenion ovalis B. Foucault & Julve ex B. Foucault & J.-M. Royer 2016, avec Cotoneaster tomentosus, Berberis vulgaris ; avant toute interprétation syntaxonomique de cette végétation, il conviendrait de revoir précisément la détermination du C. tomentosus à la lumière du traitement actuel du genre. Avec 55 relevés, les colonnes 20 à 25 révèlent un fourré à Lonicera alpigena subsp. a., L. nigra, Rosa pendulina, Clematis alpina subsp. a. (photo 3) pouvant se rattacher au Sorbo mougeotii-Lonicerion alpigenae B. Foucault 2012 (de Foucault, 2012), le Lonicero alpigenae-Rosetum pendulinae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 9 du tableau V in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte). Relevant de la même alliance, mais distinct, les colonnes 26 à 29 décrivent plutôt le Roso pendulinae-Loniceretum caeruleae ass. nov. hoc loco, relié notamment au Pino cembrae-Laricetum deciduae Julve ex B. Foucault 2018, typus nominis hoc loco : composante arbustive du relevé 15 du tableau VIII in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte).

Le tableau 2 porte sur la végétation intraforestière herbacée vivace de ces boisements de Haute-Maurienne. Quatre ensembles majeurs se partagent les vingt-et-une colonnes rassemblées ici. On doit mettre à part la colonne 1 à Trochiscanthes nodiflora (photo 4) qui relève manifestement du Veronico urticifoliae-Trochiscanthion nodiflorae B. Foucault 2020 et qui peut être définie comme Veronico urticifoliae-Trochiscanthetum nodiflorae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 24 du tableau « Hêtraies » in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte, en remplaçant Veronica latifolia par V. urticifolia).
Caractérisé par le groupe sociologique (GS) à Melampyrum nemorosum subsp. catalaunicum, dans l’ensemble formé par les colonnes 2 à 6, on peut au moins distinguer
- en 2, l’Astragalo monspessulani-Polygaletum chamaebuxi nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 4 du tableau « Hêtraies » in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte) ;
- en 3, le Cephalanthero rubrae-Polygaletum chamaebuxi nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 11 du tableau « Hêtraies » in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte) ;
- en 5, l’Aquilegio atratae-Prenanthetum purpureae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 21 du tableau « Hêtraies » in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte, en remplaçant Aquilegia atroviolacea par A. atrata).
La colonne 6 ne paraît guère floristiquement homogène, trop de liens apparaissant avec les deuxième et troisième ensembles (colonnes 7 à 12 et 13 à 17).

Le deuxième ensemble est surtout caractérisé par le GS à Globularia cordifolia, avec
- en 7, l’Asperulo oreophilae-Helianthemetum cani nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 2 du tableau I in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte, en précisant Asperula aristata par subsp. oreophila ; photo 5) ;
- en 11, le Thymo polytrichi-Seslerietum caeruleae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 10 du tableau II in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte, en remplaçant Thymus serpyllum ssp. trachs. par Th. polytrichus) ;
- en 12, le Polygalo chamaebuxi-Caricetum ferrugineae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 29 du tableau IV in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte) ; cette association peut être rapprochée du Caricion ferrugineae G. Braun-Blanq. & Braun-Blanq. in G. Braun-Blanq. 1931.

Le troisième ensemble (colonnes 13 à 17) est fortement caractérisé par Erica carnea. Les deux premières colonnes paraissent correspondre à deux variations de l’Oreoselino nigri-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 5 du tableau III in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte, en remplaçant Peucedanum oreoselinum par Oreoselinum nigrum). La colonne 15 peut être reconnue comme Carici albae–Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 19 du tableau III in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte). La colonne 17 sera alors le Valeriano montanae–Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 20 du tableau IV in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte).
Les colonnes 18 à 21 forment un quatrième ensemble caractérisé par le GS à Veronica urticifolia, avec
- en 19, le Festuco flavescentis-Adenostyletum alliariae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 16 du tableau V in Bartoli (1966, Ann. Sci. Forest. XXIII (3) hors texte) ;
- en 20, le Festuco flavescentis-Saxifragetum cuneifoliae ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 24 du tableau V in Bartoli (1966, Sci. Forest. XXIII (3) hors texte), syntaxon paraissant vicariant centre-alpin du Selaginello helveticae-Saxifragetum cuneifoliae B. Foucault 2023 des Alpes-Maritimes (de Foucault, 2023) ; photo 6.
Comme la colonne 6, la colonne 21 ne paraît guère floristiquement homogène, trop de liens apparaissant avec les deuxième et troisième ensembles (colonnes 7 à 12 et 13 à 17).

3. Les Erico-Pinetea Horvat 1959
Ces forêts résineuses de Haute-Maurienne relèvent classiquement des Erico-Pinetea Horvat 1959, à chorologie optimale centre- et est-alpine. Pourtant la synthèse de cette classe par Hölzel (1996a) pour l’Allemagne ne cite pas le travail de Bartoli, n’évoquant pour la France que l’« Onobrychido saxatilis-Pinetum sylvestris » Braun-Blanq. 1961 du Briançonnais. Ces habitats sont cités par Villaret et al. (2019 : 392 et 506). Dans cette partie, on va donc tenter d’intégrer les acquis ouest-alpins de la partie 2 dans cet ensemble centre- et est-alpin.
Pour la végétation arborescente, le tableau 3 met en évidence le Piceo abietis-Pinetum sylvestris Roulier ex B. Foucault 2018 (colonnes 1 à 6). Les colonnes 7 et 8 peuvent se rattacher à l’Abieti albae-Fagetum sylvaticae Julve ex B. Foucault 2018 dans une variation à Pinus sylvestris, la colonne 9 au Piceo abietis-Alnetum incanae Roulier ex Roulier& B. Foucault in B. Foucault 2020, la 13 au Querco pubescentis-Pinetum nigrae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 4 du tableau d’association 4 in Wagner (1941, Denksch. Akad. Wiss. Wien, Math.-Naturwiss. Kl. 104 : 63-65). Les colonnes 14 et 15 correspondent au Fraxino orni-Pinetum nigrae Martin-Bosse 1967 (Martin-Bosse, 1967). Les colonnes 16 à 21 décrivent le Pinetum sylvestri-nigrae Pawł. 1958 (apud Horvat, 1959). Les colonnes 22 à 24 sont interprétable comme Ostryo carpinifoliae-Fraxinetum orni Aichinger 1933. La colonne 25 est originale, décrivant un mélézin à Ostrya carpinifolia, Fraxinus ornus, Laburnum alpinum : Ostryo carpinifoliae-Laricetum deciduae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (E3) du relevé 1 du tableau 24 in Dakskobler et al. (2018, Hacquetia 17 (2) : 494-501). Enfin, la colonne 26 est un mélézin assez pauvre, d’ailleurs distinct du Pino cembrae-Laricetum deciduae Julve ex B. Foucault 2018 ouest-alpin, le Piceo abietis-Laricetum deciduae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (E3) du relevé 1 du tableau 1 in Dakskobler et al. (2018, Hacquetia 17 (2) : 268-277).
