
Les pelouses des sables calcaires des Landes de Gascogne (Lot-et-Garonne, Landes et Gironde)
Title
The limestone sand grasslands of the Landes de Gascogne (Lot-et-Garonne, Landes and Gironde)
Résumé
L’est des Landes de Gascogne a la particularité de posséder des sables calcaires permettant le développement d’une flore et de végétations originales pour la région Nouvelle-Aquitaine. Nous décrivons ainsi les pelouses de l’Helianthemo nummularii-Festucetum vasconcensis, du Corynephoro canescentis-Festucetum vasconcensis et de l’Erodio bipinnati-Alyssetum alyssoidis que nous replaçons dans le synsystème.
Abstract
The eastern part of the Landes de Gascogne has the particularity of possessing limestone sands allowing the development of original flora and vegetation for the Nouvelle-Aquitaine region. We thus describe the lawns of Helianthemo nummularii-Festucetum vasconcensis, Corynephoro canescentis-Festucetum vasconcensis and Erodio bipinnati-Alyssetum alyssoidis, which we place in the synsytem.
Les Landes de Gascogne constituent un immense plateau sédimentaire de 1,4 millions d’hectares composé de sables des Landes. Ces sables d’origine éolienne ont la particularité d’être très acides, de 3,5 à 5,5 unités pH (Jolivet et al., 2007 ; Trichet et al., 1999), d’être sans argile ou limon et pauvres en nutriments et notamment en phosphore. Ces propriétés résultent en partie de l’absence de stade boisé durant des siècles (en dehors de certaines marges des réseaux hydrographiques).
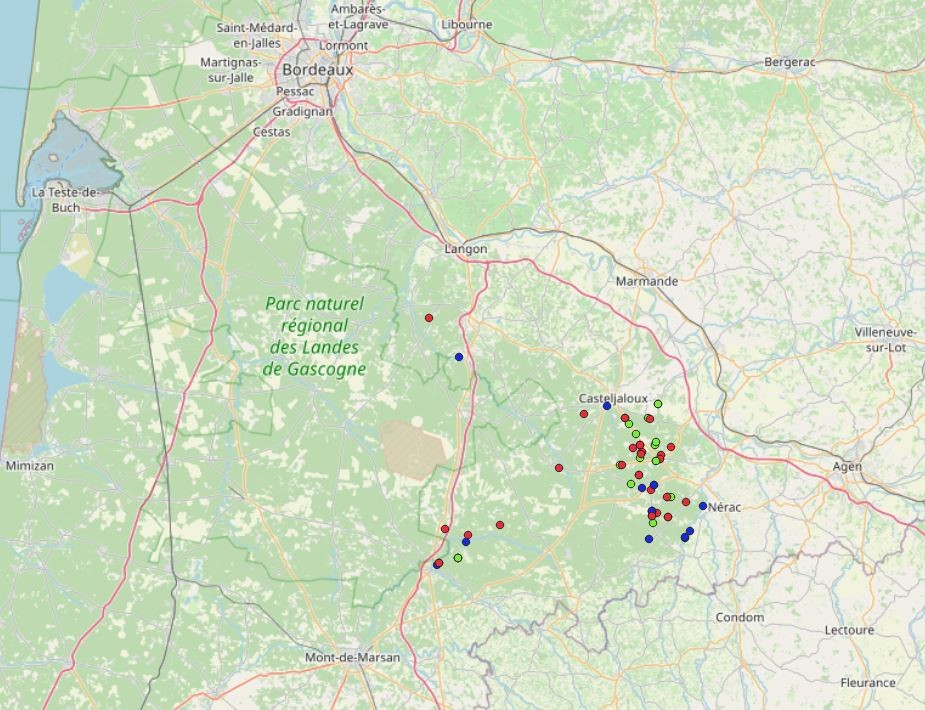
Alors que l’épaisseur de sable peut atteindre plusieurs dizaines de mètres le long du littoral aquitain, elle s’amenuise progressivement jusqu’à la pointe est du triangle landais. Dans cette zone globalement comprise entre Bazas (Gironde), Nérac et Sos (Lot-et-Garonne) (carte 2), un lent processus de brassage entre les sables éoliens et l’ensemble des molasses et calcaires sous-jacents devient alors possible, probablement grâce à des phénomènes de bioturbation et/ou de transferts hydriques (P. Becheler, comm. orale). Cette singularité permet le développement d’une flore très originale associant le cortège classique d’espèces psammophiles plus ou moins acidiphiles à un contingent d’espèces basiphiles absentes du reste des Landes de Gascogne (Euphorbia seguieriana, Artemisia campestris subsp. campestris) à l’exception de secteurs localisées sur les marges nord, de la région de Roquefort (Landes) et de certaines dunes côtières (Armeria arenaria, Euphorbia seguieriana, Koeleria arenaria, Festuca vasconcensis, Silene conica, S. otites, etc. ; carte 1).

Les milieux ouverts des sols secs et acides des Landes de Gascogne ont fait l’objet de plusieurs publications (Géhu, 1974 ; de Foucault, 1984, 1986 ; Lafon, 2019), même si leur connaissance reste encore à approfondir. En revanche, les végétations de l’est des Landes de Gascogne, plus neutrophiles, n’ont pas à notre connaissance fait l’objet d’études. Alors que ces particularités floristiques et écologiques étaient connues de longue date (notamment Lapeyrère, 1892 ; Debeaux, 1896), la mise en évidence de l’originalité de ses végétations semble récente et très partielle (Lafon et al., 2014, 2018). Cet article tente ainsi de compléter cette connaissance locale.
