
Contribution à la connaissance des communautés végétales orophiles du Massif central
Title
Contributions to the knowledge of Massif central orophilous vegetation communities
Résumé
À la suite d’un travail de trois ans sur les réserves naturelles nationales du Sancy (Le Hénaff et al., 2021), il est apparu que la connaissance de certaines végétations subalpines du Massif central restait incomplète et qu’une analyse à l’échelle de l’ensemble du Massif central était nécessaire. Après trois années d’inventaires complémentaires, l’analyse de près de 470 relevés phytosociologiques permet de proposer la description précise de quatre végétations emblématiques des sommets du Massif central. De par leur fragilité face aux changements globaux, la localisation précise de ces végétations permettra d’engager avec les partenaires une stratégie de conservation multisite.
Abstract
After three years of work on the Sancy’s national nature reserves (Le Hénaff et al., 2021), it appears that knowledge of several subalpine vegetations was incomplete and that a global analysis at the scale of Massif central was still needed. After three successive years dedicated to additional list, the analysis of 470 relevés allow us to offer a definitive description of four emblematic vegetations located on the summits of the Massif central. Because of their inherent fragility in the face of global warming, their geographical location will allow partners to engage in a strategy of multi-site management.
1. L’étage subalpin du Massif central
1.1. Généralités
Occupant une vaste zone de l’ordre de 80 000 km², le Massif central est situé à un carrefour biogéographique (aux influences atlantique, continentale et méditerranéenne). Le relief de cette région, issu en partie de l’orogénèse alpine puis de l’édification de vastes ensembles volcaniques, se caractérise par une grande diversité de roches (cristallines, sédimentaires, volcaniques). Cette diversité exerce une influence majeure sur les communautés végétales de ce territoire. L’existence d’un étage subalpin dans le Massif central est documentée depuis de nombreuses années dans la littérature. Il ne s’exprime que dans les plus hauts massifs de la région, à savoir les monts Dore, monts du Cantal, mont Mézenc, monts du Forez, ainsi que sur le mont Lozère. Le point culminant du Massif central est situé dans le département du Puy-de-Dôme, au puy de Sancy, à 1 886 m d’altitude, suivi de près par le Plomb du Cantal à 1 856 m.
Cet étage de végétation est notamment caractérisé par une pluviométrie abondante (plus de 2 000 mm/an) dans les monts Dore et les monts du Cantal. L’enneigement est assez important, mais peut connaître des variations interannuelles plus ou moins importantes selon les années. À noter que la durée d’enneigement moyenne a diminué ces dernières années entre 1980-2010 (Serre, 2015). La température moyenne annuelle est peu élevée, de l’ordre de moins de 5 °C au-dessus de 1 600 m (Thébaud & Roux, 2018). Les définitions de cet étage sont variables selon les auteurs et se base tantôt sur des paramètres climatiques (moyenne des températures annuelles), tantôt sur des limites altitudinales. Par exemple pour Rameau et al. (1993), la caractéristique principale est « une température annuelle moyenne de -2 °C à + 4 °C et une période de végétation variant entre 100 et 200 jours », tout en précisant que de nombreuses variations existent. Dans le Sancy, cette température annuelle est de l’ordre de 5,15 °C à 1 660 m. Selon Löve (in Klesczewski et al., 2020) « l’étage subalpin est l’étage naturel situé sous l’étage sans arbres, avec comme limite inférieure la forêt montagnarde dense et comme limite supérieure celle des arbres [treeline] ». Bien entendu les conditions de vent et de neige des plus hautes altitudes, la faible épaisseur des sols interdisent une présence des arbres aux plus hautes altitudes et nous considérons donc l’étage subalpin au-delà de cette limite sur le Massif central. Ces particularités écologiques permettent l’expression d’une flore subalpine dans les principaux massifs montagneux de la dition et récemment synthétisée par Klesczewski et al. (2020). Il convient de noter que l’instabilité des coulées de cendre indurées (cinérites) qui s’érodent facilement limite la fermeture du tapis herbacé et constitue un facteur favorisant le maintien d’espèces pionnières et peu compétitives, dans un contexte où le climat est moins froid et la durée d’enneigement moins longue que dans les hauts massifs alpins ou pyrénéens.
1.2. Le Cantal, le plus grand stratovolcan d’Europe
Sans rentrer dans les détails de la construction d’un tel édifice (voir les nombreuses publications de P. Nehlig sur le sujet), il convient de retenir qu’il se présente aujourd’hui comme un cône surbaissé de 70 km de diamètre et qu’il présentait originellement une altitude de plus de 3 000 m. L’encaissement des vallées et la masse de cet édifice ont permis le maintien de conditions favorables à certaines espèces subalpines qui s’expriment dans ce massif mieux que dans toute autre partie du Massif central.
1.3. La flore orophile du Massif central
Sans vouloir prétendre à effectuer une analyse approfondie de l’origine de la flore subalpine du Massif central, nous rappellerons quelques principaux éléments détaillés par Braun-Blanquet (1923). À la suite de la migration survenue durant les dernières périodes glaciaires, de nombreuses espèces pyrénéennes sont parvenues à s’implanter durablement dans le sud du Massif central (Vivarais, Cévennes, Aubrac). À partir des monts du Forez, ce front de colonisation s’épuise fortement et devient encore plus ténu vers le nord-est (monts du Beaujolais et du Lyonnais). Quelques rares espèces sont toutefois parvenues à s’implanter jusque dans les Vosges (Luzula desvauxii, Androsace halleri). Braun-Blanquet (1923) relève à juste titre que la partie sud occidentale du Massif central (incluant l’Auvergne) se rapproche tant floristiquement que du point de vue des communautés végétales des Pyrénées. C’est le cas notamment avec la présence de Silene ciliata, Sagina saginoides ou encore Jacobaea leucophylla, localisé en dehors des Pyrénées seulement sur les sommets du massif du Mézenc. Une part non négligeable de la flore orophile du Massif central est commune avec les massifs alpins et pyrénéens ; on peut citer Trifolium alpinum, Sagina saginoides, Phyteuma hemisphaericum, Helictochloa versicolor, Euphrasia minima, Bupleurum ranunculoides, Dryas octopetala, Carex curvula (photos 1 et cartes 1). De récentes études génétiques (Schönswetter et al., 2015 ; Kyrkou et al., 2015) viennent confirmer les hypothèses émises par Braun-Blanquet (1923).



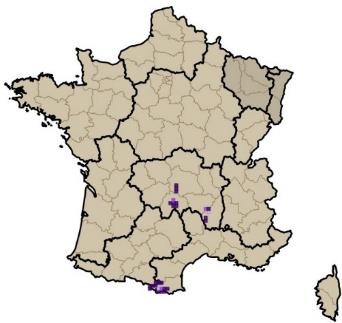
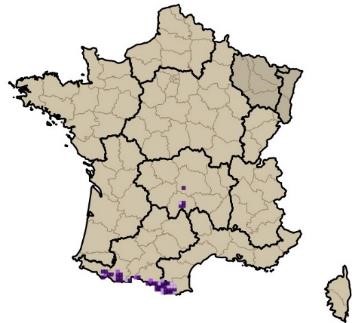
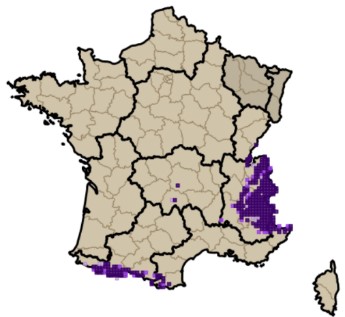
1.4. Bref rappel historique des connaissances des communautés orophiles du Massif central
Du fait de ces caractéristiques, les phytosociologues se sont intéressés depuis près de cent ans à ces communautés végétales (Braun-Blanquet, 1926 ; Luquet, 1926 ; Quézel & Rioux, 1954 ; Cusset & de Lachapelle, 1962). Ces auteurs ont principalement étudié les formations des landes montagnardes (Genisto pilosae-Vaccinion uliginosi), des mégaphorbiaies montagnardes à subalpines (Adenostylion alliariae), des pelouses subalpines thermophiles (Festucion variae) et mésophiles (Nardion strictae), des communautés chasmophytiques ou encore des formations forestières.
À la suite de ces premières vagues de travaux, il faudra attendre les études de Bock & Prelli (1975), Coquillard et al. (1994), puis de Michalet & Philippe (1996) pour avoir une connaissance satisfaisante des communautés végétales orophiles du Massif central. À la suite de Coquillard et al. (1994), ces derniers auteurs proposent de retenir la classe des Caricetea curvulae avec la description du Jasiono arvernensis-Agrostietum rupestris dont on retrouve les premières mentions dans des travaux antérieurs (Cusset & de Lachapelle, 1962 ; Bock & Prelli, 1975).
Plus récemment, les synthèses phytosociologiques menées dans le cadre de la déclinaison du Prodrome des végétations de France au niveau national ont conduit Corriol & Mikolajczak (2017) à ne pas retenir la classe des Caricetea curvulae en Auvergne, car jugeant que l’association du Sancy est floristiquement trop appauvrie et non représentative de la classe. Par analogie, la faible typicité floristique des communautés des combes à neige du Massif central a conduit Corriol & Mikolajczak (2014) à ne pas retenir la classe des Salicetea herbaceae dans la dition. Cette interprétation était déjà partagée par Bock & Prelli (1975), pour qui « il serait abusif de voir dans les combes à neige de Chaudefour un groupement des Salicetea herbaceae Braun-Blanq. 1947, la majorité des espèces les rattachent au Nardion ».