Le tableau 4 synthétise la végétation arbustive intraforestière. Les colonnes 1 à 11 relèvent à nouveau du Cotoneastro integerrimi-Amelanchierenion ovalis, où l’on peut définir pour les colonnes 1 à 9 le Rhamno saxatilis-Amelanchieretum ovalis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (Straucher) du relevé 238 du tableau 1 in Hölzel (1996b, Laufener Forsch. 3 : 171-172), et pour la 11 le Salici glabrae-Amelanchieretum ovalis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 1 in Dakskobler (1999, Wiss. Mitteil. Niederosterreichischen Landesmuseums 12 : 38-43) ; on retrouve cette association aussi bien exprimée in Rozman et al. (2020). La colonne 10 est peu distincte du Cotoneastro integerrimi-Amelanchieretuum ovalis Faber ex Korneck 1974 (ou race à Euonymus verrucosus ?). Les colonnes 12 à 18 relèvent plutôt du Berberidion vulgaris Braun-Blanq. ex Tüxen 1952.
Les colonnes 19 à 23 sont des brousses à Pinus mugo. Dans une publication de 2021, sur la base de son tableau 5, j’ai tenté une petite synthèse de ce type d’habitat ; dans le tableau 5 hoc loco, j’ai élargi celui de 2021 en y intégrant les colonnes 19 à 23 du tableau 4, ainsi que les colonnes 2 (empruntée à Dakskobler, et al., 2018 : tableau 24) et 8 à 10 (empruntées à Starlinger, 1992 : tableau 1). La colonne 2, reliée à l’Ostryo carpinifoliae-Laricetum deciduae arborescent, s’avère un intermédiaire entre le Cotoneastro integerrimi-Amelanchierenion ovalis et les Betulo-Alnetea alnobetulae : Salici appendiculatae-Amelanchieretum ovalis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 1 du tableau 24 in Dakskobler et al. (2018, Hacquetia 17 (2) : 494-501) ; plusieurs formes juvéniles des arbres potentiels y apparaissent (Larix decidua, Picea abies, Laburnum alpinum, Sorbus aucuparia subsp. a., S. aria, Fagus sylvatica, Abies alba, Ostrya carpinifolia, Fraxinus ornus).
L’ensemble des colonnes 8 à 12 peut être décrit comme Junipero communis-Rosetum pendulinae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 19 du tableau 1 in Starlinger (1992, Tuexenia 12 hors texte). Les colonnes 14 et 15 (et sans doute aussi la 13 en tant que variation) décrivent le Salici chamaemespili–Pinetum mugo ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 93 du tableau 3 in Maier (1994, Stapfia 35 : 111-114). Non loin de ces syntaxons, mais absent du tableau 5, on peut encore citer à titre informatif le Lonicero borbasianae-Pinetum mugo Horvat 1938 (Fukarek & Stefanovič, 1958 ; Šibik et al., 2010), dont la colonne 16 du tableau 5 donne un aperçu.
Pour rassembler ces brousses à Pinus mugo, a été créée la classe des Roso pendulinae-Pinetea mugo Theurillat in Theureillat et al. 1995 (Colloq. Phytosoc. XXIII : 228) ; cependant, comme le montre le tableau 5, cette végétation partage suffisamment de taxons avec celle des Betulo-Alnetea alnobetulae Rejmánek in Huml et al. 1979 pour qu’elle y soit incluse en tant qu’alliance du Pinion mugo Pawł. in Pawł. et al. 1928.
Dans ses colonnes 4, 6 à 13, 15 à 17, 28 à 33 et 42, le tableau 6 reprend les données de Hölzel (1996a), y ajoutant en tant que syntaxons 1 à 3 et 41 les données issues du tableau 2 de Haute-Maurienne (hors colonnes 1 et 6 pour le syntaxon 1, colonne 12 pour le syntaxon 2, colonne 21 pour le syntaxon 39).
Une coupure majeure passe entre les syntaxons 32-33, les syntaxons 1 à 29 s’avérant le plus souvent xérothermophiles calcicoles. Des coupures de moindre niveau passent entre les syntaxons 5-6, 15-16, 17-18 et 27-28.
Le premier ensemble (syntaxons 1 à 3) correspond à la végétation calcicole de Haute-Maurienne et montre que ces trois syntaxons ont chacun valeur d’alliance originale :
- le syntaxon 1 peut être dénommé Melampyro catalaunici-Polygalion chamaebuxi nov. hoc loco, caractérisé essentiellement par une combinaison de Melampyrum nemorosum subsp. catalaunicum, Prenanthes purpurea, Phyteuma spicatum, Euphorbia dulcis s. l., Luzula nivea, Aquilegia atrata (photo 7), Lathyrus vernus, Tanacetum corymbosum, Carex digitata, Cephalanthera longifolia, C. rubra (photo 8), Polygonatum verticillatum, Lilium martagon, Digitalis lutea, typus nominis hoc loco: le Cephalanthero rubrae-Polygaletum chamaebuxi nov. ;
- le syntaxon 2 peut être dénommé Galio corrudifolii-Arctostaphylion uvae-ursi nov. hoc loco, caractérisé essentiellement par une combinaison de Galium corrudifolium, Laserpitium siler, Coronilla minima subsp. m., Helianthemum canum, Hieracium lanatum, Anthericum liliago, Thymus polytrichus, Viola saxatilis (photo 9), Achnatherum calamagrostis, Asperula aristata subsp. oreophila, typus nominis hoc loco: l’Asperulo oreophilae-Helianthemetum cani nov. ;
- le syntaxon 3 peut être dénommé Carici albae-Ericion carneae nov. hoc loco, caractérisé essentiellement par une combinaison de Erica carnea, Luzula sylvatica subsp. sieberi, Vaccinium vitis-idaea, Orthilia secunda, Melampyrum sylvaticum, Rosa pimpinellifolia, Pyrola chlorantha, Antennaria dioica, Plantago maritima subsp. serpentina, partageant Galium corrudifolium et Laserpitium siler avec le précédent, typus nominis hoc loco: l’Oreoselino nigri-Ericetum carneae nov.



On peut rapprocher de ces trois alliances le syntaxon 4 du Briançonnais, décrit par Braun-Blanquet (1961) et repris par Hölzer (1966a) comme « Onobrychido-Pinetum », et 5 du Valais suisse, qui s’avèrent des transitions avec le syntaxon 6 à 15 :
- en 4, l’Onobrychido saxatilis-Ononidetum rotundifoliae ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 7 du tableau 4 in Braun-Blanquet (1961, Selecta 1 hors texte) ; photo 10 ;
- en 5, l’Odontito viscosi-Oreoselinetum nigri ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 8 du tableau 1 in Braun-Blanquet & Richard (1949, Murithienne 64 : 119-120, en changeant Euphrasia viscosa en Odontites viscosus et Peucedanum oreoselinum en Oreoselinum nigrum).

