
Nouvelles données sur la végétation du versant audois de la Montagne Noire
Title
New data on the vegetation of the Aude side of the Montagne Noire
Résumé
Étudié depuis une dizaine d’années, le versant audois de la Montagne Noire a fait l’objet de nombreux nouveaux relevés inédits portant sur la végétation forestière, les lisières arbustives et herbacées, les mégaphorbiaies, les landes, les pelouses vivaces et thérophytiques, les murs et les rochers, les bas-marais, définissant plusieurs associations végétales nouvelles.
Abstract
Over the last ten years or so, the Aude side of the Montagne Noire has been the subject of a large number of new unpublished relevés covering forest vegetation, shrub and herbaceous margins, tall herbs communities, heaths, perennial and therophytic lawns, walls and rocks, acid marshes, defining several new plant associations.
1. Caractères généraux de la Montagne Noire audoise
1.1. Caractères géographiques et géologiques
La Montagne Noire (orthographe suivie ici, mais parfois écrit Montagne noire ou montagne Noire) est un massif montagneux situé à l’extrémité sud-ouest du Massif central, en France (carte 1). Elle est partagée entre quatre départements, l’Aude et le Tarn en sont les principaux, tandis que la Haute-Garonne et l’Hérault en forment les bordures. Le massif, orienté est-ouest, présente deux visages : le versant nord abrupt est couvert de forêts sombres de chênes, de hêtres et de sapins ; le versant sud est moins abrupt et comprend le Cabardès au sud, qui s’étend jusqu’à Carcassonne, le Minervois à l’est et une partie du Lauragais à l’ouest. Son point culminant est le pic de Nore, à 1 211 m d’altitude dans le département de l’Aude ; le sommet est couronné d’une importante antenne TDF de 100 m de hauteur qui permet de le repérer de loin. La Montagne Noire est incluse pour partie dans le parc naturel régional du Haut-Languedoc.
Au sens strict, la Montagne Noire s’érige à la fin de l’Éocène en conséquence de la compression due au rapprochement de l’Ibérie et de la formation des Pyrénées ; c’est la faille de Mazamet qui permet cette érection sur 1 200 m. Le soulèvement du relief a permis de faire affleurer des roches anciennes, autrefois profondément enfouies : roches métamorphiques variées, gneiss (pic de Nore) et granite. Les nappes de charriage inversées (les roches les plus anciennes se trouvant au-dessus des roches les plus jeunes) au sud-ouest du massif sont très célèbres.
La Montagne Noire des géologues est nettement plus large que celle des géographes : elle s’étend presque jusqu’à Castres et Camarès, incorporant ainsi les monts de Lacaune, le Sidobre et les monts de l’Espinouse. Ce massif montagneux se divise en trois zones : un versant septentrional constitué d’écailles déversées vers le sud-est, un versant méridional composé d’un empilement de nappes-plis couchés très complexe et enfin une zone axiale constituée de dômes gneissiques (ortho- et paragneiss) et de granites encadrés par des métasédiments (sédiments métamorphisés). Il existe parfois des nappes de calcaire dévonien.
1.2. Caractères hydrologiques
La Montagne Noire est drainée par de nombreux cours d’eau, de toutes tailles, et la crête supérieure sépare le bassin versant méridional, audois, orienté vers le fleuve Aude, et le bassin septentrional, tarnais, orienté vers la Garonne. En ce qui concerne le premier, les cours d’eau principaux sont les suivants, d’ouest en est (certains sont reportés sur la carte 1).
- Le Sor naît au sud-ouest d’Arfons (Tarn), se dirige vers le lac des Cammazes, puis remonte vers le nord-ouest pour arroser Durfort, à partir duquel il draine le Lauragais jusqu’à sa confluence avec l’Agoût, à Sémalens (Tarn).
- Le Lampy naît à Arfons (Tarn), dans la Sagne du Lampy, s’élargit pour former le bassin du Lampy Neuf, puis se dirige vers l’ouest et le sud pour gagner Cennes-Monestier, Saint-Martin-le-Vieil, Alzonne où il se jette dans le Fresquel.
- L’Alzeau prend sa source en forêt de Montaut (commune d’Escoussens, Tarn), passe à l’ouest de Laprade, s’élargit pour former le lac de la Galaube, puis passe à Lacombe où il donne naissance partiellement à la rigole de la Montagne ; vers le sud, il forme le lac de Saint-Denis, puis descend encore pour former la Rougeanne en recevant la Dure.
- La Dure débute à Laprade, alimente le lac de Laprade-Basse, traverse Caudebronde, Cuxac-Cabardès, Brousses-et-Villaret et devient la Rougeanne à partir de Montolieu en s’agrégeant à d’autres ruisseaux ; la Rougeanne rejoint ensuite le Fresquel.
- L’Orbiel naît de la confluence de plusieurs sources affleurant dans la Bézalado et la Sagne Grande (commune des Martys) ; vers le sud il traverse Miraval-Cabardès, Mas-Cabardès, Les Ilhes, Lastours, Conques-sur-Orbiel et rejoint l’Aude à Trèbes.
- L’Arnette prend sa source sur le versant sud-occidental du pic de Nore, sous le nom de Rec de Cuin, lequel se dirige d’abord vers le sud en arrosant Pradelles-Cabardès, puis oblique vers l’ouest sous le nom de ruisseau de la Barthe et remonte vers le nord pour gagner Mazamet où la rivière se jette dans le Thoré, qui appartient au bassin de la Garonne.
- La Clamoux débute sur le versant nord du pic de Nore qu’il contourne par l’est, traverse Castans, Cabrespine, Villeneuve-Minervois, Villegly et rejoint l’Orbiel vers Villedubert.
- L’Argent-Double débute par une source sous le col de Salettes (commune de Lespinassière), arrose Caunes-Minervois, traverse le Minervois, passe sous le canal du Midi et rejoint l’Aude au sud-est de La Redorte.
- L’Ognon est la réunion de plusieurs ruisseaux naissant sur Cassagnoles et Félines-Minervois (Hérault) ; au sud de cette dernière commune, il oblique vers l’est, puis le sud-est pour gagner Olonzac, passer sous le canal du Midi et rejoindre l’Aude.
- La Cesse enfin naît vers Ferrals-les-Montagnes, se dirige vers le sud, puis l’est, arrose la belle cité de Minerve et Aigues-Vives, oblique à nouveau vers le sud pour gagner Bize-Minervois et Sallèles-d’Aude avant de rejoindre l’Aude.
1.3. Caractères climatiques et phytogéographiques
Le climat est assez doux dans l’ensemble, de type atlantique sur le versant nord, de type méditerranéen au sud-est, mais assez venteux. Cependant, dans le haut Cabardès et sur le versant nord, il y est plus frais. L’été est moins chaud que dans la plaine grâce aux forêts humides et l’hiver est souvent frais avec des chutes de neige régulières sur les hauteurs. Dans le Minervois, le vent est plus présent (cers et marin se relaient), mais également le soleil. La pluviosité varie entre 600 mm par an dans le bas Minervois et 1 600 mm par an aux environs du pic de Nore.
Sur le plan phytogéographique, Barreau et al. (2016) subdivisent la partie audoise principalement en Montagne Noire méditerranéenne orientale, Montagne Noire méditerranéenne occidentale, Montagne Noire haute orientale et Montagne Noire atlantique occidentale ; ce sont respectivement les régions 61 à 64 du Catalogue de la flore de l’Aude (Barreau, 2019). Dans une synthèse phytogéographique plus large (de Foucault, 2020c), les régions 61 et 62 se rapprochent de la région 34, soit les Hautes Corbières centrales, alors que les régions 63 et 64 sont rapprochées de la région 42, soit les Hautes Corbières occidentales (et non orientales comme écrit par erreur en p. 5 de la publication de 2020c).
1.4. Quelques données sur la patrimoine ethnologique
La diversité des activités humaines y a laissé des traces encore visibles dans le paysage actuel. On peut d’abord citer les terrasses qui témoignent d’activités agricoles révolues et qui ont été progressivement envahies par la végétation forestière après leur abandon, les murets de pierres sèches (photo 1) qui peuvent maintenant accueillir une végétation chasmophile, les glacières (photo 2) où était empilée et stockée la neige durant l’hiver en vue d’une conservation prolongée et d’une utilisation ultérieure dans les plaines languedociennes par les limonadiers, cafetiers, bouchers, poissonniers… Des voies charretières témoignent de la circulation ancestrale de lourds charrois qui ont fini par éroder la roche en place (photo 3).



À la limite entre Cabardès, Minervois et Carcassès, on peut encore évoquer le cimetière (ou la nécropole) wisigoth du Moural dels Morts sur la commune de Villarzel-Cabardès (photo 4), découvert en 1967, rassemblant une quarantaine de tombes.

Ce patrimoine inclut enfin des châteaux médiévaux, parfois improprement dits « châteaux cathares », surtout les quatre de Lastours (photo 5).

L’eau a joué et joue encore une fonction importante en Montagne Noire audoise, notamment depuis sa domestication (par détournement ; de Foucault, 2023b) par P.-P. Riquet suivi par Vauban pour alimenter le canal des Deux-Mers, dit aujourd’hui canal du Midi. C’est surtout le cas de la prise d’Alzeau où l’on peut observer la séparation du cours sauvage et du cours domestiqué au niveau du départ de la rigole de la Montagne (photo 6). De ce point, la rigole se dirige vers les Cammazes où elle passe sous la voûte Vauban pour rejoindre le bassin de Saint-Ferréol à partir duquel elle engendre la rigole de la Plaine qui se dirige vers le canal jusqu’au seuil de Naurouze.

1.5. Quelques contributions phytosociologiques antérieures
La connaissance de la végétation de la Montagne Noire est finalement encore assez peu avancée. On a très peu de données publiées de la session de l’Amicale phytosociologique qui se déroula en 1973 dans cette région comme l’évoque la description en nomen nudum, donc invalide, du Lavandulo stoechadis-Ericetum cinereae par Géhu (1975 : 370). Vingt ans après, Julve et de Foucault (1994) publient le compte rendu phytosociologique d’une session botanique tenue dans le Tarn et l’Aude septentrionale, puis Fallour et al. (2013) font de même pour une minisession de la Société botanique du Centre-Ouest. Le 12 juin 2021, la Société botanique d’Occitanie y a proposé une journée floristique qui a donné lieu à un compte rendu floristique (Andrieu et al., 2021), mais non phytosociologique.
À partir de 2017, plusieurs publications ont apporté des données sur ce sujet, elles seront évoquées dans la suite de cette monographie et feront parfois l’objet de compléments postérieurs.
2. Méthodes et conventions
Pour décrire la végétation observée, nous avons suivi le paradigme synusial (Gillet et al., 1991 ; de Foucault, 2020a). La nomenclature des taxons suit Flora Gallica (Tison & de Foucault, 2014). Dans les tableaux et relevés isolés, pour gagner de la place, le signe * remplace ‘subsp.’ et ‘var.’ ; le signe ‘j’ indique une forme juvénile de taxons potentiellement arbustifs ou arborescents, le signe ! indique une forme plus vigoureuse que les individus normaux du taxon, alors que ° indique une forme à vitalité plus réduite que les individus normaux du taxon. Parfois le signe / est inséré entre deux taxons très proches qu’il n’a pas été possible de séparer (par exemple Polypodium vulgare/interjectum en l’absence de sporanges).
La plupart des relevés, c’est-à-dire à l’exception des plus anciens, ont donné lieu à leur localisation spatio-temporelle (latitude, longitude, altitude, date), à l’indication du recouvrement de la végétation (en %), de la surface (en m²) et du nombre de taxons.
3. Les boisements
Les boisements de Montagne Noire ont été globalement peu étudiés. À basse altitude, ce sont des chênaies vertes, à plus haute altitude des hêtraies. Il existe aussi des plantations artificielles de divers résineux, alors que Castanea sativa y a été anciennement introduit et qu’il persiste encore bien dans les paysages actuels.
3.1. La végétation arborescente
Au niveau de la végétation arborescente, on connaît par exemple le Sorbo ariae-Fagetum sylvaticae B. Foucault 2018 observé surtout dans les Corbières (de Foucault, 2018a ; photo 7), reconnaissable aussi chez Julve & de Foucault (1994 : 32, des monts de Lacaune, Tarn) ; le relevé suivant en donne un exemple :
Pradelles-Cabardès (11), vers le pic de Nore, 13/08/2011, 4 taxons
Fagus sylvatica 5, Abies alba 3, Sorbus aria 1, S. aucuparia *a. +.

Par ailleurs, des moliniaies asséchées peuvent être colonisées progressivement par des pinèdes pionnières, le Betulo pendulae-Pinetum sylvestris B. Foucault 2018 ou le Betulo pubescentis-Pinetum sylvestris B. Foucault 2018 (de Foucault, 2018b). Le relevé suivant décrit un nouvel individu du premier :
Les Martys (11), ‘Les Moussels’, N 43° 25’ 20,5’’, E 2° 20’ 40,9’’, 850 m, 20/07/2022, 100 %, 4 taxons
Pinus sylvestris 3, Betula pendula 3, Populus tremula +, Salix atrocinerea ! +.
3.2. La végétation des sous-bois herbacés
En ce qui concerne la végétation herbacée des sous-bois, des relevés isolés ont été apportés. Des environs des Martys, on a par exemple publié des relevés de sous-bois herbacés vivaces de mull acide à Conopodium majus subsp. m., Ranunculus serpens, Sanicula europaea, Melica uniflora, Phyteuma spicatum, Stellaria holostea…, végétation qui relève des Anemono nemorosae-Caricetea sylvaticae (de Foucault, 2018b : 144, 145) où l’on peut parfois observer Crocus nudiflorus et Lactuca plumieri. On a aussi décrit une association de terrasse alluviale à mull actif à Scilla bifolia, le Tractemo lilio-hyacinthi-Erythronietum dentis-canis B. Foucault 2020 (de Foucault, 2020a). Il existe encore des sous-bois herbacés thérophytiques, correspondant le plus souvent au Moehringio trinerviae-Geranietum robertiani Gillet in Ferrez 2009 (de Foucault, 2018b : 145).
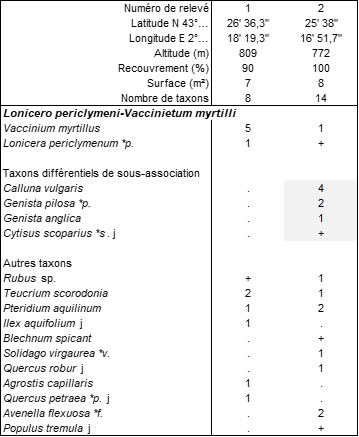
On connaît par ailleurs des sous-bois herbacés sur moder ou mor à Avenella flexuosa subsp. f., Melampyrum pratense…, voire des landes sciaphiles à Vaccinium myrtillus, le Lonicero periclymeni-Vaccinietum myrtilli B. Foucault 1974 (de Foucault, 2018b : 145 ; photo 8 ; voir aussi § 8).