Ces divergences d’interprétation sur les communautés des pelouses subalpines du massif du Sancy ont conduit Le Hénaff et al. (2021a), suivis par Le Hénaff et al. (2021b), à proposer l’alliance de l’Euphrasio minimae-Nardion strictae, afin de caractériser les pelouses acidiphiles du montagnard supérieur et du subalpin du Massif central, des plaques à neige enrichies en espèces des Caricetea curvulae. Du fait d’une appartenance incertaine à une classe donnée, le Jasiono crispae-Agrostietum rupestris a également été intégré dans cette alliance. Ce choix s’avère peu satisfaisant vis-à-vis de la définition écologique de l’Euphrasio minimae-Nardion strictae et des fortes différences floristiques qui existent entre cette association et le reste de l’alliance.
Dernièrement, Boeuf et al. (2022) proposent une nouvelle alliance du Massif central (Festucion airoidis) qui caractérise les pelouses mésoxérophiles érodées des crêtes ventées, acidiphiles, de l’étage subalpin du massif des monts Dore. Cette proposition repose à nouveau la question de la présence de la classe des Caricetea curvulae dans le Massif central et remet en cause les conclusions de Corriol & Mikolajczak (2017).
Pour terminer ce tour d’horizon, il s’avère que les études des systèmes pelousaires orophiles du Massif central se sont concentrées majoritairement sur le massif du Sancy. Les communautés du massif cantalien ont été très peu étudiées en comparaison et elles sont globalement méconnues. Dans le but d’affiner l’état des connaissances syntaxonomiques des pelouses orophiles du Massif central, nous proposons une étude globale de ces communautés végétales en y intégrant des communautés relevant des Festucetalia spadiceae (Nardetea strictae). Face aux opinions divergentes sur la présence ou non de la classe des Caricetea curvulae, cet article apporte un matériel complémentaire et inédit permettant de contribuer à éclaircir cette question.
2. Matériel et méthodes
2.1. Jeu de données et échantillonnage
Le jeu de données utilisé dans le cadre de cette étude comprend un nombre important de relevés effectués dans le Massif central, répartis au sein de plusieurs régions naturelles, allant des monts du Forez, la Montagne ardéchoise, les monts Dore et aux monts du Cantal, quelques relevés provenant du mont Aigoual. Il s’agit notamment de relevés effectués par des phytosociologues du Conservatoire botanique national du Massif central, ainsi que la bancarisation d’articles portant sur les communautés végétales orophiles du Massif central (Michalet & Philippe, 1996 ; Billy, 2002). Afin de compléter cet échantillonnage régional, nous avons également intégré des relevés publiés dans la littérature et réalisés dans le massif pyrénéen (Braun-Blanquet, 1948 ; Corriol et al., 2023). Ils se rapportent à des communautés végétales relevant de la classe des Nardetea strictae et de l’ordre des Festucetalia spadiceae, des Caricetea curvulae et des Elyno-Seslerietea pour les communautés pyrénéennes. Au total, ce sont 470 relevés phytosociologiques qui ont été mobilisés dans le cadre de cette étude.
2.2. Analyse des données
Les relevés ont été saisis dans la base de données Lobelia du Conservatoire botanique national du Massif central, hormis les publications du massif pyrénéen. Ils ont été analysés à l’aide du logiciel Juice (Tichý, 2002). Pour ce faire, plusieurs classifications ascendantes hiérarchiques (CAH) ont été réalisés (en présence-absence). Les groupes mis en évidence ont été obtenus par la méthode de Ward (mesure de distance de Jaccard). Des analyses factorielles de correspondances (AFC) ont été menées sur les groupes obtenus précédemment, ou portant sur un jeu de données partiel. La mise en évidence des espèces caractéristiques des syntaxons obtenus a été faite au travers du logiciel Juice par le calcul de l’indice de fidélité Phi (Chytrý et al., 2002). Enfin, des analyses de correspondances canoniques (CCA), couplées avec la bio-indication des espèces renseignée dans la base de données Baseflor (Julve 1997), ont permis des caractérisations écologiques.
2.3. Syntaxonomie et nomenclature
La syntaxonomie proposée ici suit le cadre conceptuel de la phytosociologie sigmatiste, reconnue à l’échelle européenne et faisant l’objet d’un code de nomenclature et régulièrement mis à jour (Theurillat et al., 2020). Cette méthodologie permet notamment de travailler dans un cadre partagé à l’échelle européenne. Elle s’appuie plus particulièrement sur le Prodrome des végétations de France et ses déclinaisons portant sur la classe des Nardetea strictae (de Foucault, 2012), Caricetea curvulae (Corriol & Mikolajczak, 2017), Elyno-Seslerietea (Corriol et al., 2023). Des ouvrages régionaux ainsi que plusieurs publications récentes ont été pris en compte dans ce travail (Thébaud et al., 2014 ; Thébaud & Roux 2018 ; Thébaud et al., 2021 ; Le Hénaff et al., 2021a et b ; Boeuf et al., 2022). Afin de ne pas alourdir le texte, les autorités des syntaxons ne sont pas mentionnées sauf dans des cas très précis. Un index syntaxonomique est présenté en annexe (tableau 11) et est consultable en ligne (https://www.cbnmc.fr/33-ressources/90-outils-pratiques/91-listes-et-referentiels).
La nomenclature taxonomique suit le référentiel TAXREF v. 16 (Gargominy et al., 2022). Face aux difficultés inhérentes à la détermination de certains genres, nous avons dû procéder à des ajustements taxonomiques. Ainsi, les observations des Hieracium ont été regroupées dans des sections. Il en va de même pour les Festuca, dont de nombreuses données anciennes ont été regroupées au sein de deux sections (F. sec. Aulaxyper et sec. Festuca). Toutefois, certaines espèces bien présentes dans la dition n’ont pas fait l’objet de tels regroupements (Festuca nigrescens, F. airoides, F. lemanii). Nous avons procédé de même pour les observations des espèces du genre Alchemilla, très hétérogènes selon les sources des relevés. Les différentes espèces mentionnées ont été regroupées dans les sections Alchemilla sect. Alpinae et A. sect. Alchemilla.
Concernant les fétuques de ces pelouses, il est à noter que, si Festuca airoides apparaît être un taxon bien circonscrit et dont les données anciennes peuvent être jugées fiables, l’existence de populations diploïdes et tétraploïdes mérite des investigations complémentaires. D’autre part, des confusions ont pu avoir lieu entre F. billyi et F. arvernensis du fait de la présence à haute altitude de populations non glauques pour cette dernière. Une étude phylogénétique inter-CBN est en cours sur l’ensemble des fétuques de France et permettra pour le Massif central de trancher sur les questions concernant l’ensemble ovina s.s./airoides/niphobia/niphobioides d’une part et l’ensemble arvernensis/billyi d’autre part. Si les niveaux de ploïdie ne sont pas les mêmes, les corrélations avec des traits morphologiques restent à affiner.
3. Résultats
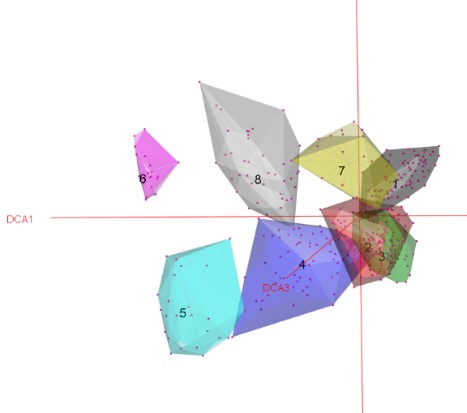
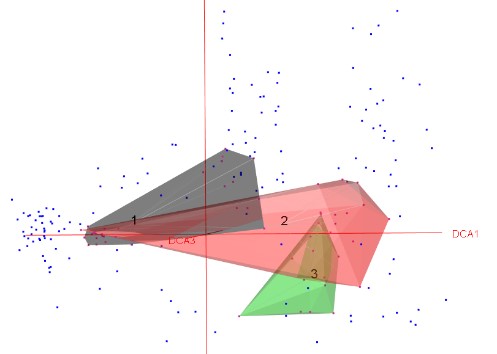
Des analyses préliminaires portant sur le jeu de données ont conduit à écarter du matériel (relevés non saturés). Ainsi ce sont près de 70 relevés, considérés comme non rattachables à un syntaxon, qui n’ont pas été utilisés pour la suite des analyses. Une analyse sur un jeu de données plus restreint (397 relevés) par une CAH permet de faire ressortir huit clusters (figure 1), répartis en deux branches principales. Cette première analyse permet de faire ressortir nettement des groupes de syntaxons floristiquement et écologiquement homogènes.

3.1. Analyse statistique
Résultats de la classification hiérarchique
Les clusters 1 à 4 vont distinguer des communautés du domaine montagnard à subalpin. Le premier embranchement regroupe l’ensemble des pelouses mésophiles structurées par Patzkea paniculata et Calamagrostis arundinacea. Le cluster 2 intègre les pelouses à Nardus stricta et se différencie négativement par rapport au troisième cluster. Ce dernier s’enrichit nettement de plusieurs espèces, à savoir Ranunculus serpens, Pilosella officinarum, Galium verum, Thymus pulegioides, Genista tinctoria et dans une moindre mesure G. sagittalis. Un tel cortège floristique indique des communautés acidiclinophiles et non strictement acidiphiles. Le cluster 4 possède une tonalité nettement plus subalpine à alpine et hygroclinophile vis-à-vis des groupes précédents, ce qui se traduit par la présence de Trifolium alpinum, Luzula desvauxii, Epikeros pyrenaeus, Plantago alpina, Mutellina adonidifolia et dans une moindre mesure Euphrasia minima. Ces groupements s’enrichissent en espèces alpines telles que Phyteuma hemisphaericum et Agrostis ruprestris. Toutefois, les liens avec les pelouses mésophiles (clusters 2 et 3) sont encore marqués, avec la fréquence importante de Nardus stricta, Avenella flexuosa, Agrostis capillaris et Galium saxatile.