Ces deux associations ne rentrent bien dans aucune des trois alliances de Haute-Maurienne, on peut considérer l’Onobrychido saxatilis-Ononidetum rotundifoliae nov. comme typus nominis hoc loco de l’Odontito viscosi-Arctostaphylion uvae-ursi all. nov. hoc loco, caractérisé par Onobrychis saxatilis (photo 11), Ononis pusilla subsp. p., Odontites viscosus subsp. v. (photo 12), Thymus praecox, Anthyllis montana, Astragalus austriacus, Viola rupestris ; un relevé en est aussi publié par Braun-Blanquet (1922 : 86-87). Ces quatre alliances plutôt ouest-alpines se distinguent assez nettement des syntaxons centre- et est-alpins décrits par Hölzel, ce qui pourrait justifier leur rapprochement dans un ordre ou un sous-ordre. Braun-Blanquet et Richard (1949) rapportent aussi d’autres relevés qui peuvent se rattacher à cet ensemble, par exemple celui de leurs pages 112-113. Par ailleurs, il faudrait vérifier si les communautés pyrénéennes et ibériques à Ononis rotundifolia peuvent aussi s’y rattacher.


Les syntaxons 6 à 15 forment le deuxième grand ensemble de cette synthèse ; la plupart d’entre eux sont originaux, mais il n’est pas toujours possible de les valider, les données détaillées étant souvent inédites ou difficiles d’accès. Le syntaxon 8 peut être défini comme Thymo polytrichi-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 30 du tableau 1 in Starlinger (1992, Tuexenia 12 hors texte). Le syntaxon 14 est défini ici comme Pulsatillo pratensis-Polygaletum chamaebuxi ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 2 du tableau 4 in Wagner (1941, Denksch. Akad. Wiss. Wien, Math.-Naturwiss. Klasse 104 : 63-65, en changeant Anemone nigricans en Pulsatilla pratensis subsp. p.). Le syntaxon 15 peut être nommé Leontodonto incani-Polygaletum chamaebuxi ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 23 du tableau 1 in Hemp (1995, Ber. Bayer. Akad. Natursch. Landsch. 19 hors texte). On peut attribuer à cet ensemble 6 à 15 la valeur d’une alliance richement caractérisée ou différenciée des alliances voisines par Festuca amethystina, Carex ornithopoda subsp. o., C. montana, Campanula cochlearifolia, Potentilla verna, Leontodon incanus, Galium anisophyllum, Thesium rostratum, Th. linophyllon subsp. montanum, Aster amellus, Noccaea montana, Cytisus nigricans…, le Festuco amethystinae-Polygalion chamaebuxi all. nov. hoc loco, typus nominis hoc loco : le Leontodonto incani-Polygaletum chamaebuxi nov. On pourra y rattacher aussi la composante cryptophyto-chaméphytique du « Calamagrostio variae-Pinetum sylvestris » et du « Dorycnio germanici-Pinetum sylvestris » décrits du sud de l’Allemagne par Oberdorfer (1957 : 368-370 et 371-372).
Les syntaxons 16 et 17 sont des Ericeta carneae surtout définis négativement relativement à leurs voisins.
Les syntaxons 18 à 23 (et 24 + 25 du Trentin italien ?), propres aux pinèdes de Pinus nigra subsp. n. de Slovénie, sont aussi très originaux :
- le 18 peut être reconnu comme Laserpitio krapfii-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 3 du tableau de l’appendice 2 in Accetto (1999, Zbornik gozdarstva in lesarstva 60 : 148-151, en remplaçant Erica herbacea par carnea) ;
- le 19 l’est comme Paederoto luteae–Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 3 du tableau 1 in Dakskobler (1999, Mitteil. Niederosterreichischen Landesmuseums 12 : 38-43 ; Paederota lutea est une Plantaginaceae voisine des Veronica, photo 13) ;
- le 20 comme Helleboro nigri-Polygaletum chamaebuxi nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 22 du tableau 1 in Rozman et al. (2020, Hacquetia 19 (1) : 36-51) ;
- le 21 l’est comme Daphno alpinae-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 13 du tableau de l’appendice 2 in Accetto (2001, Zbornik gozdarstva in lesarstva 64 : 37-39, en remplaçant Erica herbacea par carnea) ;
- le 22 est le Cyclamini purpurascentis-Ericetum carneae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 17 du tableau 2 in Rozman et al. (2020, Hacquetia 19 (1) : 52-67) ;
- sur sols acidifiés, le 23 est une transition vers les landes à Vaccinium et Rhododendron hirsutum, mais encore riche en taxons herbacés basiphiles : Valeriano saxatilis-Ericetum carneae nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 32 du tableau 1 in Rozman et al. (2020, Hacquetia 19 (1) : 36-51) ; photo 14.


Les syntaxons 24 à 27 possèdent quelques liens avec les 18 à 23, mais affaiblis ; en 24 : le Chamaecytiso purpurei-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 86 du tableau 1 in Minghetti (1996, Doc. Phytosoc., n. s., XVI hors texte) ; en 25 : le Thliphthiso purpureae-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 24 du tableau 2 in Minghetti (2003, Braun-Blanquetia 33 : 16-17, en remplaçant Asperula purpurea par Thliphthisa p.). La colonne 26 est le Rhododendro hirsuti-Juniperetum alpinae Horvat ex Horvat et al. 1974 (Surina, 2013), la 27 l’Helianthemo nitidi-Ranunculetum scutati Lakušić et al. 1979 nom. inval., validé ici comme Helianthemo glabri-Ranunculetum thorae Lakušić et al. ex ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau XX in Lakušić et al. (1979, Drugi kongres ekologa Jugoslavije 1 : 658). La 28 est l’Aquilegio nigricantis-Rhododendretum hirsuti Lakušić et al. ex Surina 1983. La valeur de cette belle unité slovèno-croate (avec irradiation vers le Trentin ?) en tant qu’alliance est confortée par un cortège floristique caractéristique et différentiel rassemblant Cyclamen purpurascens, Betonica alopecuros, Genista januensis, Gentiana symphyandra, Chamaecytisus hirsutus, Rhodothamnus chamaecistus (Ericaceae ; photo 15), Melampyrum velibiticum, Lotus germanicus, Galium lucidum, Laserpitium siler, L. peucedanoides, Leontodon incanus, Thymus polytrichus, Helleborus niger, Valeriana tripteris, Carex mucronata, Campanula cespitosa, Grafia golaka (Apiaceae), l’Aquilegio nigricantis-Rhododendrion hirsuti Čarni & Mucina 2015 (Hacquetia 14 (2) : 291). Cette alliance n’apparaît toutefois pas absolument homogène, de grands vides floristiques marquant les colonnes 26 à 28, l’une d’elles (la 28) correspondant justement au type de l’alliance. Elle devra être revue sur la base de nouveaux tableaux.