Très originale et mieux définie est la végétation développée sous chênaie verte ou parfois sous cédraie artificielle, notamment autour de Cabrespine, avec Cephalanthera rubra. Un relevé isolé en a déjà été publié (de Foucault, 2020a : 101), le tableau 1 en rapporte neuf nouveaux qui confirment l’existence du Cephalanthero rubrae-Rubietum peregrinae ass. nov. hoc loco, typus nominis hoc loco : relevé 5 du tableau 1 hoc loco (photo 9) retranscrit ci-après :
Cabrespine (11), début du chemin de la grotte du Maquis, N 43° 21’ 24,9’’, E 2° 26’ 54,3’’, 494 m, 14/06/2022, 45 %, 60 m², 16 taxons
- combinaison floristique caractéristique : Rubia peregrina *p. 2, Cephalanthera rubra 1, Osyris alba 1, Helleborus foetidus 1, Limodorum abortivum 1, Ruscus aculeatus 1, Daphne laureola +, Asplenium onopteris +, Epipactis microphylla +, Carex flacca *f. +, Piptatherum paradoxum +,
- autres taxons : Buxus sempervirens j 2, Quercus ilex j 1, pubescens j +, Clematis vitalba j +, Rhamnus alaternus *a. j +.
L’altiude moyenne de ces relevés est de 436 m et le nombre moyen de taxons par relevé est d’environ 13. On voit sur le tableau que la composante hémicryptophyto-géophytique est assez variable en dehors des deux taxons éponymes, alors que les juvéniles d’arbustes et arbres sont bien représentés. Avec Asplenium onopteris et Carex distachya, il semble que cette association se place mieux dans le Carici distachyae-Asplenion onopteridis B. Foucault 2020 et les Rubio peregrinae-Asplenietea onopteridis B. Foucault 2020 que dans les Anemono nemorosae-Caricetea sylvaticae Gillet ex Gillet & B. Foucault in B. Foucault 2020.

Annexe du tableau 1 (tous ces relevés viennent de l’Aude) – rel. 1 à 4 : divers points à Cabrespine, au-dessus du gouffre, 14/06/2022 ; rel. 5 : Cabrespine, début du chemin de la grotte du Maquis, 14/06/2022 ; rel. 6 : Cabrespine, nord du parking du gouffre, au-dessus de ‘La Gaougnasse’, 14/06/2022 ; 7 : Cabrespine, bord de la D 112, ‘Laval’, 02/06/2023 ; rel. 8 et 9 : Cabrespine, bord D 112, au-dessus du ruisseau du Cros, 02/06/2023.
4. Les lisières arbustives
En ce qui concerne les fourrés de lisière, diverses associations ont déjà été décrites, mais plusieurs nouveaux relevés viennent conforter leur définition.
Le Bupleuro fruticosi-Juniperetum phoeniceae B. Foucault 2022 a été décrit en 2021, mais correctement validé en 2022 (de Foucault, 2021a, 2022a). Il s’agit d’un fourré de corniche fortement exposé au sud, caractérisé par Juniperus phoenicea subsp. ph., Bupleurum fruticosum, Amelanchier ovalis subsp. o., Rhamnus saxatilis subsp. s., Buxus sempervirens et parfois, de manière inattendue, Erica arborea (photo 10). Le tableau 2 en rapporte six relevés nouveaux. On remarque l’absence du premier taxon éponyme dans le relevé 3 ; l’altitude serait-elle trop élevée ici pour Bupleurum fruticosum ? Un relevé de cette association a déjà été publié de l’Hérault (environs de Minerve ; Julve & de Foucault, 1994 : 36), ce qui élargit l’aire de cette association. Sans qu’il ait pu être relevé faute de temps, il a été observé sur les corniches de marbre de Caunes-Minervois (Aude ; photo 11).


Le Cytiso scoparii-Ericetum arboreae B. Foucault 2015 est un fourré acidiphile à Erica arborea, E. scoparia subsp. s. et Cytisus scoparius subsp. s. plutôt inféodé aux plaines audoises à caractère méditerranéen atténué, mais atteignant la Montagne Noire inférieure (de Foucault, 2015). Le tableau 3 en rapporte neuf relevés nouveaux, dont un provenant de la Malepère, au sud du fleuve Aude.
Le Sorbo ariae-Cytisetum scoparii B. Foucault 2019 est un fourré à Cytisus scoparius subsp. s. relayant le Cytiso scoparii-Ericetum arboreae à plus haute altitude (typiquement en moyenne 883 m ; de Foucault, 2019). Le tableau 4 en rapporte dix-sept relevés nouveaux (photo 12), avec une variation significative différenciée par Sorbus aucuparia subsp. a. j et Pinus mugo subsp. uncinata j en altitude supérieure (relevés 14 à 17 ; altitude moyenne : 1 058 m). Sous le pic de Nore, la limite altitudinale supérieure de ces fourrés est marquée par un liseré dense paucispécifique à Pinus mugo subsp. uncinata arbustif, qui évoque un peu par sa physionomie les brousses subalpines à Pinus mugo subsp. m. ou est-asiatiques à Pinus pumila (Wilmanns et al., 1985 ; de Foucault, 2021d, 2023c) :
a) Pradelles-Cabardès (11), sud du pic de Nore, N 43° 25’ 24,2’’, E 2° 27’ 44’’, 1 191 m, 08/09/2022, 100 %, 70 m², 3 taxons, photo 13
Pinus mugo *uncinata j 4, Fagus sylvatica j 3, Sorbus aria j 2.
b) Pradelles-Cabardès (11), est du pic de Nore, N 43° 25’ 27,3’’, E 2° 27’ 59,3’’, 1 173 m, 19/07/2023, 90 %, 25 m², 4 taxons
Pinus mugo *uncinata j 3, Fagus sylvatica j 3, Sorbus aria j 2, S. aucuparia *a. j 2.


Le Frangulo alni-Ilicetum aquifolii Géhu & B. Foucault in B. Foucault 2018 est un sous-bois arbustif oligotrophile associé au sous-bois herbacé à Avenella flexuosa déjà cité. Sur les croupes rocheuses chaudes, on peut parfois observer le Junipero communis-Cytisetum oromediterranei B. Foucault 2022 (de Foucault, 2022b), dont voici un nouveau relevé :
Cabrespine, chemin du Château, au-dessus de la D 9, N 43° 22’ 56,1’’, E 2° 27’ 28,6’’, 714 m, 19/07/2023, 80 %, 15 m², 9 taxons, photo 14
- combinaison caractéristique ; Cytisus oromediterraneus 3, scoparius *s. 1, Juniperus communis *c. +,
- autres taxons : Quercus ilex j 1, Erica scoparia *e. 1, Sorbus aria j 1, Crataegus monogyna +, Rubus ulmifolius +, Rosa canina +.

Juniperus communis subsp. c. colonise aussi des bas-marais acidiphiles en voie d’assèchement comme on l’a observé aux Martys (de Foucault, 2018b, tableau 12) ; on a retrouvé à nouveau cette situation dans un autre site de la même commune :
Les Martys (11), ‘La Sagne Grande’, N 43° 26’ 29,4’’, E 2° 17’ 46,7’’, 807 m, 19/07/2022, 5 taxons
Juniperus communis *c. 2, Frangula alnus *a. 1, Quercus petraea *p. j 1, Alnus glutinosa j 1, Betula pendula j +.
Dans l’étude de 2018b, ce fourré turficole avait été interprété comme une variation à J. communis de l’Osmundo regalis-Salicetum atrocinereae Braun-Blanq. & Tüxen 1952, ce n’est en tout cas pas le Junipero communis-Franguletum alni décrit des pointements de serpentines du Limousin (de Foucault, 1986b). On peut aussi rattacher à l’Osmundo–Salicetum atrocinereae le relevé suivant de fourré oligotrophile nettement plus hygrophile à Salix aurita :
Les Martys (11), ‘Les Moussels’, N 43° 25’ 20,5’’, E 2° 20’ 40,9’’, 850 m, 20/07/2022, 30 %, 50 m², 5 taxons
Salix atrocinerea 2, S. aurita 2, Frangula alnus *a. 1, Alnus glutinosa j +, Betula pendula j 1.
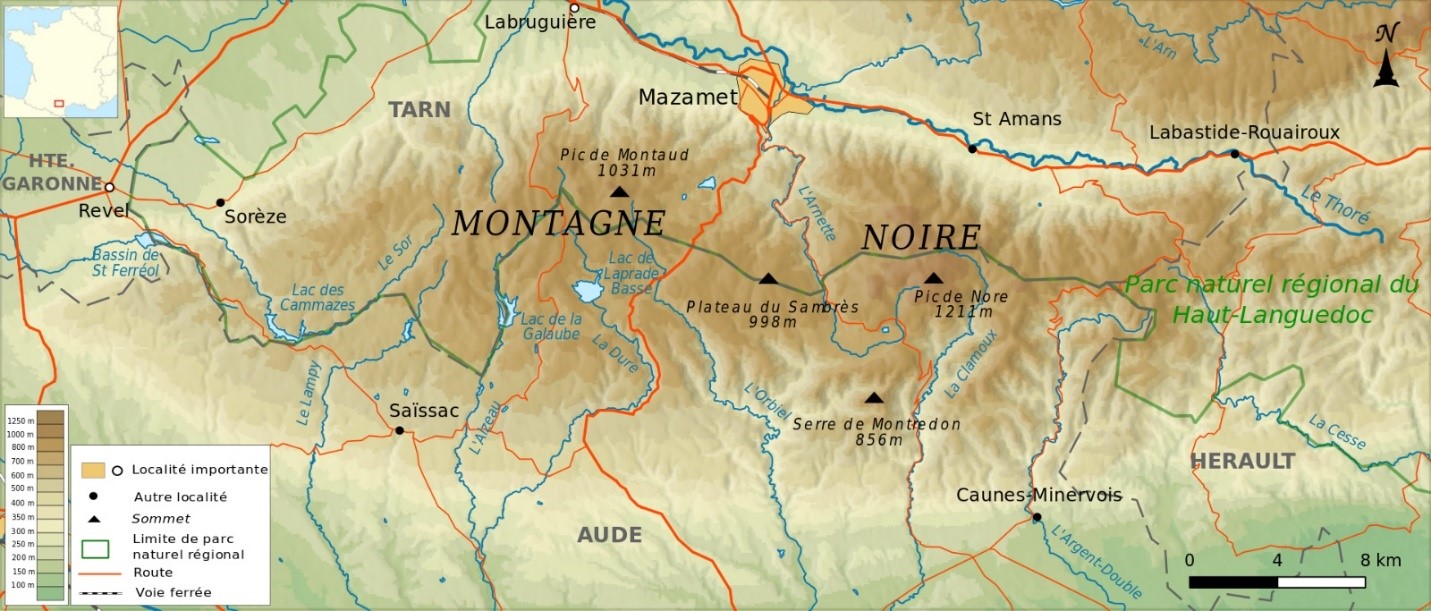
En Montagne Noire occidentale, on observe plus rarement l’Erico scopariae-Franguletum alni Géhu & Géhu-Franck 1975, ici sans doute à l’une des limites orientales de son aire générale (Géhu & Géhu, 1975 ; de Foucault & Royer, 2014) ; le tableau 5 en rapporte deux relevés, dont un inédit du Périgord dans un contexte forestier de chênaie à Quercus pyrenaica ; précédemment, il avait été noté dans le Lauragais, donc en limite occidentale de la Montagne Noire (de Foucault, 2020b).
Il faut enfin signaler la présence, au moins au voisinage de certains bâtiments agricoles, du fourré anthropophile à Sambucus nigra :
Les Martys (11), ferme ‘Les Moussels’, N 43° 25’ 30,2’’, E 2° 20’ 26,9’’, 877 m, 19/07/2022, 40 %, 30 m², 3 taxons
Sambucus nigra 2, Fraxinus excelsior j 2, Acer pseudoplatanus j 1,
à rattacher au Fraxino excelsioris-Sambucetum nigrae B. Foucault 1991 (de Foucault, 1991 ; de Foucault & Royer, 2016 ; photo 15) qui relaie à cette altitude le Fico caricae-Sambucetum nigrae B. Foucault 2015 plus méditerranéen.