Les clusters 5 à 8 regroupent l’ensemble des communautés mésophiles à thermophiles, subalpines à alpines de notre jeu de données. Les clusters 5 et 6 comprennent les communautés des Pyrénées orientales et centrales. Les deux derniers clusters vont rassembler des communautés héliophiles, oligotrophiles, acidiclinophiles à acidiphiles et mésoxérophiles du Massif central. Ils sont floristiquement distincts, le premier (cluster 7) rassemblant l’ensemble des pelouses à Festuca airoides de la dition, le dernier embranchement comportantdes pelouses à Patzkea paniculata. Là encore, il est intéressant de noter que cette analyse fait ressortir des « effets massifs » déjà mentionnés par plusieurs auteurs sur ce type de végétation (Corriol & Mikolajczak, 2017 ; Corriol et al., 2023 ; Peyre & Font, 2011).
Résultats des AFC et CCA
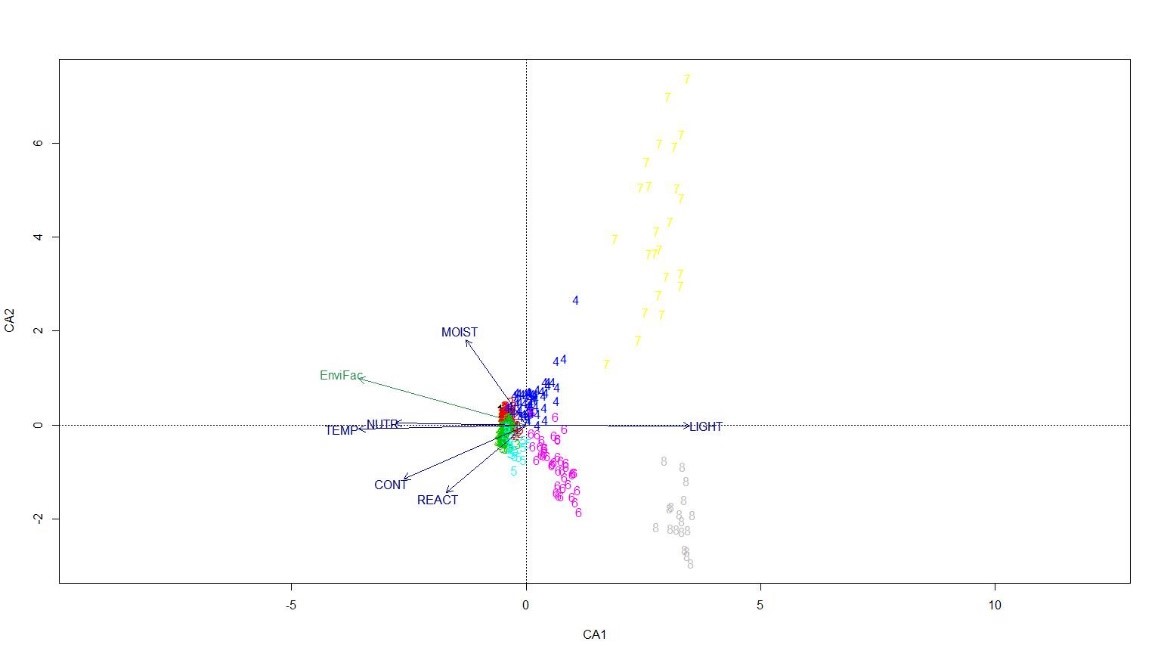
La projection sur les trois premiers axes des clusters obtenus précédemment est présentée ci-dessous (figure 2). L’axe 1 dans sa partie positive permet de dissocier les relevés comportant une flore nettement alpine à subalpine. L’axe 2 peut s’interpréter comme l’axe permettant de dissocier les clusters le long d’un gradient thermique. Dans sa partie positive, cet axe distingue des communautés xérophile, cryophile et anémophile, s’opposant dans sa partie négative à des communautés bénéficiant d’un apport hydrique plus important. Il est intéressant d’observer que les pelouses à Patzkea paniculata (mésophiles [1] et thermophiles [7]) sont relativement proches sur cette AFC, et relativement bien décorrélées des pelouses à Festuca airoides (8) du Massif central. Les principales variables écologiques permettant d’expliquer les gradients que l’on peut observer sont principalement la lumière, l’humidité édaphique, la richesse en nutriment et la température (figure 3). Là encore, on observe également une forte distinction des communautés pyrénéennes. Il est intéressant de noter une certaine convergence écologique et floristique des relevés du cluster 4 (Euphrasio minimae-Nardion strictae) vers les communautés en provenance des Pyrénées centrales. On peut émettre comme hypothèse que ces deux types de communautés végétales bénéficient toutes les deux d’un apport hydrique assez conséquent avec les chutes de neige (Michalet & Philippe, 1996 ; Corriol & Mikolajczak, 2017 , se traduisant par la présence conjointe de Trifolium alpinum, Euphrasia minima et plus marginalement d’Omalotheca supina.
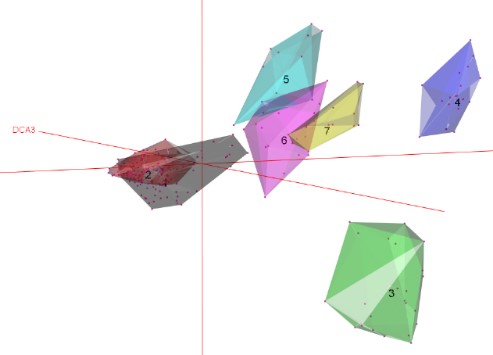
Une seconde analyse partielle (non prise en compte des communautés à Patzkea paniculata) centrée sur les communautés à Festuca airoides du Massif central, permet de mettre en évidence trois communautés végétales différentes dont la figure 4 illustre ce résultat (groupes 5, 6 et 7).
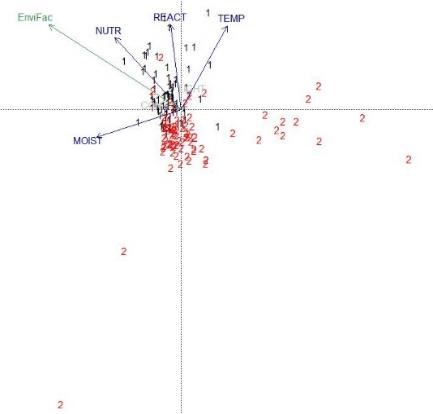
Nos analyses discriminent ainsi deux communautés (groupes 5 et 6) structurées par Festuca airoides et qui sont très liées à des secteurs géographiques distincts, à savoir les monts Dore et les monts du Cantal. La dernière communauté végétale (groupe7) est quant à elle structurée par Dryas octopetala. Une analyse centrée sur cette espèce a permis la comparaison des relevés de la dition (3) avec des végétations homologues (1 et 2) du massif pyrénéen (figure 5). Les relevés auvergnats sont plus proches du groupe 2, qui correspond à des communautés végétales mésophiles à mésoxérophiles (Corriol et al., 2023) relevant de l’alliance de l’Oxytropido-Elynion se trouvant éloignée des communautés mésophiles du Salicion pyrenaicae.
En dernier lieu, nous avons procédé à une analyse partielle portant sur les pelouses à Nardus stricta du Massif central. Les résultats obtenus avec la CAH distinguent nettement deux types de communautés. Une CCA indique que les paramètres de température, de réaction et de la richesse en nutriments sont les variables permettant d’expliquer ces différences (figure 6).
Caractérisation écologique
Une caractérisation écologique des communautés végétales a été faite en utilisant les valeurs issues de la base des traits de vie de Julve (2007), en présence-absence. Nous présentons ci-dessous les quatre principales variables étudiées (figure 7).
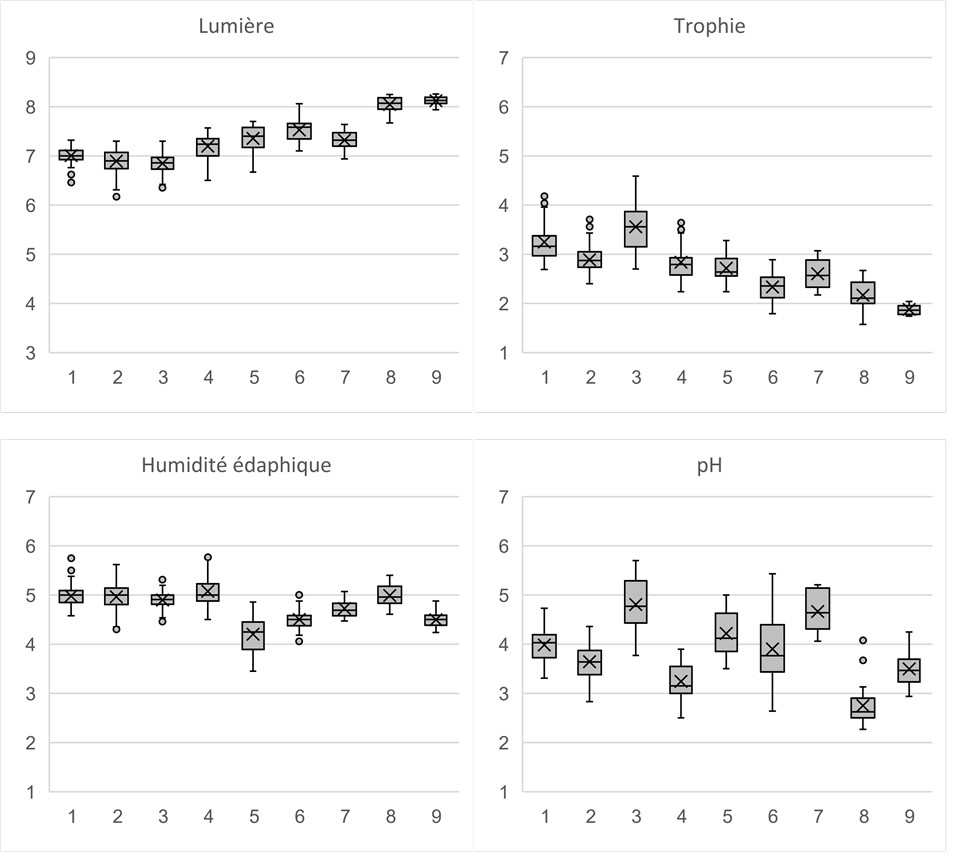
Le paramètre de luminosité est peu variable pour l’ensemble de ces communautés, il s’agit de façon générale de végétations héliophiles. On peut noter toutefois une augmentation de ce paramètre pour le Festucion variae et le Festucenion supinae du Massif central. Ils sont tous les deux situés aux plus hautes altitudes d’Auvergne, ce qui explique cette augmentation du fait de la hausse du rayonnement solaire (notamment les ultraviolets).