Le cinquième ensemble (syntaxons 29 à 33) est marqué par la grande rareté ou même l’absence des taxons caractérisant les Festuco amethystinae-Polygalion chamaebuxi et Aquilegio nigricantis-Rhododendrion hirsuti. Mais on ne peut typifier ici aucun de ces cinq syntaxons.
L’ensemble des syntaxons 1 à 33, répartis dans quatre sous-ensembles (sans compter le groupe 16-17), dont trois validés comme alliances, est hautement caractérisé par des taxons thermocalcicoles connus pour caractériser classiquement les Festuco valesiacae-Brometea erecti Braun-Blanq. & Tüxen ex Klika & Hadač 1944 et les Trifolio medii-Geranietea sanguinei T. Müll. 1962 (Anthericum ramosum, Buphthalmum salicifolium, Thymus praecox, Gymnadenia conopsea, G. odoratissima, Teucrium montanum, T. chamaedrys, Polygonatum odoratum, Oreoselinum nigrum, Vincetoxicum hirundinaria, Daphne cneorum, Sesleria caerulea, Carex humilis, Epipactis atrorubens, Hippocrepis comosa, Euphorbia cyparissias, Brachypodium rupestre, Prunella grandiflora, Globularia cordifolia, mais est aussi original par Polygala chamaebuxus, Erica carnea, Viola collina, Coronilla vaginalis, Calamagrostis varia subsp. v. Compte tenu de la complexité des deux classes citées, il est hors du propos de la présente publication de tenter d’y placer ces alliances nouvelles, et par ailleurs de définir ici une classe nouvelle pour les y inclure.
Pour la République tchèque, dans les « Erico-Pinetea », Chytrý (2013 : 368) cite encore le « Thlaspio montani-Pinetum sylvestris » serpentinicole dans lequel manquent Erica carnea, quoique celui-ci apparaisse avec une présence de II dans l’« Asplenio cuneifolii-Pinetum sylvestris » placé par l’auteur dans le Dicrano-Pinion sylvestris (Libbert 1933) Matuszkiewicz 1962. D’autres données, tant sur serpentinite que dolomie, sont encore disponibles dans le tableau II de Horvat (1959), mais ne peuvent donner lieu à validation car les données sources sont synthétiques.
L’ensemble des syntaxons 34 à 49 traduit des caractéristiques édaphiques plus fortement acides. La plupart des taxons basiphiles de l’ensemble 1 à 33 ont disparu, mais, selon les situations, certains se maintiennent en résistant à l’acidification ou en allant puiser les ions basiques nécessaires en dessous de l’horizon humifère acide. Les taxons caractéristiques sont ici des Ericaceae des genres Vaccinium et Rhododendron, ainsi que parfois Calluna vulgaris, plus rarement Kalmia procumbens, ce qui autorise leur rattachement aux Kalmio procumbentis-Vaccinietea microphylli Eggler ex R. Schub. 1960. Le syntaxon 34 est le « Rhododendro hirsuti-Pinetum mugo » Braun-Blanq. et al. 1939, qui devrait être redéfini. Le syntaxon 35 est le « Lycopodio annotini-Pinetum mugo » Starlinger 1992, dont la composante basse est redéfinie comme Lycopodio annotini-Rhododendretum hirsuti ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 6 du tableau 1 in Starlinger (1992, Tuexenia 12 hors texte). Le syntaxon 36 est le Seslerio albicantis-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 55 du tableau 3 in Minghetti (1996, Doc. Phytosoc., n. s., XVI hors texte), alors que le syntaxon 38 est le Seslerio albicantis-Polygaletum chamaebuxi ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 63 du tableau 2 in Minghetti (1996, Doc. Phytosoc., n. s., XVI hors texte), le 39 étant l’« Erico carneae-Pinetum » Braun-Blanq. 1939 selon Oberdorfer (1957 : 375, nommé Pineto-Ericetum carneae dans la figure 1). Caractérisé ou différencié relativement aux autres alliances des Kalmio-Vaccinietea par la combinaison d’Erica carnea, Polygala chamaebuxus, Rhododendron hirsutum (photo 16), Rh. ferrugineum, Juniperus communis subsp. sibirica, Daphne striata, Valeriana montana, V. tripteris, Carex digitata, Calamagrostis varia subsp. v., au moins l’ensemble 34 à 41 relève du classique Ericion carneae Rübel ex Grabherr et al. in Grabherr & Mucina 1993 (Theurillat et al., 1995 ; Mucina et al., 2016), donc à bien distinguer des « Ericion carneae » plutôt basiphiles définis dans l’ensemble 6 à 29.

Les syntaxons 42 et 43 relèvent aussi des Kalmio-Vaccinietea microphylli, mais pas de l’Ericion carneae : rassemblant deux associations bien définies, le 42 a la valeur d’une alliance, connue actuellement de Haute-Maurienne, riche en hautes herbes liées à l’Adenostylion alliariae, caractérisée ou différenciée par Veronica urticifolia, Hieracium prenanthoides, H. amplexicaule, Festuca flavescens, Adenostyles alliariae, Saxifraga cuneifolia s. l., Imperatoria ostruthium, Lactuca alpina, Actaea spicata, Avenella flexuosa subsp. f., Geranium sylvaticum, Oxalis acetosella, Prenanthes purpurea, Phyteuma spicatum, Luzula nivea, Melampyrum sylvaticum, Valeriana tripteris, Orthilia secunda, le Festuco flavescentis-Vaccinion myrtilli all. nov. hoc loco, typus nominis hoc loco : le Festuco flavescentis-Adenostyletum alliariae nov.
Pour mieux structurer cette partie Kalmio-Vaccinietea du tableau 6, on l’a extraite de ce dernier pour la reporter dans le tableau 7, tout en conservant la numérotation des quinze colonnes (la 43 n’y a pas été reprise). Les syntaxons 34 à 41 et 42 (en colonnes 1 à 3, 5 à 9 et 15) ont déjà été placés respectivement dans l’Ericion carneae et le Festuco flavescentis-Vaccinion myrtilli. On peut encore placer le 48 (colonne 4) dans l’Ericion carneae : Calamagrostio villosae-Rhododendretum ferruginei ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 21 du tableau 4 in Minghetti (1996, Doc. Phytosoc., n. s., XVI hors texte).