Annexe du tableau 2 (tous les relevés viennent de l’Aude) – rel. 1 : Cabrespine, au-dessus du gouffre géant, 12/06/2021 ; rel. 2 : id. rel. 1, ‘Las Escoles’, 12/06/2021 ; rel. 3 : id. rel. 1, roc de l’Aigle, 12/06/2021 ; rel. 4 : id. rel. 1, nord du parking du gouffre, au-dessus de ‘La Gaougnasse’, 14/06/2022 ; rel. 5 : Lastours, pech des Châteaux, 08/2021 ; rel. 6 : Caunes-Minervois, ouest ‘Les Terralbes’, 12/10/2022.
Annexe du tableau 3 (tous les relevés viennent de l’Aude, sauf le n° 3) – rel. 1 : liste de Cabrespine, 10/06/2017 ; rel. 2 : Cabrespine, 12/06/2021 ; rel. 3 : Minerve (34), vers ‘Vieulac’, bord de la D 147, 28/05/2015 ; rel. 4 : Cabrespine, bord de la D 112, nord du moulin de Laval, 14/06/2022 ; rel. 5 : Roquefère, sur la D 8005, au niveau de ‘L’Arpent’, 31/07/2022 ; rel. 6 : id. rel. 5, sur D 8005, au niveau de ‘La Bergerie d’Airole’, 31/07/2022 ; rel. 7 : Cépie, sommet du pech ; rel. 8 : Labastide-Esparbaïrenque, entre les Artigues et le Roc d’en Galière, 29/08/2022 ; rel. 9 : Cabrespine, ‘Laval Haut’ en amont du pont sur la Clamoux, 02/06/2023.
Annexe du tableau 4 (tous les relevés viennent de l’Aude, sauf les nos 2, 3 et 9) – rel. 1 : Laprade, vers le barrage, 01/09/2021 ; rel. 2 : Lacabarède (81), parc éolien de Sauveterre, 30/09/2021 ; rel. 3 : Cassagnoles (34), sud ‘Moussou’, 30/09/2021 ; rel. 4 : id. rel. 3, vers ‘Saladou’, 30/09/2021 ; rel. 5 : La Tourette-Cabardès, nord ‘Les Griffouls’, 16/06/2022 ; rel. 6 : Les Martys, ‘Les Moussels’, 20/07/2022 ; rel. 7 : Lespinassière, entre la source de l’Argent-Double et ‘Les Pierres Blanches’, 14/07/2022 ; rel. 8 : id. rel. 7, sud-ouest ‘Les Pierres Blanches’, 14/07/2022 ; rel. 9 : id. rel. 3, entre ‘Les Pierres Blanches’ et ‘Manciès’, 14/07/2022 ; rel. 10 : Pradelles-Cabardès, ‘Las Tailladès’, 2/08/2022 ; rel. 11 : Pradelles-Cabardès, sud-est du Roc de Nouret, 15/09/2022 ; rel. 12 : Labastide-Esparbaïrenque, sud de la D 8006, 21/09/2022 ; rel. 13 : Pradelles-Cabardès, sud-ouest du Roc de Nouret, 15/09/2022 ; rel. 14 : Cuxac-Cabardès, ‘Laprade-Basse’, au-dessus du marais, sous-bois arbustif, 14/10/2019 ; rel. 15 : Pradelles-Cabardès, sous le pic de Nore, 31/07/2022 ; rel. 16 : Pradelles-Cabardès, sud du pic de Nore, bord de la D 87 ; rel. 17 : id. rel. 16, nord-ouest du pic de Nore, sud-ouest du Roc du Couillou, 15/09/2022.
Annexe du tableau 5 – rel. 1 : Saint-Denis (11), au-dessous du barrage, bord de la D 308, 01/09/2021 ; rel. 2 : Saint-Julien-de-Crempse (24), ‘Lavergne’, bord de la D 107, 21/06/2017.
5. Les lisières herbacées vivaces
Dans les lisières herbacées vivaces, on peut distinguer les ourlets et les coupes forestières. En ce qui concerne les premiers, il faut d’abord signaler celui de la chênaie verte, relié au Cephalantero rubrae-Rubietum peregrinae intraforestier :
Cabrespine (11), au-dessus de ‘Costo Granié’, N 43° 21’ 52,7’’, E 2° 28’ 18’’, 532 m, 23/04/2017, 24 taxons
- Trifolio-Geranietea sanguinei: Teucrium chamaedrys 2, Hieracium glaucinum 2, Clematis flammula 1, Rubia peregrina *p. 1, Melittis melissophyllum *m. 1, Vincetoxicum hirundinaria +, Origanum vulgare *v. +, Helleborus foetidus +, Cephalanthera longifolia +, Brachypodium sylvaticum +,
- autres taxons : Quercus ilex j 2, Pilosella officinarum 2, Asparagus acutifolius j 1, Carex halleriana 1, Lonicera implexa 1, Poa bulbosa *b. 1, Brachypodium retusum 1, Bupleurum fruticosum j 1, Aphyllanthes monspeliensis +, Globularia bisnagarica +, Coronilla minima *lotoides +, Cistus albidus j +, Cephalaria leucantha +, Teucrium polium *p. +.
Au-dessus du gouffre de la même commune, la lisière herbacée est caractérisée par Digitalis lutea, Cephalanthera rubra à nouveau, Rubia peregrina subsp. p., Origanum vulgare subsp. v., Clinopodium vulgare, Polygala vulgaris subsp. v., Galium album… ; on est loin ici des véritables Trifolio medii-Geranietea sanguinei, et de telles végétations sont trop mal connues pour d’ores et déjà donner lieu à des interprétations.
Il faut signaler ensuite celui à Lactuca plumieri (photo 16). Le tableau 6 rassemble dix relevés de cet ourlet de la hêtraie, dont un des monts de Lacaune (Tarn), ourlet qui combine surtout L. plumieri, Teucrium scorodonia, Solidago virgaurea subsp. v., Pteridium aquilinum, Avenella flexuosa subsp. f., Digitalis purpurea subsp. p., Hieracium lachenalii, Holcus mollis subsp. m., Stellaria holostea et, dans la forme typique, Linaria repens, Knautia arvernensis, Clinopodium vulgare. Il s’agit d’un ourlet acidiphile à affinités atlantiques teintées de nuances montagnardes que l’on peut dénommer Lactuco plumieri-Teucrietum scorodoniae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 6 hoc loco, retranscrit ci-dessous
Cuxac-Cabardès (11), vers ‘Le Jassot’ entre ‘Laprade-Basse’ et Lacombe, bord de la D 203, N 43° 25’ 53,7’’,
E 2° 15’ 33’’, 867 m, 01/09/2021, 70 %, 40 m², 23 taxons, photo 17
- Melampyro pratensis-Holcetea mollis: Pteridium aquilinum 2, Avenella flexuosa *f. 2, Teucrium scorodonia 1, Solidago virgaurea *v. 1, Hieracium lachenalii 1, Lactuca plumieri +, Viola riviniana +, Lonicera periclymenum *p. +, Luzula sylvatica *s. +,
- taxons différentiels du typicum: Centaurea decipiens 2, Knautia arvernensis 1, Clinopodium vulgare 1, Silene vulgaris *v. 1, Dactylis glomerata *g. +, Linaria repens +,
- autres taxons : Calluna vulgaris 1, Cruciata laevipes 1, Prunella hastifolia 1, Ranunculus bulbosus 1, Fagus sylvatica j 1, Jasione montana +, Silene nutans *n. +, Brachypodium sylvaticum +.


À côté de la forme typique, il existe une variation d’ambiance intraforestière dépourvue des taxons plus ou moins héliophiles différenciant cette dernière (relevés 9 et 10). Ce Lactuco plumieri-Teucrietum scorodoniae paraît vicariant du Tephroserido helenitidis-Knautietum arvernensis Corriol et al. 2022 décrit des monts de Lacaune et du versant tarnais de la Montagne Noire (Corriol et al., 2022).
De la Montagne Noire héraultaise, on a aussi décrit le Polygonato odorati-Paeonietum microcarpae B. Foucault 2021, magnifique ourlet qui toutefois ne passe pas dans le département de l’Aude (de Foucault, 2021d).
Les talus à fougères peuvent être aussi considérés comme des ourlets, en l’occurrence plus internes ; l’association oligotrophile la plus classique est l’Athyrio filicis-feminae-Blechnetum spicant B. Foucault 1995, présent aussi en Montagne Noire :
Les Martys (11), ‘Les Moussels’, N 43° 25’ 20,5’’, E 2° 20’ 40,9’’, 850 m, 20/07/2022, 60 %, 10 m², 6 taxons
Blechnum spicant 3, Vaccinium myrtillus 2, Juncus effusus 2, Athyrium filix-femina 1, Lonicera periclymenum *p. 1, Rubus sp. 1.
Une association venant sur humus plus actif est constituée par l’Asplenio onopteridis-Polystichetum setiferi B. Foucault 2020 :
Roquefère (11), au sud de la chapelle Saint-André, N 43° 22’ 42’’, E 2° 23’ 25,1’’, 473 m, 29/08/2022, 75 %, 10 m², 6 taxons
Polystichum setiferum 3, Hedera helix 2, Asplenium onopteris 1, Quercus ilex j 1, Erica scoparia *s. j +, Ruscus aculeatus +.
Les coupes forestières herbacées vivaces sont surtout caractérisées par Linaria repens et Digitalis purpurea subsp. p., se rattachant ainsi au Linario repentis-Digitalietum purpureae Ghestem & Descubes-Gouilly 1977 (Ghestem & Descubes-Gouilly, 1977 ; de Foucault & Catteau, 2015). Décrite initialement du Limousin, cette association à répartition atlantique se retrouve aussi dans les monts de Lacaune (Tarn ; Julve & de Foucault, 1994 : 31 ; Guitton et al., 2017a : 315). Le relevé suivant illustre la composition de cette association en Montagne Noire :
Cuxac-Cabardès (11), est du lac de Laprade-Basse, bord de la D 101, N 43° 25’ 11,6’’, E 2° 17’ 03,6’’, 758 m, 19/07/2023, 75 %, 80 m², 27 taxons
- combinaison caractéristique : Digitalis purpurea *p. 2, Teucrium scorodonia 2, Carex pilulifera *p. 2, Linaria repens 1, Avenella flexuosa *f. 2, Pteridium aquilinum 2, Hypericum pulchrum 1, Lonicera periclymenum *p. +,
- jeunes ligneux : Ilex aquifolium j +, Sorbus aria j +, Cytisus scoparius *s. j,
- autres taxons : Vinca minor 2, Rubus 2, Agrostis capillaris *c. 2, Festuca filiformis +, Veronica officinalis 1, Betonica officinalis *o. 1, Convallaria majalis +, Senecio inaequidens +, Jasione montana +, Blechnum spicant 1, Vaccinium myrtillus +, Carex laevigata +, Silene vulgaris *v. +, Solidago virgaurea *v. +, Anthoxanthum odoratum +, Calluna vulgaris +.
Dans la composante ‘jeunes ligneux’, on reconnaît la combinaison caractéristique du Sorbo ariae-Cytisetum scoparii qui envahira la coupe par dynamique végétale progressive. Cette association herbacée vivace (photo 18) est en lien spatial avec une végétation de coupe thérophytique, à valeur d’ourlet, évoquée dans un paragraphe ultérieur (§ 7).

Annexe du tableau 6 – rel. 1 et 2 : Cuxac-Cabardès (11), ‘Laprade-Basse’, non loin du barrage, 01/09/2021 ; rel. 3 : Cuxac-Cabardès (11), vers ‘Le Jassot’ entre ‘Laprade-Basse’ et Lacombe, bord de la D 203, 01/09/2021 ; rel. 4 et 5 : Pradelles-Cabardès (11), sous le pic de Nore, 13/08/2011 ; rel. 6 : Lespinassière (11), sud-est du col de Salettes en allant vers la Serre d’Alaric, 14/07/2022 ; rel. 7 : monts de Lacaune (81), relevé in Julve & de Foucault (1994 : 33, bas de page) ; rel. 8 : id. rel. 1 et 2, non loin du parking du barrage, 19/07/2023 ; rel. 9 : id. rel. 1 et 2, en sous-bois de hêtraie, 14/10/2019 ; rel. 10 : id. rel. 4 et 5, 13/08/2011.
6. Les mégaphorbiaies et les microphorbiaies
Les mégaphorbiaies ont été peu étudiées en Montagne Noire. À Cabrespine, sur les rives de la Clamoux (‘Laval Haut’), sans le relever, on a constaté la présence du Carici pendulae-Equisetetum telmateiae B. Foucault 2021 (de Foucault, 2021c).
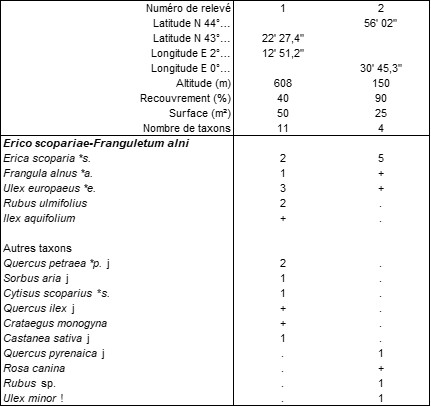
Il faut surtout signaler une autre association, caractérisée par Ranunculus aconitifolius, en ambiance plutôt forestière, vicariant altitudinal de l’Athyrio filicis-feminae-Scirpetum sylvatici B. Foucault 2011 (de Foucault, 2011b), dont le tableau 7 rapproche deux relevés et une liste floristique. Sont associés à la Renoncule Cirsium palustre, Filipendula ulmaria, Eupatorium cannabinum subsp. c., Athyrium filix-femina, Scirpus sylvaticus, Convolvulus sepium. De faibles affinités montagnardes sont affirmées par R. aconitifolius, ce qui amène à proposer le rattachement de cette mégaphorbiaie au groupe d’associations acidiphiles du Filipendulo ulmariae-Chaerophyllion hirsuti (de Foucault, 2011b), mais aucun des six syntaxons rapprochés dans ce groupe ne paraît y correspondre. On décrit ici un syntaxon nouveau, l’Eupatorio cannabini-Ranunculetum aconitifolii ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 7 hoc loco (photo 19). Le relevé 3 correspond à une variation sur substrat plus ou moins tourbeux différenciée par Molinia caerulea et Succisa pratensis.