Hormis pour le Galio-Patzkeion, ces communautés sont oligotrophiles, voir peroligotrophiles pour les pelouses à Festuca airoides du Massif central et des Pyrénées. Le Galio-Patzkeion présente une plus grande variation sur ce paramètre, certaines associations sont à la charnière avec les mégaphorbiaies mésophiles mésotrophiles du Calamagrostietum arundinaceae.
Toutes les pelouses dominées ou co-dominées par Nardus stricta sont mésophiles, de même que l’Euphrasio-Nardion (communautés des zones d’accumulation de neige et bénéficiant ainsi d’un apport en eau plus conséquent). Les communautés du Festucion variae et du Festucenion airoidis du Massif central sont situées dans des contextes plus contraignants et sont mésoxérophiles à mésophiles (constat similaire dans les Pyrénées). Les communautés à Dryas octopetala sont quant à elles plutôt mésophiles.
Le pH est le paramètre qui présente les variations les plus importantes. Il est intéressant de constater que les syntaxons du Massif central du Festucenion airoidis et du Festucion variae sont acidiclinophiles, ce qui est rarement mentionné dans la littérature (Corriol & Mikolajczak 2017 ; de Foucault 2012). Il en va de même pour le Galio-Patzkeion, considéré comme acidiphile par de Foucault (2016), mais qui s’avère être plutôt acidiclinophile. Concernant les communautés à Dryas octopetala, nous pouvons émettre comme hypothèse qu’un humus brut s’est développé au fil des années, acidifiant le substrat. Toutefois elles se développent bien sur des roches volcaniques (pyroclastites) basiques.
3.2. Présentation détaillée des communautés végétales inédites
Syntaxon relevant du Festucion supinae
Sileno ciliatae-Festucetum airoidis ass. nov. hoc loco (photos 2 et 3), typus nominis : rel 2631781 (voir tableau 1) ; ce relevé type est retranscrit ci-dessous :
- Silene ciliata 2, Pulsatilla vernalis 1, Pedicularis comosa +, Androsace halleri 1, Festuca airoides 2, Agrostis rupestris 1, Luzula spicata 1, Alchemilla transiens 1, Phyteuma hemisphaericum 1, Helictochloa versicolor 1, Cerastium alpinum 1, arvense subsp. strictum +, Alchemilla flabellata r, Dianthus gratianopolitanus +, Antennaria dioica 2, Leucanthemum delarbrei r, cf. Alchemilla saxatilis r, Dianthus hyssopifolius i, Galium saxatile r, Carex caryophyllea 1, Thymus pulegioides +, Galium pumilum +, Avenella flexuosa +, Calluna vulgaris +, Genista pilosa 1, Vaccinium myrtillus +, Valeriana tripteris i.


Le tableau 1 propose quatorze relevés inédits de cette association. Cette pelouse occupe les secteurs des crêtes ou des vires rocheuses peu inclinées de l’étage subalpin supérieur, des sols superficiels issus de roches volcaniques (basanite et trachyandésite) des monts du Cantal. La durée d’enneigement est plus faible que sur d’autres secteurs du fait de son positionnement sur les crêtes (peu propices à l’accumulation de la neige sur une longue période). La forte luminosité estivale, combinée à des sols peu profonds et l’action du vent, induit une dessiccation importante de cette pelouse dès le début du mois d’août. La variante type est enrichie en espèces des pelouses acidiclinophiles des Nardetalia strictae et des Festucetalia spadiceae (Carex caryophyllea, Thymus pulegioides, Lotus corniculatus, Galium pumilum, Campanula rotundifolia, Gentiana lutea, Noccaea caerulescens, Thesium alpinum). Elle se développe sur des roches faiblement acides de type basanite, ce qui permet l’expression de ce cortège floristique (facteur édaphique moins contraignant). Elle assure également la transition avec des pelouses de l’ordre des Festucetalia spadiceae (Nardetea strictae). La variante appauvrie est dépourvue des espèces citées précédemment et se développe sur des roches légèrement plus acides, de type trachyandésite, ce qui pourrait expliquer cette différente et une richesse spécifique moindre. Des recherches complémentaires seraient nécessaires afin de statuer sur le statut syntaxonomique à accorder à ces deux variantes (sous-associations éventuelles). Nous mettons en évidence une dernière variation à Koeleria pyramidata et Festuca arvernensis qui correspond semble-t-il à des communautés se développant en contrebas des crêtes et qui sont donc moins soumises à l’action contraignante du vent.
Il est frappant de constater que cette association possède de nombreuses similitudes écologiques et floristiques avec le Hieracio pumili-Festucetum supinae décrit par Braun-Blanquet (1948) des Pyrénées orientales. Cette dernière s’en distingue par un cortège d’espèces endémiques lié à l’étage alpin (absent du Massif central) de la chaîne des Pyrénées (tableau 7). Nous faisons le choix d’élever au rang d’association la communauté cantalienne, dès lors que le Hieracio pumili-Festucetum est circonscrit uniquement aux Pyrénées orientales (Corriol & Mikolajczak 2017). Le Sileno cilatae-Festucetum peut donc être considéré comme une association vicariante, floristiquement appauvrie, traduisant des liens existant entre ces deux massifs lors des phases de migration de la flore pyrénéenne en dehors de ce massif.
Cette pelouse semble se maintenir naturellement dans certains contextes, tels que les vires rocheuses. Toutefois, sur certains secteurs, elle est située dans des parcs de pâturage extensif. Les animaux peuvent donc avoir une action de blocage dynamique, sous réserve que la pression de pâturage soit très faible pour éviter un surpiétinement et une déstructuration du tapis végétal. Cette pelouse semble évoluer très progressivement vers une lande typique des crêtes du Cantal, le Pulsatillo vernalis-Cytisetum decumbentis (tableau 5 ; photo 4), dont on retrouve plusieurs espèces caractéristiques de la pelouse étudiée ici. Il s’agit selon Thébaud et al. (2021) d’une lande xérophile, cryophile et anémophile. Sur des sols plus profonds, cette pelouse peut être en contact avec des landes montagnardes mésophiles et cryophiles du Genisto pilosae-Vaccinion uliginosi (Vaccinietum uliginoso-myrtilli ; photo 5), mais principalement avec des pelouses acidiphiles ou acidiclinophiles montagnardes relevant du Galio saxatilis-Potentillion aureae ou du Nardo-Agrostion tenuis.


Syntaxon de l’alliance du Nardo-Agrostion tenuis
Ranunculo serpentis-Nardetum strictae (Billy ex Thébaud, C. Roux, C.-E. Bernard & Delcoigne 2014) stat. nov. hoc loco (basionyme : Diantho sylvatici-Leontodontetum helvetici galietosum veri Billy ex Thébaud, C. Roux, C.-E. Bernard & Delcoigne in Thébaud et al. 2014, Guide d’identification des végétations du nord du Massif central : 257, rel. G664) photos 6 et 7). Le relevé G664 correspond dans notre tableau 2 au numéro 2080299, que nous retranscrivons ci-dessous :
- Galium verum 1, Ranunculus serpens 1, Thymus pulegioides +, Meum athamanticum 1, Scorzoneroides pyrenaica 1, Viola lutea 1, Dianthus seguieri pseudocollinus +, Potentilla aurea +, Gentiana lutea 1, Poa chaixii 1, Epikeros pyrenaeus 1, Trifolium alpinum 1, Thesium alpinum (X), Nardus stricta 2, Agrostis capillaris 1, Potentilla erecta 1, Avenella flexuosa 1, Galium saxatile 1, Jasione laevis 1, Pilosella officinarum +, Anthoxanthum odoratum 1, Festuca nigrescens 1, Betonica officinalis +, Campanula rotundifolia +, Luzula multiflora +, Euphrasia officinalis subsp. rostkoviana +, Carex leporina +, Stellaria graminea +, Achillea millefolium +, Veronica officinalis +, Rhinanthus minor +, Trifolium repens +, T. pratense 1, Centaurea nigra 1, Bistorta officinalis 1, Rumex acetosa +, Cynosurus cristatus (X), Sanguisorba officinalis 1.
Cette pelouse occupe les estives d’altitude de l’étage montagnard supérieur, plus rarement à l’étage subalpin inférieur, sur des sols modérément acides d’origine volcanique, oligotrophes. Elle occupe également des secteurs mieux exposés que les communautés du Galio-Potentillion, permettant l’expression d’une flore thermoclinophile. On observe un cortège d’espèces différentielles des Festuco-Brometea important et constant (Genista sagittalis, Lotus corniculatus, Thymus pulegioides, Briza media, Genista tinctoria, Galium pumilum).