L’ensemble des colonnes 10 à 13 de ce tableau 7 partage plusieurs taxons avec l’Ericion carneae (Erica carnea, Rhododendron hirsutum, Polygala chamaebuxus, Rhodothamnus chamaecistus), mais en est par ailleurs fortement distinct. Il est notamment différencié par plusieurs fougères (Athyrium distentifolium, A. filix-femina, Dryopteris villarii, D. dilatata, D. filix-mas, Polystichum aculeatum, P. lonchitis, Gymnocarpium dryopteris, Oreopteris limbosperma) traduisant une humidité atmosphérique notable dans ce massif du Dachstein culminant à 2 995 m (sud-est de Salzburg, en Autriche ; Maier, 1994), ainsi que plusieurs herbes de mégaphorbiaies et microphorbiaies orophiles ; il est caractérisé aussi par des taxons plus originaux tels que Oreojuncus monanthos, Homogyne discolor, Pteroselinum austriacum, Gentiana pannonica, Alchemilla anisiaca, Heracleum austriacum, Cardamine enneaphyllos, Betonica alopecuros, Jacobaea abrotanifolia, Aposeris foetida, Knautia dipsacifolia auxquels on peut ajouter Achillea clavennae absent du tableau. Il s’agit clairement d’une alliance distincte des précédentes que l’on peut dénommer Knautio dipsacifoliae-Rhododendrion hirsuti all. nov. hoc loco. Pour typifier cette alliance, il faut en décrire quelques associations :
- en colonne 10, le Gymnocarpio dryopteridis-Lycopodietum annotini ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 65 du tableau 3 in Maier (1994, Stapfia 35 : 111-114) ;
- en colonne 11, le Cardamino enneaphylli-Huperzietum selaginis ass. nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 39 du tableau 3 in Maier (1994, Stapfia 35 : 111-114, en remplaçant Dentaria enneaphyllos par Cardamine e.) ;
- en colonne 12, le Gentiano pannonicae-Rhododendretum hirsuti nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 60 du tableau 3 in Maier (1994, Stapfia 35 : 111-114) ;
- en colonne 13, l’Oreojunco monanthi-Rhododendretum hirsuti nov. hoc loco, typus nominis hoc loco: composante cryptophyto-chaméphytique du relevé 80 du tableau 3 in Maier (1994, Stapfia 35 : 111-114, en remplaçant Juncus monanthos par Oreojuncus m.).
Cet inventaire permet alors de retenir le Cardamino enneaphylli-Huperzietum selaginis nov. comme typus nominis hoc loco du Knautio dipsacifoliae-Rhododendrion hirsuti nov. Morton (1993) en publie aussi des relevés isolés (relevés 1, 4, 9d, 15, 19, 22e, 25…). En colonne 14, le syntaxon 46 est assez éloigné des précédents, le Calamagrostio villosae-Vaccinietum vitis-idaeae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 53 du tableau 5 in Minghetti (1996, Doc. Phytosoc., n. s., XVI hors texte).
Il reste encore à traiter le large ensemble allant des colonnes 16 à 33 de ce tableau 7 correspondant surtout à des syntaxons décrits des Alpes juliennes, vers la Slovénie, notamment étudiées par I. Dakskobler et divers coauteurs, le cœur de cette unité étant principalement formée des colonnes 17 à 31.
La colonne 16 est le Dryopterido dilatatae-Rhododendretum hirsuti ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 7 du tableau 1 in Dakskobler (2016, Hacquetia 15 (1) : 76-80). Avec 110 relevés, l’ensemble 17 à 25 paraît relever d’une seule association pourvue de variations, le Paederoto luteae-Rhododendretum hirsuti ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 12 du tableau 1 in Dakskobler et al. (2018, Hacquetia 17 (2) : 268-277). Les colonnes 26 et 27 relèvent du Festuco nitidae-Vaccinietum myrtilli ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 2 du tableau 15 in Dakskobler et al. (2018, Hacquetia 17 (2) : 422-431). De même, les colonnes 28 à 30 peuvent se rattacher à un unique Homogyno discoloris-Dryadetum octopetalae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 3 du tableau 21 in Dakskobler et al. (2018, Hacquetia 17 (2) : 477-481). La colonne 31 s’avère être un Knautio drymejae-Ericetum carneae ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 1 du tableau 23 in Dakskobler et al. (2018, Hacquetia 17 (2) : 489-493). Les colonnes 32 et 33 forment le Campanulo carnicae-Rhododendretum hirsuti ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 18 du tableau 24 in Dakskobler et al. (2018, Hacquetia 17 (2) : 494-501).
Tout cet ensemble de dix-huit colonnes possède la valeur d’une alliance des Alpes de Slovénie caractérisée ou différenciée des alliances voisines par de nombreux taxons, dont Aconitum lamarckii, A. angustifolium, Dryopteris villarii, D. expansa, Festuca calva, F. nitida, Aquilegia nigricans, Arabis procurrens subsp. vochinensis, Knautia drymeja, Paederota lutea, Gymnocarpium robertianum, Cyclamen purpurascens (photo 17), Anemone nemorosa, A. trifolia, Galium laevigatum, Phegopteris connectilis, Astrantia carniolica (photo 18), A. bavarica, Cirsium erisithales, Saxifraga cuneifolia s. l., Calamagrostis arundinacea, Senecio cacaliaster, Luzula luzuloides, L. luzulina, Geum rivale, Rumex arifolius, Laserpitium peucedanoides, Veratrum album subsp. lobelianum, Homogyne sylvestris, H. discolor (photo 19), Heliosperma alpestre (photo 20)… D’autres comme Aposeris foetida (photo 21), Betonica alopecuros (photo 22), Helleborus niger… sont communs à cette unité et au Knautio dispsacifoliae-Rhododendrion hirsuti. Cette belle alliance slovène peut être dénommée Homogyno sylvestris-Rhodothamnion chamaecisti all. nov. hoc loco, typus nominis hoc loco : le Paederoto luteae-Rhododendretum hirsuti nov.
Les cinq alliances du tableau 7 partagent notamment Erica carnea, Polygala chamaebuxus, Rhododendron hirsutum, Calamagrostis villosa, Rhodothamnus chamaecistus, Juniperus communis subsp. sibirica, un ensemble de taxons suffisamment original pour justifier un ordre distinct des Rhododendro-Vaccinietalia Braun-Blanq. in Braun-Blanq. & Jenny 1926 au sein des Kalmio-Vaccinietea microphylli et pour reprendre les Rhododendro hirsuti-Ericetalia carneae Grabherr et al. in Grabherr & Mucina 1993 (Die Pflanzengesellschaften Osterreichs, II non consulté). Il n’y a pas lieu de séparer cet ordre dans une classe à part, celle des Rhododendro hirsuti-Ericetea carneae (Schubert et al., 2001).