Annexe du tableau 7 – rel. 1 : Pradelles-Cabardès (11), route du pic de Nore, 13/08/2011 ; rel. 2 : Cuxac-Cabardès (11), ‘Laprade-Basse’, atterrissement au nord-est du lac, 11/10/2021 ; rel. 3 : id. rel. 2, atterrissement au nord-ouest du lac, 10/10/2022.
Au voisinage des mégaphorbaies, on peut assez souvent observer des microphorbiaies. Elles ont été peu étudiées sur la dition, citons simplement l’observation du Cardaminetum raphanifoliae Braun-Blanq. 1948 au niveau de rochers suintants entre Miraval-Cabardès et les Martys (N 43° 24′ 01,6″, E 2° 20′ 17,2″, 555 m, 07/05/2016), avec C. raphanifolia, Chrysosplenium oppositifolium, Lotus pedunculatus, Hypericum androsaemum, H. tetrapterum, Athyrium filix-femina, Galium palustre.
7. Les ourlets à thérophytes
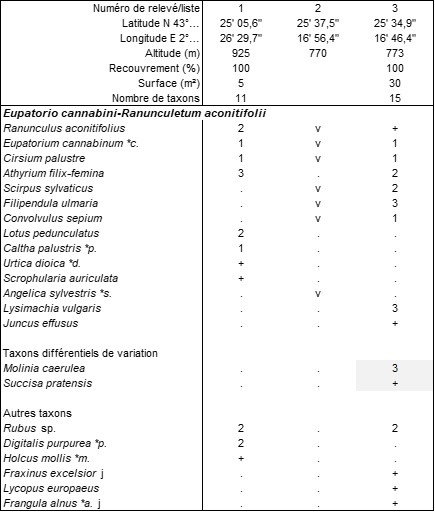
Plusieurs associations d’ourlets thérophytiques ont déjà été décrites localement, ainsi l’Arabidopsio thalianae-Myosotidetum balbisianae B. Foucault 2017, surtout cévenol mais connu aussi de Montagne Noire, à 690 m d’altitude (Fallour et al., 2013 ; de Foucault, 2017a), le Geranio lucidi-Cardaminetum hirsutae B. Foucault & Frileux 1983 vers 750 m d’altitude, le Chaerophyllo nodosi-Alliarietum petiolatae Rivas Mart. & Mayor in Fuentes 1986 vers 450 m et le Senecioni lividi-Moehringietum pentandrae B. Foucault 2017 vers 380 m. Le tableau 8 présente deux relevés inédits de ce dernier provenant du Lauragais voisin ; en ambiance plus forestière (relevé 1 de ce tableau), le Senecioni-Moehringietum pentandrae accueille des taxons plus psychrophiles comme Moehringia trinervia. Sur ces trois derniers syntaxons, voir de Foucault (2017b). Ces associations relèvent des Cardaminetea hirsutae Géhu 2000.
À plus haute altitude, à une moyenne de 730 m, la composition floristique de l’ourlet se modifie, avec la disparition des taxons plus thermophiles. Le tableau 9 en rapporte quinze relevés et une liste partielle, accueillant 5 taxons par relevé en moyenne. Cet ourlet thérophytique combine essentiellement Galeopsis ladanum, Lapsana communis subsp. c, Moehringia trinervia, Geranium robertianum et, plus rarement, Torilis arvensis subsp. anthriscoides. On le décrit sous le nom de Lapsano communis-Galeopsietum ladani ass. nov. hoc loco, typus nominis hoc loco : relevé 6 du tableau 9 hoc loco (photo 20) restranscrit ci-dessous
Lespinassière (11), sud-est du col de Salettes, N 43° 25’ 04,5’’, E 2° 34’ 31,6’’, 885 m, 14/07/2022, 35 %, 2 m², 7 taxons
Geranium robertianum 2, Galeopsis ladanum 2, G. tetrahit 1, Torilis arvensis *anthriscoides 1, Galium aparine 1, Lapsana communis *c. +, Alliaria petiolata +.
Cette association relève plus des Galeopsio tetrahit-Senecionetea sylvatici H. Passarge ex B. Foucault 2011 (de Foucault, 2011a), au sein du Lapsano communis-Geranion robertiani Tüxen & Brun-Hool 1975. Elle semble bien exister aussi dans le Sidobre comme en témoigne le relevé in Guitton et al. (2016 : 319, bas de colonne de droite), à une altitude de seulement 247 m, mais alors enrichie en taxons plus thermophiles (Geranium rotundifolium, G. purpureum, Andryala integrifolia…, ces deux derniers se retrouvant d’ailleurs aussi dans notre relevé 14) ; il s’agit sans doute d’un ourlet à distinguer du Lapsano communis-Galeopsietum ladani et à rattacher plutôt aux Cardaminetea hirsutae Géhu 2000. Le Lapsano-Galeopsietum serait alors à y rechercher à une altitude supérieure.
Les relevés/liste 1 à 15 décrivent essentiellement des ourlets linéaires et d’ailleurs pas toujours faciles à percevoir. Le relevé 16 est en revanche spatialement et largement étendu, c’est là que l’on observe Senecio sylvaticus qui en différencie une variation de coupe forestière en mosaïque avec la végétation vivace du Linario repentis-Digitalietum purpureae (cf. supra, § 5).

De ces végétations herbacées thérophytiques, on peut rapprocher la végétation commensale des cultures, notamment de céréales. En août 2011, on a parcouru une belle moisson non loin du lac de Pradelles-Cabardès, malheureusement, avec la base de loisirs qui s’est développée depuis autour du lac, il n’a pas été possible de la retrouver, on devra se contenter de la liste floristique suivante
~ N 43° 24’ 07,9’’, E 2° 26’ 19,6’’, 794 m, 13/08/2011
- composante thérophytique : Scleranthus annuus s. l., Cyanus segetum, Anthemis arvensis, Viola arvensis, Chenopodium album, Senecio vulgaris, Lapsana communis *c., Erodium cicutarium *c., Ornithopus perpusillus, Galeopsis segetum, Stellaria media, Silene gallica, Fallopia convolvulus, Capsella bursa-pastoris, Legousia speculum-veneris, Lathyrus aphaca…,
- composante vivace : Rumex acetosella,
représentant incomplet d’une végétation du Scleranthion annui (Kruseman & J. Vlieger 1939) G. Sissingh in V. Westh. et al. 1946.
Annexe du tableau 8 – rel. 1 : Labécède-Lauragais, à l’est du centre de vol à voile, sous la D 334, sous boisement ouvert, en mosaïque avec un sous-bois herbacé vivace ouvert à Conopodium majus, 13/05/2017 ; rel. 2 : id. rel. 1 mais sur un talus voisin plus éclairé.
Annexe du tableau 9 (tous les relevés viennent de l’Aude, sauf le n° 3) LGl – rel. 1 : Cuxac-Cabardès, ‘Laprade-Basse’, vers le parking du lac, bord de la D 101, 1/09/2021 ; rel. 2 : Cuxac-Cabardès, entre ‘Laprade-Basse’ et Lacombe, bord de la D 203, 01/09/2021 ; rel. 3 : Lacabarède (81), vers ‘Ravaillé’, 30/09/2021 ; rel. 4 : liste à Cabrespine, vers ‘Belle Bruyère’, 24/09/2014 ; rel. 5 : Cabrespine, vers ‘Fount Ferrouzo’, bord de la D 112, 14/06/2022 ; rel. 6 : Lespinassière, sud-est du col de Salettes, 14/07/2022 ; rel. 7 : Les Martys, près de ‘Laurens’, 18/07/2022 ; rel. 8 : Les Martys, haut de la Bézalado, 19/07/2022 ; rel. 9 : Roquefère, sur la D 8005, au niveau de ‘L’Arpent’, 31/07/2022 ; rel. 10 : Roquefère, sur la D 8005, au niveau de ‘Caberliéro’, 31/07/2022 ; rel. 11 : id. rel. 10, sur la D 8005, sous la chapelle Saint-Sernin, au niveau du départ du sentier pédestre vers Labastide-Esparbaïrenque, 31/07/2022 ; rel. 12 : Roquefère, sous Saint-Julien, 29/08/2022 ; rel. 13 : Pradelles-Cabardès, est ‘Les Jouys’, 21/09/2022 ; rel. 14 : Cabrespine, au-dessus de la D 112, ‘Lanoux’, 19/07/2023 ; rel. 15 : Villardonnel, prolongement du chemin de la Coste, au-dessus du ruisseau de Villande, 08/08/2023 ; rel. 16 : Cuxac-Cabardès (11), est du lac de Laprade-Basse, bord de la D 101.
8. Les landes
Les landes de la Montagne Noire sont favorisées par les roches acides. Parmi elles, il y a d’abord le Lavandulo stoechadis-Ericetum cinereae Géhu ex B. Foucault 2021 (de Foucault, 2020b, 2021b) qui déborde des limites de la Montagne Noire inférieure vers les plaines audoises plus méridionales sur substrats acides, jusqu’au Tarn (Julve & de Foucault, 1994 : 42). Une association voisine a été décrite de régions plus septentrionales, le Cisto salviifolii-Ericetum cinereae Guitton et al. 2017 ; pour bien les distinguer, le tableau comparatif 10 s’impose, étant ainsi construit :
- colonne CEc-a: tableau 1 in Guitton et al. (2017b, relevés 1 à 10) ;
- colonne CEc-b: tableau 2 in Guitton et al. (2016) ;
- colonnes LEc-a à -c: tableau 13 in de Foucault (2020b, resp. relevés 3 à 9, 10 à 13 et 14 à 42).
Il montre que le Lavandulo-Ericetum cinereae (LEc) se distingue positivement bien du Cisto-Ericetum cinereae (CEc) par Erica arborea j, E. scoparia subsp. s. j, Rubia peregrina subsp. p., Arbutus unedo j, Lotus dorycnium, Genista pilosa subsp. p., ainsi que des taxons différentiels du typicum (Lavandula stoechas subsp. s., Cistus albidus j, C. monspeliensis j). Ce même tableau 10 rapporte trois nouveaux relevés du Lavandulo-Ericetum cinereae.
Au sein du Cisto-Ericetum cinereae, une variation présente dans le Sidobre (colonne CEc-b) se différencie par Asphodelus albus et Leucanthemum subglaucum. En définitive, le Cisto salviifolii-Ericetum cinereae est apparemment propre à la Vendée et à une partie des Cévennes, étant relayé vers le sud par le Lavandulo-Ericetum cinereae à partir des étages inférieurs de la Montagne Noire. Toutefois, comme le montrent les deux relevés suivants, le Cisto-Ericetum cinereae paraît atteindre le Lauragais :
a) Labécède-Lauragais (11), sur l’aérodrome du centre de vol à voile de la Montagne Noire, N 43° 24’ 26’’,
E 1° 59’ 15’’, 443 m, 13/05/2017, 90 %, 15 m², 11 taxons
- Cisto salviifolii-Ericetum cinereae et unités supérieures : Calluna vulgaris 4, Erica cinerea 2, Cistus salviifolius 2, Ulex europaeus *e. j 2, Erica scoparia *s. j +,
- autres taxons : Trifolium ochroleucon 1, Pilosella officinarum 1, Festuca ovina 1, Anacamptis morio *m. 1, Scorzonera humilis +, Brachypodium rupestre *r. + ;
b) Verdun-en-Lauragais (11), sud du camping du Bout du Monde, N 43° 23’ 29,6’’, E 2° 04’ 39,9’’, 453 m, 18/09/2019, 90 %, 15 m², 8 taxons
- Cisto salviifolii-Ericetum cinereae et unités supérieures : Cistus salviifolius 3, Erica cinerea 2, Calluna vulgaris 2, Ulex europaeus *e. j 2,
- autres taxons : Silene nutans *n. 2, Rubus 1, Teucrium scorodonia 1, Cytisus scoparius *s. j +.
Dans le Cisto salviifolii-Ericion cinereae Géhu in Bardat et al. 2004, ces deux associations côtoient notamment le Festuco vasconcensis-Ericetum cinereae Géhu & Géhu-Franck 1975 et le Cisto pouzolzii-Callunetum vulgaris B. Foucault 2021.
Mais d’autres compositions floristiques sont un peu différentes, reliées à des landes d’altitude supérieure. Une des plus originales est celle décrite par les vingt relevés du tableau 11 (la date tardive de certains relevés n’a pas permis de distinguer Carex pilulifera et C. leporina en l’absence d’épis ♀). Cette lande montagnarde combine Calluna vulgaris, Genista pilosa subsp. p., Festuca arvernensis, Vaccinium myrtillus, Avenella flexuosa subsp. f., Serratula tinctoria, plus rarement Genista anglica, à une altitude moyenne de 970 m, avec une moyenne d’environ 13 taxons par relevé. On propose de la dénommer Festuco arvernensis-Callunetum vulgaris ass. nov. hoc loco, typus nominis hoc loco : relevé 19 du tableau 11 hoc loco, restranscrit ci-dessous :
Pradelles-Cabardès (11), sous le pic de Nore, N 43° 25’ 27,8’’, E 2° 26’ 59,5’’, 1 104 m, 31/07/2022, 100 %, 30 m², 14 taxons
- Festuco arvernensis-Callunetum vulgaris et unités supérieures : Calluna vulgaris 5, Festuca arvernensis 2, Genista pilosa *p. 2, Vaccinium myrtillus 2, Avenella flexuosa *f. 2, Genista anglica 1,
- taxon différentiel du typicum: Meum athamanticum 1,
- autres taxons : Cytisus scoparius *s. j 2, Agrostis capillaris 2, Solidago virgaurea *v. 1, Sorbus aria j 1, Centaurea decipiens +, Rubus +, Fagus sylvatica j +.
Cette association varie en
- typicum, différencié par Meum athamanticum, à haute altitude, surtout autour du pic de Nore, vers 1 200 m (photo 21) ;
- ericetosum cinereae subass. nov. hoc loco, différencié par Erica cinerea, Jasione montana, Campanula rotundifolia r., moins souvent Plantago holosteum, à une altitude moyenne d’environ 884 m, particulièrement répandu sur les croupes des Soulanes de Nore, ainsi colorées de rose en été (photo 22) ; typus nominis hoc loco: relevé 5 du tableau 11 hoc loco, retranscrit ci-dessous
Lespinassière (11), sud-ouest ‘Les Pierres Blanches’’, N 43° 24’ 07,3’’, E 2° 35’ 24,9’’, 887 m, 14/07/2022, 95 %, 80 m², 16 taxons
- Festuco arvernensis-Callunetum vulgaris et unités supérieures : Calluna vulgaris 4, Genista pilosa *p. 2, Avenella flexuosa *f. 2, Serratula tinctoria 2, Festuca arvernensis 1, Vaccinium myrtillus +, Genista anglica +,
- taxons différentiels de l’ericetosum cinereae: Erica cinerea 5, Jasione montana +, Campanula rotundifolia *r. +, Plantago holosteum +,
- autres taxons : Solidago virgaurea *v. 2, Carex pilulifera/leporina *l. 1, Achillea millefolium +, Sorbus aria j +, Juniperus communis *c. j.