Elle présente de nombreuses similarités écologiques et floristiques avec le Diantho pseudocollini-Festucetum nigrescentis. En effet il s’agit de pelouses oligotrophiles, des sols modérément acides, ce qui avait conduit Le Hénaff et al. (2021b) à l’inclure dans la variabilité de cette association. Toutefois, afin d’assurer une certaine cohérence globale du synsystème régional, il semble nécessaire de conserver la distinction de deux associations caractérisant différents sous-étages du domaine montagnard (supérieur et inférieur). Le Ranunculo serpentis-Nardetum strictae correspond aux pelouses du montagnard supérieur enrichi en espèces orophiles (Scorzoneroides pyrenaica, Potentilla aurea, Trifolium alpinum) contrairement au Diantho pseudocollini-Festucetum nigrescentis. Nous avons intégré à titre de comparaison les colonnes synthétiques de cette dernière association dans le tableau 2, ainsi que celle du Diantho pseudocollini-Meetum athamantici décrit par de Foucault (1986) de l’Aubrac. Cette dernière association est à la limite des Festucetalia spadiceae et possède un fond floristique commun avec les deux autres associations. Elle possède encore des espèces pelousaires collinéennes (Avenula pubescens, Prunella hastifolia[1], Potentilla verna, Poterium sanguisorba, Trifolium ochroleucon, Pimpinella saxifraga, Linum catharticum, Anthyllis vulneraria, Armeria arenaria), justifiant son intégration dans le Violion caninae. On peut distinguer au sein du Ranunculo serpentis-Nardetum une variante d’ordre trophique avec une augmentation du cortège des espèces mésotrophiles des Arrhenatheretea.


Syntaxon de l’Oxytropido–Elynion myosuroidis
L’examen des relevés à disposition permet de faire ressortir un lot important d’espèces possédant des affinités avec la classe des Elyno myosuroidis-Seslerietea caeruleae avec la présence de Bistorta vivipara, Dryas octopetala, Cerastium alpinum, Sabulina verna, Gentiana verna et Alchemilla alpigena. Du fait du caractère nettement cryophile de ce groupement, il possède des affinités floristiques et écologiques fortes avec l’ordre des Oxytropido-Elynetalia myosuroidis. En effet les Seslerietalia caeruleae et les Astragaletalia sempervirentis regroupent des communautés subalpines à alpines, mésophiles à xérophiles. Au sein des Oxytropido-Elynetalia myosuroidis, ces communautés à Dryas octopetala relèvent potentiellement de deux alliances : le Salicion pyrenaicae et l’Oxytropido-Elynion myosuroidis. La première alliance, considérée comme strictement inféodée au massif pyrénéen, se distingue de la seconde par des communautés uniquement psychrophiles et cryophiles. L’Oxytropido-Elynion myosuroidis comporte des végétations xérophiles, car exposées aux vents. La présence dans nos relevés de Luzula spicata, Euphrasia minima et plus ponctuellement de Phyteuma hemisphaericum plaide pour une intégration du syntaxon auvergnat dans l’alliance de l’Oxytropido-Elynion myosuroidis. En l’état actuel de nos connaissances, il ne semble pas possible, du fait d’un appauvrissement floristique évident par rapport aux communautés des Alpes et Pyrénées, d’intégrer le syntaxon auvergnat dans l’une des sous-alliances décrites par Corriol et al. (2023). A noter l’introgression d’espèces chasmophytiques telles que Saxifraga fragosoi, S. paniculata et S. lamottei, indiquant des liens avec des communautés du Dianthion gratianopolitani. Cette particularité a conduit Le Hénaff et al. (2021a) à intégrer les communautés à Dryas octopetala dans le Saxifragetum lamottei, en y distinguant plusieurs variantes. Selon eux, la variante à Dryas octopetala « se localise au niveau d’éperons rocheux très fracturés. Cette dernière variante, plus pelousaire, se retrouve généralement au niveau de pentes moins prononcées ; elle est très riche en espèces ». Nous désignerons par la suite cette végétation sous le nom de groupement à Luzula spicata et Dryas octopetala (tableau 6 ; photo 8).

Syntaxon de l’Arctostaphylo-Cetrarion nivalis
Parmi les relevés disponibles, on peut observer quatre relevés qui se distinguent par la présence et une dominance des chaméphytes plus importantes (Vaccinium uliginosum, Empetrum nigrum), combinée avec la forte dominance de Dryas octopetala. En l’état, il semble délicat d’intégrer ces relevés dans une classe pelousaire. Du fait du caractère psychrophile et neutroclinophile à acidiclinophile de ce groupement, il semble pertinent d’intégrer ces relevés dans la classe des landes des Loiseleurio procumbentis-Vaccinietea microphylli, l’ordre des Rhododendro ferruginei-Vaccinietalia microphylli et l’alliance de l’Arctostaphylo-Cetrarion nivalis. Cette dernière rassemble les landes artico-alpines calcicoles, dans lequel semble pouvoir s’intégrer ce groupement. Nous le désignerons par la suite sous le nom de groupement à Vaccinium uliginosum et Dryas octopetala (tableau 6 ; photo 9).

4. Discussions
Les analyses menées confirment l’originalité et la pertinence du Galio saxatilis-Patzkeion paniculatae (de Foucault, 2016). Il est intéressant de souligner que cette alliance présente une signature écologique analogue à celle du Caricion ferrugineae (Elyno myosuroidis-Seslerietea caeruleae), à savoir des pelouses montagnardes et subalpines, riches en hautes herbes. Le Galio-Patzkeion possède un fond floristique des Nardetalia strictae et des Festucetalia spadiceae. Il s’enrichit en espèces des mégaphorbiaies mésophiles subalpines du Calamagrostion arundinaceae (Mulgedio-Aconitetea), et plus particulièrement du Calamagrostietum arundinaceae décrit par Luquet (1926)[2] (photo 10). On peut le considérer comme un vicariant acidiclinophile à neutroclinophile des pelouses basiphiles du Caricion ferrugineae. Il serait intéressant de rechercher cette alliance en dehors du Massif central, notamment dans les Pyrénées et éventuellement les Alpes afin d’améliorer la chorologie globale de cette alliance. Il existe en revanche une dissymétrie dans le traitement de ce type de communautés. En contexte acide, elles sont intégrées dans les Nardetea strictae et non les Caricetea curvulae, contrairement aux pelouses basiphiles homologues, qui relèvent des Elyno-Seslerietea et non des Festuco-Brometea. Deux tableaux synthétiques sont disponibles en annexe, présentant en détail les associations qui y sont intégrées.

Le branchement dans notre CAH du Festucion variae à proximité des communautés du Festucion supinae (Pyrénées et Massif central) ne remet pas en cause selon nous les conclusions de de Foucault (1994, 2012) sur la place de cette alliance. Au regard des travaux menés par cet auteur, ainsi que ceux de Corriol & Mikolajczak (2017), cette alliance s’intègre bien à l’ordre des Festucetalia spadiceae. Ce choix est fondé sur le vaste tableau synthétique de cette classe proposé par de Foucault (1994), présenté en annexe (tableau 8) auquel nous y avons intégré de nouvelles alliances décrites a posteriori. À notre connaissance, le Festucion variae n’a pas été lectotypifié. La description d’origine de Braun-Blanquet (1926) étant invalide (art. 2b), le type doit être désigné dans Guinochet (1938) qui la valide. Du fait que le Centaureo uniflorae-Festucetum spadiceae Lippmaa 1933 (syn. Centaureo uniflorae-Festucetum spadiceae Guin. 1938) est versé par de Foucault (2012) dans l’Hyperico richeri-Festucion paniculatae, le type ne peut être que le Festuco scabriculmis-Potentilletum valderiae. Toutefois une telle lectotypification devrait théoriquement entraîner une correction du nom de l’alliance, ce qui dépasse le cadre de notre étude. Nous présentons ici un tableau inédit de relevés rapportés à cette association (tableau 3) provenant du massif du Cantal, auquel nous avons ajouté le relevé historique de Braun-Blanquet. La composition floristique indique nettement une communauté thermophile, oligotrophile et acidiclinophile. Nous mettons également en évidence un groupement à Patzkea paniculata et Festuca arvernensis (photos 11 et 12), floristiquement et écologiquement proche du Leucanthemo delarbrei-Patzkeetum paniculatae. Festuca arvernensis est l’espèce qui domine physionomiquement cette végétation, ou plus rarement Festuca billyi. Nous pouvons observer une diminution des espèces du Festucion variae, de même que certaines espèces des Festucetalia spadiceae et des Nardetalia strictae. L’étude de ce groupement sera à poursuivre dans les années à venir, il pourrait s’agir d’une sous-association du Leucanthemo delarbrei-Patzkeetum paniculatae ou un syntaxon autonome. Ce groupement est lié à des secteurs rocheux bien exposés, chauds, sur des sols très superficiels. Des communautés similaires semblent exister dans le Mézenc et dont l’étude devra être poursuivie.


Les relevés rapportés par Michalet et Philippe (1996) des monts Dore au Leucanthemo delarbrei-Patzkeetum paniculatae (chamaespartietosum sagittalis) ne peuvent selon nous s’y rapporter. En effet les contextes stationnels sont très différents de ceux de nos observations et de celles de Braun-Blanquet en 1926 (pentes rocheuses chaudes). Dans le massif du Sancy, de nombreux relevés ont été faits sur des cinérites, ce qui se traduit par une flore plus mésophile (Linaria vulgaris, Knautia basaltica, Hypericum maculatum, Silene vulgaris, Solidago virgaurea). Ces relevés doivent être rapportés à une communauté du Galio saxatilis-Patzkeion. La mention du Festucion variae dans le Massif central doit être réservée aux situations chaudes, et non mésiques. C’est pourquoi nous revenons sur la proposition de Le Hénaff (2021a) d’intégrer le Trifolio villosi-Avenuletum pubescentis dans le Festucion variae, cette association se plaçant beaucoup mieux au sein du Galio saxatilis-Patzkeion. La présence de telles communautés dans le massif des monts Dore est très rare, elles s’expriment davantage dans les monts du Cantal. La présence du Leucanthemo delarbrei-Patzkeetum paniculatae (photo 13) dans le massif du Mézenc (Lemée, 1953) serait à confirmer par la réalisation de nouveaux relevés, ainsi que dans la montagne ardéchoise. Un tableau synthétique présentant la différence entre ces deux alliances est disponible en annexe (tableaux 7 et 10).