4. Quelques aspects écologiques
En plus des facteurs chorologiques, on peut tenter d’expliquer la diversité de la végétation du sous-bois de ces pinèdes par des facteurs édaphoclimatiques.
Un des pionniers dans l’étude des relations entre phytosociologie et pédologie, Pallmann (1948) met en relation les unités phytosociologiques connues à son époque avec des facteurs climatiques. Ainsi les figures 2 et 3 replacent ces unités selon l’exposition et par suite les facteurs photiques et thermiques ; on voit la situation du « Pineto-Caricetum humilis » en exposition sud, celle des variations du « Mugeto-Ericetum » plutôt à l’est et à l’ouest, celle du « Mugeto-Rhodoretum hirsuti » plutôt au nord. Selon l’approche développée ici, le « Pineto-Caricetum humilis » correspond aux sous-bois cryptophyto-chaméphytiques à aspect de pelouses basiphiles du type Melampyro catalaunici-Polygalion chamaebuxi, Galio corrudifolii-Arctostaphylion uvae-ursi et Onobrychido saxatilis-Ononidion rotundifoliae des Alpes occidentales, Festuco amethystinae-Polygalion chamaebuxi des Alpes centro-orientales. Le « Mugeto-Rhodoretum hirsuti » correspond aux sous-bois cryptophyto-chaméphytiques à aspect de landes acidiphiles à Vaccinium. Cette diversité est à mettre en relation avec l’activité biologique du sol selon le facteur thermique et ses conséquences sur le développement de l’humus : forte activité en exposition sud avec humus réduit de type mull calcique, faible activité en exposition nord avec accumulation d’humus de type moder ou mor, notamment sous l’influence du dépôt d’aiguilles de résineux acidifiantes.
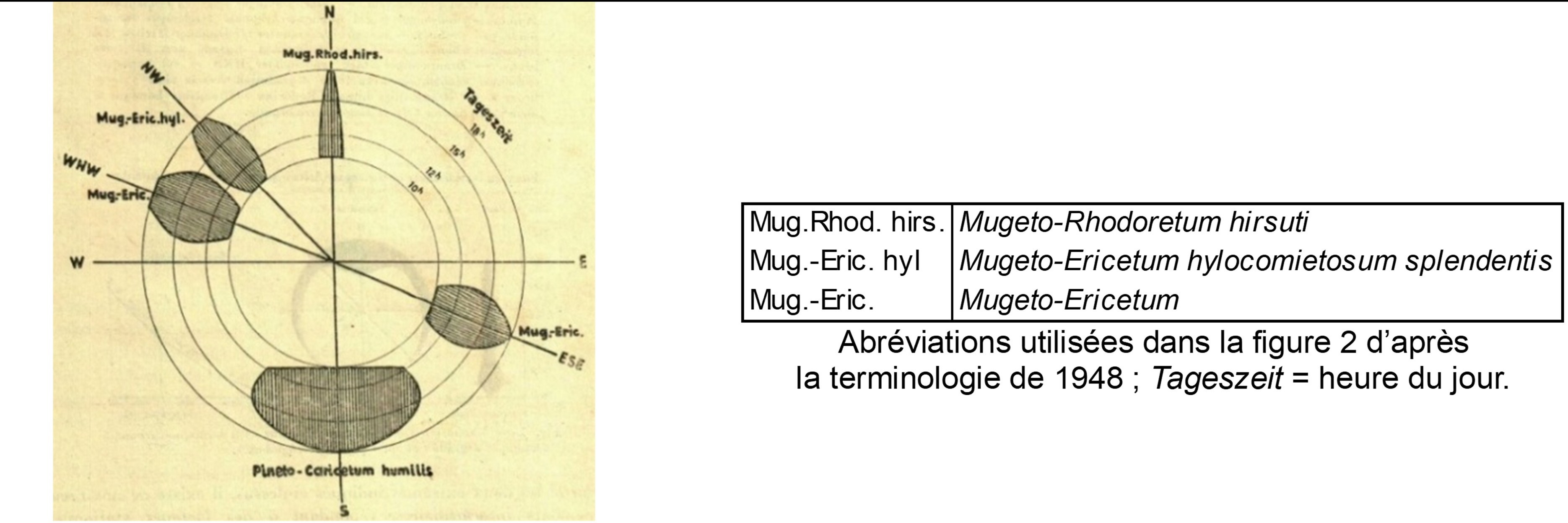
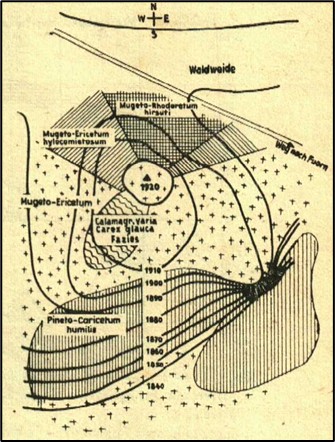
En 1966, Bartoli précise aussi ces relations entre végétation et sols. En exposition sud, le sol est de type rendzine jeune à brunifiée, alors qu’en exposition nord, en outre sur roches mères silicatées (schistes, grès, quartzites), sous la végétation rattachée ici au Festuco flavescentis-Vaccinion myrtilli, il est plutôt de type sol brun plus ou moins lessivé à mull-moder, voire un sol podzolique ferrugineux ou un podzol.