La taxonomie suit le référentiel taxonomique national pour la faune, la flore et la fonge de France métropolitaine et d’outre-mer (Gargominy et al., 2019), élaboré et diffusé par le Muséum national d’histoire naturelle dans le cadre de la mise en œuvre du Système d’information sur la nature et les paysages. Seules les sous-espèces différentes de celles de l’espèce ont été précisées.
Plusieurs espèces de nos relevés appartiennent à des groupes taxonomiques complexes et leur détermination n’est pas toujours garantie. Ainsi, Thymus drucei a pu être confondu avec Th. longicaulis auct. sensu Flora Gallica (Tison & de Foucault, 2014). Festuca lemanii pose encore de nos jours de nombreux problèmes, à la fois sur sa description biométrique, sa chorologie et sa niche écologique, il est possible que certains individus notés ici sous ce nom correspondent à F. auquierii voire à F. ovina subsp. guestfalica. De même, Festuca nigrescens a pu être confondu avec un infrataxon du groupe de F. trichophylla.
Le système phytosociologique suit le catalogue des végétations de Nouvelle-Aquitaine (Lafon et al., 2020) qui intègre les travaux récents de phytosociologie sigmatiste français et européens et pour lesquels nous proposons des modifications. Nous avons également actualisé les autorités synomenclaturales et les noms de syntaxons en suivant le Code international de nomenclature phytosociologique (Theurillat et al., 2020).
Les végétations bryolichéniques n’ont pas été relevées avec la végétation trachéophytique en accord avec les préconisations d’auteurs multiples (Boullet, 1986 ; Deil, 1995 ; Bardat et al., 2012 ; de Foucault, 2015, 2018) ayant relevé que ces deux grands ensembles ne partagent pas le même niveau d’homogénéité. Toutefois, dans certains cas et lorsque ces espèces étaient connues, elles ont été intégrées pour information.
Les tableaux phytosociologiques sont présentés dans le texte pour faciliter leur lecture mais ne comportent pas toutes les espèces accidentelles. Pour cela, les tableaux complets sont disponibles en fichiers annexes au format Excel.
1. Résultats phytosociologiques
1.1. La pelouse à Hélianthème nummulaire et Fétuque de Gascogne : Helianthemo nummularii-Festucetum vasconcensis ass. nov. hoc loco
Typus nominis hoc loco : relevé 403930 du tableau 1 hoc loco
Cortège caractéristique et position syntaxonomique
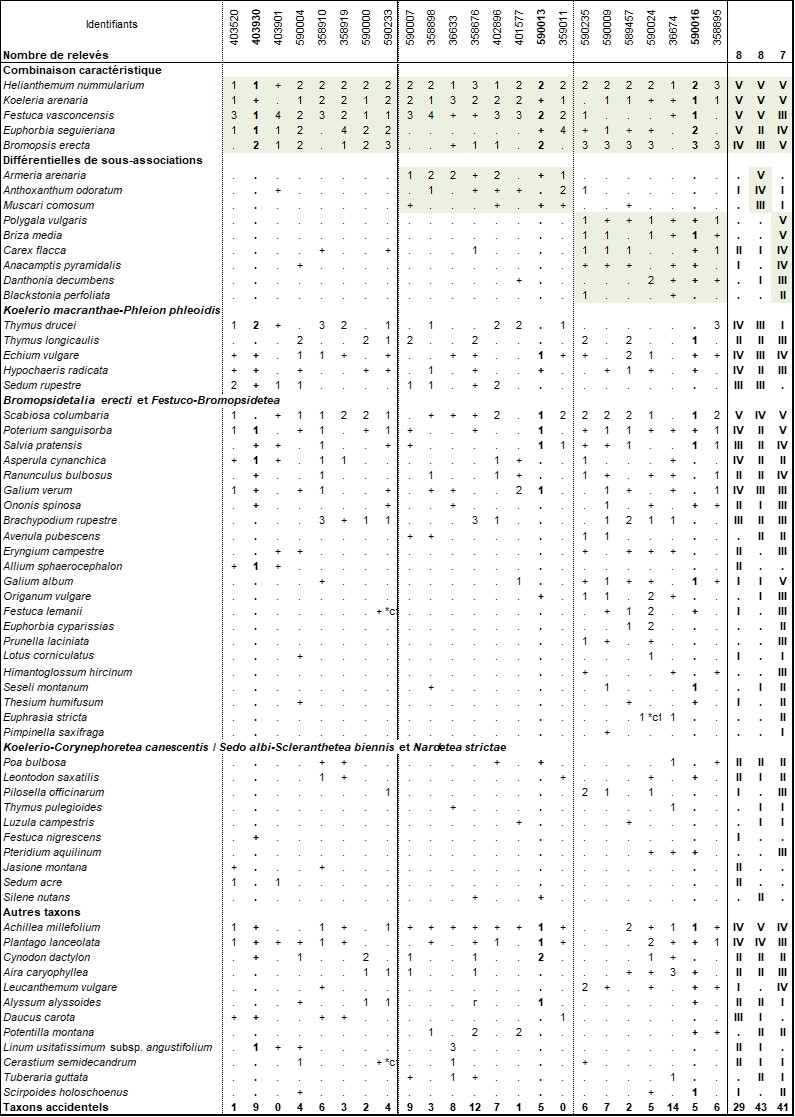
Cette pelouse plus ou moins fermée (photos 1 et 2) se caractérise par Helianthemum nummularium, Festuca vasconcensis, Koeleria arenaria, Euphorbia seguieriana et Bromopsis erecta (rare dans le secteur). La présence de Poterium sanguisorba, Scabiosa columbaria, Salvia pratensis et l’important contingent d’espèces des Bromopsidetalia erectae nous permettent de ranger cette végétation dans les pelouses calcicoles des Festuco-Bromopsidetea. La présence constante d’espèces des pelouses sableuses comme Koeleria arenaria, Sedum rupestre, Festuca vasconcensis et Thymus drucei (parfois remplacé par Th. longicaulis auct., mais dont