La première description d’une pelouse acidiclinophile du montagnard supérieur à Nardus stricta dans le Massif central semble avoir été faite par Braun-Blanquet (1926). L’unique relevé (n° 1710123 dans notre tableau 2) comporte en effet Ranunculus serpens, Genista sagittalis, Lotus corniculatus et Thymus pulegioides. Nous ne pouvons en revanche retenir le Festuco rubrae-Nardetum strictae Braun Blanq. 1926 cité par de Foucault (1994 ; 2012). En effet, Braun Blanquet (1926) ne décrit pas une telle association mais propose une sous-association à Festuca rubra du Plantagini alpinae-Nardetum strictae. Or cette association est un homonyme du Plantagini alpinae-Nardetum strictae Luquet 1926 et est rejetée par Thébaud et al. (2014) qui la lectotypifie dans les travaux de Luquet (1926). Le Festuco rubrae-Nardetum strictae Braun Blanq. 1926 que l’on retrouve dans de nombreuses publications est un nom fantôme.
La thèse de Luquet (1926) ne semble pas pouvoir être utilisée, les sous-associations proposées (« sous-association à Nardus stricta et Festuca rubra », « sous-association à Deschampsia flexuosa et Agrostis capillaris ») du Plantagini alpinae-Nardetum strictae sont illégitimes et doivent être rejetées (art. 13, 34c). À noter que, bien que ces sous-associations soient illégitimes, elles restent néanmoins valides. La composition floristique de ces nardaies les rapproche du Diantho pseudocollini-Scorzoneroidetum pyrenaicae (photo 14). En l’absence de tableaux effectifs de relevés, il est difficile de savoir si Luquet (1926) a réellement étudié une nardaie acidiclinophile. Il est probable que quelques relevés aient été réalisés dans ce contexte. Nous tenons à signaler qu’en théorie le principe de priorité reviendrait à retenir l’autorité de Luquet et non Lachapelle pour désigner ces pelouses acidiphiles du montagnard supérieur. Toutefois, il nous semble important d’assurer une relative stabilité nomenclaturale, du fait que ce syntaxon est largement reconnu par les phytosociologues régionaux et nationaux. Il conviendra à l’avenir de proposer à la commission de nomenclature un nom conservé pour éviter toute ambiguïté.

Les travaux de Cusset et Lachapelle (1962) ne peuvent pas davantage être exploités. En effet, ils proposent de façon invalide et illégitime le Nardeto-Leontodontetum pyrenaici, un homonyme antérieur ayant déjà été décrit dans la littérature (Leontodonto helvetici-Nardetum strictae Bartsch 1940). On peut noter toutefois que la colonne synthétique disponible semble proche de la communauté relevée dans le Cantal par Braun Blanquet (1926). Quant à la proposition de Cusset & Lachapelle (1962) du Nardeto-Festucetum rubrae, elle n’est reprise dans aucun travail régional ou national. Ils indiquent qu’il s’agirait d’une nardaie artificielle et le tableau synthétique fourni indique assez nettement une communauté acidiphile du Galio saxatilis-Potentillion aureae. Elle ne peut donc pas être synonyme du Plantagini alpinae-Nardetum strictae festucetosum rubrae de Braun-Blanquet. La présence avec des fréquences plus ou moins importantes d’Alchemilla vulgaris, Ajuga reptans, Rumex acetosella, Trifolium pratense, T. repens, Leontodon autumnalis laisse supposer qu’il s’agit bien d’une communauté dégradée par l’action du surpâturage, ou des relevés effectués sur des contextes écologiques non homogènes. Ce constat était également partagé par Billy (2000) et nous pouvons qu’abandonner cette association.
Coquillard (1993) étudie dans le massif du Sancy une nardaie riche en espèces des Festuco-Brometea et qu’il place dans le Violion caninae. Toutefois cette description est invalide (art. 1) et le nom proposé (Polygalo vulgaris-Nardetum strictae) a déjà été publié antérieurement (Oberdorfer, 1957). Elle occupe des sols d’origine volcanique (doréites, trachy-andésites, cinérites et basaltes) modérément acides. Ce type de roches permet l’expression d’un cortège floristique acidiclinophile caractéristique de cette association. Il faudra néanmoins attendre les travaux de Billy (2002) sur cette pelouse de l’étage montagnard supérieur pour stabiliser la nomenclature phytosociologique. Il y distingue un type nettement acidiphile et une sous-association plus neutroclinophile.
Avec la mise en évidence du Nardo-Agrostion tenuis dans le Massif central, il nous semble important de revenir à la diagnose d’origine de cette alliance et de son association type publié par Sillinger (1933). Cet auteur y décrit une communauté acidiclinophile de l’étage montagnard (Potentilla aurea, Viola lutea, Phleum alpinum, Poa alpina, Antennaria dioica) des Carpates qu’il nomme Nardetum montanum. Ce syntaxon est illégitime et devrait être rejeté (art. 34a). L’auteur y distingue deux variantes, la première sur des sols enrichis en bases et la seconde sur des sols siliceux. L’ensemble des relevés comporte des taxons affines des sols légèrement acides à neutres (Lotus corniculatus, Pimpinella saxifraga, Briza media, Galium verum, Pilosella officinarum). La première variante s’enrichit quant à elle d’espèces nettement calcicoles (Plantago media, Carlina vulgaris). Cette description a conduit de Foucault (1994, 2012) à considérer le Nardo-Agrostion tenuis comme une alliance caractérisant des pelouses acidiclinophiles de l’étage montagnard. Il s’agit d’une alliance similaire au Violion caninae, réservé à l’étage collinéen et submontagnard. La mention de cette nouvelle alliance dans le Massif central permet ainsi une meilleure cohérence globale du synsystème, à savoir une structuration analogue entre l’étage collinéen et montagnard :
| Collinéen | Montagnard | |
| Acidiclinophile | Violion caninae | Nardo-Agrostion tenuis |
| Acidiphile | Galio saxatilis-Festucion filiformis | Galio saxatilis-Potentillion aureae |
Dans le Massif central, cette alliance comprend donc deux associations, le Ranunculo serpentis-Nardetum strictae et le Diantho pseudocollini-Festucetum nigrescentis, anciennement intégrées dans le Galio saxatilis-Potentillion aureae. Cette dernière alliance est à réserver aux syntaxons strictement acidiphiles (tableaux 7 et 8). À noter que de Foucault (1994, 2012) n’a pas la même conception de cette alliance que Mucina et al. (2016), pour qui il s’agit de communautés mésophiles, montagnardes à subalpines d’Europe centrale et des Balkans. Nous suivons ici l’interprétation de Foucault (1994 ; 2012), qui fait primer les facteurs édaphiques plutôt que chorologiques.
Le rejet du Plantagini alpinae-Nardetum strictae Braun-Blanq. 1926 par Thébaud et al. (2014) entraîne de facto le rejet du Nardion strictae Braun-Blanq. 1926, largement repris dans la littérature européenne (Mucina et al., 2016). Cet espace syntaxonomique pour caractériser ces pelouses liées aux systèmes des plaques à neige tardive du Massif central a conduit Le Hénaff et al. (2021a) à proposer l’Euphrasio minimae-Nardion strictae (photo 15). On trouvera en annexe les tableaux synthétiques (tableaux 7 et 9), présentant en détail cette alliance et les associations qui lui sont subordonnées. Il serait intéressant d’étudier la présence potentielle de cette alliance en dehors du Massif central. Il est fait mention dans les Vosges (Collaud et al., 2016) d’un syntaxon (Sibbaldio procumbentis-Nardetum strictae Carbiener in Collaud et al. 2016), écologiquement et floristiquement proche de certaines associations placées dans l’Euphrasio-Nardion telles que le Plantagini alpinae-Nardetum strictae Luquet 1926. Cette association pourrait être intégrée dans cette alliance plutôt que dans le Galio saxatilis-Potentillion aureae.

La présence des Caricetea curvulae dans le Massif central ne fait pas consensus (Michalet & Philippe, 1996 ; Thébaud et al., 2014 ; Corriol & Mikolajczak, 2017 ; Bœuf et al., 2022). Jusqu’à récemment, cette divergence d’opinion se basait uniquement sur l’association décrite du Sancy, qui est de façon indiscutable floristiquement très appauvrie. Les investigations menées dans le massif cantalien ont mis en évidence une association nouvelle, floristiquement mieux caractérisée. Le Sileno ciliatae-Festucetum comporte des espèces des Nardetealia et Festucetalia spadiceae. Cette particularité est en contradiction avec Corriol et Mikolajczak (2017), pour qui cette classe est totalement dépourvue de ces espèces. Nous tenons à rappeler que, dans le Massif central, il n’existe pas d’étage alpin et que l’association du Cantal se trouve notamment sur des roches volcaniques moyennent acides (basaltes) permettant à ces espèces de s’exprimer plus aisément, auquel cas il aurait fallu verser cette association (et par ricochet le Jasiono arvernensis-Agrostietum rupestris) dans la classe des Nardetea strictae. Une intégration dans l’ordre des Festucetalia spadiceae n’étant pas possible du fait des trop fortes différences floristiques et écologiques, nous aurions dû proposer un nouvel ordre pour caractériser ces pelouses. Selon nous cette solution n’est pas pertinente et complexifierait à outrance la nomenclature phytosociologique pour délimiter ces pelouses orophiles. C’est pourquoi nous retenons la classe des Caricetea curvulae comme étant présente dans le Massif central, rejoignant les conclusions de plusieurs travaux régionaux (Michalet & Philippe, 1996 ; Thébaud et al., 2014 ; Thébaud & Roux, 2018 ; Bœuf et al., 2022).
