
Un panorama formalisé des ethnosciences naturelles
Title
A formalized panorama of the natural ethnosciences
Résumé
L’auteur présente ici un large panorama illustré (près de 250 photos) des ethnosciences naturelles (ethnobotanique, ethnozoologie, ethnomycologie, ethnominéralogie, ethnophytosociologie…) en utilisant leur formalisation par les opérateurs relationnels introduite en 1987.
Abstract
The author presents here a large illustrated panorama (nearly 250 photos) of natural ethnosciences (ethnobotany, ethnozoology, ethnomycology, ethnominerology, ethnophytosociology…) using their formalization by relational operators introduced in 1987.
1. Introduction
Lors d’un voyage en Guadeloupe vers le milieu des années 1980, j’ai été comme « saisi » par un concept pourtant très classique, celui de la dualité structure-fonction, en entrevoyant, outre son application à la botanique, celle de la formalisation de l’ethnobotanique, puis plus largement des ethnosciences naturelles (Barrau, 1971, 1990 ; de Foucault, 1992a et b ; les ethnosciences autres, telles que ethnomédecine, ethnomathématique, ethnolinguistique, ethnopsychiatrie…, resteront en dehors de ce propos). Sur environ trente-quatre ans, plus de quarante publications ont été consacrées à ces disciplines, soit approximativement 10 % de ma production scientifique, celle-ci étant surtout dédiée à la phytosociologie ; il y eut aussi des cours et des conférences. Ces publications étant très éparpillées, il est apparu qu’une synthèse s’avérait nécessaire en y ajoutant de nouveaux développements. Cette synthèse constitue donc l’objet de cet article qui permet de balayer la plupart des ethnosciences naturelles.
Les publications qui viennent d’être évoquées pêchent par leur manque d’illustrations, ce qui est bien dommage pour ces disciplines mettant en avant la diversité des productions humaines. Cela est aujourd’hui bien plus facile dans Cahiers botaniques et on profite de cette opportunité pour joindre au texte de nombreuses illustrations. Malheureusement (rien n’est parfait…), nombre de celles-ci sont des scans des diapositives réalisées dans mes premières années de réflexion sur ce thème, avant l’avènement des appareils photographiques numériques, d’où une qualité parfois un peu faible des photos associées à ce texte. Pour gérer au mieux ces photos, elles sont numérotées par grand chapitre, par exemple « photo 5-3 » pour la photo 3 du chapitre 5 ; si ce chapitre ne contient qu’une unique photo, l’extension est omise (exemple « photo 6 »).
On trouvera beaucoup de faits ethnoscientifiques dans le Journal d’agriculture tropicale et de botanique appliquée qui fut précédemment nommé Revue de botanique appliquée et d’agriculture coloniale (1921 à 1945), puis Revue internationale de botanique appliquée et d’agriculture tropicale (1946 à 1953), édité par le laboratoire d’ethnobotanique et d’ethnozoologie du Muséum national d’histoire naturelle de Paris ; le contenu en est intégralement disponible sur la plateforme Persée/Sciences de la vie jusque l’année 2004 (volume 42).
Pour faciliter l’accès à ce panorama dense, le tableau en annexe 3 rapproche les opérateurs, les symboles et les abréviations utilisés.
2. Définitions à propos des ethnosciences naturelles
On définit ici une ethnoscience naturelle (terme abrégé en « e.science ») comme l’étude des relations entre les hommes (H) et un groupe d’éléments naturels (E) de leur environnement. En spécifiant plus strictement E, on peut alors distinguer plusieurs e.sciences :
- l’e.botanique (avec E = une plante P) ;
- l’e.zoologie (E = un animal Z; Pujol & Carbone, 1990) ;
- l’e.mycologie (incluant l’e.lichénologie; E = un champignon ou un lichen C ; on rappelle que les lichens sont maintenant inclus dans le règne fongique) ;
- l’e.minéralogie (E = une roche ou un minéral R; Haudricourt, 1968 ; Gonthier, 1990) ;
- l’e.pédologie (E = un sol S) ;
- l’e.météorologie (E = un élément météorologique M) ;
- l’e.astronomie (E = un élément de l’espace A) ;
- l’e.phytosociologie (E = une communauté végétale V ; de Foucault, 1990a)…
L’ensemble e.botanique + e.zoologie + e.mycologie peut être dénommé e.biologie (Castetter, 1944 ; Anderson, 1971). Par ailleurs, pour le règne minéral, je propose d’introduire une nouvelle e.science naturelle, l’ethnogéomorphologie où E est un élément conséquent G du relief, pas seulement des roches isolées ; quelques illustrations en sont déjà offertes par les châteaux du pays cathare (parfois improprement dénommés « châteaux cathares »), dans l’Aude, qui furent édifiés sur des promontoires abrupts pour se protéger et surveiller les éventuels mouvements qui se dérouleraient dans les plaines en contrebas (photo 2-1), par des sites difficilement accessibles (photo 2-2), par les abris en falaise occupés par des groupes humains… De nombreuses grottes en falaise (Chauvet, Cosquer aujourd’hui submergée…) furent utilisées comme abris par les premiers Hominidés et parfois encore actuellement, d’autres sont considérées comme relevant du sacré et donnent lieu à des cultes (cas du littoral marocain de Salé ; Claisse, 1997). De tels sites ont accueilli aussi des moines dès les débuts du bouddhisme à Sri Lanka (photo 2-3).
Il est possible d’étendre ces sciences relationnelles à d’autres types de relations ne faisant plus intervenir directement H en tant acteur (mais il peut rester observateur) :
- zoo-botanique: étude des relations entre animaux et plantes pour nourrir, soigner (avec le cas particulier de la zoopharmacologie ; Huffman, 1995), abriter (nids, fourmilières), fixer (cocon), poser des pièges, utiliser un outil (cas bien connu de l’oiseau des Galapagos utilisant une épine de cactus), se cacher par mimétisme (phasme, phyllie)… ; on peut considérer que c’est une branche de l’éthologie animale ;
- phyto-zoologie: étude des relations entre plantes et animaux pour la reproduction (pollinisation : cas bien connu des Ophrys…), la dispersion (myrmécochorie)… ; on peut considérer que c’est une branche de l’éthologie végétale ;
- zoo-mycologie: étude des relations insectes-champignons (cas de la culture des champignons dans les termitières).



3. Dualité structure-fonction
La dualité structure-fonction est fondamentale pour notre propos comme on l’exposera ci-après, mais déborde aussi vers la sociologie et l’anthropologie (Ledrut, 1990 : 61).
3.1. En biologie
Il est classique en biologie de distinguer une structure anatomique dotée d’une fonction pour l’être vivant auquel appartient cette structure. Plus précisément, cette fonction peut être qualifiée de primaire. En termes sémiologiques, ce binôme, cette dualité peut être considéré(e) comme un sème (double donnée d’un signifiant et d’un signifié selon de Saussure, 1915) *structure = signifiant U/fonction = signifié F* (F pouvant normalement être désigné par un verbe à l’infinitif ; on reconnaîtra des sèmes en ce qu’ils sont notés entre deux astérisques, ses deux termes étant séparés par une barre oblique). En général, il peut y avoir des sèmes *U/0*, où 0 désigne l’absence de fonction (fonction nulle) ; mais en principe il ne peut y avoir de sème *0/F*, c’est-à-dire de fonction ne s’appuyant pas sur une structure matérielle (nous en verrons toutefois une exception dans un cas très particulier ; cf. § 14).
On peut illustrer de tels sèmes primaires par des exemples pris en botanique : *tige/conduire les sèves*, *vrille/accrocher*, *feuille/respirer et réaliser la photosynthèse*, *fleur/reproduire* (photo 3-1).

Une réflexion plus étendue a permis de relier cette dualité à une autre, très classique aussi, la dualité analogie-homologie (de Foucault, 1997b). À partir de multiples exemples botaniques, une loi évolutive assez générale a été dégagée : une fonction donnée est remplie par un organe optimal à un certain stade de l’évolution végétale, mais cette même fonction peut se rencontrer chez des organismes plus primitifs où cet organe n’existe pas, elle y est alors remplie par un organe analogue, mais non homologue, de cet organe optimal ; la fonction précède donc la structure optimale. La photo 3-2 illustre le cas des Torreya, Taxacées dont les graines sont complètement enveloppées par un arille charnu, l’ensemble rappelant ainsi des drupes, dont elles sont analogues mais non homologues.

3.2. En e.sciences
Dans la continuité de la dualité biologique qui vient d’être introduite, on s’aperçoit que beaucoup de faits e.scientifiques associent bien une structure E’ et une fonction F’ attribuée par H à ce E’ pour ses propres besoins, F’ étant alors qualifiée de fonction secondaire. Et l’on peut reconnaître des sèmes de la forme *E’/F’*. E’ dérive en général d’une structure naturelle initiale E et la formalisation des e.sciences (de Foucault, 1987a) propose d’identifier la relation de H à E comme la transformation que H fait subir à E pour la transformer en son dérivé E’ : E’ = aE, où a désigne un opérateur général de transformation (toujours marqué en gras), symbolisant la relation H–E.
Un fait e.scientifique s’inscrit dans un cadre spatial, une aire géographique A, plus ou moins étendue (Amérique du Nord, Togo…), qui peut d’ailleurs être distincte de l’aire naturelle spontanée de E (plus petite, comme dans le cas d’un fait e.scientifique limité à une aire restreinte, donc endémique au sens naturaliste ; ou plus grande, comme dans le cas d’une plante cultivée en des régions où elle n’est pas spontanée). Il s’inscrit aussi dans un cadre temporel T ; on objective ainsi des faits e.scientifiques propres à certaines périodes historiques, la notion de désuétude… À partir de ces concepts, on peut représenter le fait e.scientifique global sous la forme symbolique
{E’ = aE, F’, A, T }.
Cette écriture est déjà en fait une abstraction, car elle résume une collection de faits individuels, concrets, reconnus dans un espace-temps A x T.
3.3. Discussion
Certains ethnologues (par exemple Sigaut, 1991) introduisent entre E’ et F’ le fonctionnement, c’est-à-dire l’opération par laquelle le mécanisme réalise sa fonction. F’ est alors l’ensemble complet des finalités exactes pour lesquelles E’ est mis en œuvre ; c’est ce qui le relie au système dont il n’est qu’un élément. Le rapport entre E’ et F’ est ainsi médiatisé par le fonctionnement : à quoi sert E’, comment il sert, pourquoi il est ainsi. Si on ignore le système, comment donner un sens à E’, remonter de E’ à F’ ? Par exemple, si le couteau sert en coupant, sa fonction F’ dépend du système ethnologique. Ces idées rejoignent celles concernant le lien de la forme à la fonction énoncées par Ledrut (1990 : 80), autrement dit le mode d’action.
3.4. Notion de chaîne opératoire
Il est rare qu’un seul a élémentaire intervienne dans un fait donné ; plus souvent plusieurs se combinent de façon ordonnée pour constituer une transformation plus complexe ; pour représenter celle-ci, on considérera le produit (normalement non commutatif) des opérateurs relationnels. Le cas d’une feuille alimentaire empruntée (e) à une plante, puis préparée (p) avant d’être cuite (c) pour être consommée offre la relation globale d’opérateur cpe. Ce processus technique, suite d’opérations élémentaires, est dénommé chaîne opératoire (CO) et va « du matériau brut jusqu’à l’objet fini » (M. Mauss) ; Maget (1953, cité par Pélegrin et al., 1988 : 57) parle de « chaînes de fabrication », découpées en « phases » puis en « gestes élémentaires ». Au cours d’une CO, les changements d’opération ou de technique scandent des étapes.
3.5. Conséquences sur les enquêtes
Pour réaliser l’étude assez complète d’un fait e.scientifique, il convient d’interroger les H sur les différentes composantes de la formule et fixer cette enquête dans l’espace-temps :
- connaissance des E ; par exemple, si l’enquêté donne des noms vernaculaires de P, tenter de les déterminer scientifiquement (à l’aide de flores, d’herbiers, de spécialistes) ;
- connaissance de E’ ; si c’est une forme complexe, en distinguer les parties, recueillir le nom et la fonction partielle de celles-ci ;
- connaissance des a : recueillir les modes de passage de E à la forme finale étudiée E’, préciser les CO ;
- connaissance des F’ ;
- si possible tenter de reconnaître son extension géographique A ;
- et si possible aussi tenter d’en reconstituer l’histoire (T).
4. Diversité des classifications des faits e.scientifiques
4.1. Principe d’une classification
Ayant posé plus haut la forme symbolique du fait e.scientifique global, une classification consistera à privilégier un des symboles X de la formule et à rassembler dans une même catégorie tous les faits caractérisés par le même X, invariant caractéristique de la catégorie. Selon le choix de ce X, on pourra donc distinguer plusieurs types de classification, chacun à valeur de monographie :
- X = E : monographie spécifique, consacrée à un être naturel ou à un groupe un peu plus large (exemple d’une monographie e.botanique des bambous ; Dupont & de Foucault, 1995) ;
- X = E’ : monographie ethnographique, consacrée à un fait e.scientifique, une forme précise, par exemple des barrières de prairie (de Foucault, 1991b ; cf. § 2) ;
- X = F’ : monographie fonctionnelle, très couramment et inconsciemment utilisée quand on s’intéresse aux plantes alimentaires, tinctoriales, médicinales (par exemple Arnold, 1985), à balai (de Foucault, 1997a ; cf. § 8), aux animaux de trait… ; notons en passant que ces F’ ont pu être à la source de nombreux qualificatifs de noms de plantes : edulis, esculentus, tinctoria, scoparia, officinalis… ;
- X = A : monographie régionale (par exemple Martin, 1971 ; Lormée et al., 2011 ; voir aussi § 2);
- X = T : monographie historique ; cas de l’e.botanique de l’Égypte ancienne.
4.2. Monographie ethnobotanique d’une région donnée
Dans le domaine de l’e.botanique, une monographie peut décrire l’ensemble des ressources e.botaniques offertes par une région donnée grâce à l’établissement de spectres. On illustrera cette approche dans le cas d’une petite région d’Espagne (d’après Benítez Cruz et al., 2010).
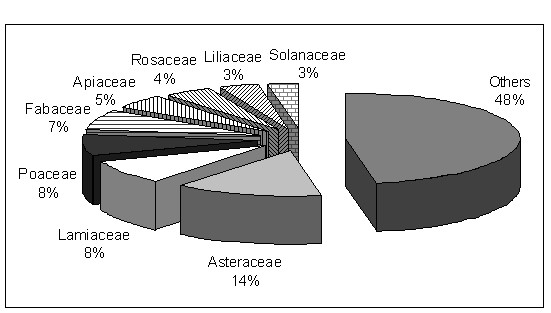
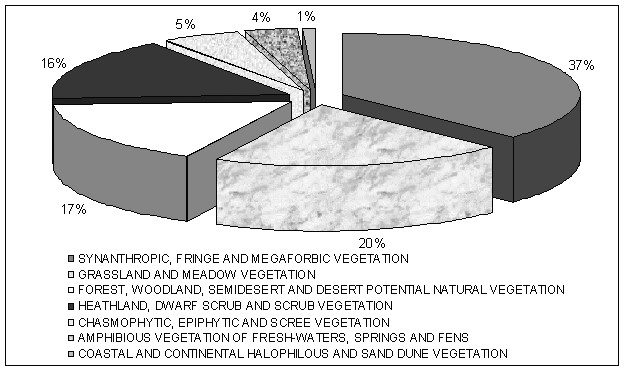
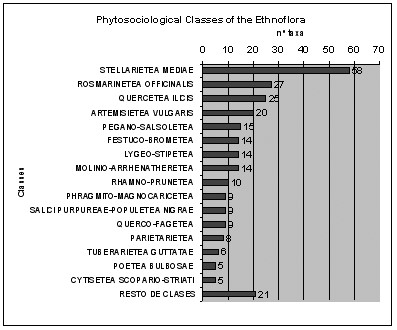
On peut d’abord répartir les plantes utilisées selon un spectre systématique (figure 1), mettant en évidence les familles d’importance ethnologique ; on peut encore rapporter ces plantes à la partie de la flore non utilisée dans les principales familles (figure 2). On peut aussi établir un spectre phytogéographique (figure 3), alors qu’un spectre des habitats et un spectre phytosociologique pourront mettre en évidence les habitats ou plus précisément les classes de végétation d’importance ethnologique (figures 4 et 5). On pourrait encore imaginer d’autres types de spectres, par exemple biologique…