la distinction n’est pas toujours aisée) permet un rattachement au Koelerio macranthae-Phleion phleoidis. Nous intégrons par défaut notre association au Festucenion longifolio-lemanii par la présence de Thymus drucei et de Festuca vasconcensis. Cette fétuque cespiteuse de la section Festuca (groupe ovina) peut être considérée comme le vicariant structurel de F. longifolia ou de F. arvernensis dominants dans les végétations du Festucenion longifolio-lemanii du Massif central et de la vallée de la Loire (Royer & Ferrez, 2020).


Variations
En plus du typicum, deux sous-associations ont pu être mises en évidence. La première, que l’on propose de nommer armerietosum arenariae subass. nov. hoc loco (typus nominis hoc loco : relevé 590013 du tableau 1 hoc loco), se différencie par un ensemble plus important de taxons acidiphiles (Armeria arenaria, Anthoxanthum odoratum, etc.) et fait la transition vers le Corynephoro canescentis-Festucetum vasconcensis présenté ci-après. La seconde, dénommée brizetosum mediae subass. nov. hoc loco (typus nominis hoc loco : relevé 590016 du tableau 1 hoc loco), est plus riche en espèces des Festuco-Bromopsidetea (Briza media, Carex flacca, Blackstonia perfoliata, Anacamptis pyramidalis, etc).
Écologie
Cette pelouse se développe sur des sables stabilisés encore riches en calcaire. Il est également probable que ces sables soient bien pourvus en nutriments, la biomasse y étant plus importante que dans le Corynephoro canescentis-Festucetum vasconcensis.
Ces végétations se développent particulièrement bien sur les talus routiers. Il est probable qu’elle ait été aussi présente de façon surfacique au sein de certains airiaux (unités familiales d’exploitation propres au plateau landais, formant des clairières au sein de la pinède) ou plus largement des parcours pastoraux ancestraux, mais la disparition progressive de ces modes d’exploitation traditionnels couplée à la sylviculture intensive du Pin maritime en a fait disparaître les pelouses qui lui étaient associées.
La végétation pionnière qui précède l’Helianthemo nummularii-Festucetum vasconcensis dans la succession végétale n’a pas été formellement identifiée, car elle est particulièrement dynamique. Toutefois, le relevé 589999 du tableau 3 pourrait en être un représentant, mais cela reste à étudier. L’Erodio bipinnati-Alyssetum alyssoidis pourrait également participer à la succession sur les sables plus acides.
1.2. La pelouse à Corynéphore blanchâtre et à Fétuque de Gascogne : Corynephoro canescentis-Festucetum vasconcensis ass. nov. hoc loco
Typus nominis hoc loco : relevé 590210 du tableau 2 hoc loco
Cortège caractéristique et position syntaxonomique
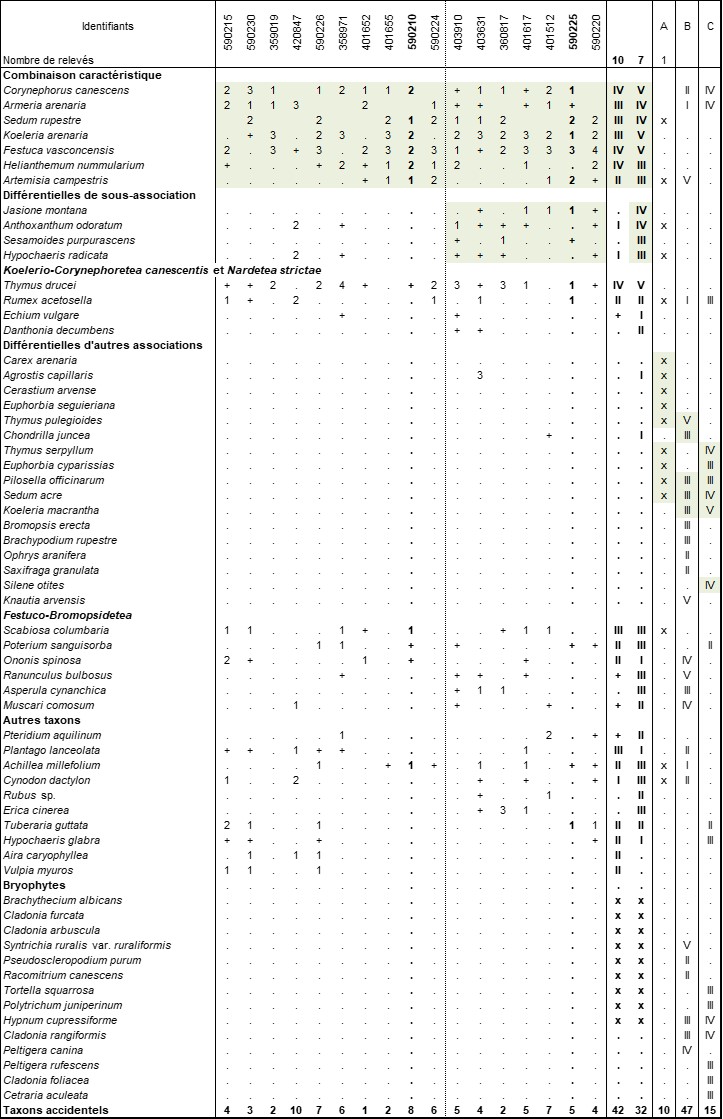
Cette pelouse le plus souvent ouverte (photo 3) se caractérise par la présence d’Armeria arenaria, Corynephorus canescens, Festuca vasconcensis, Helianthemum nummularium, Koeleria arenaria, Sedum rupestre et plus ponctuellement de Rumex acetosella et Artemisia campestris subsp. campestris, rare dans la région.