La présence d’hémicryptophytes et géophytes dans cette dominance de chaméphytes incite à poser l’hypothèse de l’existence d’une pelouse systémiquement liée à ce Festuco-Callunetum. C’est le cas notamment pour la variation typicum : le 13 août 2011, une prospection au pic de Nore avait montré la présence d’une pelouse acidiphile à Meum athamanticum, Dianthus seguieri subsp. pseudocollinus (photo 23), Serratula tinctoria, Betonica officinalis subsp. o., Solidago virgaurea subsp. v., Succisa pratensis, Jasione laevis, Carex pilulifera subsp. p., Agrostis capillaris, Potentilla erecta, Pilosella officinarum, Avenella flexuosa subsp. f., Galium saxatile, Scorzonera humilis…, accompagnés de quelques chaméphytes plus discrets (Calluna vulgaris, Genista pilosa subsp. p., Vaccinium myrtillus) là où en 2022 on n’a observé que la lande dense. Cette pelouse nettement acidiphile rappelle le Diantho sylvatici-Meetum athamantici décrit d’Aubrac (de Foucault, 1986a), lequel est toutefois plus neutrophile et relève du Violion caninae Schwick. 1944. Julve et de Foucault (1994 : 39) ont publié un relevé complet de cette pelouse du pic de Nore avec Festuca arvernensis, Viola lutea subsp. l., Senecio doronicum. La liste suivante en offre un nouvel exemple :
Pradelles-Cabardès (11), sud du pic de Nore, N 43° 25’ 24,2’’, E 2° 27’ 44’’, 1 191 m, 15/09/2022, photo 24
- Meum athamanticum, Lotus corniculatus *c., Carex pilulifera *p., Serratula tinctoria, Potentilla erecta, Festuca arvernensis, Scorzonera humilis, Hypochaeris radicata, Nardus stricta, Crocus nudiflorus, Betonica officinalis *o.,
- chaméphytes de la lande : Vaccinium myrtillus, Calluna vulgaris, Genista pilosa *p.,
- autres taxons : Centaurea decipiens, Achillea millefolium, Teucrium scorodonia, Plantago lanceolata, Solidago virgaurea *v., Sorbus aria j…


Finalement on peut définir ici le Festuco arvernensis-Meetum athamantici ass. nov. hoc loco, typus nominis hoc loco : relevé in Julve & de Foucault (1994, Bull. Soc. Bot. N. France 47 (4), § Une pelouse acidophile…, tiers supérieur de la p. 39), en précisant Dianthus seguieri coll. en D. seguieri subsp. pseudocollinus, association pouvant se rattacher au Galio saxatilis-Potentillion aureae B. Foucault 1994 au sein des Nardetea strictae et des Festucetalia spadiceae (de Foucault, 1994, 2012).
Dans le même ordre d’idées, signalons encore la description du Violo bubanii-Prunelletum hastifoliae sur les Martys, à plus basse altitude, rattaché au Violion caninae (de Foucault, 2021b).
Entre le Lavandulo stoechadis-Ericetum cinereae et le Festuco arvernensis-Callunetum vulgaris, semble s’insérer une autre lande dont le tableau 12 rapporte cinq relevés, un provenant de l’Ardèche. Il s’agit encore d’une lande à Calluna vulgaris et Erica cinerea, mais les espèces plus thermophiles comme Lavandula stoechas et Cistus salviifolius manquent, ainsi que les taxons plus psychrophiles comme Vaccinium myrtillus, alors qu’apparaît Centaurea pectinata. On propose le nom de Centaureo pectinatae-Ericetum cinereae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 12 hoc loco (photo 25). Elle semble occuper une aire allant de la Montagne Noire aux Cévennes ardéchoises.

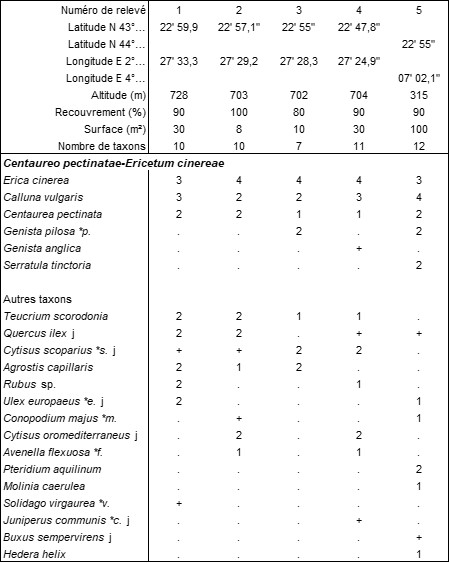
Dans la monographie des sagnes de la Bézalado et de la Grande Sagne (de Foucault, 2018b), on a enfin évoqué une lande plus ou moins turficole à Genista anglica qui se rapproche du Genisto anglicae-Callunion vulgaris Glemarec et al. La colonne 47d du tableau 13 en synthétise les sept relevés déjà connus (d’après de Foucault, 2018b, tableau 9) ; à sa droite, les relevés 1 et 2 sont inédits, alors que les colonnes 47a à 47c synthétisent les trois variations du Molinio caeruleae-Ericetum tetralicis (Géhu & Wattez 1975) Glemarec, Delassus & Boullet, que les colonnes 48a à 48c synthétisent les trois variations du Carici trinervis-Callunetum vulgaris B. Foucault & Géhu 1978 (ces deux syntaxons d’après Glemarec et al., à paraître) et que la colonne HGa synthétise l’Helictochloo marginatae-Genistetum anglicae B. Foucault 2019 du pays de Sault (d’après de Foucault, 2019, tableau 16). On voit que les landes turficoles à G. anglica de Montagne Noire (colonnes 47d, relevés 1 et 2) se rapprochent du Molinio caeruleae-Ericetum tetralicis sans s’y rattacher (absence significative de Festuca filiformis, Erica tetralix, Agrostis canina, Ulex europaeus subsp. e. j, alors que Gentiana pneumonanthe, Scorzonera humilis et Juniperus communis subsp. c. l’en différencient faiblement), de sorte qu’on peut définir ici le Molinio caeruleae-Genistetum anglicae ass. nov. hoc loco, typus nominis hoc loco : relevé 52 du tableau 9 in de Foucault (2018b, Evaxiana 4 : 169).
Il faut enfin évoquer le classique Lonicero periclymeni-Vaccinietum myrtilli B. Foucault 1994, une lande intra- à subforestière (photo 8) dont le tableau 14 rapporte deux nouveaux relevés, le n° 1 correspondant au typicum intraforestier, le n° 2 au callunetosum vulgaris subforestier, en clairière, marqué par la régression quantitative de V. myrtillus et l’apparition de chaméphytes des landes plus héliophiles.
Annexe du tableau 10 (tous les relevés viennent de l’Aude) – rel. 1 : Labastide-Esparbaïrenque, sud du Roc d’En Galière, chemin des Villages Perchés, 29/08/2022 ; rel. 2 : id. rel. 1, entre la Madeleine et le Vigné, 29/08/2022 ; rel. 3 : Fournes-Cabardès, vers ‘Sériès’, sud de la D 401, 18/01/2023.
Annexe du tableau 11 (tous les relevés vienent de l’Aude, sauf les nos 9 et 10) – rel. 1, 2 : Lespinassière, sud de la source de l’Argent-Double, 14/07/2022 ; rel. 3 : id. rel. 1 et 2, entre la source et ‘Les Pierres Blanches’, 14/07/2022 ; rel. 4 : id. rel. 1 et 2, près de ‘Les Pierres Blanches’’, 14/07/2022 ; rel. 5 : id. rel. 1 et 2, sud-ouest ‘Les Pierres Blanches’’, 14/07/2022 ; rel. 6, 7 : id. rel. 1 et 2, sud de la route forestière des Rives Hautes, 14/07/2022 ; rel. 8 : id. rel. 1 et 2, nord-ouest ‘Les Pierres Blanches’, 14/07/2022 ; rel. 9 : Cassagnoles (34), entre ‘Les Pierres Blanches’ et ‘Mancès’, 14/07/2022 ; rel. 10 : id. rel. 9, sud-est de la source de l’Argent-Double, 14/07/2022 ; rel. 11 : Cabrespine, vers le Roc de l’Aigle, 10/06/2017 ; rel. 12 : Miraval-Cabardès, Roc du Bougre, 26/09/2018 ; rel. 13 : Pradelles-Cabardès, Roquo d’Astié, 15/09/2022 ; rel. 14 : Pradelles-Cabardès, vers le cairn Jean-Ané, 21/09/2022 ; rel. 15 à 19 : Pradelles-Cabardès, différents points autour du pic de Nore, 31/07/2022 ; rel. 20 : Pradelles-Cabardès, sous le Roc de Nouret, 15/09/2022.
Annexe du tableau 12 (tous les relevés viennent de l’Aude, sauf le n° 5) – rel. 1 à 4 : Cabrespine, chemin du Château, ouest de la D 112, 19/07/2023 ; rel. 5 : Brahic (07), route de Malbosc, sud du Mas de l’Ayre, bord de la D 216, 09/09/2020.
Annexe du tableau 13 – rel. 1 : Les Martys (11), ‘Les Moussels’, 20/07/2022 ; rel. 2 : Les Martys (11), la Sagne Grande, 19/07/2022.
Annexe du tableau 14 – rel. 1 : Les Martys (11), ‘La Bézalado’, lande intraforestière d’une boulaie oligotrophile, 11/07/2016 ; rel. 2 : Les Martys (11), nord du lac de Laprade-Basse, clairière forestière, 10/10/2022.
9. Les tonsures à thérophytes
La végétation des tonsures à thérophytes se différencie essentiellement selon la topographique. Ainsi, les bas niveaux assez longuement inondables et plutôt eutrophes se rencontrent notamment sur les berges de lacs ou réservoirs (Laprade-Basse, Les Cammazes…). Le tableau 15 rapporte six relevés de cette rare végétation paucispécifique à phénologie tardi-estivale à automnale, caractérisée par Corrigiola littoralis, Gnaphalium uliginosum, Laphangium luteoalbum, Cyperus fuscus, Juncus bufonius, J. tenageia, à rapprocher de l’Eleocharition ovatae G. Phil. 1968 (de Foucault, 2013, sub Eleocharition soloniensis), le Gnaphalio uliginosi-Corrigioletum littoralis ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 15 hoc loco (photo 26).
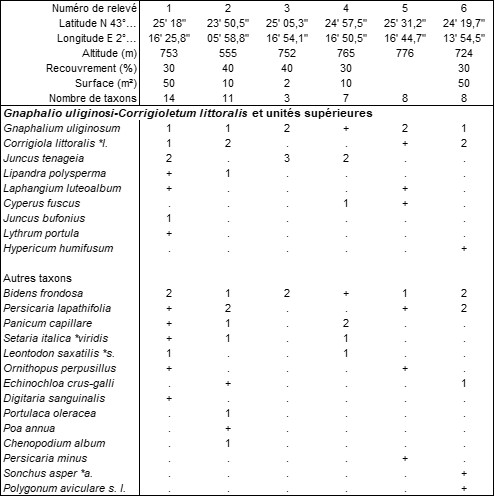
Aux Cammazes, cette végétation thérophytique est associée à la végétation vivace suivante : Mentha pulegium 2, Plantago major s. l. 2, Equisetum arvense 1, Senecio inaequidens 1, nouveau représentant du Plantagini majoris-Menthetum pulegii B. Foucault in J.-M. Royer et al. 2006 (photo 27).