5. La classification relationnelle
Dans la diversité des classifications possibles, il reste encore celle s’appuyant sur le cas X = a, qui paraît plus essentielle que les précédentes, car elle met en avant la nature même des e.sciences, la relation E-H. Elle revient à ranger dans une même catégorie les faits e.scientifiques offrant la même relation avec les H, donc des faits isomorphes au sens du structuralisme (cf. annexe 1). Les E incriminés dans des faits isomorphes sont alors homologues. Cette classification aboutira à une logique assez différente des précédentes, car elle écartera dans des catégories différentes des faits rapprochés par leur F’, par exemple les plantes alimentaires crues ou cuites, les clôtures végétales vives ou mortes.
5.1. Les relations e.scientifiques
La possibilité d’une telle classification tient au fait que les a possèdent un certain degré d’universalité, étant indépendantes des E sur lesquelles elles s’exercent et des H qui les mettent en œuvre. On peut dresser une liste des opérateurs élémentaires, sans distinguer celles qui ont les mêmes effets sur les E. Toutes ces relations ne s’appliquent pas obligatoirement à tous les E envisageables. Comme on l’a déjà précisé, les opérateurs a pourront souvent apparaître comme des produits de quelques-uns de ces opérateurs élémentaires.
- 0 : transformation nulle, qui n’agit pas sur E
- 1 : identité, qui laisse E inchangé (s’applique à P, Z, C, R, S, M, V, G)
- -1 : relation d’éloignement, répulsion (P, Z, C, S, M, V, G)
- b : brûlage, réduction en cendres (P, Z, C, R, S, V)
- c : cuisson (P, Z, C, R) (photo 5-1)
- d : dessiccation (P, Z, C, S, V)
- e : emprunt (P, Z, C, (R), S, V) ; si on souhaite préciser l’organe qui a été emprunté, on peut l’indiquer en indice, par exemple efeuilleP
- g : amélioration, par exemple génétique (P, Z, S, V)
- h : homothétie (P, Z, C, V, G)
- i : production organique (P, Z, (C)) (photo 5-2)
- m : mise à mort (P, Z)
- o : ornementation
- p : préparation par des transformations diverses (P, Z, C, R, V)
- r : rôtissage, torréfaction (P, Z)
- s : stylisation (P, Z, C, (R), M, V) (photo 5-3)
- t : travail sur E (P, Z, R, S, G)
- v : tressage, vannerie (P, Z) (photo 5-4)
- δ : déformation
- Σ : structuration (P, Z, C, R, S, V)
- Δ : déstructuration (P, Z, C, R, S) (photo 5-5)
- Λ : redressement d’un objet initialement couché, horizontal (R) (photo 5-6)






Dans cet essai, on ne détaille pas l’opérateur p qui cache en fait plusieurs transformations physico-chimiques. L’opérateur e doit être détaillé à part, car il cache deux faits e.scientifiques : l’un porte sur la partie utilisée E’1, l’autre (parfois inconscient) sur son complément E’2 dans E’ (avec E’ = E’1 ∪ E’2 ; ∪ réunion au sens de la théorie des ensembles). Ceci se passe par exemple lors de l’épluchage d’un légume, quand on garde la partie utile pour la consommation et qu’on délaisse ou élimine les parties retirées, à moins que, récupérées, elles ne viennent alimenter des animaux domestiques de l’Hortus (cf. § 15.3) ou un compost (photo 5-7).

On peut d’ores et déjà détailler quelques CO, par exemple la fabrication du vin de palme (pi ; photo 15-81), de l’huile d’olive (ie), du charbon de bois (be), la préparation de graines en guise de café (par exemple chez Cassia occidentalis, pre), l’empierrement pour un chemin pavé (ΣtR). Le cas des ruches traditionnelles du Queyras pour abriter des abeilles, fabriquées à partir d’un tronc de mélèze (Larix decidua) et d’une lauze de schiste lustré, correspond à la chaîne Σ[tdeLarix, 1Schiste] (photo 5-8) ; dans les Cévennes, le mélèze est remplacé par le châtaignier (Castanea sativa), le schiste lustré par un autre type de schiste (photo 5-9). La ruche picarde (photo 5-10) renvoie à la formule Σ[vdeOsier, Σ(iVache, 1Argile)], décrivant un cas où le corps de la ruche, fait de rameaux d’osier tressés, est étanchéifié par un mélange d’argile et de bouse de vache. Les abeilles elles-mêmes étanchéifient leur ruche avec de la propolis, gomme qu’elles récoltent sur les bourgeons de divers arbres (nouveau cas de relation zoo-botanique).



Dans les développements à venir, on sera peut-être amené à introduire d’autres opérateurs, moins fréquents que les précédents. On peut notamment penser à la domestication (D), qui s’applique surtout aux P et Z, chez lesquels H induit des modifications morphologiques, physiologiques, éthologiques… pour des finalités diverses (production, énergie…). Ainsi l’histoire du maïs (Zea mays) montre l’évolution de la taille de l’infrutescence au cours du temps à partir des maïs primitifs (photo 5-11). Mais on peut l’étendre aussi à l’e.minéralogie à travers l’eau, roche à l’état liquide à la température ambiante. En effet, celle-ci peut être captée puis acheminée par des conduites forcées, des canalisations, éventuellement mise en réserve dans des barrages où elle acquiert de l’énergie potentielle. La domestication touche aussi le feu attaché à la maison, la domus.

La courte CO tG décrit les sites exceptionnels que sont Petra (Jordanie), Lalibela (Éthiopie), les églises troglodytiques de Cappadoce (Turquie ; photo 5-12), la nécropole de Darius à Persépolis (Iran), les mines néolithiques de Spiennes (photo 5-13) ; les bouddhas de Bâmiyân (Afghanistan) en relevaient aussi, mais ils ont été détruits à l’explosif par les talibans en mars 2001. Le déplacement du monument d’Abou Simbel (Égypte) suite à la construction du barrage d’Assouan a donné lieu à une déstructuration fortement encadrée suivie de sa reconstruction exacte quelques dizaines de mètres au-dessus, soit le produit ΣΔ = 1 ; un cas très semblable concerne le gedige de Nalanda à Sri Lanka (photo 5-14). Le cas d’Uluru (photo 5-15) est à la limite e.minéralogie/e.géomorphologie selon qu’on le considère comme un monolithe (alors un des plus massifs au monde) ou un relief.
L’érection des mo’ai (têtes monumentales) de Rapu Nui (Chili, île de Pâques ou Easter Island) a fait appel à des matériaux volcaniques, surtout issus du volcan Rano Raraku ; détachés, déplacés et rapprochés sur une aire cérémonielle appelée ahu, les mo’ai ont enfin été redressés : ΛΣ(tG).




5.2. Notion de relation indirecte
Il arrive que H utilise un produit E’ non délibérément fabriqué par lui ; il existe donc une relation indirecte entre H et E ; elle sera représentée sous la forme |a|. Un premier exemple est celui de la récolte de bois sec qui peut être collecté tel quel sans avoir été séché volontairement par H, soit |de|P (ici a est le produit de deux relations indirectes, emprunt e puis dessiccation d) ; l’utilisation de brindilles pour baliser des pistes peut être décrite comme Σ|de|P, la structuration Σ étant bien assurée directement par H.
Ce concept sera aussi utile pour décrire certaines chaînes commerciales, comme dans le cas du bouquet de muguet acheté associant un fleuriste à un client, ce dernier n’ayant qu’une relation indirecte aux fleurs achetées :
{ΣeP, rapporter financièrement} → {|Se|P, orner ou porter bonheur} →(fanaison)→ {|Σe|P, 0}
La fanaison étant une évolution normale de la fleur coupée, elle n’est pas de nature e.scientifique ; par contre elle influence la fonction secondaire.
De telles relations entrent aussi en jeu dans le cas des fossiles végétaux et des paléobotanistes qui les étudient. Ici, la transformation est le produit d’un emprunt e (P n’est jamais intégralement conservée) indirect par une fossilisation. Cette dernière est une transformation aussi indirecte qui ne laisse invariantes que les relations organiques, guère les éléments eux-mêmes ; en cela elle est très proche de la stylisation s, soit la relation e.botanique globale proposée sous la forme {|se|P, être étudié ou témoigner d’une flore disparue}.
L’épizoochorie des semences par H peut être décrite de manière analogue sous la forme {|e|P, 0}. En haute Tinée (Association des Stéphanois, 1987), H recueille les brindilles de mélèze (Larix decidua) accumulées par les fourmis pour en faire de la litière : {|Σ|e||P, pailler} ; on notera que l’emprunt est déjà indirect du mélèze aux fourmis.
Le « buc » est une ruche des Pyrénées audoises (plateau de Sault) réalisée à partir d’une lauze et d’un tronc de sapin creux suite à une pourriture de cœur due à l’action d’un champignon : Σ[e|t|Abies alba, 1R], |t| étant assuré par un C (photo 5-16).