Les récentes propositions de Bœuf et al. (2022) de retenir pour les Pyrénées orientales le Festucion niphobiae Braun-Blanq. 1948 nom. corr. Boeuf, Lazare, S. Riba & Thébaud in Bœuf (2022) (cf. art. 43) [basionyme : Festucion supinae Braun Blanq. 1948 nom. inept.] ajoutent un flou syntaxonomique sur la caractérisation écologique de ces pelouses. Selon ces auteurs, cette alliance regroupe les associations de pelouses cryophiles, de climax climatiques, se développant à l’étage alpin et pouvant comporter ou non Festuca airoides. Ils considèrent que cette alliance est vicariante du Festucion airoidis R. Michalet & Th. Philippe ex Boeuf, Thébaud & Lazare in Bœuf 2022, qui caractérise « la zone des plus hautes crêtes ventées érodées, acidoxériques des monts Dore au-dessus de 1600 m, sujette à une dynamique secondaire d’origine anthropique ». Au-delà du problème de validité de cette alliance (l’association type désignée est invalide [art. 16]), il nous semble judicieux de mettre en perspective la pertinence écologique de cette nouvelle alliance vis-à-vis du type (Jasiono crispae-Agrostietum rupestris). Cette association est un syntaxon très appauvri, essentiellement composée d’espèces de la classe et plus rarement du niveau de l’ordre et de la sous-alliance. En l’état, cela reviendrait à considérer le Festucion airoidis comme une alliance d’appauvrissement, sans espèces différentielles, ce qui nous semble peu souhaitable. Nous jugeons préférable d’inclure les syntaxons du Massif central au sein du Festucion supinae et qu’il s’agit d’associations d’appauvrissement chorologique dont le Jasiono crispae-Agrostietum rupestris représenterait l’extrême irradiation septentrionale de cette alliance d’optimum pyrénéo-cantabrique. De plus, Bœuf et al. (2022) précise que la proposition de mutation de Rivas-Martínez et al. (2002) du Festucion supinae en Festucion airoidis Braun-Blanq. 1948 ne semble pas avoir été examinée par la commission de nomenclature. Selon eux, le Festucion supinae est un nom. inept. et doit être abandonné. Les problèmes soulevés par ces auteurs devront être éclaircis dans les années à venir afin de stabiliser la nomenclature de ces pelouses. Dans l’attente d’un avis officiel de la commission de nomenclature, nous suivrons le synsystème proposé par Corriol & Mikolajczak (2017).
Nous sommes également amenés à reconsidérer la place du Biscutello arvernensis-Festucetum airoidis, (association des vires rocheuses à Festuca airoides) publié par Billy (2002) et validé par Thébaud et al. (2014), qui l’intègrent dans le Dianthion gratianopolitani, décrit par Focquet (1982). Plus récemment Bœuf et al. (2022) l’intègrent dans l’Asarinion procumbentis, alliance des communautés chasmophytiques des roches siliceuses riches en fougères rupicoles (Mady et al., 2022). Il est important dans un premier temps de rappeler les réserves émises par Billy (2002) sur l’intégration de cette association à la classe chasmophytique des Asplenietea trichomanis. Il précise que « les individus d’association que j’ai réunis ici ne sont plus aussi nettement saxicoles. À vrai dire, leur habitat mérite plutôt d’être qualifié de rochers herbeux et pourtant ce sont ceux où Dianthus gratianopolitanus est le mieux représenté. […] Pour les phytosociologues, l’appartenance de ce groupement aux Asplenietea doit paraître assez douteuse et l’on pourrait aussi penser au Sedo-Scleranthion mais la place prise par l’œillet et la raiponce me semble autoriser de le classer au moins provisoirement à la suite des deux précédents. » Une comparaison des associations de cette alliance avec le Jasiono arvernensis-Agrostietum rupestris et le Sileno ciliatae-Festucetum airoidis (Le Gloanec, 2023) montre clairement que le Biscutello arvernensis-Festucetum airoidis ne relève pas des Asplenietea trichomanis (absence totale d’espèces typiques de la classe) mais bien des Caricetea curvulae. Les relevés effectués par Billy (2000) ont probablement été réalisé sur de faibles surfaces et dans des contextes stationnels favorisant l’expression de Dianthus gratianopolitanus (très faible épaisseur du sol). La seule présence de D. gratianopolitanus n’est pas un argument floristique suffisamment important pour permettre un rattachement aux Asplenietea. En effet, cette espèce est fréquente dans le Diantho gratianopolitani-Melicion ciliatae, caractérisant des communautés climaciques rupicoles des bords des falaises et des rochers mésothermes. Cette alliance est intégrée aux Festuco-Brometea par Royer (1991). De fait, nous considérons que cette association est devenue source d’ambiguïté et doit être rejetée : Biscutello arvernensis-Festucetum airoidis Billy ex Thébaud, C. Roux,
C.-E. Bernard & Delcoigne 2014 nom. amb. propos. (art. 36).
L’interprétation des relevés à Dryas octopetala est très délicate du fait que ces communautés sont extrêmement rares dans le Massif central et occupent des surfaces très restreintes. En réalité, le cortège d’espèces caractéristiques de ces végétations n’est pas bien exprimé et largement incomplet en comparaison avec les massifs alpins et pyrénéens. La distinction entre communautés landicoles et pelousaires est peu aisée et se base avant tout sur la dominance globale des chaméphytes et est donc subjective. On peut toutefois noter une fréquence a priori plus importante des espèces pelousaires (Caricetea curvulae et Elyno-Seslerietea) au sein du groupement à Luzula spicata et Dryas octopetala que dans le groupement landicole. Là encore, le choix d’intégrer ces relevés au sein de l’alliance de l’Oxytropido-Elynion est sujet à interprétation et se base avant tout plus sur des considérations écologiques que floristiques. En effet, une part non négligeable d’espèces caractéristiques de cette alliance (Corriol et al., 2023) est absente du Massif central, exception faite de quelques espèces différentielles (Euphrasia minima, Luzula spicata, Phyteuma hemisphaericum), mais aussi d’ordre et de classe. Ces communautés du Massif central représentent des isolats géographiques, très appauvris dont l’appartenance phytosociologique est et restera incertaine.
Mise à jour synomenclaturale de divers syntaxons de pelouses du Massif central
Dans le cadre du catalogue des végétations agropastorales du Massif central (Le Hénaff et al., 2021), il a été proposé de faire évoluer les noms des syntaxons en accord avec la nomenclature botanique actuelle. L’utilisation de l’article 44 n’était pas correcte au sens du code de nomenclature phytosociologique, l’évolution du nom dans les cas présentés ici relevant en fait de l’article 45 relatif à la mutation des noms, en accord avec l’évolution de la nomenclature. Afin d’assurer l’homogénéité des noms utilisés dans le référentiel des végétations du Massif central (CBN Massif central 2021) les mutations sont ici proposées.
Les pelouses du Galio saxatilis-Patzkeion paniculatae
- Arnico montanae-Patzkeetum paniculatae Michalet & T. Philippe ex Thébaud, C. Roux,
C.-E. Bernard & Delcoigne 2014 nom. mut. nov. hoc loco (art. 45) ; photos 16 et 17 - Trollio europaei-Avenelletum flexuosae Michalet & T. Philippe ex Thébaud, C. Roux,
C.-E. Bernard & Delcoigne 2014 nom. mut. nov. hoc loco (art. 45) - Vicio orobi-Patzkeetum paniculatae Michalet & T. Philippe ex Thébaud, C. Roux,
C.-E. Bernard & Delcoigne 2014 nom. mut. nov. hoc loco (art. 45)
Les pelouses du Festucion variae
- Leucanthemo delarbrei-Patzkeetum paniculatae Braun-Blanq. 1926 mut. nov. hoc loco (art.45)


5. Perspectives
Les systèmes pelousaires orophiles du Massif central comportent une importante diversité floristique ainsi que des communautés végétales. Nous espérons que cette contribution permettra une meilleure connaissance de ces systèmes patrimoniaux pour le Massif central. Au-delà des aspects typologiques développés dans cet article, il serait souhaitable d’avoir des actions de conservation de ces milieux naturels, menacés à plus ou moins long terme par diverses menaces (changements des pratiques agropastorales, surfréquentation par les randonneurs, changement climatique).
[1] Cette espèce peut être considérée comme caractéristique locale, car très présente dans le sud du Massif central, et devient nettement plus rare dans le centre de l’Auvergne.
[2] Bien que citée des monts du Cantal dans la littérature (Thébaud et al., 2014), il ne semblerait qu’aucun relevé provenant de ce massif ait été publié jusqu’à présent. Le tableau 4 en annexe propose six relevés rapportés à cette association, attestant de sa présence de manière effective dans le massif cantalien.
Bibliographie
Billy F., 2002. Végétations pionnières en Basse-Auvergne. Bulletin de la Société botanique du Centre-Ouest, n. s., n° spécial 22 : 1-197.
Bock Ch. & Prelli R., 1975. Notice explicative de la carte des groupements végétaux du Cirque de Chaudefour (Mont-Dore). Avernia Biologica, n. s., 1-26.
Bœuf R., Hardion L., Šmarda P., Lazare J.-J., Thébaud G., Gregor T., Portal R., Höcker R., Riba Mazas S., Besnard G., & Holveck P., 2022. Des Carpates aux Pyrénées, centrées sur l’Alsace et les Vosges, analyses génétiques de quelques fétuques à feuilles fines (Festuca sect. Festuca) estimées d’intérêt régional à européen : considérations taxinomiques et syntaxinomiques. Botanique 8 : 37-133.
Braun-Blanquet J., 1923. L’origine et le développement des flores dans le Massif central de la France. Paul Lechevalier, Paris, 282 p.
Braun-Blanquet J., 1926. Le climax complexe des landes alpines. In Études phytosociologiques en Auvergne, rapport sur une excursion inter-universitaire, Arvenia 2 : 29- 48.