Dans les niveaux topographiques moyens, sous le pic de Nore (commune de Pradelles-Cabardès, 11), à deux reprises (étés 2011 et 2022) a été observée une végétation paucispécifique à Juncus bufonius et Illecebrum verticillatum, parfois Hypericum humifusum, Ornithopus perpusillus ; elle peut se rattacher aux Nanocyperetalia flavescentis sans qu’il soit possible de descendre plus bas dans la hiérarchie synsystématique.
Les niveaux supérieurs, guère marqués par l’eau, sont le lieu de prédilection des pelouses thérophytiques xérophiles à Tuberaria guttata. Elles n’ont pas été étudiées en détail, deux relevés de Montagne Noire audoise sont rapprochés dans le tableau 16 en 1 et 2. Ces relevés se rattachent au groupe des pelouses de corniche rocheuse sur sol rocailleux à Micropyrum tenellum, qui s’oppose à d’autres pelouses thérophytiques voisines mais développées sur sol sablonneux ; ce groupe compte jusqu’à présent six syntaxons rapprochés dans ce tableau synthétique 16 :
- colonne A: Micropyretum tenelli Korneck 1975 mut. hoc loco, eu- à subatlantique ; Korneck (1975, synthèse des trois colonnes du tableau 1, sub Narduretum lachenalii) ; de Foucault (1979, tableau 3 ; 1981, tableau III) ;
- colonne B: Vulpio muyri-Micropyretum tenelli Felzines & Loiseau 2004 inv. (Felzines & Loiseau, 2004, tableau IX), subatlantique ;
- colonne C: Arnoserido minimae-Micropyretum tenelli Foucault 1999 nom. mut. hoc loco (initialement sub Arnoserido minimae-Narduretum aristati), thermo-atlantique (de Foucault, 1988, tableau 10, sub Narduretum lachenalii ; Julve & de Foucault, 1994, liste p. 42 ; de Foucault, 2017a, tableau 4 ; Guitton et al., 2016, tableau 6 : relevés J2/01 à J2/06) ;
- colonne D: groupement à Periballia involucrata (de la Fuente, 1985, tableau 26, sub Trisetario ovatae-Agrostietum truncatulae) ;
- relevés 1 et 2 : deux relevés inédits franco-méditerranéens ;
- colonne E: groupement à Micropyum tenellum (López, 1978, tableau 29) ;
- colonne F: groupement à Aira cupaniana (Rivas Goday, 1958, tableau 12 : relevés 5 à 10).
Ces associations se rattachent soit au Thero-Airion praecocis Tüxen ex Oberd. 1957 (A et B), soit au Tuberario guttatae-Airion praecocis B. Foucault 1999 (C et D), soit au Tuberarion guttatae Braun-Blanq. in Braun-Blanq. et al. 1940 (1, 2 et F) (sur ces alliances, voir des données récentes in de Foucault, 2023a) ; l’association de Montagne Noire (1 et 2) peut être décrite sous le nom de Tolpido umbellatae-Micropyretum tenelli ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 16 hoc loco, retranscrit ci-dessous :
au-dessus de Roquefère (11), N 43° 22’ 28,8’’, E 2° 22’ 45,6’’, ~ 341 m, 30/05/2015, 35 %, 2 m², 11 taxons
- Tuberarietea guttatae: Micropyrum tenellum 2, Tolpis umbellata 2, Trifolium arvense 2, Briza maxima 2, Leontodon saxatilis *rothii 2, Vulpia bromoides 1, Ornithopus compressus 1, Logfia minima 1, Aira caryophyllea 1, Filago germanica +,
- autre taxon : Andryala integrifolia
Le relevé R13 du tableau 3 publié par Guitton et al. (2017a) et provenant des monts de Lacaune correspond aussi à cette association. Rappelons que Felzines et Loiseau (2004) ont défini une sous-alliance Micropyrenion tenelli au sein du Thero-Airion praecocis ; la large extension de ces végétations à Micropyrum tenellum à travers trois alliances de la classe des Tuberarietea guttatae laisse cependant planer un doute sur l’intérêt de cette petite unité. Il est clair qu’une large synthèse de cette classe sera bien utile.
Il existe aussi des pelouses thérophytiques xérophiles relevant du Tuberarion guttatae mais dépourvues de M. tenellum :
Villardonnel (11), ‘Les Combals’, N 43° 20’ 06,9’’, E 2° 18’ 02,9’’, 372 m, 26/05/2020, 21 taxons
- Tuberarietea guttatae: Ornithopus compressus 2, Vulpia ciliata s. l. 2, Filago germanica 2, Lupinus angustifolius 2, Logfia gallica 2, L. minima 1, Aira caryophyllea 1, armoricana 1, Tolpis umbellata 1, Lotus angustissimus 1, Tuberaria guttata +, Aphanes australis +, Trifolium arvense +, Silene gallica +, Briza maxima +,
- autres taxons : Andryala integrifolia 2, Trifolium campestre +, Anthemis arvensis +, Galactites tomentosa +, Ervilia hirsuta +, Briza minor +.
Sur calcaires dévoniens, par exemple aux environs de Cabrespine, la végétation xérophile thérophytique relève des Stipo capensis-Brachypodietea distachyi Brullo in Brullo et al. 2001 ; l’unique relevé suivant en offre un exemple :
Cabrespine (11), chemin de Brescou, N 43° 21’ 49,7’’, E 2° 28’ 13,7’’, 516 m, 23/04/2017, 10 taxons
Brachypodium distachyon 3, Helianthemum salicifolium 2, Galium parisiense 2, Valantia muralis 2, Lysimachia linum-stellatum 1, Euphorbia exigua 1, Saxifraga tridactylites 1, Chaenorrhinum rubrifolium *r. 1, Minuartia hybrida *laxa +, Hornungia petraea +.
Annexe du tableau 15 – rel. 1 : Cuxac-Cabardès (11), lac de Laprade-Basse, rive nord vers l’aire de pique-nique, 26/09/2019 ; rel. 2 : Les Cammazes (81), rive sud du lac, sous la D 629, 18/09/2019 ; rel. 3 et 4 : id. rel. 1, nord-est du barrage, plage de la Dure, 01/09/2021 ; rel. 5 : id. rel. 1, 10/10/2022 ; rel. 6 : Lacombe (11), rive orientale de la Galaube.
Annexe du tableau 16 – rel. 1 : au-dessus de Roquefère (11), 30/05/2015 ; rel. 2 : Villardonnel (11), la Barthe Haute.
10. Les murs et les rochers
Les rochers affleurants offrent divers habitats originaux, qu’ils soient naturels ou qu’ils soient structurés en murs. Tout d’abord, les fentes des pans subverticaux acides accueillent une végétation chasmophile souvent riche en ptéridophytes. Ainsi, le tableau 17 illustre une végétation à Asplenium foreziense que l’on peut rattacher à l’Asplenietum septentrionali-foreziensis Mateo 1983 ; les relevés 1 à 4 reprennent le tableau 28 de Mateo Sanz (1983), les relevés 5 à 7 sont inédits [rel. 5 : Cabrespine (11), ‘Maïssantoul’, rive gauche du ruisseau de Serremijeanne, le 23/04/2017 ; rel. 6 : Roquefère (11), sur la D 8005, vers l’Arpent, 29/08/2022 ; rel. 7 : id. rel. 5, mur entre ‘Laval-Haut’ et le Cabanial, 02/06/2023], le relevé 8 est la reprise du relevé J1/21 du tableau 8 in Guitton et al. (2016, du Sidobre). On note que la composante française de cette végétation (photo 28) se distingue cependant de la composante ibérique par l’absence de Sedum brevifolium et Poa flaccidula, ce dernier taxon absent de l’Aude.

À côté de cette association à A. foreziense, le même tableau 17 fournit en plus cinq relevés (nos 9 à 13) de l’Asarino procumbentis-Sedetum hirsuti B. Foucault 2017 initialement décrit des Cévennes (de Foucault, 2017a ; le relevé 9 est la reprise du relevé 12 du tableau 1 de cette publication, de Montagne Noire : Mas-Cabardès, chemin des Douilhos), localement enrichi en Micranthes clusii (photo 29). Le relevé 13 correspond à une variation marquée par une légère eutrophisation favorisant Parietaria judaica (photo 30).


Récemment, en Montagne Noire, on a aussi défini l’Umbilico rupestris-Cystopteridetum fragilis (de Foucault, 2022b), dont le tableau 17 rapporte quatre nouveaux relevés (nos 14 à 17 ; photo 31). Le relevé 14 correspond à une variation marquée par une légère eutrophisation favorisant Cymbalaria muralis.

Non loin du relevé 11 de ce tableau 17 (voir localisation spatio-temporelle à l’annexe de ce tableau), un rocher suintant a donné lieu au relevé suivant :
N 42° 24’ 00,4’’, E 2° 20’ 17,1’’, 551 m, 02/06/2023, 10 m², 50 %, 5 taxons
Hypericum androsaemum 2, H. tetrapterum 2, Athyrium filix-femina 2, Lotus pedunculatus 2, Eupatorium cannabinum *c. 1 ;
par sa situation et son écologie, il rappelle l’Hyperico androsaemi-Adiantetum capilli-veneris, toutefois sans le second taxon éponyme, qui paraît manquer en Montagne Noire (Barreau, 2019).
La plupart des associations précédentes sont développées sur roches acides. Sur les substrats rocheux où se mêlent des matériaux siliceux et des calcaires, par exemple aux environs de Cabrespine, on a observé une végétation assez différente :
Cabrespine (11), chemin de Saint-Martin, est ‘La Fraou’, N 43° 20’ 41,2’’, E 2° 27’ 40,2’’, 462 m, 29/09/2019, 30 %, 10 m², 8 taxons
- combinaison caractéristique : Plantago holosteum 2, Saxifraga fragosoi +, Sedum hirsutum 1, album +, Dianthus pungens +,
- autres taxons : Buxus sempervirens j 1°, Senecio inaequidens 1, Cistus albidus j +.
Complétons toutes ces associations par l’Asarino procumbentis-Parietarietum judaicae B. Foucault 2017 présent sur les murs anthropisés de plusieurs villages, décrit initialement des Cévennes et de Montagne Noire, puis retrouvé dans les Albères et en Cerdagne ; le relevé suivant en offre un nouvel exemple :
Labastide-Esparbaïrenque (11), nord de la rue du Tint, N 43° 22’ 59,1’’, E 2° 23’ 42,4’’, 406 m, 29/08/2022, 60 %, 5 m², 6 taxons
- combinaison caractéristique et Parietarietea judaicae: Umbilicus rupestris 3, Parietaria judaica 2, Asarina procumbens +,
- autres taxons : Epilobium lanceolatum 1, Pseudoturritis turrita 1, Senecio inaequidens
Riche en taxons xérophiles, dont des Crassulaceae, la végétation chomophytique est celle qui colonise les dalles et les pointements rocheux subhorizontaux. Ainsi, à Cabrespine (11 ; le 17/06/2022), quelques dalles plus ou moins siliceuses sont colonisées par Sedum anglicum, Plantago holosteum, Jasione montana, Rumex acetosella, Anthemis cretica subsp. saxatilis, Centaurea pectinata, Scleranthus perennis, parfois quelques chaméphytes issus de landes de contact (Erica cinerea, Calluna vulgaris) ; une telle végétation pourvue d’A. cretica subsp. saxatilis ne semble pas apparaître dans la synthèse récente de Royer & Ferrez (2018). Il existe bien par ailleurs l’Anthemido saxatilis-Plantaginetum holostei B. Foucault, Menand & Corriol in Corriol et al. 2021, mais cette association manque de Sedum anglicum (Corriol et al., 2021). Le contact thérophytique est à base de Micropyrum tenellum, Logfia gallica, Hypochaeris glabra… (probablement un fragment de l’Arnoserido minimae-Micropyretum tenelli B. Foucault 1999).
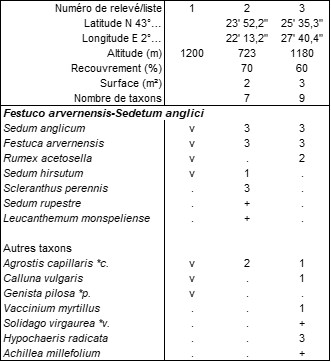
Mieux caractérisée est la pelouse vivace décrite par les relevés et listes du tableau 18, caractérisée par la combinaison de Sedum anglicum (photo 32), S. hirsutum, Festuca arvernensis, Rumex acetosella, développée sur les hauteurs du massif, jusqu’au pic de Nore. Cette combinaison paraît exister aussi dans le Tarn, au Roc de Montalet, autour de 1 200 m, avec un Festuca incomplètement indéterminé (F. gr. ovina ; Julve & de Foucault, 1994 : 35). Pour apprécier son originalité, le tableau 19 reprend le tableau 2 de la synthèse de Royer & Ferrez (2018) actualisé, modifié et complété (le syntaxon n° 11 de cette synthèse n’y est vraiment pas à sa place et n’est pas repris ici) ; il fait apparaître deux groupes d’associations (sauf précisions, ces données sont reprises de la synthèse de 2018) :
- le premier surtout différencié par Hypericum linariifolium et Prospero autumnale, des plaines et collines
- Hyperico linariifolii-Sedetum reflexi B. Foucault 1979.
- Hyperico linariifolii-Sedetum anglici B. Foucault ex B. Foucault, J.-M. Royer & Ferrez 2018.
- Scillo autumnalis-Ranunculetum paludosi B. Foucault 1988 ex 2008, qui peut être muté en Prosperini autumnalis-Ranunculetum paludosi nom. mut. hoc loco.
- Plantagini holostei-Sesamoidetum purpurascentis B. Foucault 1988 ex 2008.
- Festuco trachyphyllae-Sedetum anglici Clément & Touffet 1978 ; la détermination du premier taxon éponyme devrait être revue.
- Festuco tenuifoliae-Sedetum anglici Clément & Touffet 1978 qui peut être muté en Festuco filiformis– Sedetum anglici nom. mut. hoc loco.
- Sedo anglici-Scilletum vernae Bioret 1994 qui peut être muté en Sedo anglici-Tractemetum vernae nom. mut. hoc loco.
- Dactylido marinae-Sedetum anglici Géhu, Géhu-Franck & Caron 1978.
CSa. Corynephoro canescentis-Sedetum anglici B. Foucault 1999 (Géhu, 1964, tableau 4, composante vivace).
- « Tuberario maritimae-Romuleetum columnae G. Lemée ex Provost 1978 », dont la composante vivace a été distinguée en Romuleo coronatae-Sedetum anglici B. Foucault ex Provost & B. Foucault in B. Foucault 2018.
- le second surtout différencié par Sedum hirsutum et Scleranthus perennis, plus alticole
- « Spergulo morisonii-Sedetum hirsuti Billy ex Thébaud, Ca. Roux, Bernard & Delcoigne 2014 ».
FSa. le syntaxon du tableau 18 hoc loco.
L’association FSa de la Montagne Noire est donc suffisamment originale pour être distinguée comme Festuco arvernensis-Sedetum anglici ass. nov. hoc loco, typus nominis hoc loco : relevé 2 du tableau 18 hoc loco (photo 33).


Le « Spergulo morisonii-Sedetum hirsuti Billy ex Thébaud et al. 2014 » d’Auvergne (Billy, 2002, tableau p. 48 : colonne 1 ; Thébaud et al., 2014) est un ensemble biologiquement complexe où la composante thérophytique est formée de Spergula morisonii (IV sur six relevés), Micropyrum tenellum (III), Hypochaeris glabra (II), Logfia minima (II), Teesdalea nudicaulis (II) ; d’après la synthèse précédente de la végétation thérophytique à M. tenellum (§ 9 et tableau 16), cette dernière coïncide avec le Micropyretum tenelli Korneck 1975. La composante vivace se distingue de notre pelouse de Montagne Noire par Festuca filiformis au lieu de F. arvernensis et l’absence de S. anglicum. On peut renommer la pelouse d’Auvergne en Festuco filiformis-Sedetum hirsuti Billy ex ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 718 du tableau X in Billy (2002, Bull. Soc. Bot. Centre-Ouest, n. s., n° sp. 22 : 168).
Mais surtout, la Montagne Noire est par ailleurs le domaine d’une espèce endémique, Armeria malinvaudii (photo 34), dont l’aire s’étend sur l’Aude nord-orientale et l’Hérault. Dans le tableau 20 sont réunis neuf relevés de cette végétation originale dans laquelle A. malinvaudii est souvent accompagné de Festuca arvernensis et Plantago holosteum ; étant topographiquement associée au Festuco arvernensis-Callunetum vulgaris ericetosum cinereae (photo 35), cette pelouse accueille aussi divers chaméphytes de cette lande. Étonnamment, on n’y a pas observé Rumex acetosella, mais ce taxon est peut-être présent en d’autres stations de l’Armeria. En moyenne, on compte environ 7,5 taxons par relevé, à une altitude moyenne de 910 m. Cette association originale peut être dénommée Armerio malinvaudii-Festucetum arvernensis ass. nov. hoc loco, typus nominis hoc loco : relevé 9 du tableau 20 hoc loco (photo 36 ; voir aussi photo in Plassart et al., 2016 : 123). L’injection de ce tableau 20 dans le tableau synthétique 1 de la synthèse de Royer & Ferrez (2018) mène à inclure cette association dans le Sedo albi-Scleranthion biennis, ce qui est conforme avec l’altitude moyenne des relevés.
En définitive, le Festuco arvernensis-Sedetum anglici occupe la partie occidentale du massif, alors que l’Armerio malinvaudii-Festucetum arvernensis en occupe la partie orientale.