5.3. Les autorelations
Si on considère le comportement de l’araignée sécrétant un produit pour tisser sa toile (ici Σ plutôt que v) ou la sécrétion du ver à soie construisant un cocon, ils font apparaître i sous la forme d’une autorelation que l’on peut noter i↑. Un exemple voisin se présente avec la salive de certaines femmes utilisée pour faire fermenter un glucide (yucca en Amazonie péruvienne pour préparer le masato ; manioc en Amazonie guyanaise pour préparer le cachiri ; photo 5-17).
Les autorelations jouent aussi un rôle important dans les rapports entre H et son propre corps (Chippaux, 1990) : amputations diverses (dont l’autocastration : e↑), scarifications (t↑), déformations d’organes (δ↑), voire même le suicide (m↑). Dans des conditions extrêmes, une personne peut être amenée à boire sa propre urine pour lutter contre la déshydratation (i↑). Parfois l’autorelation passe par un acteur extérieur, elle n’est alors qu’indirecte : |a↑|.

5.4. Opérateurs et outils
Les opérateurs décrivent des transformations de E en E’ ; elles peuvent être remplies par la main humaine (e…) ou plus souvent par des techniques, dont des outils, qui apparaissent alors comme des relais entre H et la matière (Leroi-Gourhan, 1992b), par exemple la hache pour e et m, le métier à tisser pour v, du matériel et des produits diversifiés pour p, des liens variés pour Σ. On déplore en passant l’abus actuel du terme de technologie comme synonyme de technique sophistiquée, alors que ce mot désigne bien une science (radical –logie), celle des techniques (Haudricourt, 1987).
On voit aussi incidemment qu’un fait e.scientifique de la forme {aE1, F’} peut donner lieu à un second de la forme {a’E2, F’ = assurer a}. Quelques exemples : repellent d’origine végétale (Melia azedarach, M.a.) pour insectes
{-1insectes, 0} et {peM.a., F’ = assurer -1} ; séchage du café ou de briques d’argile {deCoffea ou dtArgile, F’} et {1Soleil, F’ = assurer d} (photos 5-18 et 5-19), ces deux derniers exemples avec deux faits de nature e.astronomique ; pour un arbre P ombrageant protégeant du soleil, on peut écrire la forme suivante : {1P, -1Soleil}.


6. Quelques premières illustrations
On va illustrer ici quelques premiers faits du type {a, F’}. La classe {0, 0}, en quelque sorte le zéro des e.sciences, rassemble des faits où les E n’ont aucune relation avec H, par exemple des P et Z sauvages et/ou inférieurs, habituellement ignorés de H.
D’après ce qui a été dit plus haut (à savoir qu’il n’y aurait pas de fonction sans structure matérielle), la classe {0, F’} doit être vide (cf. cependant en mythologie, § 14).
La classe {a, 0} est possible avec a = -1 (E = Hippomane mancinella, le Mancenillier ; photo 6), a = m (E = nuisibles), a = e (anthropochorie ; épluchage, photo 5-7).
La classe {1, F’} est extrêmement riche : P supports (F’ = porter), E étudiés par les chercheurs, E = signifiants U (alors F’ = signifier)…
La classe {1, F’ = F}, autrement dit l’utilisation de E non modifié pour un F’ identique à sa fonction primaire, peut être illustrée par les plantes d’aquarium utilisées pour oxygéner l’eau, abriter des poissons ; autre exemple : utilisation volontaire de dattiers mâles pour féconder les dattiers femelles.
La classe {a, F’ = F} porte sur des E modifiés pour une F’ identique à leur fonction primaire. Si E est un être vivant suffisamment complexe pour être formé d’organes eux-mêmes dotés d’une fonction primaire F1, il est possible d’emprunter (opérateur e) un de ces organes pour l’utiliser de la même manière que dans E. Par exemple, si la peau animale protège les organes internes, elle peut aussi être utilisée après la mort de l’animal (pdmZ) pour réaliser un vêtement à usage humain et encore à fonction de protection ; en relève en outre l’utilisation par les Inuits de la graisse de phoque pour lutter contre le froid : {pem, F’ = F}.
7. Un premier cas concret : la cabana du Limousin
Un exemple d’étude e.botanique assez complète a porté sur les cabanes végétales du feuillardier limousin (de Foucault, 1994a). Dans cette région, celui-ci exploite des taillis de châtaignier (P = Castanea sativa, Fagaceae ; photo 7-1) pour fabriquer des feuillards destinés principalement à cercler les tonneaux pour vins et liqueurs (surtout de Bordeaux, Bourgogne et Cognac) et s’abrite dans une cabane.

Il y a d’abord une composante structurelle. Lors de la fabrication de l’abri forestier, interviennent plusieurs étapes mettant en jeu des parties possédant chacune une fonction partielle, par opposition à la fonction globale de l’objet final :
- jeunes pousses de P courbées et entrecroisées en vue de façonner des arceaux qui porteront des barres : {vP, porter 4 barres horizontales) (photo 7-2) ;
- quatre barres porteront les branches {eP} ; elles sont fixées aux arceaux par des liens {veP} (photo 7-3) et parfois consolidées par des fourches (photo 7-4) ;
- les branches soutiendront la couverture, elles ont valeur de chevrons ;
- la couverture est réalisée progressivement à partir des résidus laissés par le travail, des copeaux {e2P} accumulés en tas (photo 7-5) ; ils sont posés sur les chevrons grâce à une fourche de bois {teP} (photo 7-6).





La fonction de la cabana finale (photo 7-6) ne se réduit pas à la somme des F’ partielles précédentes, il y a émergence d’une fonction globale : abriter temporairement le feuillardier en cas de mauvais temps, protéger ses outils, entreposer les feuillards finis.
L’extension spatiale (variable A de la formule générale) de ce fait e.botanique correspond actuellement à la partie sud du département de la Haute-Vienne, mais devait jadis s’étendre sur le Périgord (Sarladais notamment), le Nontronnais, la Charente limousine et peut-être même l’Auvergne. L’extension temporelle (T) fait apparaître que ce métier serait apparu vers 1770, aurait eu son optimum entre 1880 et 1930 ; aujourd’hui il s’éteint progressivement. Souvent, maintenant, les feuillards sont préparés dans les fermes, de sorte que la cabana disparaît plus vite que la profession elle-même.
D’autres cabanes végétales existaient ou existent encore en France, associées à de petits métiers du bois ou de la forêt ; citons le carcahoux des bûcherons de Haute-Normandie (Sannier, 1985 ; de Foucault & Legrand, 1994 ; photo 7-7), les loges des cercliers de Charente (de Foucault, 1994a ; photo 7-8), les cabanes de sabotiers du Nord-Est et de Wallonie (où elles sont dénommées baraques ; Calozet, 1945). En République centrafricaine, on peut rencontrer des cabanes de chasseurs pygmées (photo 7-9). On illustre ainsi l’ethnovicariance, lorsque des objets de même fonction mais de structure différente se remplacent selon les régions géographiques.