La présence de Rumex acetosella, Corynephorus canescens, Sedum rupestre, Thymus drucei, Artemisia campestris permet un rattachement aux Koelerio-Corynephoretea canescentis. Il nous semble qu’un rattachement au Sileno conicae-Cerastion semidecandri, qui rassemble les pelouses sablo-calcaires annuelles (Korneck, 1974) et dont la discussion est entreprise dans l’association suivante est à écarter. Sissingh et Tideman (1960) ont défini aux Pays-Bas le Sedo-Cerastion arvensis, rassemblant les pelouses vivaces sur sables calcaires du domaine atlantique à subatlantique. Mucina et al. (2016) considèrent le Sedo-Cerastion arvensis comme une alliance des pelouses fermées mésoxérophiles des sols sableux acides à neutres des plaines (sub)atlantiques d’Europe (vicariant de l’Armerion elongatae Pötsch 1962 continental). L’association type du Sedo-Cerastion arvensis est l’Euphorbietum seguieriano-cyparissiae Sissingh & Tideman 1960 (basé sur un relevé unique) et la définition qu’en donnent Sissingh et Tideman (1960) semble davantage plaider pour une restriction à la partie basiphile à neutroclinophile des pelouses sur sables ; les végétations plus acidiphiles peuvent en outre prendre place dans le Festucion guestfalico-filiformis Loiseau & Felzines in J.-M. Royer, Felzines, Misset & Thévenin 2006, d’ailleurs mis en synonyme du Sedo-Cerastion arvensis par Mucina et al. (2016).
La proximité du Corynephoro canescentis-Festucetum vasconcensis avec l’Euphorbietum seguieriano-cyparissiae est d’ailleurs évidente (colonne A du tableau 2) avec la présence commune de Rumex acetosella, Artemisia campestris subsp. campestris, Thymus gr. pulegioides (= Th. drucei ?), Sedum rupestre, Scabiosa columbaria, Hypochaeris radicata, Anthoxanthum odoratum, etc. Pour ces raisons, nous proposons donc de rattacher cette association au Sedo-Cerastion arvensis.
Afin de confirmer l’originalité de notre association, nous l’avons comparée avec l’Artemisietum campestris G. Lemée 1937 (colonne B du tableau 2) et le Sileno conicae-Koelerietum macranthae Paul & Y. Rich. ex Loiseau & Felzines 2010 (colonne C du tableau 2). Le Corynephoro canescentis-Festucetum vasconcensis y apparaît nettement différencié par la présence de Sedum rupestre, Koeleria arenaria, Festuca vasconcensis, Helianthemum nummularium et l’absence de Chondrilla juncea, Euphorbia cyparissias, Pilosella officinarum, Sedum acre, Koeleria macrantha, etc.
Enfin, la synusie bryolichénique n’a pas été relevée systématiquement mais quelques taxons semblent être assez fréquents : Polytrichum juniperinum, Hypnum cupressiforme, Cladonia furcata, Racomitrium canescens et plus ponctuellement Brachythecium albicans, Peltigera rufescens, Cladonia arbuscula, Syntrichia ruralis var. ruraliformis et Pseudoscleropodium purum.
Variations
En plus du typicum, un jasionetosum montanae subass. nov. hoc loco (typus nominis hoc loco : relevé 590225 du tableau 2 hoc loco ; photo 4) des sols plus acides est mis en évidence. Il se différencie par la présence de Jasione montana, Sesamoides purpurascens et Anthoxanthum odoratum. Il fait la transition vers l’Astrocarpo purpurascentis-Corynephoretum canescentis Braun-Blanq. 1967, dont Géhu (1974) a démontré le premier la présence dans les Landes de Gascogne.