Braun‑Blanquet J., 1948. La végétation alpine des Pyrenées‑Orientales. Monografia de la estacion de estudios pirenaicos y del instituto español de edafologia, ecologia y fisiologia vegetal 9 : 1‑306.
Chytrý M., Tichý L., Holt J. & Botta-Dukát Z., 2002. Determination of diagnostic species with statistical fidelity measures. Journal of Vegetation Science 13 : 79-90.
Coquillard P., 1993. Dynamique des systèmes agro-pastoraux de l’étage montagnard du massif du Sancy et de la chaîne des Puys : variations biologiques et fonctionnelles ; exemple d’application au modèle à Calluna vulgaris. Thèse de doctorat universitaire UAM 3, Faculté de Saint-Jérôme, 266 p.
Coquillard P., Gueugnot J., Julve Ph., Michalet R. & Michelin Y., 1994. Carte écologique du Massif du Sancy au
1/25 000. Ecologia mediterranea XX (1-2) : 9-57.
Collaud R., Ferrez Y., Simler N. & Voirin M., 2017. Typologie phytosociologique des végétations agropastorales du Massif des Vosges. Les Nouvelles Archives de la Flore jurassienne et du nord-est de la France 14 : 139-174.
Corriol G. & Mikolajczak A., 2014. Contribution au Prodrome des végétations de France : les Salicetea herbaceae Braun-Blanq. 1948. Journal de botanique de la Société botanique de France 68 : 15-49.
Corriol G. & Mikolajczak A., 2017. Contribution au prodrome des végétations de France : les Caricetea curvulae Braun-Blanq. 1948 nom. conserv. propos. Journal de botanique de la Société botanique de France 77 : 57-96.
Corriol G., Sanz Th. & Van Es J., 2023. Contribution au Prodrome des végétations de France : les Elyno myosuroidis-Seslerietea caeruleae Braun-Blanq. 1948. Carnets botaniques 121 : 1-86.
Cusset G. & de Lachapelle B., 1962 (1961). Études botaniques dans les monts Dore. Revue de la Société d’histoire naturelle d’Auvergne, n. s., 27 (1-4) : 2-82.
Focquet P., 1982. La végétation des parois siliceuses de la vallée de la Vésubie (Alpes-Maritimes, France). Documents phytosociologiques, n. s., VII : 5-188.
Foucault B. (de), 1986. Contribution à une étude systémique des prairies de l’Aubrac (Massif central français). Documents phytosociologiques, n. s., X (1) : 255-305.
Foucault B. (de), 1994. Essai synsystématique sur les pelouses sèches acidophiles (Nardetea strictae, Caricetea curvulae). Colloques phytosociologiques XXII, Syntaxonomie typologique des habitats : 431-454.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Nardetea strictae Rivas Goday in Rivas Goday & Rivas-Mart. 1963. Journal de botanique de la Société botanique de France 59 : 241-344.
Foucault B. (de), 2016. Errata et compléments à propos du Prodrome des végétations de France, quatrième note. Journal de botanique de la Société botanique de France 75 : 63-68.
Gargominy O., Tercerie S., Régnier C., Ramage T., Dupont P., Daszkiewicz P. & Poncet L., 2022. TAXREF, référentiel taxonomique pour la France : méthodologie, mise en œuvre et diffusion. Rapport PatriNat (OFB-CNRS-MNHN), Muséum national d’histoire naturelle, Paris. 47 pp.
Klesczewski M., Pouget L., Lecoq M., Oudot M., Bossaert M. & Kelian G., 2020. Contribution à la connaissance de l’étage subalpin dans le massif du mont Aigoual (Gard, Lozère) : synthèse bibliographique, délimitation, caractérisation floristique et comparaison à l’échelle du Massif central. BIOM 1 (1) : 49-68.
Kyrkou I., Iriondo J.-M. & García-Fernández A., 2015. A glacial survivor of the alpine Mediterranean region: phylogenetic and phylogeographic insights into Silene ciliata Pourr. (Caryophyllaceae). PeerJ. 3 : e1193.
Julve Ph., 2017. Baseflor, http://philippe.julve.pagesperso-orange, fr/catminat.htm.
Le Gloanec V., 2023. Étude des pelouses subalpines à Festuca airoides du Massif central. Caractérisation typologique et habitats. Conservatoire botanique national du Massif central / Région-Auvergne-Rhône-Alpes, 42 p. + annexes.
Le Hénaff P.-M., Hostein C., Dumont M. & Pradinas R., 2021a. Catalogue des végétations des réserves naturelles nationales du massif du Sancy. Conservatoire botanique national du Massif central / Direction régionale de l’environnement, de l’aménagement et du logement Auvergne-Rhône-Alpes, 215 p.
Le Hénaff P.-M., Galliot J.-N., Le Gloanec V. & Ragache Q., 2021b. Végétations agropastorales du Massif central, catalogue des milieux ouverts herbacés. Conservatoire botanique national du Massif central, 531 p.
Lemée G., 1953. Observations sur la végétation actuelle et son évolution postglaciaire dans le massif du Mézenc. Bulletin de la Société botanique de France 100 : 67-77.
Luquet A., 1926. Essai sur la géographie botanique de l’Auvergne. Les associations végétales du massif des monts Dore. Thèse, Paris, 267 p.
Mady M., Theurillat J.-P. & Lazare J.-J., 2022. Nomenclatural note on the Antirrhinetum asarinae Braun-Blanquet 1915. Botanique 8 : 199-202.
Michalet R. & Philippe Th., 1996. Les landes et les pelouses acidiphiles de l’étage subalpin des monts Dore (Massif central français) : syntaxonomie et potentialités dynamiques. Colloques phytosociologiques XXIV, Fitodinamica: I differenti aspetti della dinamica vegetale : 434-471.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Y.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M. & Tichý L., 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Journal of Vegetation Science 19 (1) : 3-264.
Oberdorfer E., 1957. Süddeutsche Pflanzengesellschaften. Pflanzensoziologie 10 : 1-564.
Peyre G. & Font X., 2011. Syntaxonomic revision and floristic characterization of the phytosociological alliances corresponding to subalpine and alpine grasslands of the Pyrenees and Cantabrian Mountains (classes Caricetea curvulae, Carici‑Kobresietea, and Festuco‑Seslerietea). Plant Biosystems 145, suppl.1 : 220‑232.
Quézel P. & Rioux J., 1954. L’étage subalpin dans le Cantal (Massif central de France). Vegetatio 4 (6) : 345-378.
Royer J.-M., 1991. Synthèse eurosibérienne, phytosociologique et phytogéographique de la classe des Festuco-Brometea. Dissertationes Botanicae 178 : 1-296 + tableaux.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal, addenda to the syntaxonomical checklist of 2001, I. Itinera Geobotanica 15 (1) : 5-432.
Schönswetter P., Magauer M. & Schneeweiss G., 2015. Androsace halleri subsp. nuria Schönsw. & Schneew. (Primulaceae), a new taxon from the eastern Pyrenees (Spain, France). Phytotaxa 201 : 227-232.
Sillinger P., 1933. Monografická studie o vegetaci Nízkych Tater. Library of the research board for Slovakia and Ruthenia of the Slav institute 6 : 1-339.
Serre F., 2015. Les singularités du climat et son évolution récente dans le massif des monts Dore et dans la réserve naturelle nationale de Chastreix-Sancy. Réserve naturelle nationale de Chastreix-Sancy / Direction régionale de l’Aménagement, de l’Environnement et du Logement Auvergne, 48 p.
Thébaud G., Roux C., Bernard C.-É. & Delcoigne A., 2014. Guide d’identification des végétations du nord du Massif central. Associations végétales et habitats naturels. Presses universitaires Blaise-Pascal, Clermont-Ferrand, 296 p.
Thébaud G. & Roux C., 2018. Végétations montagnardes et subalpines du Massif central français. Botanique 4 : 111-138.
Thébaud G., Choisnet G. & Roux C., 2021. Contribution to the survey of the heathlands of the French Massif central (habitats 4030 and 4060): analysis of phytosociological data. BIOM 2-2021 : 62-109.
Theurillat J.-P., Willner W., Fernández‐González F., Bültmann H., Čarni A., Gigante D., Mucina L. & Weber H., 2020. International code of phytosociological nomenclature, 4th edition. Applied Journal of Vegetation Science, https://doi.org/10.1111/avsc.12491.
Tichý L., 2002. JUICE, software for vegetation classification. Journal of Vegetation Science 13 : 451-453.
Remerciements
L’étude des végétations du Massif central ne pourrait se faire sans le soutien continu de la direction régionale de l’Environnement, de l’Aménagement et du Logement et de la Région Auvergne-Rhône-Alpes au travers de différents outils (CVB – Contrats vert et bleu, CPO, Natura 2000, etc.). Les présents résultats sont issus pour une bonne partie d’une campagne de terrain spécifique réalisée dans le cadre du CVB du Parc naturel régional des Volcans d’Auvergne, ayant reçu le soutien financier de la Région Auvergne-Rhône-Alpes et du Feder Massif central. L’Europe s’engage dans le Massif central avec le Fonds européen de développement régional (Feder). Merci aussi à Olivier Argagnon (CBN méditerranéen) pour la transmission de relevés en provenance des Pyrénées orientales, à Emmanuel Catteau (CBN de Bailleul) pour avoir accepté de relire notre manuscrit et avoir répondu à nos interrogations, ainsi qu’à Luce Mansot (CBN Massif central) pour sa relecture finale. Nous remercions également Gilles Corriol (CBN Pyrénées et Midi-Pyrénées) pour avoir transmis des relevés provenant des Pyrénées centrales et avoir pris le temps d’examiner ce travail, nous permettant une meilleure compréhension et analyses de ces systèmes d’altitude.