La dimension temporelle se manifeste encore dans les modifications de l’objet global et/ou de ses parties. De telles monographies doivent donc aussi intégrer des investigations d’ordre dynamique. On peut les classer en considérant divers invariants à travers ces changements :
- à E’ invariant : la cabana étant en voie de disparition, pour conserver des traces de ce patrimoine ethnologique, on en remonte comme témoignage de ce mode de vie et de cette profession en déclin ; c’est donc une dynamique à E’ invariant : {E’, abriter} → {E’, témoigner} (photo 7-7) ;
- à F’ invariant : il existe aussi des dynamiques de la forme {E’1, F’} → {E’2, F’} ; par exemple les liens végétaux (photo 7-3) peuvent être remplacés par du fil de fer, plus facile à utiliser ; les copeaux de la couverture (photo 7-6) sont progressivement substitués en tôles ou feuilles de plastique plus durables.
Mais il existe aussi des cas de dynamique sans une telle invariance ; ainsi les matériaux végétaux d’une cabana concrète délaissée après la saison de travail ont pu être récupérés pour allumer le feu familial durant l’hiver.
8. Un second cas concret : les balayettes et balais traditionnels
Le formalisme présenté ici a servi aussi pour décrire et classer des objets de la vie courante que sont les balais et balayettes (de Foucault, 1997a), dont certains sont encore fabriqués à partir d’éléments naturels, végétaux, mais aussi parfois animaux. Au-delà de leur grande diversité de formes, ils possèdent le plus souvent une même fonction, celle de réunir en un volume réduit des éléments dispersés sur une surface plus ou moins vaste, des éléments inutiles dépourvus de fonction (poussière, déchets par exemple) ou utiles qu’il serait fastidieux de réunir un à un (des grains de blé par exemple). Parfois, au contraire, surtout les balayettes, ils permettent d’écarter les poussières d’un objet fragile (époussetage). On peut aisément distinguer ces deux types d’objet, les balais sont de plus grande taille, l’usager étant debout à légèrement courbé ; la balayette est de plus petite taille, forçant l’utilisateur à être fortement incliné s’il œuvre sur le sol, cet objet étant alors mieux adapté au travail en hauteur.
L’étude de leur architecture amène à les ordonner selon une complexification croissante, mettant en jeu des CO de plus en plus élaborées. Dans la suite ‘b.’ représentera indifféremment un balai ou une balayette. Les illustrations sont extraites de la monographie citée.
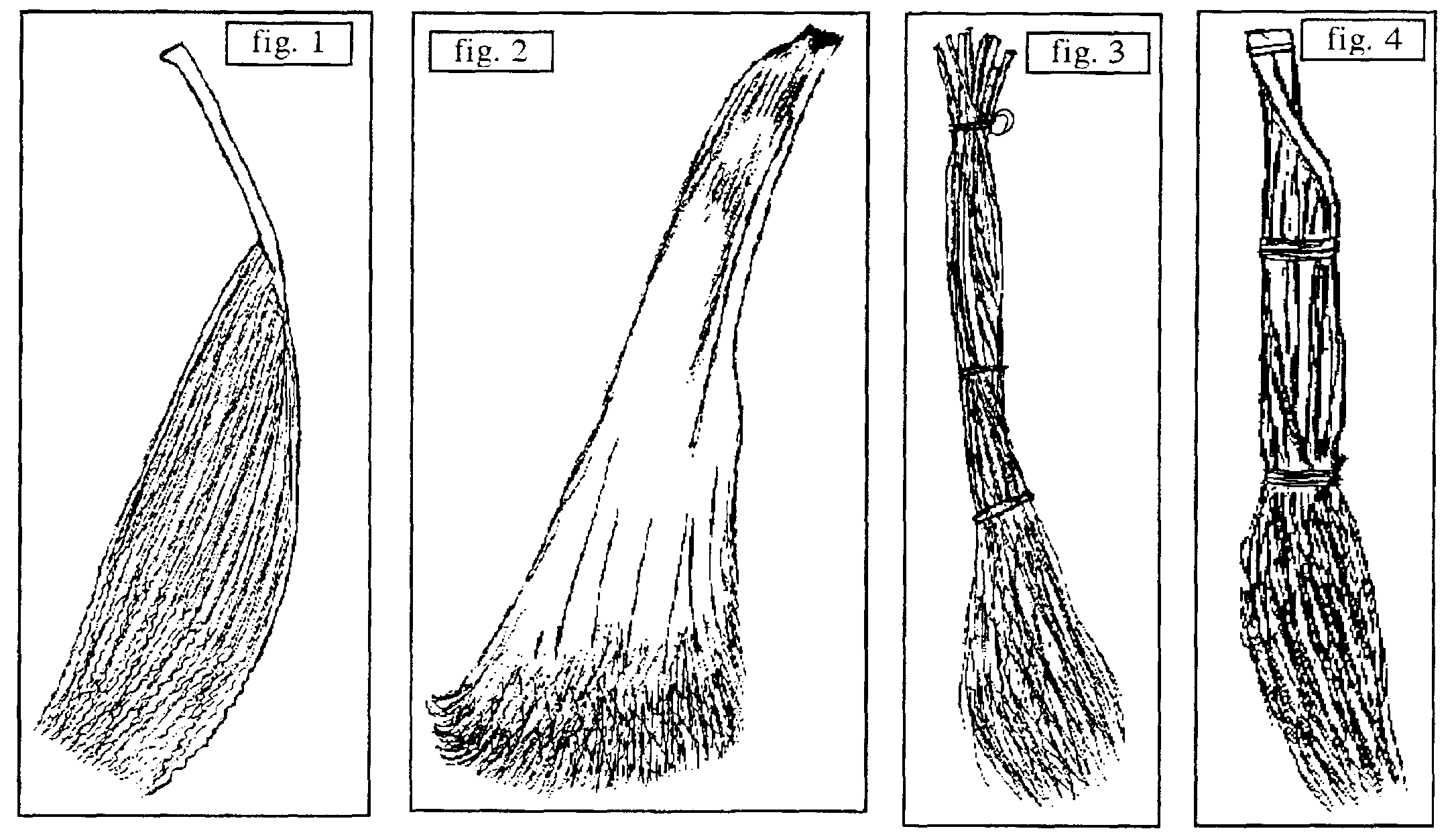
A. b. élaborés sans structuration, étant simplement constitués d’un élément végétal unique suffisamment volumineux pour remplir sa fonction (eP) : b. à base d’inflorescence femelle (spadice) de Phoenix dactylifera (figure 6) et b. faite de la base d’un pétiole de feuille du même dattier (figure 7), deux objets recueillis en Égypte du Nord par J.-M. et J. Géhu en 1986, le premier ayant aussi été observé au Maroc (Ourzazate, avril 1992) et en Tunisie méridionale (Chebika, Tozeur, avril 1993). Un b. intermédiaire entre cette catégorie A et la suivante a été observé à Tamegroute (Maroc, avril 1992) : deux fragments de feuilles de Ph. dactylifera le constituent mais ne sont structurés ensemble que lors de l’usage, lorsque la main humaine les prend et les réunit le temps de s’en servir.
B. b. élaborés avec structuration Σ à partir d’éléments séparés, Σ étant assuré grâce à des liens de nature diverse, traditionnellement végétaux.
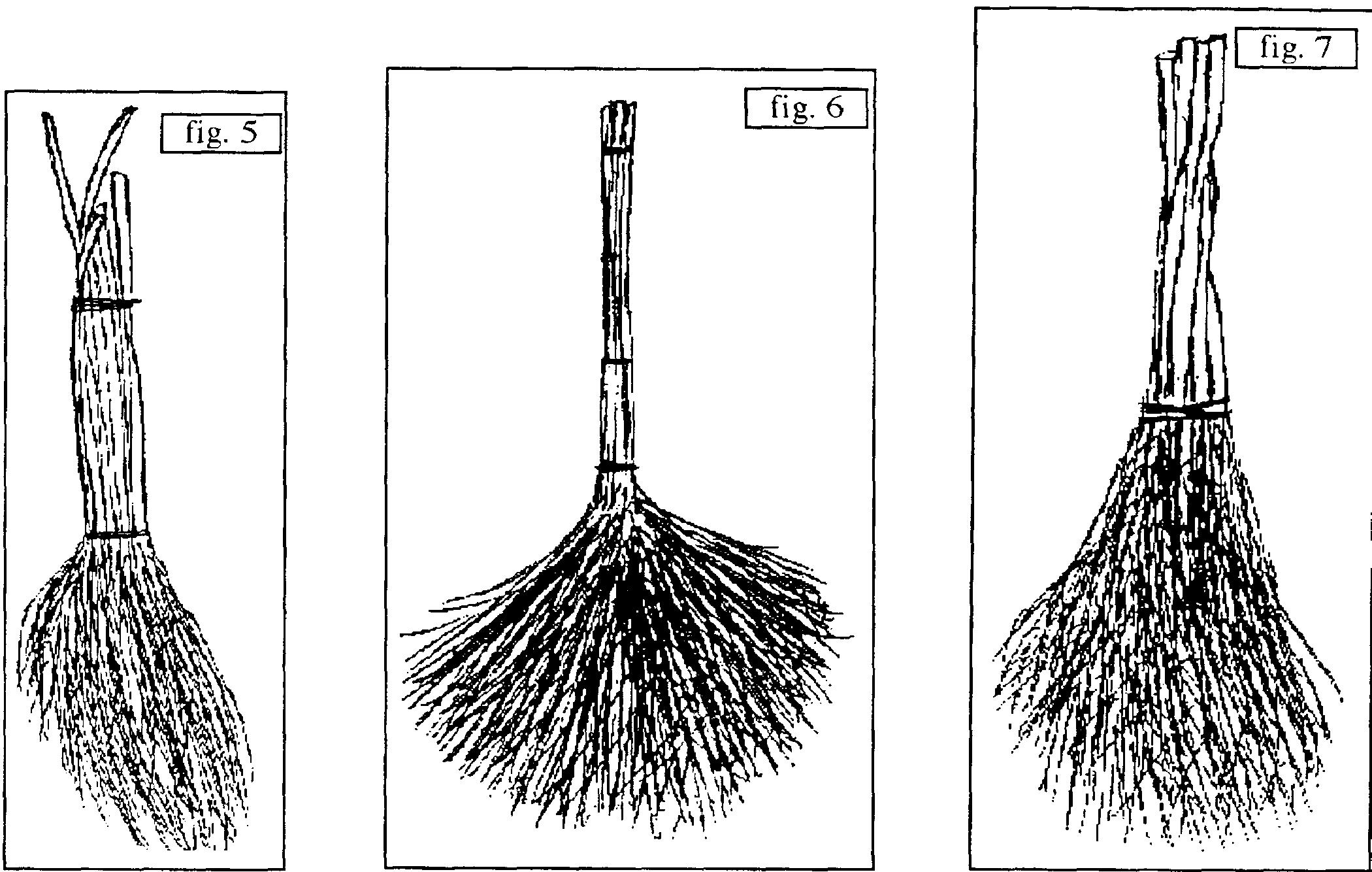
- B.1. sans manche : Σe ou ΣdeP : b. d’Amelanchier ovalis du Gard (figure 8 ; Durand-Tullou, 1972) pour nettoyer bergeries et aires de battage (Σ est apparemment assuré par du fil de fer) ; b. de Cytisus scoparius du sud du Massif central (figure 9 : Montroc, Tarn ; figure 10 : Douch, Hérault) (Σ est assuré par de la ficelle et/ou du fil de fer) ; certains de ces b. furent utilisés pour sortir les cendres des fours et balayer la sole sur laquelle étaient placées les tourtes dans le Massif central (Lapeyre & Dodinet, 1982) ; b. de Betula pendula structuré au moyen de fil de fer (figure 11, Beynat, « Espagnagol », Corrèze, janvier 1991) ; b. de Spartium junceum ou de Genista cinerea (Rémuzat, Drôme, août 1994, resp. dits localement « grand genêt » et « petit genêt »), structurés par des cordelettes (il faudrait comparer ceux à G. cinerea avec les b. faits du même matériau signalés en Haute-Tinée ; Association des Stéphanois, 1987) ; b. de Cytisus oromediterraneus (figure 12, Douch, Hérault, printemps 1987) actuellement structuré par du fil de fer ; b. de chaume de riz (Oryza sativa) observé à Sade (environ de Kuta, Lombok, Indonésie, août 1993) pour le ménage de l’ethnie Sasak, structurée par un lien végétal .
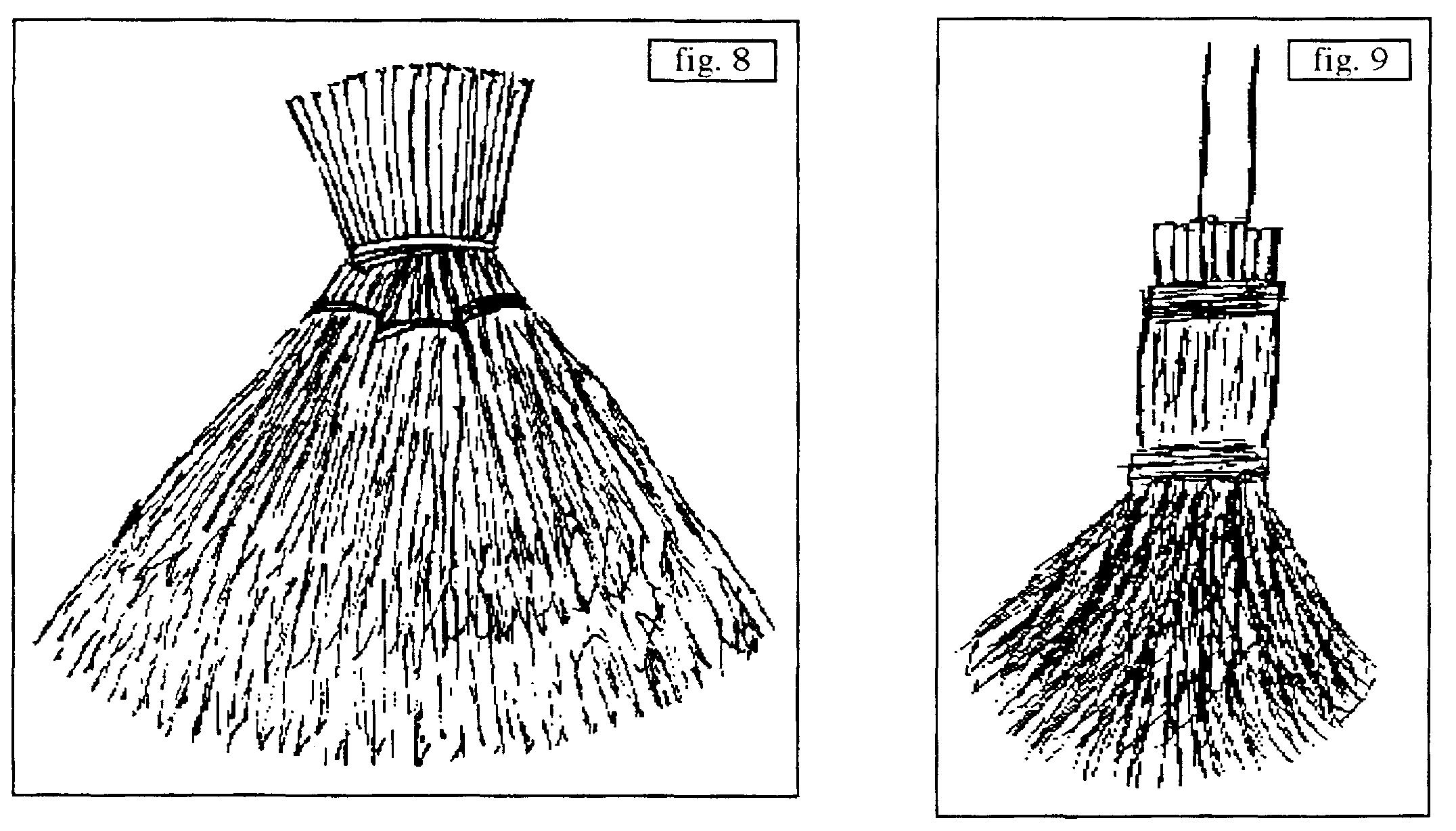
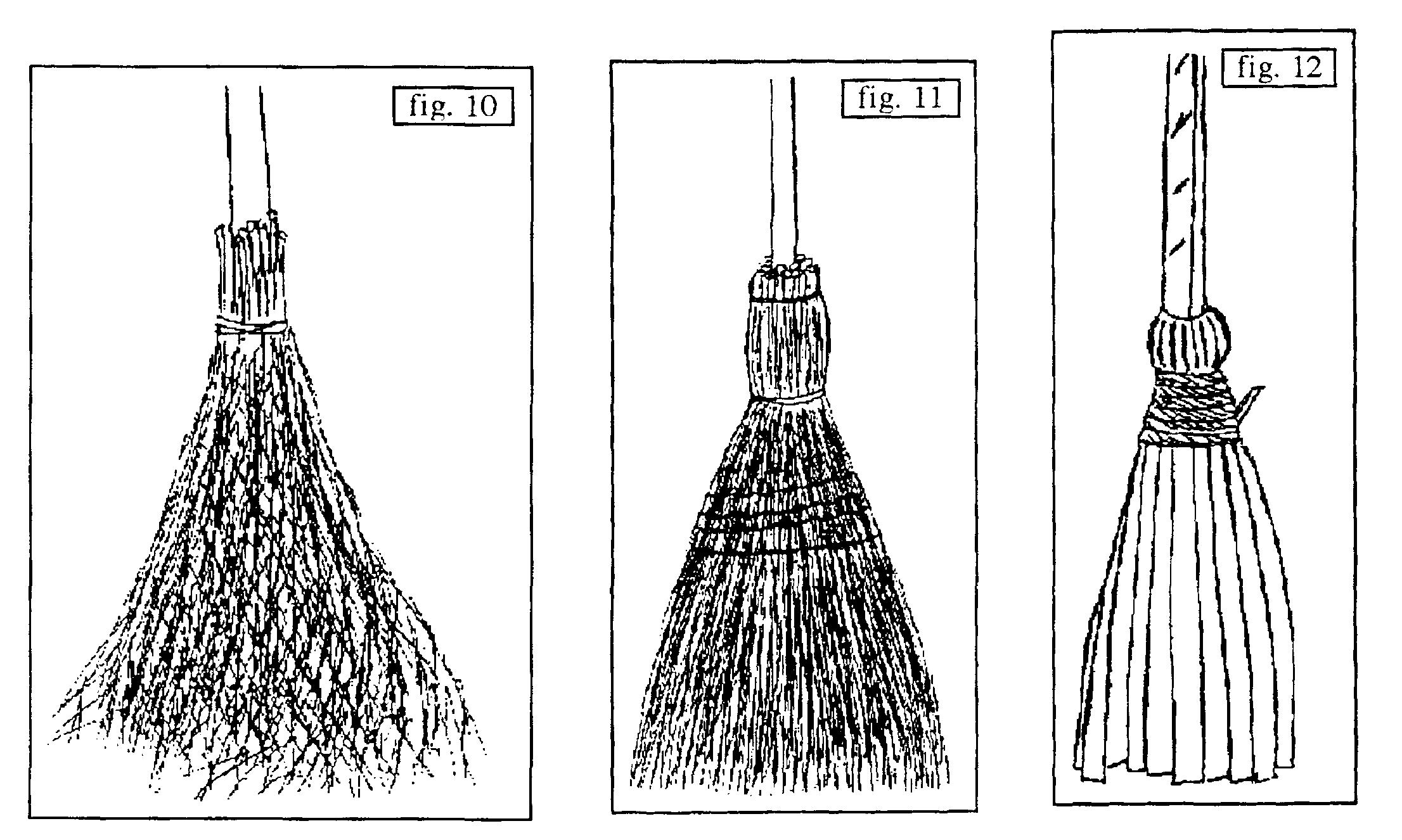
- B.2. [avec manche : un manche externe est ajouté, Σ[(e ou deP1), a P2], P2 désignant la plante utilisée pour élaborer ce manche ; b. de Calluna vulgaris (figure 14, Montroc, Tarn, structuré grâce à une ficelle) ; b. de Betula pendula (figure 15, Le Fuilet, Maine-et-Loire) structuré par du fil de fer ; ce b. est ou était l’un des plus fréquents en Europe du Nord, signalé en Picardie (Valcke, 1989), en Creuse, réalisé en grand en Corrèze (Beynat, « Espagnagol » pour les services de la voirie de Brive-la-Gaillarde en 1991) ; Brosse (1989 : 47) l’évoque pour la Russie : « Selon les proverbes russes […] le bouleau […] nettoie […] ; de ses branches, on fait des torches, car elles donnent de grandes flammes claires, ainsi que des balais. » ; en Morvan, il est réservé aux étables et cours, alors qu’un b. de Cytisus scoparius, plus souple, le remplace pour l’intérieur des maisons (Crosnier, 1990) ; b. d’Amelanchier ovalis proche d’un précédent cité en B.1 mais réservé aux dalles et à la terre battue (Durand-Tullou, 1972) ; b. de Prunus spinosa ou de Crataegus monogyna selon des informateurs de Beynat (Corrèze, janvier 1991) pour balayer les feuilles mortes des prés en hiver (usage aujourd’hui désuet), le manche était à base d’Ilex aquifolium, Castanea sativa ou Corylus avellana ; divers b. de Symphoricarpos albus en Morvan, de Sorghum halepense (figure 16) en Bresse… (Crosnier, 1990) ; b. de Phoenix canariensis en Grande-Canarie (figure 17) : ici le corps du b. et le lien structurant sont empruntés au limbe, le manche, entré en force, est extrait du rachis d’une feuille ; b. de Cocos nucifera avec manche en bois varié selon les individus, pour balayer les sols humides en Inde ; b. d’Arenga pinnata (palmier à sucre), manche de bambou et lien végétal (Tanah Lot, Bali, août 1993).
D’autres cas, illustrés ou non, sont à retrouver dans la publication de 1997. Présentons encore le jharoo, b. d’Inde du Nord utilisé pour la poussière sèche et réalisé à partir de l’inflorescence d’une grande graminée, probablement Thysanolaena maxima, structurée au moyen d’une cordelette de Corchorus capsularis (photo 8).
Suite à l’analyse de cette diversité de b. végétaux, ajoutons qu’il existe aussi des balayettes élaborées à partir d’éléments d’origine animale, des « plumeaux ». L’un des plus frustes est certainement celui que j’ai connu en Basse-Normandie dans les années 1950, élaboré à partir d’une aile d’oie désarticulée au niveau du coude, puis séchée, soit la chaîne opératoire e.zoologique demOie, qui relève de la catégorie A précédente, plumeau très efficace pour nettoyer les coins d’escalier. Plus élaboré est le plumeau réalisé à partir de plumes, dont l’ensemble est pourvu d’un court manche (catégorie B.2), soit Σ(e ou emZ, aP), pratique pour nettoyer les objets délicats dans les magasins, les musées…