Écologie
Cette pelouse se développe sur des sables peu mobiles et à richesse modérée en carbonates. Ces sables semblent plus acides que pour l’association précédente, ce dont témoignent l’absence de la plupart des espèces des Festuco-Bromopsidetea, la présence de Rumex acetosella et, dans une moindre mesure, de Corynephorus canescens. En effet, Paul et Richard (1968) ont démontré la tolérance de cette dernière pour les sables calcaires dans certaines conditions. La fréquence d’Armeria arenaria témoigne également du fait que ces sables ne sont plus très riches en calcaires (Paul, 1969). Cette pelouse peut ainsi être qualifiée d’acidiclinophile. La biomasse est aussi plus faible que pour l’Helianthemo nummularii-Festucetum vasconcensis, témoignant d’une plus faible richesse en nutriments dans le sol et/ou de sables plus filtrants en raison d’une importance moindre en éléments limoneux. Seule une étude pédologique permettrait de valider ces hypothèses.
1.3. La pelouse à Bec-de-grue poilu et Passerage à calices persistants : Erodio bipinnati-Alyssetum alyssoidis ass. nov. hoc loco
Typus nominis hoc loco : relevé 590011 du tableau 3 hoc loco
Cortège caractéristique et position syntaxonomique
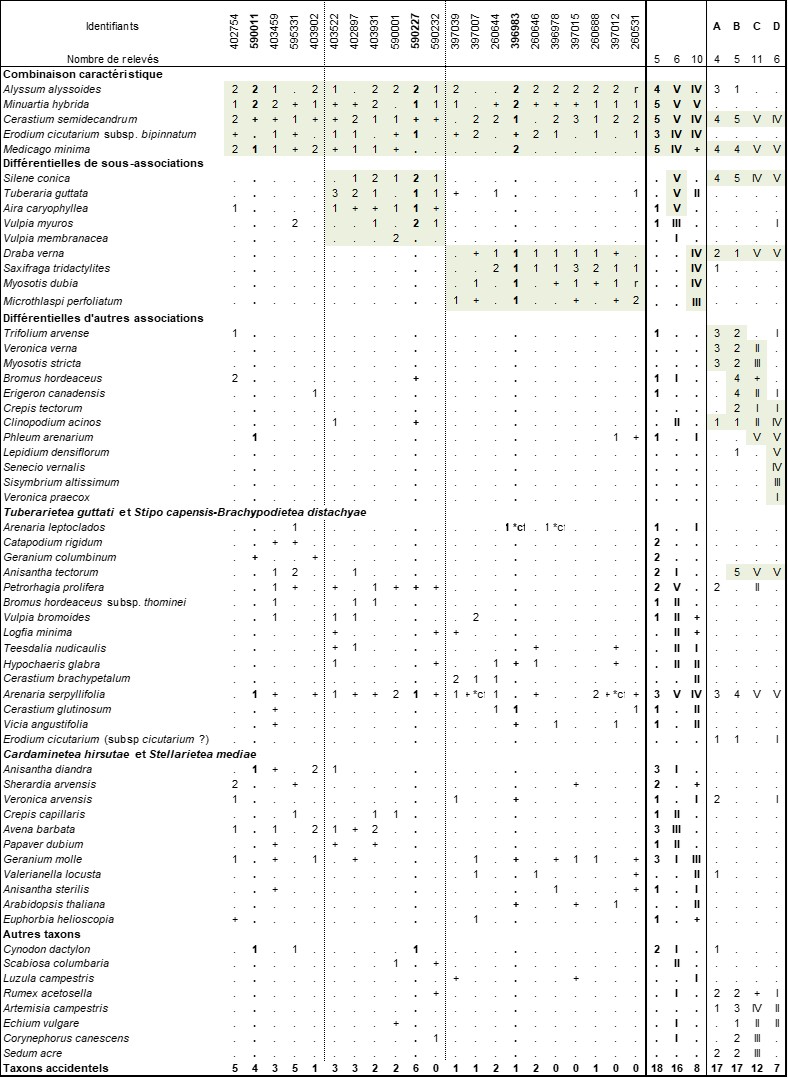
Cette pelouse thérophytique très ouverte (photo 5) se caractérise par la présence de taxons calcicoles comme Alyssum alyssoides, de taxons sabulicoles comme Cerastium semidecandrum, Erodium cicutarium subsp. bipinnatum et des taxons thérophytes fréquents des Landes de Gascogne comme Minuartia hybrida, Medicago minima ou Arenaria serpyllifolia.

Cette association semble bien s’intégrer dans la conception du Sileno conicae-Cerastion semidecandri qu’en donne son auteur (Korneck, 1974). En effet, Korneck la définit comme rassemblant des « communautés pionnières annuelles subméditerranéennes et subatlantiques caractérisées par Silene conica subsp. conica, Cerastium semidecandrum, Medicago minima et dans une moindre mesure Phleum arenarium et Bromus tectorum ». Il précise également que ces végétations sont parfois en contact de végétations vivaces à Koeleria glauca. Il décrit deux associations thérophytiques au sein de cette alliance : le Bromo tectorum-Phleetum arenarii (colonnes C et D du tableau 3) et le Sileno conicae-Cerastietum semidecandri (colonnes A et B du tableau 3). Dengler et al. (2003) définissent cette dernière association comme le lectotype du Sileno conicae-Cerastion semidecandri.