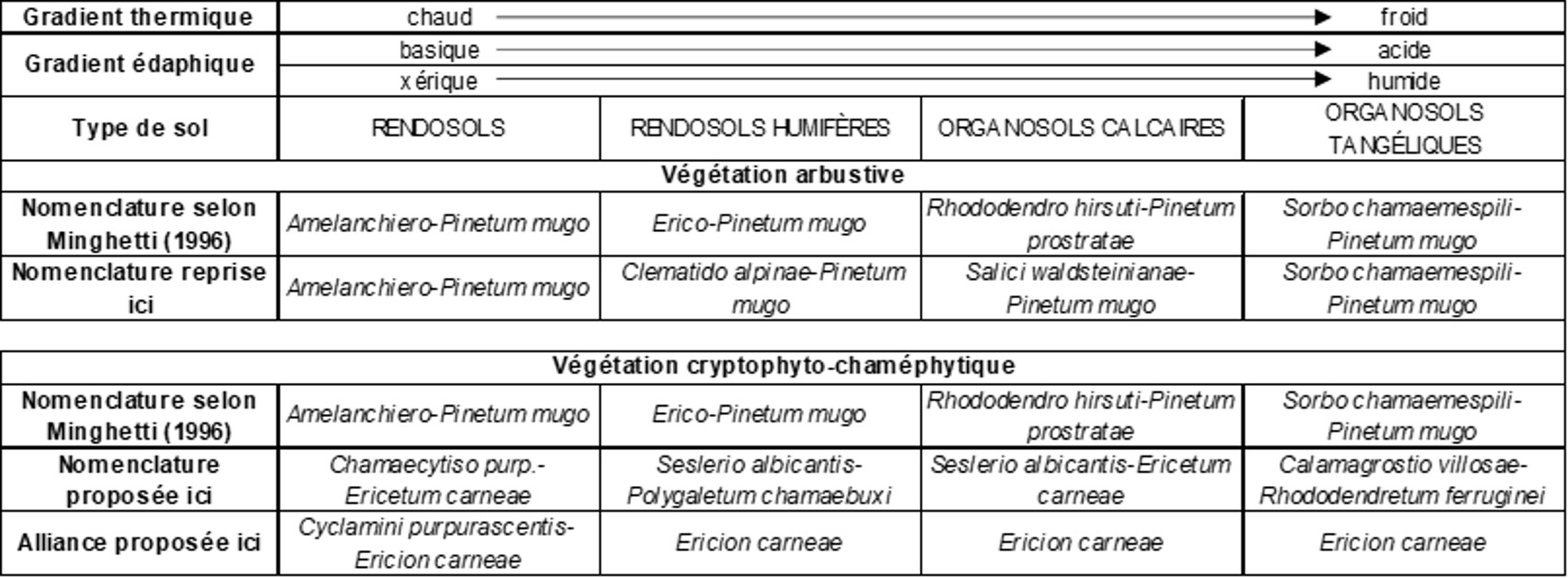
En 1996, Minghetti accompagne la description de ses cinq Pineta mugo de précisions écologiques, en particulier en ordonnant les associations qu’il a reconnues selon des gradients. On laissera de côté le « Rhododendro ferruginei-Pinetum prostratae » développé sur substrats cristallins (schistes, paragneiss, granites, tonalites, granodiorites) et volcaniques (rhyolites, rhyodacites) pour ne considérer que les quatre autres sur roche mère plus ou moins homogène pour la végétation (calcaires durs, dolomies, calcaires marneux), abordant ainsi une approche systémique. Ainsi ses figures 16 à 18 les placent selon des gradients traduits ici dans le tableau 8, en distinguant la végétation arbustive (reprise d’après de Foucault, 2021, § 2) et la végétation cryptophyto-chaméphytique ; les types de sols sont repris de Minghetti et al. (1997) qui ont adopté la nomenclature du Référentiel pédologique de l’AFES (1992, noms en majuscules selon la recommandation de ce document).
La série pédologique spatiale allant des RENDOSOLS modaux aux ORGANOSOLS TANGÉLIQUES a donc aussi la valeur d’une série temporelle menant des premiers aux derniers sous l’influence des facteurs édaphoclimatiques, ainsi que le met aussi en évidence Duchaufour (1983) ; c’est un nouvel exemple d’isomorphisme spatio-temporel, c’est-à-dire de symétrie entre phénomènes se déroulant dans l’espce et phénomènes se déroulant dans le temps (de Foucault, 2014), appliqué ici à la pédologie. Cette dynamique permet alors d’interpréter la présence du groupe sociologique à Rubus saxatilis du tableau 7 : ces taxons pourraient être relictuels du stade initial de pelouses sur RENDOSOLS dans les landes finales acidiphiles.
5. Conclusion
Cette monographie phytosociologique consacrée aux pinèdes à Erica carnea et Polygala chamaebuxus revues selon le paradigme synusial a donc permis de reconnaître une cinquantaine d’associations originales. Les syntaxons cryptophyto-chaméphytiques sont replacés dans neuf alliances, six plutôt basiphiles, qui devraient pouvoir se rattacher aux Festuco-Brometea erecti ou aux Trifolio-Geranietea sanguinei, et trois acidiphiles venant s’ajouter au classique Ericion carneae dans les Kalmio-Vaccinietea microphylli. Dans l’avenir, il faudra sans doute ajouter des travaux antérieurs non utilisés ici et il reste à espérer que cet échantillon de publications et les tableaux qui en ont été retirés pourront aider dans l’avenir à une meilleure compréhension de ces fabuleux habitats et à la définition de nouvelles unités.
Bibliographie
Accetto M. 1999. Asociacija Carici sempervirentis-Pinetum nigrae Accetto (1996) 1999 nom. nov. v Sloveniji. Zbornik gozdarstva in lesarstva 60 : 107-151 (en slovène).
Accetto M., 2001. Asociacija Daphno alpinae-Pinetum nigrae ass. nova v Sloveniji. Zbornik gozdarstva in lesarstva 64 : 5-39 (en slovène).
AFES (D. Baize & M.-C. Girard, coord.), 1992. Référentiel pédologique, principaux sols d’Europe. Inra Éditions, Paris, 222 p.
Bartoli C., 1966. Études écologiques sur les associations forestières de la Haute-Maurienne. Annales des sciences forestières XXIII (3) : 431-741.
Braun-Blanquet J., 1922. Une reconnaissance phytosociologique dans le Briançonnais. Bulletin de la Société botanique de France 69, rapports d’excursion de la session extraordinaire tenue dans le Briançonnais en juillet 1922 : 77-103.
Braun-Blanquet J., 1961. Die inneralpine Trockenvegetation. Geobotanica Selecta 1 : 1-273.
Braun-Blanquet J. & Richard F., 1949. Groupements végétaux et sols du bassin de Sierre. Bulletin de la Murithienne (bulletin annuel de la Société valaisanne des sciences naturelles) 64 : 106-134.
Čarni A. & Mucina L., 2015. Validations and typifications of some south European syntaxa. Hacquetia 14 (2) : 289-299.
Chytrý M., 2013. Vegetace České republiky, 4 – Lesní a křovinná vegetace (Vegetation of the Czech Republic, 4 – Forest and scrub vegetation). Academia, Praha, 551 p. (en grande partie en tchèque, reste en anglais).
Dakskobler I., 1999. Contribution to the knowledge of the association Fraxino orni-Pinetum nigrae Martin-Bosse 1967. Wissenschaftliche Mitteilungen des Niederosterreichischen Landesmuseums 12 : 25-32.
Dakskobler I., 2016. Two new pioneer communities of Sorbus aucuparia and Sorbus aria in the southern Julian Alps. Hacquetia 15 (1) : 67-91.
Dakskobler I., Seliškar A. & Rozman A., 2018. Phytosociological analysis of European larch forests in the Southeastern Alps. Hacquetia17 (2) : 245-519.
Duchaufour Ph., 1983. Pédologie, I – Pédogénèse et classification. Masson, Paris, 491 p.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Betulo carpaticae-Alnetea viridis Rejmánek in Huml, Lepš, Prach & Rejmánek 1979. Journal de botanique de la Société botanique de France 60 : 47-68.