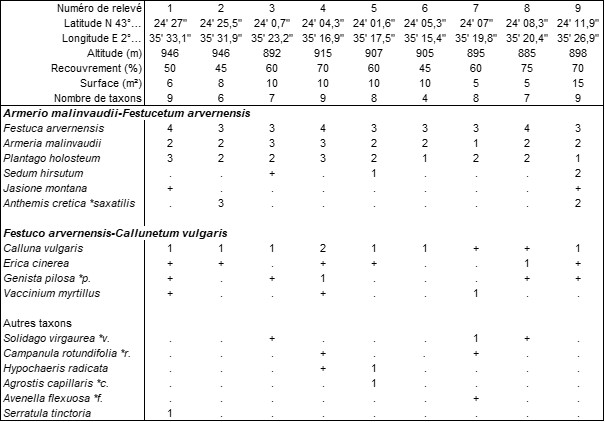
Sur les affleurements calcaires, la végétation chomophytique est évidemment assez différente de celle observée sur roches acides, bien plus fréquentes, mais elle n’a été jusqu’à présent que très imparfaitement étudiée.
Au-dessus du gouffre de Cabrespine, il existe par exemple quelques dalles calcaires à Sedum album, S. sediforme, S. ochroleucum, Helianthemum apenninum, Lactuca perennis (alliance du Sedion micrantho-sediformis au sein des Sedo albi-Scleranthetea biennis).
Au-dessus du même village, tant du côté ouest (au-dessus du gouffre) que du côté est (au-dessus de ‘Châlet’), s’étendent de belles dalles sur calcaire dont le tableau 21 rapproche quatre relevés qui partagent Helianthemum canum, Thymus vulgaris subsp. v., Helichrysum stoechas subsp. s., Ononis minutissima, Fumana ericifolia, Globularia bisnagarica, Carex halleriana. Il montre qu’on peut opposer le relevé 4 (qui semble plutôt se rattacher à l’Helianthemo–Aphyllanthion monspeliensis Díez Garretas et al. 1998) aux relevés 1 à 3, ces derniers se distinguant par Arenaria aggregata subsp. a. (photo 37), Iberis saxatilis subsp. s. (photo 38), Trinia glauca subsp. g. (photo 39), Galium corrudifolium, Helianthemum apenninum, Inula montana, Koeleria vallesiana subsp. v., Teucrium aureum, Anthyllis montana subsp. m. (photo 40), Minuartia capillacea (taxon patrimonial pour l’Aude selon Plassart et al., 2016 ; photo 41), Ononis striata (photo 42)… Cette combinaison de taxons oriente nettement cette végétation vers les Ononidetalia striatae Braun-Blanq. et al. ex Braun-Blanq. 1950 et sans doute l’Ononidion striatae Braun-Blanq. & Susplugas 1937, bien qu’aucune des associations connues de cette alliance n’y corresponde vraiment, y compris l’association type de l’alliance, le Genisto corbariensis-Seslerietum caeruleae Braun-Blanq. & Susplugas 1937 décrit des hautes Corbières (Braun-Blanquet & Susplugas, 1937, sub Genisto corbariensis-Seslerietum elegantissimae ; Royer & Ferrez, 2019). Il semble donc bien que l’association définie par les relevés 1 à 3 du tableau 21, développée autour de 630 m d’altitude en moyenne, avec une moyenne de 23 taxons par relevé, soit originale et puisse être décrite sous le nom de Trinio glaucae-Arenarietum aggregatae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 21 hoc loco, retranscrit ci-dessous
Cabrespine (11), col de la Condamine, N 43° 21’ 58,8’’, E 2° 28’ 31,7’’, 611 m, 23/04/2017, 26 taxons
- Ononidion striatae et unités supérieures : Ranunculus gramineus 3, Iberis saxatilis *s. 2, Galium corrudifolium 2, Helianthemum apenninum 2, canum 2, Valeriana tuberosa 2, Sedum ochroleucum 2, Teucrium chamaedrys 2, Narcissus assoanus 2, Poa bulbosa *b. 2, Thymus vulgaris *v. 2, Helichrysum stoechas *s. 2, Inula montana 2, Arenaria aggregata *a. 1, Allium sphaerocephalon *s. 1, Anthyllis montana *m. 1, Globularia cordifolia 1, Trinia glauca *g. +, Ononis minutissima +, Festuca lemanii 1, Carex halleriana 1, C. humilis +,
- autres taxons : Anacamptis morio *m. +, Linum usitatissimum *angustifolium +, Muscari neglectum 1, Quercus ilex j +.