9. Transformations fonctionnelles
On a vu les nombreuses transformations structurelles E → E’ ; on peut se demander maintenant si on peut raisonner de la même manière pour le passage F → F’ et alors écrire F’ = bF. En dehors de b = 1 (soit F = F’) déjà évoqué précédemment, il semble que ces transformations fonctionnelles soient plus rares ou alors plus difficiles à mettre en évidence. On peut pourtant penser d’abord à la consécration ou la sacralisation (bénédiction…) qui dote E’ d’un signifié sacré, sans véritable modification structurelle ; ce n’est qu’au regard des H du groupe que ce signifié apparaît. Quelquefois la modification structurelle existe, mais est très faible : bandes colorées (sur Couroupita, Ficus chez les bouddhistes ; opérateur o ; photo 9-1). La sacralisation peut mettre en jeu un fait e.botanique utilisant par exemple l’encens (issu d’un Boswellia), le khul et le henné pour les moutons de l’Ayd El Kébir. Les « arbres à esprits » d’Afrique noire sont aussi sacralisés après un rituel de consécration (Bonnet, 1990 : 705 ; voir aussi ci-après) et ce caractère peut être discrètement marqué par des cordons d’herbes aromatiques, des cercles de plumes de volatiles sacrifiés (emZ) lors du rituel, des ornementations colorées (o) sur le tronc (Bonnet, 1990 : 704).

Dans la continuité de cette idée, on en déduit inversement la désacralisation (ou laïcisation) et la conversion. Dans nos régions ouest-européennes, la conversion d’un arbre initialement dédié à un culte païen peut passer par la simple installation sur son tronc d’un petit oratoire (photo 9-2) : « Il en est des bois sacrés comme des gentils : on n’extermine pas ces derniers, on les convertit, on les change. De même, on ne coupe pas les bois sacrés, on fait mieux : on les consacre à Jésus-Christ. » (saint Augustin). L’histoire de la basilique Sainte-Sophie d’Istanbul (photo 9-3) illustre un cas de dynamique où interviennent successivement ces transformations fonctionnelles (de Foucault, 1993, actualisé). À Sainte-Sophie, la conversion du ive siècle a été matérialisée par l’installation de minarets, du miqrāb (ou mihrab, indiquant la direction de la Mecque), du minbar (chaire pour l’imam), de vastes calligraphies de versets du Coran…


On peut ensuite penser au rituel, défini comme un ensemble de rites : là encore E’ n’est pas modifié, pourtant la signification de la forme ritualisée est complètement modifiée (de Foucault & Claisse, 1999 ; Claisse & de Foucault, 1999a). Un rite est donc sous-tendu d’efficacité, de signification, et le concret du rituel fait apparaître diverses pratiques :
- actives ou positives :
- l’actant lui-même est mis en condition physique et préparé ;
- le rite est fixé dans l’espace (lieu, orientation, approche de E…) et le temps (période, position selon la Lune ou le Soleil… ; e.astronomie) ;
- incantations magiques, prières… ;
- interventions de « fluides » divers : eau, vapeur, magnétisme… ;
- répétition de règles stéréotypées, de gestes donnant une efficacité opérante ;
- négatives :
- interdits ;
- secret, marginalité.
Mauss (1947/1989 : 237) reconnaît aussi l’existence de rites positifs et négatifs, ainsi que (en p. 240) des rites oraux. Des rituels marocains permettent par exemple de doter la nourriture d’un caractère sacré dans le cas de situations dans lesquelles une partie des nourritures est détournée pour devenir remède grâce à un rituel dotant le repas d’un attribut sacrificiel, l’autre partie étant partagée entre humains ou entre humains et génies (Diouri, 1994 ; Claisse & de Foucault, 1999a). Un rapprochement avec la communion de confession catholique est instructif : le repas sacrificiel rapproche des convives consommant ensemble une victime qui s’est elle-même offerte ou immolée, le Christ.
10. Dynamique des faits e.scientifiques
10.1. Généralités
L’exemple de la cabana nous a permis d’illustrer quelques points de dynamique, que l’on peut maintenant chercher à généraliser. En s’appuyant à nouveau sur la formulation simplifiée {E’ = aE, F’}, on peut illustrer l’évolution de faits en considérant T comme variable pour décrire des séries temporelles (de Foucault, 1990b), l’ethnovicariance ayant montré un exemple de série spatiale.
Il existe d’abord des séries à un paramètre variant :
- à E ou E’ invariant ; revient à l’étude de la variation de F’ pour un même objet ; pour ce cas, Poirier (1990 : 957) évoque ce qu’il appelle une loi de transposition fonctionnelle : « un objet est utilisé pour un autre usage que celui originellement prévu» ;
- à F’ invariant ; revient à l’étude de la variation de E’ pour une même fonction ;
- F’1 → F’2; par exemple l’encens issu de Burseraceae était, semble-t-il, initialement utilisé pour ôter les mauvaises odeurs dues aux sacrifices animaux dans les lieux de culte ; après leur abolition, il est devenu rituel ;
- 0 → F’2, cas de la récupération d’une forme de F’ nul pour une fonction non nulle ; ainsi des mauvaises herbes {1P, 0} sont ultérieurement devenues des céréales utiles {aP, nourrir} ; récupération de déchets ménagers pour un compost (photo 5-7) ;
- F’1 → 0, cas d’un E’ achevant son rôle (fleurs fanées…), de désuétude… ;
- {-1P, 0} → {mP, 0} : une plante dangereuse dont on s’écarte peut être finalement détruite pour éviter tout incident ou accident (cas de l’éradication du mancenillier, Hippomane mancinella, des plages à touristes aux Antilles françaises ; photo 6) ;
- les notions de succédané et d’ersatz apparaissent lorsque E’ initial devient rare ou manque et est remplacé par un autre E’ souvent de moindre qualité ; ainsi les caryopses d’orge torréfiés ont pu remplacer le café en période de pénurie, des plantes diverses peuvent pour la même raison remplacer le tabac (Cuénot, 1967, bien plus qu’une « note préliminaire » en dépit du titre), d’autres ont remplacé diverses matières premières (textiles, colorants, caoutchouc…) en période de restriction lors de conflits armés (Decary, 1968).
Il existe aussi des séries sans paramètre : la notion de falsification peut par exemple apparaître lorsque E’ et F’ varient simultanément : {aP1, F’1} → {a’P2, F’2} ; elle touche notamment des plantes médicinales (Adonis vernalis falsifié par d’autres Adonis…), des épices (stigmates de Crocus sativus falsifiés par des fleurons de Carthamus tinctorius).
La monographie déjà citée des balais et balayettes (cf. § 8) a aussi donné lieu à de telles études dynamiques, en recherchant les invariants que laissent ces modifications.

10.2. Cas des balais et balayettes
À F’ invariant
On peut mettre en évidence le remplacement dans le temps de certaines formes de b. (balais, balayettes) par d’autres. On pense surtout d’emblée aux b. végétaux remplacés par des b. en matière synthétique. Le grand dommage de cette dynamique structurelle est une banalisation : aux divers b. traditionnels d’aire géographique plus ou moins limitée, réalisés avec du matériel local, se substitue un b. pratiquement unique sur une grande partie de notre planète (de Foucault, 1995a) ; par exemple aux élégants b. thaïlandais (photo 10) se substituent des b. que l’on retrouve à l’identique en Europe.