Le Sileno conicae-Cerastietum semidecandri est très proche de l’Erodio bipinnati-Alyssetum alyssoidis (colonnes A et B du tableau 3) dont il se différencie principalement par la présence de Veronica verna, Trifolium arvense, Myosotis stricta et l’absence de Minuartia hybrida, Erodium cicutarium subsp. bipinnatum. Notre association peut ainsi être considérée comme le vicariant thermo-atlantique, voire atlantique, du Sileno conicae-Cerastietum semidecandri subatlantique à continental. La synusie thérophytique de l’Artemisietum campestris G. Lemée 1937 semble très proche de notre association, ce qui pourrait laisser penser à sa présence dans le Perche.
Le rattachement du Sileno conicae-Cerastion semidecandri à une unité supérieure apparaît plus complexe. Bardat et al. (2004) l’intègrent aux Corynephoretalia canescentis et aux Koelerio-Corynephoretea canescentis alors que Korneck (1974) ou Mucina et al. (2016) le rattachent aux Sedo albi-Scleranthetalia biennis et aux Sedo albi-Scleranthetea biennis. Mais l’intégration d’une alliance thérophytique à des unités à dominante d’espèces vivaces sur la base de la présence de taxons hémicryptophytiques issus de leur succession dynamique est largement discutable, les espèces thérophytiques communes étant le plus souvent assez faibles.
Un rattachement aux classes de végétations thérophytiques des Tuberarietea guttatae (Braun-Blanq. ex Rivas Goday 1958) Rivas Goday & Rivas Mart. 1963 nom. mut. ou des Stipo capensis-Brachypodietea distachyi Brullo in Brullo, Scelsi & Spamp. 2001 nom. mut. ne nous paraît pas évident non plus dans leurs conceptions actuelles. En effet, les thérophytes acidiphiles qui caractérisent les Tuberarietea guttatae sont trop rares dans les relevés des associations du Sileno conicae-Cerastion semidecandri rassemblées (tableau 3). Les Stipo capensis-Brachypodietea distachyi qui représentent l’équivalent des Tuberarietea guttatae pour les sols calcaires à neutres ont une définition méditerranéenne où le Sileno conicae-Cerastion semidecandri s’intègrerait assez mal. En 1999, de Foucault proposait une conception des Stipo capensis-Brachypodietea distachyi plus large mais faiblement caractérisée floristiquement : Medicago minima, Arenaria serpyllifolia, Catapodium rigidum, Trifolium scabrum et T. campestre. Il y intégrait ainsi le Sileno conicae-Cerastion semidecandri dans le nouvel ordre des Phleo arenarii-Cerastietalia semidecandri (Glowacki 1988) B. Foucault 1999. Cette proposition tentante resterait à étudier sur la base de la connaissance actuelle, mais elle dépasse le cadre de cet article.
Variations
En plus du typicum, deux sous-associations peuvent être mises en évidence. La première est restreinte aux sables plus rudéralisés et se différencie par la présence de taxons nitrophiles des Cardaminetea hirsutae : Draba verna, Saxifraga tridactylites, Microthlaspi perfoliatum et Myosotis dubia et par la rareté de Medicago minima. Nous proposons de la nommer saxifragetosum tridactylitae subass. nov. hoc loco (typus nominis hoc loco : relevé 396983 du tableau 3 hoc loco).
La seconde sous-association, des sols plus pauvres en calcium et faisant transition vers les pelouses acidiphiles des Tuberarietea guttatae, se différencie par la présence de taxons de cette classe (Tuberaria guttata, Aira caryophyllea, Vulpia myuros), ainsi que de Silene conica ou Petrorhagia prolifera. Nous proposons de la nommer silenetosum conicae subass. nov. hoc loco (typus nominis hoc loco : relevé 590227 du tableau 3 hoc loco).
Écologie
L’Erodio bipinnati-Alyssetum alyssoidis est une tonsure vernale (optimum au mois d’avril) qui se développe sur des sables moyennement riches en calcaires et qui entre dans la dynamique du Corynephoro canescentis-Festucetum vasconcensis avec lequel elle partage la même écologie. Ces deux végétations se retrouvent régulièrement en contact spatial. Elle pourrait également rentrer dans la dynamique de l’Helianthemo nummularii-Festucetum vasconcensis (au moins pour le silenetosum conicae).
2. Synsystème des associations décrites
Festuco-Bromopsidetea Braun-Blanq. & Tüxen ex Klika & Hadač 1944 nom. mut. nov. hoc loco P. Lafon & E. Henry (= Festuco-Brometea Braun-Blanq. & Tüxen ex Klika & Hadač 1944)
Bromopsidetalia erecti W. Koch 1926 nom. mut. nov. hoc loco P. Lafon & E. Henry (= Brometalia erecti W. Koch 1926)
Koelerio macranthae-Phleion phleoidis Korneck 1974
Festucenion longifolio-lemanii Loiseau & Felzines 2010
Helianthemo nummularii-Festucetum vasconcensis
typicum
armerietosum arenariae
brizetosum mediae
Koelerio-Corynephoretea canescentis Klika in Klika & V. Novák 1941
Corynephoretalia canescentis Klika 1934
Sedo-Cerastion arvensis Sissingh & Tideman 1960
Corynephoro canescentis-Festucetum vasconcensis
typicum
jasionetosum montanae
Sileno conicae-Cerastion semidecandri Korneck 1974
Erodio bipinnati-Alyssetum alyssoidis
typicum
saxifragetosum tridactylitae
silenetosum conicae
3. Conclusion
Cette étude centrée sur la pointe est des Landes de Gascogne a révélé la présence de trois associations nouvelles :
- l’Helianthemo nummularii-Festucetum vasconcensis, pelouse vivace des sables stabilisés encore riches en calcaire ;
- le Corynephoro canescentis-Festucetum vasconcensis, pelouse vivace ouverte sur sables peu mobiles et légèrement enrichis en carbonates;
- l’Erodio bipinnati-Alyssetum alyssoidis, pelouse thérophytique vernale psammophile en contact spatial avec les deux premières végétations.