Annexe du tableau 17 – rel. 1 à 4 : Mateo-Sanz (1983), tableau 28 ; rel. 5 : Cabrespine (11), ‘Maïssantoul’, rive gauche du ruisseau de Serremijeanne, le 23/04/2017 ; rel. 6 : Roquefère (11), sur la D 8005, vers l’Arpent, 29/08/2022 ; rel. 7 : id. rel. 5, mur entre ‘Laval-Haut’ et le Cabanial, 02/06/2023 ; rel. 8 : reprise du relevé J1/21 du tableau 8 in Guitton et al. (2016, du Sidobre) ; rel. 9 : reprise du relevé 12 du tableau 1 in de Foucault (2017a) ; rel. 10 : Cabrespine (11), Sermijeanne, route de la Clamoux, 23/04/2017 ; rel. 11 : La Tourette-Cabardès (11), sur la D 9, 02/06/2023 ; rel. 12 : Miraval-Cabardès (11), sur la D 101, Le Campmas, 02/06/2023 ; rel. 13 : id. rel. 10, ‘Laval’, 02/06/2023 ; rel. 14 : Caudebronde (11), impasse Font d’Izard, 02/06/2023 ; rel. 15 : id. rel. 14, sous la route de la Baute, 02/06/2023 ; rel. 16 : id. rel. 14, entrée du village sur la D 662, 02/06/2023 ; rel. 17 : Cuxac-Cabardès (11), Laprade-Basse, mur près de l’église Saint-Roch-en-Cabardès, 19/07/2023.
Annexe du tableau 18 – rel. 1 : liste au pic de Nore, 31/08/2011 ; rel. 2 : Roquefère (11), ‘Cubserviès’, au nord de la cascade, 30/05/2015 ; rel. 3 : Pradelles-Cabardès (11), pic de Nore, 19/07/2023.
Annexe du tableau 20 – Divers relevés effectués au sud-est du col de Salettes, Lespinassière (11) et Cassagnoles (34), 14/07/2022.
Annexe du tableau 21, tous les relevés viennent du territoire de la commune de Cabrespine (11) – rel. 1 : au-dessus du gouffre, sous le Roc de l’Aigle, 12/06/2021 ; rel. 2 : id. rel. 1, vers le sommet du Roc de l’Aigle, 12/06/2021 ; rel. 3 : col de la Condamine, 23/04/2017 ; rel. 4 : chemin de Brescou, 23/04/2017.
11. Les bas-marais acidiphiles
En achevant la monographie phytosociologique de la sagne de la Bézalado (de Foucault, 2018b, 2022b), on concluait sur la valeur de modèle de ce système tourbeux montagnard pour une bonne partie de la Montagne Noire et des massifs voisins (Sidobre, monts de Lacaune). De fait, cette sagne offre une belle diversité d’associations oligotrophiles à caractère atlantique comme le montre la liste synsystématique suivante : Trocdario verticillati-Juncetum acutiflori (G. Lemée 1937) Oberd. ex B. Foucault 2023 (présent aussi dans le Tarn : Julve & de Foucault, 1994 ; Guitton et al., 2017a), Trocdario verticillati–Molinietum caeruleae (G. Lemée 1937) J.-M. Royer, Felzines, Misset & Thévenin 2006, Trocdario verticillati-Narthecietum ossifragi B. Foucault 2018, Trocdario verticillati-Juncetum squarrosi B. Foucault & Philippe ex B. Foucault & Philippe in J.-M. Royer et al. 2006, Carici binervis-Agrostietum caninae B. Foucault 2008 (pour l’autorité des deux premières associations, voir de Foucault, 2023d). Le relevé suivant offre un nouvel exemple de Trocdario-Narthecietum ossifragi :
Les Martys (11), ‘Les Moussels’, N 43° 25’ 20,5’’, E 2° 20’ 40,9’’, 850 m, 20/07/2022, 90 %, 60 m², 29 taxons
- Trocdario verticillati-Narthecietum ossifragi et unités supérieures : Narthecium ossifragum 3, Molinia caerulea 3, Eriophorum angustifolium *a. 3, Rhynchospora alba 2, Viola palustris 2, Succisa pratensis 2, Lysimachia tenella 2, Trocdaris verticillatum 2, Carex panicea 2, echinata *e. 2, C. demissa 1, Drosera rotundifolia 1, Juncus acutiflorus +, Dactylorhiza maculata +, Wahlenbergia hederacea +, Potentilla erecta +,
- autres taxons : Genista anglica 2, Potamogeton polygonifolius 2, Calluna vulgaris 1, Eleocharis multicaulis 1, Salix atrocinerea j 1, aurita j +, Cirsium palustre +, Briza media *m. +, Lotus pedunculatus +, Polygala serpyllifolia +, Juncus bulbosus +, Holcus lanatus *l. +, Betula pendula j +,
alors que le suivant illustre le Trocdario verticillati-Juncetum acutiflori
Les Martys (11), ‘Les Moussels’, N 43° 25’ 20,5’’, E 2° 20’ 40,9’’, 850 m, 20/07/2022, 90 %, 60 m², 26 taxons
- Trocdario verticillati-Juncetum acutiflori et unités supérieures : Molinia caerulea 4, Juncus acutiflorus 3, Eriophorum angustifolium *a. 2, Viola palustris 2, Trocdaris verticillatum 2, Wahlenbergia hederacea 2, Carex panicea 2, echinata *e. 2, C. demissa 1, Succisa pratensis 1, Potentilla erecta 1, Dactylorhiza maculata 1, Luzula multiflora *m. 1, Lysimachia tenella 1, Agrostis canina 1, Scorzonera humilis +, Scutellaria minor +, Narthecium ossifragum +,
- autres taxons : Calluna vulgaris 2, Genista anglica 1, Lotus pedunculatus 1, Holcus lanatus *l. +, Cirsium palustre +, Briza media *m. +, Salix atrocinerea j +, Angelica sylvestris *s. +.
Sous le pic de Nore (commune de Pradelles-Cabardès), donc vers 900 à 1 000 m d’altitude, le 15 septembre 2022, on a aussi observé en bordure de piste une pelouse mésohygrophile sur substrat plus ou moins tourbeux tassé rappelant le Trocdario verticillati-Juncetum squarrosi avec les taxons suivants : Carex binervis, Juncus squarrosus, Danthonia decumbens subsp. d., Nardus stricta, Agrostis capillaris, Anthoxanthum odoratum, Calluna vulgaris, Erica cinerea. Le relevé suivant en offre un exemple plus précis :
Pradelles-Cabardès (11), pic de Nore, bord de la D 87, N 43° 25’ 39,8’’, E 2° 28’ 12,9’’, 1 177 m, 19/07/2023, 90 %, 2 m², 11 taxons
- Nardo strictae-Juncion squarrosi et unités supérieures : Juncus squarrosus 3, Carex binervis 3, Potentilla erecta 2, Luzula multiflora *m. 2, Nardus stricta 1, Veronica officinalis +,
- autres taxons : Molinia caerulea 3, Calluna vulgaris 2, Agrostis capillaris 2, Festuca arvernensis 1, Serratula tinctoria +.
La tonalité faiblement atlantique due à l’altitude conduit à rattacher cette végétation au Carici binervis-Nardetum strictae (Pethybridge & Praeger 1905) Braun-Blanq. & Tüxen 1952, décrit d’Irlande et cité du Tarn par Julve & de Foucault (1994 : 32, sub Nardo strictae-Caricetum binervis) plutôt qu’au Trocdario verticillati-Juncetum squarrosi (de Foucault, 2012).
Affine des bas-marais acidiphiles, la végétation amphibie de bas niveau topographique relevant des Littorelletea uniflorae Braun-Blanq. & Tüxen ex V. Westh. et al. 1946 est représentée en Montagne Noire au moins par l’Hyperico elodis-Potametum polygonifolii Braun-Blanq. & Tüxen 1952 des flaques d’eau acide oligotrophe et l’Eleocharitetum multicaulis (P. Allorge 1922) Tüxen 1937 des bordures amphibies de pièces d’eau (photo 43).
12. Les systèmes pastoraux de Peyremale (commune de Montolieu)
Le piémont méridional de la Montagne Noire offre encore un patrimoine végétal original comme en témoigne la monographie phytosociologique du domaine de Peyremale, sur la commune de Montolieu (de Foucault & Klesczewski, 2021), avec une extension vers Villardonnel, Verdun-en-Lauragais, Saissac, qui a donné lieu à la description des Dioscoreo communis-Pyretum spinosae, Anacamptido laxiflorae-Serapiadetum linguae, Ranunculo paludosi-Serapiadetum linguae, Junco acutiflori-Bellevalietum romanae, Ranunculo paludosi-Isoetetum duriei, et plus récemment (de Foucault, 2023d) du Serapiado linguae-Scorzoneretum humilis. Le relevé suivant offre un nouvel exemple du Ranunculo paludosi-Serapiadetum linguae :
Villardonnel (11), ‘Les Combals’, N 43° 20’ 05’’, E 2° 17’ 58,5’’, 357 m, 26/05/2020, 70 %, 30 m², 10 taxons
- Ranunculo paludosi-Serapiadetum linguae et unités supérieures : Ranunculus paludosus 2, Gaudinia fragilis 2, Serapias lingua 1, Lychnis flos-cuculi *f. +,
- autres taxons : Fraxinus angustifolia *a. j 1, Holcus lanatus *l. 1, Centaurium erythraea *e. 1, Hypericum perforatum +, Juncus conglomeratus +, Trifolium dubium +,
en mosaïque avec une pelouse thérophytique à Vulpia bromoides, Aira armoricana, Briza minor, Lathyrus nissolia, L. setifolius… accompagnés de taxons hygrophiles traduisant un engorgement temporaire du substrat (Juncus bufonius, Ranunculus sardous, Lysimachia minima, Lythrum hyssopifolia, Hypericum humifusum).
13. Conclusion
Durant une bonne dizaine d’années, on a donc poursuivi la connaissance phytosociologique du versant audois de la Montagne Noire. Treize associations végétales nouvelles ont été définies ici, dans différentes formations végétales. Il faut toutefois bien reconnaître que certaines sont sous-représentées (boisements, pelouses de dalles calcaires…). Il reste à souhaiter que cette contribution suscite de nouvelles investigations en Montagne Noire audoise, mais aussi tarnaise, et de prolonger ces études vers la Montagne Noire orientale, héraultaise.
D’un autre côté, il faudrait poursuivre la définition de certaines associations insuffisamment documentées, l’Eupatorio cannabini-Ranunculetum aconitifolii, le Centaureo pectinatae-Ericetum cinereae, le Tolpido umbellatae-Micropyretum tenelli, le Festuco arvernensis-Meetum athamantici, le Festuco arvernensis-Sedetum anglici et le Trinio glaucae-Arenarietum aggregatae.
Bibliographie
Andrieu F., Barreau D., Bossier X. & Vizcaïno D., 2021. Compte rendu de l’herborisation de la SBOcc au Roc de l’Aigle et à la Serre de Montredon (Montagne Noire, Aude). Carnets botaniques 78 : 1-22.
Barreau D., 2019. Catalogue de la flore de l’Aude, Société d’études scientifiques de l’Aude, Carcassonne.
Barreau D., Plassart C. & Vizcaïno D., 2016. Régions biogéographiques. In C. Plassart, D. Barreau & F. Andrieu, Atlas de la flore patrimoniale de l’Aude, Biotope, Mèze : 63-81.
Billy F., 2002. Végétations pionnières en basse-Auvergne. Bulletin de la Société botanique du Centre-Ouest, n. s., n° sp. 22 : 1-197.
Braun-Blanquet J. & Susplugas J., 1937. Reconnaissance phytogéographique dans les Corbières. Bulletin de la Société botanique de France 84 : 669-685.
Corriol G., Kessler F. & Menand M., 2021. Étude phytosociologique de quelques végétations vasculaires lithophiles, silicicoles du sud-ouest de la France. Carnets botaniques 48 : 1-10 + 6 tableaux, https://doi.org/10.34971/60JH-BB76.
Corriol G., Prud’homme F., Laigneau F. & Kessler F., 2016, publié 2022. Validation de quelques associations végétales inédites des Pyrénées et de Midi-Pyrénées. Le Monde des plantes 519 : 3-19.
Fallour D., Guitton H., Henry É., Thomassin G. & de Foucault B., 2013. Minisession phytosociologique synusiale en Montagne noire (Tarn et Aude), 18-20 mai 2013. Bulletin de la Société botanique du Centre-Ouest, n. s., 44 : 905-940.
Felzines J.-C. & Loiseau J.-E., 2004. Groupements thérophytiques printaniers acidiphiles médio-ligériens ; contribution à la structure de l’alliance Thero-Airion et de l’ordre des Helianthemetalia guttati (classe des Tuberarietea). Bulletin de la Société botanique du Centre-Ouest, n. s., 35 : 3-54.
Foucault B. (de), 1979. Observations sur la végétation des rochers arides de la Basse-Normandie armoricaine. Documents phytosociologiques, n. s., IV : 267-277.
Foucault B. (de), 1981. Cartographie chorologique et étude complémentaire de quelques associations végétales des pointements de roches précambriennes et primaires de Basse-Normandie continentale. Bulletin de la Société linnéenne de Normandie 108 : 61-70.
Foucault B. (de), 1986a. Contribution à une étude systémique des prairies de l’Aubrac (Massif central français). Documents phytosociologiques, n. s., X (1) : 255-305.
Foucault B. (de), 1986b. Données phytosociologiques sur la végétation observée lors de la douzième session de la S.B.C.O. en Limousin et Marche. Bulletin de la Société botanique du Centre-Ouest, n. s., 17 : 291-308.
Foucault B. (de), 1988. Contribution à la connaissance phytosociologique des corniches rocheuses de la vallée de l’Argenton, entre Argenton-Château et Massais (Deux-Sèvres). Bulletin de la Société botanique du Centre-Ouest, n. s., 19 : 39-64.
Foucault B. (de), 1991. Introduction à une systémique des végétations arbustives. Documents phytosociologiques, n. s., XIII : 63-104.
Foucault B. (de), 1994. Essai synsystématique sur les pelouses sèches acidophiles (Nardetea strictae, Caricetea curvulae). Colloques phytosociologiques XXII, Syntaxonomie typologique des habitats : 431-454.
Foucault B. (de), 2011a. Synthèse phytosociologique sur la végétation observée lors de la 146e session de la SBF dans les Ardennes. Bulletin de la Société d’histoire naturelle des Ardennes 101 : 33-50.
Foucault B. (de), 2011b. Contribution au prodrome des végétations de France : les Filipendulo ulmariae-Convolvuletea sepium Géhu & Géhu-Franck 1987. Journal de botanique de la Société botanique de France 53 : 73-137.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Nardetea strictae Rivas Goday in Rivas Goday & Rivas-Mart. 1963. Journal de botanique de la Société botanique de France 59 : 241-344.
Foucault B. (de), 2013. Contribution au prodrome des végétations de France : les Isoëtetea velatae de Foucault 1988 et les Juncetea bufonii de Foucault 1988 (« Isoëto–Nanojuncetea bufonii »). Journal de botanique de la Société botanique de France 62 : 35-70, 63 : 63-109.
Foucault B. (de), 2015. Quelques associations arbustives nouvelles pour la Malepère (Aude centrale). Bulletin de la Société botanique du Centre-Ouest, n. s., 46 : 244-260.
Foucault B. (de), 2017a. Données phytosociologiques nouvelles sur la végétation des Cévennes occidentales (département de la Lozère) et du Haut-Languedoc (département de l’Hérault). Bulletin de la Société botanique du Centre-Ouest, n. s., 48 : 334-359.
Foucault B. (de), 2017b. Quelques ourlets thérophytiques de l’Aude. Botanique 2 : 23-36.
Foucault B. (de), 2018a. La sortie phytosociologique du 28 mai 2017 à la Fontaine salée et ses environs en forêt domaniale de l’Eau salée (commune de Sougraigne). Bulletin de la Société d’études scientifiques de l’Aude CXVII : 189-196.
Foucault B. (de), 2018b. Caractérisation des habitats de la Bézalado et de la Sagne Grande, commune des Martys (Aude, France). Evaxiana 4 : 143-171.
Foucault B. (de), 2019. Contribution à la connaissance phytosociologique de la végétation du pays de Sault (Aude, France). Evaxiana 6 : 29-129.
Foucault B. (de), 2020a. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2020b. Données phytosociologiques sur les forêts de la Malepère (Aude, France) et leurs lisières. Carnets botaniques 9 : 1-15 et 15 tableaux, https://doi.org/10.34971/5nt2-zy92.
Foucault B. (de), 2020c. Essai de synthèse phytogéographique de l’Aude sur la base du Catalogue 2019 de sa flore. Carnets bot. 23 : 1-10 et annexes, https://doi.org/10.34971/ZBV5-3252.
Foucault B. (de), 2021a. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de), 2021b. Miscellanées phytosociologiques : quelques syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 44 : 1-23 + 28 tableaux, https://doi.org/10.34971/AKGG-T733.
Foucault B. (de), 2021c. Quelques données phytosociologiques sur les ripisylves languedociennes et certains de leurs satellites. Carnets botaniques 42 : 1-14 + 10 tableaux, https://doi.org/10.34971/V2QK-XN03.
Foucault B. (de), 2021d. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2022a. Validation de syntaxons invalidement publiés en 2021. Carnets botaniques 79 : 1-12, https://doi.org/10.34971/dxjk-t193.
Foucault B. (de), 2022b. Miscellanées phytosociologiques III : syntaxons nouveaux, validés ou complétés. Carnets botaniques 112 : 1-22, https://doi.org/10.34971/6AX1-0K40.
Foucault B. (de), 2023a. Miscellanées phytosociologiques IV : syntaxons nouveaux, validés ou complétés. Carnets botaniques 123 : 1-17, https://doi.org/10.34971/1d8e-ky97.
Foucault B. (de), 2023b. Un panorama formalisé des ethnosciences naturelles. Carnets botaniques 123 : 1-100, https://doi.org/10.34971/w804-7n97.
Foucault B. (de), 2023c. Miscellanées phytosociologiques V : syntaxons nouveaux, validés ou complétés. Carnets botaniques 154 : 1-15 + 6 tableaux, https://doi.org/10.34971/M5RZ-7B62.
Foucault B. (de), 2023d. Retour sur la structuration synsystématique des alliances du Molinion caeruleae et du Juncion acutiflori. Carnets botaniques 146 : 1-26 + tableaux 1 à 18d, https://doi.org/10.34971/86S0-XR84.
Foucault B. (de) & Royer J.-M., 2014. Contribution au prodrome des végétations de France : les Franguletea alni Doing ex V. Westh. in V. Westh. & Den Held 1969. Journal de botanique de la Société botanique de France 66 : 83-106.
Foucault B. (de) & Catteau E., 2015. Contribution au prodrome des végétations de France : les Epilobietea angustifolii Tüxen & Preising in Tüxen 1950. Journal de botanique de la Société botanique de France 70 : 63-88.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Foucault B. (de) & Klesczewski M., 2021. Contribution à une monographie phytosociologique du domaine de Peyremale (commune de Montolieu, Aude). Carnets botaniques 47 : 1-18 et tableaux 2 à 17, https://doi.org/10.34971/B79J-2H81.
Fuente V. (de la), 1985. Vegetación orófila del occidente de la provincial da Guadalajara (España). Lazaroa 8 : 123-219.
Géhu J.-M., 1975. Essai pour un système de classification phytosociologique des landes atlantiques françaises. Colloques phytosociologiques II, La végétation des landes d’Europe occidentale : 361-377.
Géhu J.-M. & Géhu J., 1975. Les fourrés à Erica scoparia et Frangula alnus d’Aquitaine (Scopario-Franguletum alnae). Documents phytosociologiques 9-14 : 117-120.
Ghestem A. & Descubes-Gouilly Ch., 1977. Phytosociologie de Digitalis purpurea L. en Limousin (France) (premiers documents). Documents phytosociologiques, n. s., I : 125-133.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Glemarec E., Delassus L. & Boullet V., à paraître. Contribution au prodrome des végétations de France : les Calluno vulgaris – Ulicetea minoris Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, 3e série.
Guitton H., Henry É., Puig S. & de Foucault B., 2016. Sur quelques communautés végétales originales du Sidobre (Tarn, France) ; compte rendu de la minisession phytosociologie synusiale du 23 au 25 mai 2015. Bulletin de la Société botanique du Centre-Ouest, n. s., 47 : 317-328.
Guitton H., Henry É., Puig S. & de Foucault B., 2017a. Sur quelques communautés végétales originales des monts de Lacaune (Tarn, France) : compte rendu de la minisession de phytosociologie synusiale (10 au 12 juin 2016). Bulletin de la Société botanique du Centre-Ouest, n. s., 48 : 308-329.
Guitton H., Juhel C. & Julve Ph., 2017b. Une nouvelle association de lande thermoatlantique à Cistus salviifolius et Erica cinerea en France. Documents phytosociologiques, 3e série, 6 : 202-211.
Julve Ph. & de Foucault B., 1994. Phytosociologie synusiale dans le Tarn. Bulletin de la Société de botanique du nord de la France 47 (4) : 23-47.
Korneck D., 1975. Das Narduretum lachenalii, eine neue Thero-Airion-Assoziation. Beiträge zur naturkundlichen Forschung in Südwestdeutschland 34 : 161-166.
López G., 1978. Contribución al conocimiento fitosociologico de la Serrania de Cuenca, II. Anales del Instituto botánico A.J. Cavanilles 34 (2) : 597-702.
Mateo Sanz G., 1983. Estudio sobre la flora y vegetación de las sierras de Mira y Talayuelas. I.C.O.N.A. Monografias 31 : 1-290.
Plassart C., Barreau D. & Andrieu F., 2016. Atlas de la flore patrimoniale de l’Aude. Biotope, Mèze, 432 p.
Rivas Goday S;, 1958. Nuevos ordenes y allianzas de Helianthemetea annua Br.-Bl. Anales del Instituto botánico A.J. Cavanilles 15 : 539-651.
Royer J.-M. & Ferrez Y., 2018. Contribution au prodrome des végétations de France : les Sedo albi – Scleranthetea biennis Braun-Blanq. 1955. Documents phytosociologiques, 3e série, 7 : 180-281.
Royer J.-M. & Ferrez Y., 2019. Contribution au prodrome des végétations de France : les Festuco – Brometea Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, 3e série, 13 : 4-382.
Thébaud G., Roux C., Bernard C.-E. & Delcoigne A., 2014. Guide d’identification des végétations du nord du Massif central, associatins végétales et habitats naturels. Presses universitaires Blaise-Pascal, Clermont-Ferrand, 274 p.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.
Wilmanns O., Bogenrieder A. & Nakamura Y., 1985. Vergleichende Studien des Pinus-Krummholzes in den japanischen und europäischen Alpen. Tuexenia 5 : 335-358.