Des parties de b. peuvent aussi présenter une telle dynamique, essentiellement les liens qui structurent la plupart des b. étudiés, avec l’invariant « F’ = assurer Σ » : cette dynamique remplace les liens végétaux (genêt ou osier tressés) par du fil de fer, autre cas de banalisation. Par ailleurs ce passage temporel d’un lien végétal au fil de fer se déroule aussi dans l’espace, entre les ethnies restées plus ou moins traditionnelles (Canaries, Thaïlande, Malaisie, Indonésie, Inde) et les pays plus avancés (France) ; on est là en présence d’un exemple d’isomorphisme spatio-temporel, de symétrie entre espace et temps (de Foucault, 2014a).
À P’ invariant
Les b. peuvent aussi présenter une dynamique à structure invariante et fonction secondaire variante. Ainsi, à l’origine, ils étaient de fonction rituelle. Brosse (1989) précise que, dans le rituel d’initiation, un candidat chaman sibérien est purifié grâce un b. fait de rameaux de Betula, trempé dans une marmite contenant un mélange bouilli de Thymus, Juniperus communis et écorce de Pinus. Toujours selon ce même auteur, sa fonction fut aussi magique : le balayage était un acte cultuel, pour purifier le sol des souillures externes, profanes ; cet acte magique serait encore utilisé chez les Berbères de Kabylie. Rappelons-nous par ailleurs du b. magique de L’Apprenti sorcier (nouvelle de Goethe mise en musique par P. Dukas puis en film animé par Walt Disney dans Fantasia). Cette dynamique reliant ces actes cultuels à l’acte profane est une désacralisation comme évoqué au § 9.
On pourrait prendre encore l’exemple des chaînes commerciales, qui associent un vendeur, pour qui le b. possède une fonction de rapport pécunier, à un acheteur, pour qui celle-ci est d’ordre ménager.
En annexe 2, on trouvera un autre cas de dynamique de balai faisant intervenir une relation indirecte.
11. Interrelations entre e.sciences
Nous avons vu incidemment que deux e.sciences ou plus peuvent intervenir dans un fait donné, ce que nous allons détailler et illustrer à nouveau.
11.1. Les galles
Une galle est le résultat d’une piqûre (nouvel opérateur k) d’un P par un insecte Z ; la fonction primaire F est la reproduction de Z. D’un autre côté, le résultat kP ou akP peut acquérir pour H une fonction secondaire F’ : colorer (galle de Pistacia terebinthus), tanner (galles de Quercus, Tamarix), écrire (galle de Quercus transformée en encre). Un cas assez particulier concerne la manne : kTamarix nilotica par Trabutina manipara induit une exsudation i sucrée qui, desséchée, fournit ce produit alimentaire : {d|ik|P, nourrir}, k étant assuré par Trabutina.
11.2. Quelques élevages
Plusieurs interrelations peuvent être décrites par la structure relationnelle suivante :
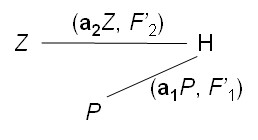
selon les cas suivants
- Z = ver à soie malgache (chenille du Lépidoptère Borocera cajani), P = Tapia (Uapaca bojeri, Phyllanthaceae arborescente des hauts plateaux malgaches), avec a1 = 1, a2 = vpi, F’1 = nourrir Z, F’2 = orner, vêtir (Verheggen et al., 2013) ; dans le même ordre d’idées, on peut avoir Z = Bombyx mori et P = Morus alba, nigra… ; photos 11-1 et 11-2 ;
- Z = cochenille (Dactylopius coccus), P = Nopalea cochenillifera (Cactaceae), a1 = 1, a2= p, F’1 = nourrir Z, F’2 = colorer ;
- avec a1 = a2 = 1, F’1 = nourrir Z, on décrit le fait des P semées pour nourrir le gibier Z des chasses de Sologne ;
- avec a1 = 1 ou e, F’1 = appâter ou anesthésier Z, on décrit le fait des P utilisées pour attirer des animaux Z dans des pièges, les narcotiques du poisson… ; F’2 peut être 0, nourrir… ;
- avec a2 = pem, F’1 = tanner, F’2 = vêtir, on a le fait des P utilisées pour tanner les peaux animales, celles-ci étant utilisées pour vêtir.


Dans le même ordre d’idées, de Léon (1963) évoque la soie issue du ver à soie du ricin (Samia cynthia sur Ricinus communis) en Israël.
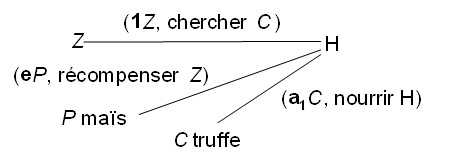
Des interrelations peuvent s’avérer plus complexes, par exemple quand H utilise une truie Z pour détecter la truffe C, puis la récompense avec quelques grains de maïs P (photo 11-3) ; la relation simple H–P du schéma précédent se dédouble ici en H–C et P :
Les élégantes fleurs artificielles de Guyane formées à partir de plumes d’oiseaux Zi offrent encore une interrelation originale pouvant s’analyser ainsi : Σi(e ou em)Zi ≡ seP, les Zi offrent leurs plumes de leur vivant ou non (e ou em), alors structurées (Si) en un ensemble ressemblant (s) à une partie (efleurs) de P.

Le peintre italien Arcimboldo (dit aussi Arcimboldi, xvie siècle) est bien connu pour ses tableaux représentant des assemblages divers structurant des formes de complexité supérieure. Notamment Les Quatre Saisons constitue un ensemble de quatre assemblages de parties de plantes (racines, feuilles, fleurs, fruits, inflorescences…) représentant un buste humain : sΣi(ePik) ≡ seHk ; Hk est un type humain, k est l’indice de la saison et varie de 1 à 4 (photo 11-4 pour k = 1, Le Printemps). En outre les ePik ne sont pas choisis au hasard, mais sont reliés à la saison symbolisée, de sorte que les deux termes de cette équivalence sont les signifiants de la saison, signifiant relativement arbitraire dans le cas du terme de droite (pas vraiment de point commun entre la saison k et seHk sauf peut-être l’image de la jeunesse pour Le Printemps, celui de la vieillesse pour L’Hiver), mais absolument pas arbitraire dans le cas du terme de gauche, car il existe des invariants entre un ePik et la saison k : fleurs et jeunes feuilles/printemps, légumes, couronnes de céréales et fruits estivaux/été, champignons et fruits automnaux/automne, tronc gris et lierre (sempervirent)/hiver.
D’autres tableaux dus aussi à cet artiste constituent des portraits de profession (cuisinier, amiral, jardinier…) structurés à partir d’attributs de ces professions ; ainsi L’Amiral l’est à partir de poissons et de coquilles marines :
sΣi(emZi) ≡ seH, H étant l’amiral.

11.3. Notions de recette et de produit
En définitive, une grande partie de l’e.science naturelle se transcende dans la notion de produit, forme complexe à fonction précise, dont la structuration fait intervenir plusieurs règnes, minéral, animal, végétal, fongique. Une recette peut être alors définie comme l’énoncé (écrit, ou parfois oral dans le cas de traditions elles-mêmes dites « orales ») des règles qui permettent de transformer les initiaux Ei en un produit complexe final. Un aEi peut être appelé un ingrédient et est soumis après préparations diverses (p) à une structuration (Σi, quoiqu’une structure soit plus que la somme des parties). Le produit final est alors de la forme Σi(peEi), pouvant être précédée d’un autre opérateur si une intervention suit la structuration (cuisson c par exemple). Selon la F’ on peut en distinguer divers types.
Produit et recette culinaire
Si F’ = nourrir, alimenter. Le règne minéral y est faiblement représenté (sel, eau, bicarbonate de soude). En passant évoquons le cas particulier de la bûche de Noël, produit structuré puis cuit (c) et finalement diversement ornementé (o) pour simuler une partie de végétal (seP).
Produit et formule pharmaceutique
Si F’ = soigner, guérir. La recette est ici plutôt appelée formule. Dans ce cas, les divers règnes interviennent significativement : borate de sodium, oxyde de zinc, hydrate de chloral, divers sels… (minéral), huiles végétales, essences, teintures diverses… (végétal), blanc de baleine, cire, huiles d’origine animale. De bons exemples de telles formules sont fournis par les électuaires de Cratevas (mythridaticon), à 54 ingrédients (plantes, animaux dont venins), d’Andromachus (thériaque) à 64 ingrédients (entre 40 et 60 selon Clapier-Valladon, 1990, mais ce produit s’avère variable selon les régions ; voir aussi Chast, 2010). La boisson indienne du sacrifice ou parahome, où interviennent la transfomation fonctionnelle b et l’haoma, un produit quelque peu secret (d’origine végétale ou fongique ?), peut s’analyser ainsi

En haute Tinée (Alpes sud-occidentales françaises), on utilisait un crin de mulet frotté sur une tige de Berberis vulgaris décortiquée, l’ensemble étant alors serré autour d’une verrue pour l’éliminer (Association des Stéphanois, 1987).
Produit et recette de poison
Au Nord-Congo, une recette de poison de chasse ressortit au schéma suivant (Strophanthus et Parquetina relèvent des Apocynaceae, famille bien connue pour ses alcaloïdes) :
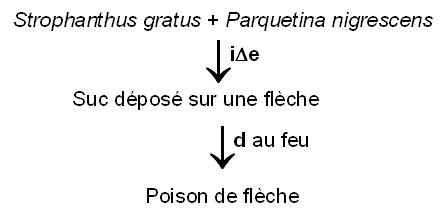
On trouvera bien d’autres CO débouchant sur de tels poisons dans Yongo et al. (2014).
Abri et recette architecturale
Pour F’ = abriter, protéger. On peut évoquer les abris de pierre sèche fréquents en zone méditerranéenne et associés au mode de vie pastoral ou agricole et temporairement occupés. En France, selon les régions pastorales, ils portent des noms variés : gariotes (Quercy, Périgord ; photo 11-5), orris (Pyrénées ; photo 11-6), cazelles (Lot ; photo 11-7), bories (Vaucluse ; photo 11-8), capitelles (Gard, Aude ; photo 11-9), cazotes (Albères ; photo 11-10) ; en Italie ce sont les nuraghi (Sardaigne), les trulli (Pouilles)… Les plus simples répondent au fait ΣR. Évidemment le seul opérateur Σ ne suffit pas pour rendre pas compte de toute la diversité de la structuration architecturale.






D’autres abris font intervenir un matériau végétal, comme les baraques de la côte languedocienne (Salses, Le Barcarès) en Phragmites australis (localement senills ou sanills) et Arundo donax (canyas) (François, 2014).
Parmi de nombreux autres exemples, citons les cases du pays Masaï et des Himba (photos 11-11 et 11-12) :
Σ[{iZ = bouse}, {1R = terre}, {deC = herbes de savane}].


Pour F’ = abriter les abeilles, illustrons le cas de la ruche landaise Σ[iZ, 1R, vdeP] (photo 11-13 ; sur la diversité des ruches, voir de Foucault, 2010a ; voir aussi supra, photos 5-8 à 5-10, et Baumann, 1984 : photo 346).

Instruments et recette instrumentale
Un type de recettes des plus importantes à la vie des hommes, souvent de tradition orale, aboutit à la structuration d’instruments dotés de fonctions utilitaires. Ainsi des haches néolithiques (F’ = couper) faisaient intervenir trois règnes quand, par exemple, le manche était d’érable (P), la gaine en bois de cerf (Z), la lame en pierre (R) polie (t), soit Σ[teP, te ou temZ, tR].
Une hache actuelle du Hoggar (photo 11-14) illustre le fait Σ[{|p|R = lame}, {tdeP = manche}]. Pour un outil mésolithique à pointe d’os avec lames lithiques tranchantes, c’est plutôt Σ[tR, temZ].

Certains outils atteignent une complexité e.scientifique élevée, tel ce couteau des Indiens Haïda (Amérique du Nord-Ouest) à lame de fer (L = pR) et manche d’ivoire de morse Z1 sculpté et incrusté d’haliotis Z2 : Σ[temZ1, ΔemZ2] = M
représentant une tête d’ours Z3 surmontée d’une grenouille Z4 : M ≡ sΣ[eZ3, 1Z4], d’où le produit final Σ(L, M), la structuration ultime de liaison lame-manche étant assurée par des plaques de bois (aP) et des lanières de cuir (aZ) (exposition Les Amériques de Claude Lévi-Strauss, musée de l’Homme, Paris, 1989).
Outre les outils, on peut encore évoquer les instruments de musique accompagnant des rites, distrayant les auditeurs… : clarinette primitive du Maghreb à une ou deux pièce(s) d’un roseau ajustée(s) dans une corne de vache (photo 11-15), caremère du sud-ouest de la France à anche d’Arundo donax, tuyau de buis travaillé et pavillon de corne animale (photo 11-16), bumpa du Burkina Faso en tige de millet aux extrémités de laquelle sont fixées deux calebasses à fonction résonatrice (fonction très courante de ces fruits de Cucurbitaceae ou de Crescentia dans les instruments traditionnels), tiktiri indien à une calebasse et deux tiges de roseau, cornemuse utilisant la peau animale, flûte guyanaise en os travaillé orné de plumes d’oiseaux (photo 11-17), hochets à graines secouées, lithophones… (Diagram Group, 1978).



12. Êtres naturels et communication
De nombreux faits e.scientifiques ont pour finalité une communication entre les hommes. Déjà divers objets astronomiques portent quelques messages ; ce fut le cas des comètes dont le passage fut longtemps considéré comme un présage néfaste. Ainsi la Tapisserie de Bayeux (s. d. : volets 32-33) rappelle un tel message lors du passage de la comète de Halley dans le ciel européen relativement à la lutte qui opposa Guillaume le Conquérant et Harold en 1066 (photo 12-1).

12.1. Supports de communication
Tout d’abord, divers E transformés ou non peuvent servir de support à un message. C’est l’exemple classique des arbres ou des rochers (gravures et peintures rupestres) avec les opérateurs t (si l’arbre P ou le rocher R est travaillé pour inscrire le message ; gravures rupestres ou sur tronc ; photos 12-2 et 12-3) ou 1 si on estime que l’écriture est finalement une relation physique très limitée par rapport à t (cas des arbres supports de balise de randonnée, photo 12-4), ou o dans le cas des peintures rupestres (photo 12-5) ; dans ce dernier cas, la recette de la peinture utilisée serait bien intéressante à connaître. L’une des plus célèbres pierres gravées est la pierre de Rosette découverte par des soldats français, mais aujourd’hui propriété du Royaume-Uni, avec ses trois versions d’un même texte qui ont permis le déchiffrement des hiéroglyphes par Champollion.