Leur position dans le synsystème a été discutée. Elle a permis de remettre en avant le Sedo-Cerastion arvensis, tout en précisant les contours de ses enveloppes écologique et chorologique. La chorologie du Festucenion longifolio-lemanii a également été étendue au plateau landais.
Ces trois associations sont actuellement considérées comme endémiques de l’est des Landes de Gascogne, avec parfois des irradiations sur les marges nord jusqu’en Gironde (carte 2). Leurs répartitions sont assez similaires et centrées entre Beauziac, Ambrus et Fargues-sur-Ourbise, ainsi qu’en quelques stations périphériques à l’est de Roquefort (Landes) et au sud-ouest de Langon (Gironde). Le Corynephoro canescentis-Festucetum vasconcensis est probablement l’association la plus rare, car l’Helianthemo nummularii-Festucetum vasconcensis, qui occupait sans doute au siècle dernier des biotopes moins rudéraux, trouve aujourd’hui refuge le long des linéaires routiers.
Toutes ces pelouses sur sables plus ou moins calcaires sont d’une exceptionnelle patrimonialité en raison de l’originalité écologique de leurs habitats. Elles contiennent de plus des espèces protégées ou exceptionnelles pour la région (Euphorbia seguieriana, Artemisia campestris subsp. campestris, etc.). Elles ont probablement historiquement régressé en raison de la fermeture du milieu et de la sylviculture de Pins maritime qui acidifie le milieu.
Les caractéristiques écologiques et dynamiques de ces associations restent encore à préciser, notamment par la réalisation d’analyses pédologiques des zones de relevé.
Les prospections de terrain ont permis de réaliser le contour des parties les plus xériques de ce secteur au sein duquel se trouve le principal noyau de présence des trois associations. Il a donné lieu à la création d’une ZNIEFF de type 2 Pinède à chêne liège de l’est du triangle landais (https://inpn.mnhn.fr/zone/znieff/720030121) qui concentre de nombreuses végétations originales comme le groupement à Cistus umbellatus et Cistus lasianthus subsp. alyssoides P. Lafon, Le Fouler & Caze 2015, ainsi que plusieurs espèces végétales très originales pour le secteur (parmi celles que nous n’avons pas encore citées pour la xérosère : Airopsis tenella, Neoschischkinia elegans, Ophrys incubacea, Galium boreale, Cytinus hypocistis, Agrostis castellana, Anemone rubra, Daphne cneorum, Polycnemum majus, etc.). De nouvelles découvertes sont encore à y faire, tant au niveau floristique que phytosociologique !
Bibliographie
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Collection Patrimoines naturels 61 : 1-171.
Bardat J., Boullet V. & Hugonnot V., 2012. La place des bryophytes dans la synsystématique des végétations vasculaires. 1 – La problématique. Communication inédite au colloque de Saint-Mandé 2012, Prodrome et cartographie des végétations de France.
Boullet V., 1986. Les pelouses calcicoles (Festuco-Brometea) du domaine atlantique français et ses abords au nord de la Gironde et du Lot. Essai de synthèse phytosociologique. Thèse, Université des Sciences et Techniques de Lille, 333 p. + annexes.
Deil U. 1995. The class Adiantetea in the Mediterranean area; an approach from vegetation history and community evolution. Colloques phytosociologiques XXIII : 241-258.
Debeaux O., 1896. Flore du Lot-et-Garonne ou catalogue méthodique des plantes vasculaires de ce département. 645 p.
Dengler J., Berg C., Eisenberg M., Isermann M., Jansen F., Koska I., Löbel S., Manthey M., Päzolt J., Spangenberg A., Timermann T. & Wollert H., 2003. New descriptions and typifications of syntaxa within the project ‘Plant communities of Mecklenburg-Vorpommern and their vulnerability’, I. Feddes Repertorium 114 (7-8) : 587-631.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse, Université de Rouen, Rouen, 675 p. et annexes.
Foucault B. (de), 1986. Données systémiques sur la végétation prairiale mésophile du Pays Basque et des Landes de Gascogne. Documents phytosociologiques NS 10 (1) : 203-219.
Foucault B. (de), 1999. Nouvelle contribution à une synsystématique des pelouses sèches à thérophytes. Documents phytosociologiques,NS, XIX : 47-105.
Foucault B. (de), 2015. Contribution au prodrome des végétations de France : les Adiantetea capilli-veneris Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952. Acta Botanica Gallica: Botany Letters 162 (4) : 375-403.