Foucault B. (de), 2014. Les isomorphismes spatio-temporels : approche transdisciplinaire et valeur heuristique. Bulletin de la Société botanique du Centre-Ouest, n. s., 45 : 438-445.
Foucault B. (de), 2018. Contributions à la flore des Alpes, compte-rendu de la 151e session extraordinaire de la Société botanique de France en Maurienne (Savoie) : quelques données phytosociologiques sur la haute Maurienne. Journal de botanique de la Société botanique de France 81 : 54-61.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2023. Miscellanées phytosociologiques V : syntaxons nouveaux, validés ou complétés. Carnets botaniques 154 : 1-16 + 6 tableaux, https://doi.org/10.34971/M5RZ-7B62.
Fukarek P. & Stefanović V., 1958. Prašuma Peručica i njena vegetacija. Radovi Šumarskog fakulteta Univerziteta u Sarajevu 6 (3) : 93-146 (en slovène).
Grabherr G. & Mucina L. (eds.), 1993. Die Pflanzengesellschaften Osterreichs, II. Gustav Fischer Verlag, Jena.
Hemp A., 1995. Die landschaftsökologische Bedeutung der Dolomit kiefernwälder (Buphthalmo-Pinetum) in der Frankenalb. Berichte der Bayerischen Akademie für Naturschutz und Landschaftspflege 19 : 205-248.
Hölzel N., 1996a. Synopsis der Pflanzengesellschaften Deutschlands, I – Erico-Pinetea (H6), alpisch-dinarische Karbonat-Trocken-Kiefernwälder. Floristisch-soziologische Arbeitsgemeinschaft und die Reinhold-Tüxen-Gesellschaft, Göttingen : 11-49.
Hölzel N., 1996b. Schneeheide-Kiefernwälder in den mittleren Nördlichen Kalkalpen. Laufener Forschungsbericht 3 : 1-192.
Horvat I., 1959. Sistematski odnosi termofilnik hrastovih i borovih suma Jugoistocn Europe (Wärmeliebende Eichen- und Kiefernwäldern Südosteuropas in systematischer Betrachtung). Biološki Glasnik 12 : 1-40 (en croate).
Lakušić R., Pavlović D., Adžić S., Kutleša L., Mišić L., Redžić S., Maljević D. & Bratović S., 1979. Struktura i dinamika ekosistema planine Vranice u Bosni. Drugi kongres ekologa Jugoslavije 1 : 605-714 (en croate).
Maier F., 1994. Die Waldvegetation an der Dachstein-Nordabdachung (Oberösterreich), Pflanzensoziologie, Floristik, Naturschutz. Stapfia 35 : 1-117.
Martin-Bosse H., 1967. Schwarzföhrenwälder in Kärnten. Angewandte Pflanzensoziologie XX : 1-89.
Minghetti P., 1996. Analisi fitosociologica delle pinete a Pinus mugo Turra del Trentino (Italia). Documents phytosociologiques, n. s., XVI : 461-503.
Minghetti P., 2003. Le pinete a Pinus sylvestris del Trentino-Alto Adige (Alpi italiane): tipologia, ecologia e corologia. Braun-Blanquetia 33 : 1-95.
Minghetti P., Sartori G. & Lambert K., 1997. Relations sol-végétation dans les pinèdes à Pinus mugo Turra du Trentin (Italie). Revue d’écologie alpine 4 : 23-34.
Morton F., 1933. Pflanzensoziologische Untersuchungen im Gebiet des Dachsteinmassivs, Sarsteins und Höllengebirges. Feddes Repertorium LXXI : 1-33.
Mucina L., Bültmann H. et al., 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 19, suppl. 1 : 3-264.
Oberdorfer E., 1957. Süddeutsche Pflanzengesellschaften. Pflanzensoziologie 10 : 1-564.
Pallmann H., 1948. Pédologie et phytosociologie (conférence donnée au Congrès international de pédologie de Montpellier, 3 mai 1947). In Comptes rendus de la Conférence de pédologie méditerranéenne, Association française pour l’étude du sol, réunions dans la métropole : 3-36.
POWO, Plant of the World online, https://powo.science.kew.org.
Rameau J.-C., 2001. Habitats forestiers. Cahiers d’habitats Natura 2000, La Documentation française 1 (2), 423 p.
Rozman A., Dakskobler I & Šilc U., 2020. Phytosociological analysis of basophilic Scots pine forests in the Southeastern Alps. Hacquetia 19 (1) : 23-80.
Schubert R., Hilbig W. & Klotz S., 2001. Bestimmungsbuch der Pflanzengesellschaften Deutschlands. Spektrum Akademischer Verlag, Heidelberg, 472 p.
Šibik J., Šibiková I. & Klement J., 2010. The subalpine Pinus mugo-communities of the Carpathians with a European perspective. Phytocoenologia 40 (2-3) : 155-188.
Starlinger F., 1992. Rotföhren- und Spirkenwälder am Fernpaß (Tirol). Tuexenia 12 : 67-91.
Surina B., 2013. Heaths with dwarf ericaceous shrubs and Alpine juniper (Juniperus alpina) in the Dinaric Alps: a nomenclatorial and synsystematic re-appraisal. Acta Botanica Croatica 72 (1) : 113-132.
Theurillat J.-P., Aeschimann D., Küpfer Ph. & Spichiger R., 1995. The higher vegetation units of Alps. Colloques phytosociologiques XXIII, Large area vegetation surveys : 189-239.
Thévenin S. & Royer J.-M., 2001. Les groupements végétaux de la Champagne crayeuse. Bulletin de la Société d’étude des sciences naturelles de Reims, n° spécial : 1-32.
Vanden Berghen C., 1963. Étude de la végétation des Grands Causses du Massif central de France. Mémoires de la Société royale de botanique de Belgique 1 : 1-285.
Villaret J.-C., Van Es J. et sept autres auteurs, 2019. Guide des habitats naturels des Alpes, du Jura méridional à la Haute Provence et des bords du Rhône au Mont-Blanc, description, écologie, espèces diagnostiques, conservation. Naturalia Publications, Turriers, 639 p.
Wagner H., 1941. Die Trockenrasengesellschaften am Alpenostrand. Denkschriften der Akademie der Wissenschaften Wien, Mathematisch-Naturwissenschaftliche Klasse 104 : 1-81.
Zöttl H., 1951. Die Vegetationsentwicklung auf Felsschutt in der alpinen und subalpinen Stufe des Wettersteingebirges. Jahrbuch des Vereins Schultze der Alpenpflanzen und -tiere 16 : 10-74.
Remerciements
Boštjan Surina (Musée d’histoire naturelle de Rijeka, Croatie) m’a aidé à acquérir certaines publications croates et Franck Le Driant m’a autorisé à utiliser des photos de sa base photographique FloreAlpes (https://www.florealpes.com) pour illustrer cette synthèse, je leur en suis très reconnaissant.