Quelquefois, le support de communication est fixé sur un autre support, par exemple une planche portant une information clouée sur un tronc d’arbre. Dans certains jardins botaniques, le nom vernaculaire ou scientifique de l’arbre est lisible directement sur celui-ci.
Le support peut être cependant plus complexe, car il nécessite une préparation p :
- cas d’une tablette d’argile cuite recevant des caractères cunéiformes : ctpR (photo 12-6), l’argile R étant préparée puis travaillée pour inscrire le message, enfin cuite pour fixer définitivement celui-ci ;
- cas du papier à base de végétal : papyrus (photo 12-7), mûrier ou érable à papier (photo 12-8)… : dpeP, avec P = Cyperus papyrus, Broussonetia papyrifera, Acer griseum;
- cas des parchemins et velins d’origine animale (pemZ ; photo 12-9) ;
- cas du recyclage du papier pour fabriquer du papier pour des journaux, des revues, des ouvrages…




Le papier dit antaimoro (ou parfois antemoro) a été introduit par les Arabes au viie siècle à Madagascar pour copier et diffuser le Coran ; récemment, en y déposant des fleurs séchées, c’est devenu un élément décoratif. Adapté de la préparation du papyrus égyptien, il est élaboré à partir de l’écorce d’avoha (Gnidia danguyana, mais peut-être aussi d’autres Thymelaeaceae) bouillie, puis rincée et écrasée à l’aide d’un maillet pour obtenir une pâte (photos 12-10a et 12-10b). Celle-ci, mélangée à de l’eau, est ensuite soigneusement tendue sur un cadre en bois ; les fleurs sont déposées avant le séchage complet (photos 12-10c et 12-10d).




À Sri Lanka, les feuilles du palmier talipot Corypha umbraculifera, célèbre par ailleurs pour sa monocarpie (modèle architectural de Holttum) et sa floraison ultime spectaculaire (photo 12-11), sont préparées en papier ola qui pourra recevoir des textes sacrés (photo 12-12).
Dans le cas des tatouages, c’est le corps humain lui-même qui forme un support à des ornements divers (o), après un éventuel travail préalable (t) sur la peau.
Dès l’Antiquité, des murs et autres montants verticaux ont servi et servent encore de support aux graffiti.
Pour écrire sur les supports (Jackson, 1982), il faut disposer de calames, pinceaux, plumes…, initialement formés à partir de plantes (Juncus maritimus, Phragmites australis par exemple) et d’animaux (surtout oie), ainsi que d’encre (à base de suie, d’eau, de résine, d’oxydes minéraux).


12.2. Des repères du temps
Molet (1990a) donne à lire de très intéressantes considérations sur les êtres naturels pouvant rythmer le temps à travers divers groupes humains de notre planète en fonction de leur état de développement : retour, ponte, éclosion, envol de divers oiseaux, apparition de certains insectes ou poissons, état de l’eau (gel, dégel), force du Soleil, position de certains astres, développement de certaines plantes bien identifiées… Pendant longtemps, le cadran solaire, utilisant la lumière du Soleil (e.astronomie ; photo 12-13), a rythmé le temps dans de nombreuses villes.
Rappelons encore les fossiles marqueurs des périodes géologiques, du Primaire au Tertiaire (Trilobites, Ammonites, Nummulites).

12.3. Des repères de l’espace
Des E peuvent constituer des repères dans l’espace. Un des plus célèbres fut l’arbre du Ténéré (Acacia tortilis subsp. raddiana), aujourd’hui disparu suite à un choc par un chauffeur sans doute ivre et qui servit longtemps de repère sur la route des caravanes traversant le désert éponyme, au Niger (Anonyme, 1974 ; Le Floc’h & Grouzis, 2003 ; photo 12-14). D’autres plus anonymes servent certainement encore aujourd’hui à se repérer dans les zones sahariennes et autres déserts (Frérot, 2011 : 78).

Des arbres ont aussi « offert » leur nom pour créer des toponymes locaux (photo 12-15). D’ailleurs, parfois l’arbre a disparu, il ne reste alors que le toponyme pour conserver une mémoire végétale.
Les cairns sont de petits amas de pierre (ΣR) servant de points de repère le long de sentiers.
D’autre part, des astres, dont l’Étoile polaire pour l’hémisphère nord et la Croix du sud pour l’hémisphère sud, ont pu servir à guider la navigation (du moins en l’absence de nuages) avant l’invention des appareils électroniques.


12.4. Ethnobiologie hiéroglyphique
La communication peut par ailleurs utiliser des modèles vivants, soit {seE} pour constituer des caractères ou des idéogrammes ; c’est notamment le cas des hiéroglyphes égyptiens (Granet & Mathieu, 1990 ; photo 12-17) qui usent de modèles végétaux – roseau fleuri/Phragmites australis (photo 12-18), papyrus/Cyperus papyrus, plant de lotus/Nelumbo nucifera ?, « jonc »…) – et animaux – scarabée, vautour percnoptère, poussin, vipère à cornes, chouette, cobra, jabiru, poissons, carnard pilet et caneton, faon de bubale, lion, ibis, hirondelle, pintade, cormoran… –, voire astronomiques (soleil levant).


12.5. La bioindication
L’observation de quelques E peut par ailleurs renseigner sur des phénomènes d’une autre nature utiles à H. Il existe ainsi des P ou des Z indices (relation 1) : la floraison d’un Liatris (Asteraceae) indiquait aux Indiens nord-américains l’arrêt de la chasse au bison car le maïs arrive à maturité ; en Nouvelle-Calédonie, l’arrivée des baleines à bosse indique aux agriculteurs le moment de planter l’igname.
D’autres indices utilisent l’opérateur e. Ainsi la carline coupée, séchée (deCarlina acanthifolia) et placée sur la porte d’une bergerie (photo 12-19) peut servir d’hygromètre (et non de baromètre comme on le dit trop souvent…). Au Togo, les limites des champs et les propriétés peuvent être indiquées par un emprunt à une communauté végétale (eV), de même pour indiquer aux conducteurs un incident ou un accident sur une route (balisage de sécurité ; photo 12-20). Des petits indicateurs de « bar de brousse » utilisent des ingrédients d’origine végétale (photo 12-21).



D’une manière générale, la flore, la végétation, la faune, la fonge apparaissent comme des médias qui délivrent des informations sur la qualité du milieu dans lequel elles vivent. Les plans d’eau, les sols, les arbres ou phorophytes sont alors les supports du message délivré. Ainsi la végétation des classes phytosociologiques des Lemnetea minoris et des Potametea pectinati informe sur la qualité de l’eau (pollution, concentration en sel, dureté de l’eau ; photo 12-22). Les lichens épiphytes ont été utilisés pour estimer la qualité de l’air environnant (photo 12-23). Au niveau de la végétation terrestre, la bonne connaissance de la synécologie permet aussi de recueillir des informations sur l’environnement, par exemple dans le cas des prairies dont la flore dépend fortement de la richesse trophique du sol. Stehlé (1963) utilise les associations végétales des bananeraies pour orienter le choix et diriger les bonnes pratiques sur les cultures : « la lecture raisonnée du tapis végétal est donc susceptible d’aider considérablement à la connaissance des moyens d’améliorer le sol et sa fertilité ». De même la prospection archéologique peut être facilitée par l’observation de la végétation qui contraste avec la végétation naturelle normale de la station d’étude, une végétation calcicole ou nitrophile dans un contexte globalement acide ou oligotrophe par exemple (Couderc, 1985 ; Ghestem, 2002 ; Ghestem et al., 2003). Cette démarche heuristique est notamment rendue possible parce que la dynamique de la végétation initiale vers la végétation eutrophisée ou neutrophile à basiphile présente un phénomène d’hystérésis comparable à celui qui existe en physique, lequel permet une conservation assez longue, une mémoire de l’histoire qui s’est déroulée sur un site, la dynamique inverse étant bien plus lente pour le retour à l’état initial que la dynamique directe (de Foucault, 2010b).


12.6. Le langage des plantes
On aborde ici les faits e.scientifiques du type {aE, transmettre un message B}, E étant ici une plante P. On va montrer qu’il existe souvent quelque chose en commun, un invariant entre E et B, ce qui permet par ailleurs de classer ces sèmes messagers dans des catégories dénommées « structures » (de Foucault, 1991a, qui offre de bien plus nombreux exemples que ceux retenus ici ; voir aussi l’annexe 2). Comme on le verra, le passage de P à B fera parfois appel à des sèmes tels que *P/N(P)* (N = nom de P), *P/écologie de P*, *P/fonction secondaire e.botanique de P*.
Auparavant, on peut analyser brièvement la comptine consistant à effleurer (verbe parfois impropremernt remplacé par effeuiller) les « pétales » de la marguerite ordinaire en énonçant il (ou elle) m’aime un peu, beaucoup, passionnément, à la folie, pas du tout : ΔecapituleLeucanthemum ircutianum, ces « pétales » étant en fait les fleurs ligulées périphériques du capitule de la plante.
Les structures botaniques
Dans un premier ensemble de structures, P est considéré dans sa totalité, globalement. Une première structure relie directement P à son message : Utricularia/je guette le moment propice, Immortelle (Helichrysum)/toujours, Sensitive (Mimosa pudica)/pudeur… Dans une deuxième structure, intervient un trait intermédiaire comme le port général de P : Glaïeul (Gladiolus, à feuilles ensiformes)/provocation, Aristolochia/étreinte, Lonicera/lien d’amour… Une troisième met en jeu l’écologie de P, alors une association d’idées (AI) permet de passer de cette écologie au signifié : Joubarbe (Sempervivum, plante frugale des milieux déficitaires en eau)/je me contente de peu. Une quatrième fait intervenir un sème e.botanique *aP/F’* et une AI permet de passer de F’ au message : Achillée (Achillea millefolium, utilisé en médecine populaire pour soigner et guérir)/je soulage, Châtaignier (Castanea sativa, jadis arbre de justice)/rendez-moi justice…
Dans un deuxième ensemble de structures, le message n’est relié qu’à une partie de P, d’où l’intervention de l’opérateur e. L’une d’elles est basée sur une qualité de cette partie : Potiron (Cucurbita maxima, à fruits énormes)/grosseur, Balsamine (Impatiens, à fruits à brusque déhiscence élastique)/impatience… Une autre est associée à un objet puis utilise l’AI : Campanule (Campanula, à fleurs rappelant les cloches des animaux)/surveillance.
Les structures linguistiques
Ici, P n’intervient plus physiquement, mais seulement par son nom N(P), éventuellement à travers des jeux de mots et quelques catégories de ces objets linguistiques (de Foucault, 1988b).
Une première structure fait intervenir l’homonymie : Souci (Calendula) homonyme de souci (ennui)/peine, Pensée (Viola)/souvenir. Une deuxième joue sur l’homographie, via notamment des personnages mythiques ; ainsi Narcisse (Narcissus) homographe de Narcisse (fils de Céphise)/égoïsme, puis Adonis/souvenir douloureux, Euryale/amitié à toute épreuve… Une troisième joue sur l’homophonie, rejoignant alors le calembour : Circée (Circaea) homophone de la magicienne Circé/sortilège ; un peu plus complexe, le message Sésame (Sesamum indicum)/ouvrez-moi votre cœur fait intervenir une homographie, une AI et un changement dans la forme conjuguée du verbe ouvrir.
Les structures mixtes
Il peut exister enfin des structures mixtes faisant intervenir des relations botaniques et des relations linguistiques ; ainsi l’Ortie (Urtica dioica, à piqûre douloureuse)/douleur cuisante, avec une homonymie selon le sens propre et le sens figuré du message final, puis Gattilier (Vitex agnus-castus, anaphrodisiaque)/froideur, Lycopode (Lycopodium clavatum)/flamme ardente… La Digitale (Digitalis purpurea, qui soigne les affections cardiaques) pourrait guérir aussi des peines de cœur, d’où le message de consolation.
Cette approche réduite ici au langage des plantes peut s’étendre à l’e.zoologie ; ainsi le poisson représentant le Christ H est le terme final de la suite sémique suivante :
H → N(H) Iesus Christos Theou Uio Soter (pour Jésus-Christ fils de Dieu sauveur) → Ichthus par siglaison
terme homographe de N(Z) où Z est le poisson.
Les bouquets messagers
Plusieurs végétaux peuvent être réunis dans un bouquet pour délivrer un message plus complexe : si on dispose en ce bouquet plusieurs parties de Pi messagères de Bi, alors le bouquet {ΣiaPi} délivrera le message ∪iBi (∪ symbole de réunion dans la théorie des ensembles) ; toutefois ∪ n’inclut évidemment pas la syntaxe du message qui devra être reconstituée par le récepteur.
Le grand ethnobiologiste français J. Barrau (1970) rapporte l’interprétation acquise par enquête d’un remarquable bouquet de Nouvelle-Calédonie (noms des taxons actualisés selon Catalogue of life, www.catalogueoflife.org). Il se compose d’un sommet feuillé de Cordyline fruticosa (qui signifierait parole d’honneur), de fragments de palme de Cocos nucifera et d’un rameau d’Araucaria columnaris (séjour paisible), d’un rhizome feuillé de Chrysopogon aciculatus et d’un rameau de Ficus obliqua (invitation à partager le séjour paisible), une tige fleurie de Strobilanthes alternata (ne pas douter de la sincérité de ce message), le tout réuni par un lien de Lygodium reticulatum (sérieux de ce message), d’où le message global librement traduit ainsi par l’ethnobiologiste : « N’ayez aucune crainte, je vous donne ma parole de façon la plus durable que vous pouvez venir partager la paix de notre village et y parler avec nous en toute sécurité ». En introduction à cette petite étude il citait aussi l’observation par M. Leenhardt, autre ethnologue français, de messages faits de nœuds de tissus d’écorce ou de paille, qui font alors intervenir notamment l’opérateur v pour la réalisation du nœud.
Dans les campagnes, il était de tradition que les ouvriers offrissent au maître un bouquet à la fin des moissons, souvent fait de brins de céréales tressés, par exemple {veBlé ou Orge}.
On peut rapprocher des bouquets le rite iranien des haft sîn (rite des « sept S ») comme figure allégorique d’un tableau symbolique représentant en quelque sorte ce qu’il y a d’immatériel et de transcendant dans notre vie, opposé à ses aspects purement existentiels (Bellakhdar, 2022) : « l’observation de ce rituel consiste à dresser une table comportant sept éléments comestibles et sept autres non comestibles, chacun de ces quatorze éléments commençant par la lettre S et ayant une valeur symbolique qui lui est attachée en propre. Les sept éléments comestibles sont généralement les suivants : des germes de blé ou de lentille (Sabzeh) symbolisant la Renaissance, un pudding sucré (Samanu) symbolisant l’Abondance, le fruit de l’argousier (Senjed) symbolisant l’Amour, l’ail (Sîr) symbolisant la Médecine, la pomme (Sîb) symbolisant la Beauté et la Santé, le fruit du sumac (Somaq) symbolisant l’Éclat du soleil, le vinaigre (Serkeh) symbolisant la Patience et la Sagesse, vertus inhérentes à l’âge. Les éléments non comestibles traditionnellement présents sur cette table sont eux aussi au nombre de sept : des jacinthes (Sonbol) pour le Printemps, de l’argent pour la Prospérité, des œufs peints pour la Fertilité, un poisson rouge dans son bocal pour la Vie, un miroir avec un œuf posé au dessus pour le Reflet de la vie, des bougies pour le Feu et la Lumière et enfin un livre sacré (le Coran chez les musulmans, l’Avesta chez les zoroastriens ou encore un livre de l’un des grands poètes persans, selon la tradition de chaque famille) pour la Science .».
On peut rapprocher des bouquets messagers l’ikebana japonais (ΣiePi), art floral valorisant globalement le vase, les tiges, les branches, les feuilles, les fleurs (contrairement à l’art du bouquet occidental qui privilégie seulement les fleurs), symbolisant le ciel, la terre et l’humanité (photo 12-24).