Foucault B. (de), 2018. Contribution au prodrome des végétations de France : les Montio fontanae – Cardaminetea amarae Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques,série 3, 7 : 6-86.
Gargominy O., Tercerie S., Régnier C., Ramage T., Dupont P., Daszkiewicz P. & Poncet L., 2019. TAXREF v13, référentiel taxonomique pour la France : méthodologie, mise en œuvre et diffusion. Muséum national d’histoire naturelle, Paris, Rapport Patrinat, 63 p.
Géhu J.-M., 1974. L’Astrocarpo-Corynephoretum canescentis Br.-Bl. 1967 dans les landes de Gascogne. Documents phytosociologiques 6 : 43-45.
Jolivet C., Augusto L., Trichet P. & Arrouays D., 2007. Les sols du massif forestier des Landes de Gascogne : formation, histoire, propriétés et variabilité spatiale. Revue forestière française LIX (1) : 7-30.
Korneck D., 1974. Xerothermvegetation in Rheinland-Pfalz und Nachbargebieten. Schriftenreihe für Vegetationskunde 7 : 1-196, 158 tab. h.t.
Lafon P., 2019. La succession végétale dans les Landes de Gascogne et la position de l’Avoine de Thore (Pseudarrhenatherum longifolium). Evaxiana 6 : 131-150.
Lafon P., Leblond N. & Caze G., 2014. État des lieux sur la présence en Aquitaine de l’habitat d’intérêt communautaire UE 9330 « Forêts à Quercus suber » en contexte non littoral. Audenge, Conservatoire botanique national Sud-Atlantique, DREAL Aquitaine, 22 p. + annexes.
Lafon P., Henry E., Caze G. & Bébot P., 2018. Étude cartographique pour l’intégration dans le réseau Natura 2000 de l’habitat d’intérêt communautaire UE 9330 « Forêts à Quercus suber » en contexte non littoral. Conservatoire botanique national Sud-Atlantique, 22 p. + annexes.
Lafon P. (coord.), Mady M., Corriol G. & Belaud A. 2020. Catalogue des végétations de Nouvelle-Aquitaine. Classification, chorologie et correspondances avec les habitats européens. Conservatoire botanique national Sud-Atlantique / Conservatoire botanique national Massif central / Conservatoire botanique national des Pyrénées et de Midi-Pyrénées, 272 p.
Lapeyrère E., 1892. Flore du département des Landes. Imp. Hazael Labèque, Dax, 510 p.
Lemée G., 1937. Recherches écologiques sur la végétation du Perche. Thèse, Paris, Librairie générale de l’enseignement, 385 p.
Loiseau J.-E. & Felzines J.-C., 2007. Les groupements des pelouses à Corynephorus canescens des vallées de l’Allier et du cours moyen de la Loire (Auvergne, Bourgogne, Centre, France). Nouvelle composition des Corynephoretalia canescentis. Journal de botanique de la Société botanique de France 39 : 57-77.
Loiseau J.-E. & Felzines J.-C., 2010. Les groupements du Koelerio-Phleion des alluvions de l’Allier et de la Loire (Auvergne, Bourgogne, Centre, France). Apports synsystématiques. Journal de botanique de la Société botanique de France 40 : 281-350.
Mucina L., Bültmann H., Dierben K., Theurillat J.-P., Raus Th., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Y.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M. & Tichý L., 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 19 (1) : 3-264.
Paul P., 1969. Études expérimentales sur le déterminisme de la composition floristique des pelouses xérophiles. II – L’action autoécologique du sol en conditions naturelles et controlées. Acta Oecologia IV : 47-70.
Paul P. & Richard Y., 1968. Études expérimentales sur le déterminisme de la composition floristique des pelouses xérophiles. I – L’action autoécologique du sol sur les espèces calcicoles et calcifuges. Acta Oecologia III : 29-48.
Royer J.-M., Felzines J.-C., Misset C. & Thévenin S., 2006. Synopsis commenté des groupements végétaux de la Bourgogne et de la Champagne-Ardenne. Bulletin de la Société botanique du Centre-Ouest,NS, numéro spécial 25 : 1-394.
Royer J.-M. & Ferrez Y., 2020. Contribution au prodrome des végétations de France : les Festuco – Brometea Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, série 3, 13 : 5-304.
Sissingh G. & Tideman P., 1960. De Plantengemeenschappen uit de omgeving van Didam en Zevenaar. 30 p.
Theurillat J.-P., Willner W., Fernández-González F., Bültmann H., Čarni A., Gigante D., Mucina L. & Weber H., 2020. International Code of Phytosociological Nomenclature, 4th edition. Applied Vegetation Science 24 (2) : 1-62.
Tison J.-M. & Foucault B. (de) (coords), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Trichet P., Jolivet C., Arrouays D., Loustau D., Bert D. & Ranger J., 1999. Le maintien de la fertilité des sols forestiers landais dans le cadre de la sylviculture du pin maritime. Étude et gestion des sols 6 (4) : 197-214.
Remerciements
Nous avons le plaisir de remercier Pierre-Marie Le Hénaff (CBNMC) et Mickaël Mady (CBNMC) pour leur relecture de cet article. Nous remercions la DREAL Nouvelle-Aquitaine pour le soutien aux études de connaissances des végétations de la région dont cet article valorise certains relevés acquis dans ce cadre.