Les parfums et les mutilations
Au moyen d’alambics, les molécules volatiles de pétales et les huiles essentielles peuvent être extraites pour préparer des parfums : {ieP} (par exemple lavandin, Pelargonium, vetiver Vetiveria, ylang ylang Cananga odorata, eau de fleur d’oranger ; extension possible aux animaux par le musc). Rituellement, ces parfums sont souvent des messagers des H vers les dieux. Plus récemment, on peut se demander si le parfum ne serait pas aussi un message volatil à l’adresse d’un partenaire pour l’attirer, le séduire…
On a présenté précédemment (§ 5.3) diverses autorelations chez H dont beaucoup paraissent aussi avoir valeur de message destiné à autrui ; Chippaux (1990) cite la mutilation (représentée par e↑), la déformation du crâne, des pieds, du cou… (δ↑ ou parfois simplement δ), les incisions de la peau ou scarifications (t↑, ou t si intervient un acteur humain extérieur), l’arrangement de la chevelure (v↑ ou Σ↑), les tatouages, les amputations ; des peintures corporelles sont évoquées par Zahan (1990). Derrière tout cela, il y a une motivation érotique ou religieuse, de défi social, mais aussi d’initiation chez les adolescents.
12.7. Êtres naturels et symboles
De nombreux faits e.scientifiques sont attachés à des symboles matérialisés par des drapeaux, des armoiries ou des emblèmes, souvent de collectivités (territoires, États…) et mettent en scène, entre autres, des animaux et/ou des végétaux plus ou moins stylisés (opérateur s).
En ce qui concerne les drapeaux, on peut évoquer cinq pays qui y ont fait intervenir de tels E : le Liban (sCedrus libani), le Canada (avec seAcer saccharinum ; photo 12-25), la Dominique (un oiseau stylisé), l’Ouganda (une grue royale, Balearica regulorum), la Papouasie-Nouvelle-Guinée (un oiseau de paradis, famille des Paradisaeidae).

Au niveau des emblèmes et armoiries, cette représentation d’êtres vivants est bien plus riche ; on y retrouve le cèdre pour le Liban et l’oiseau de paradis pour la Papouasie-Nouvelle-Guinée. L’Arabie saoudite a retenu le palmier dattier (Phoenix dactylifera), l’Australie un kangourou (Macropodidae) et l’émeu (Dromaius novaehollandiae), la Barbade un poisson, un oiseau (pélican ?) et un hibiscus, la Bolivie un condor des Andes (Vultur gryphus, qui apparaît aussi dans les armoiries de la Colombie et de l’Équateur), le Botswana deux zèbres (Equus sp.), la Côte-d’Ivoire un éléphant de savane (Loxodonta africana), l’Erythrée un dromadaire (Camelus dromedarius), le Guatemala le quetzal (Pharomachrus mocinnus), les Maldives le cocotier (Cocos nucifera), le Mexique un aigle saisi d’un serpent et posé sur un Opuntia, la Namibie, le Qatar et le sultanat d’Oman un oryx (Oryx sp. ; photo 12-26), le Népal un rhododendron (Rhododendron sp.), les Seychelles le palmier endémique (Loidoicea maldavica) et une tortue, Trinité-et-Tobago un ibis rouge (Eudocimus ruber) et un oiseau Cracidae, le Zimbabwe deux koudous (Tragelaphus strepsiceros) et un oiseau, la Tasmanie le thylacine (Thylacinus cynocephalus, actuellement considéré comme éteint), les Terres australes et antarctiques françaises le chou des Kerguélen (Pringlea antiscorbutica), la langouste (Palinuridae) et le manchot royal (Aptenodytes patagonicus). En France, on connaît bien la « fleur de lys » (en fait un Iris), l’abeille napoléonienne.

L’arbre de la liberté (photo 12-27) symbolise la vertu éponyme depuis la période de la Révolution française et, en tant qu’arbre de la vie, la continuité, la croissance, la force et la puissance. Il est devenu au cours du xixe siècle un des symboles de la République française avec Marianne ou la semeuse. De tels arbres sont installés en général dans un endroit fréquenté, apparent, comme signes de joie et symboles d’affranchissement. Ils étaient considérés comme monuments publics. Des inscriptions en vers et en prose, des couplets, des strophes patriotiques attestaient la vénération des populations locales pour ces emblèmes révolutionnaires. Des lois spéciales protégèrent leur consécration, mais tombèrent en désuétude sous le Consulat ; les arbres de la liberté qui survécurent au gouvernement républicain perdirent leur caractère politique. Mais la tradition populaire conserva le souvenir de leur origine.

L’arbre de vie est un symbole d’immortalité dans le monde mésopotamien (photo 12-28).

Le coquelicot (Papaver rhoeas) et le bleuet (Cyanus segetum), deux plantes colonisant en pionnières des terrains plus ou moins dénudés, notamment des surfaces pilonnées lors des combats, ont été associés au souvenir des soldats tués lors de la Première Guerre mondiale, le rouge de la fleur du coquelicot étant relié au sang versé, rappelant aussi, avec le bleu du second végétal, deux des couleurs du drapeau français (de Foucault & Valcke, 1996).
Pour mieux s’identifier, plusieurs sociétés ou associations créent un logo (abréviation usuelle de logotype), emblème signifiant, parfois à base d’éléments naturels plus ou moins largement stylisés (photos 12-29 à 12-31).



Les champignons, les animaux et les fleurs peuvent être utilisés pour orner des timbres en philatélie (Artielle, 1986 ; Peyron, 1960 ; Roussel, 1993).
On propose de rattacher à ce chapitre les « mesures de grandeur ». Par exemple, la monnaie peut servir à mesurer la valeur financière d’un objet ; alors les cauris, coquilles en forme de porcelaine des gastéropodes Cypraeoidae (|em|Monetaria moneta, un binôme sans doute en rapport avec cette fonction ; photo 12-32) dans le monde indo-pacifique, des pierres précieuses (quartz, ambre gris, néphrite…) ou des métaux précieux (or, argent) ont servi de monnaie. Ou encore, le poids peut être considéré comme une autre mesure d’un objet ; alors la graine du caroubier a pu servir pour des opérations de pesage ; le nom de cet arbre est d’ailleurs apparenté au terme carat encore utilisé en joaillerie. Ces faits sont plus originaux que les simples représentations stylisées (s) de E sur des pièces métalliques et des billets de banque (cas des cauris sur des billets des Maldives).

12.8. La théorie des signatures
Dans diverses cultures apparaît l’idée que la nature émet des signes vers H, notamment pour qu’il y trouve de quoi soigner quelques-uns de ses maux. Cette origine presque universelle et sa durée plaident en faveur de sa nécessité, de sa situation de passage obligé dans l’évolution des connaissances humaines, un invariant de la pensée inconsciente de H.
La théorie des signatures (Claisse & de Foucault, 1999b) pose que tout mal doit trouver son remède dans l’univers naturel et que ce remède pourrait être reconnu à travers des signes envoyés vers celui qui sait les capter, signes ayant des analogies avec le mal à soigner (« médecine par analogie »). La nature apparaît donc comme un livre qu’il convient de déchiffrer en y repérant les signes déposés par les puissances créatrices. Selon l’approche formelle développée ici, il s’agit de trouver un invariant entre E et F’ :
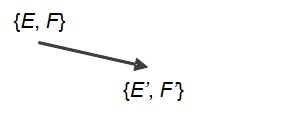
Le tableau 1 rapporte plusieurs exemples de tels invariants. Ils ne préjugent évidemment pas de l’efficacité réelle du remède ainsi repéré. Dans les cas reconnus comme tels, on peut d’ailleurs se demander si la signature n’a pas été recherchée après, comme moyen mnémotechnique de conservation et de transmission du savoir acquis au groupe.
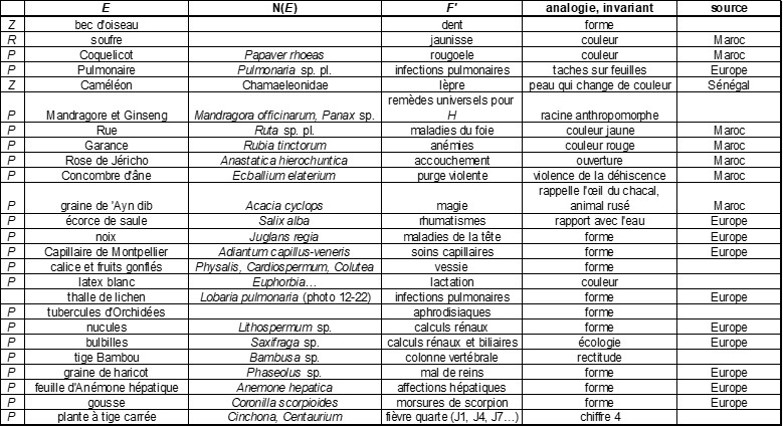
13. Êtres naturels et énergie
Pour leurs besoins quotidiens les hommes doivent bien souvent recourir à une source d’énergie : cuisson des aliments, chauffage, éclairage, déplacements…, et vont trouver dans leur environnement toute une diversité de moyens pour s’en procurer. En fait, il y a plusieurs types d’énergie (lumineuse, mécanique, électrique, calorifique…) qui peuvent passer de l’un à l’autre, mais il est important d’avoir à l’esprit sa conservation quantitative lors de ses transformations.
13.1. L’énergie d’origine végétale
Une des premières de ces sources est le bois, élaboré par les plantes ligneuses au cours de leur croissance. Les chaînes opératoires font intervenir les opérateurs e (photo 13-1), d (séchage), Δ (déstructuration en bûches ou déchiquetage en copeaux selon son intensité ; photos 5-5 et 13-2). L’émondage (e) offre de petits rameaux et dote ultérieurement les arbres ainsi touchés d’un port particulier, dit en « têtard », sur les saules (Salix alba ; photo 13-3), les charmes (Carpinus betulus), les érables champêtres (Acer campestre), les frênes (Fraxinus excelsior)…, tout en offrant parfois aussi un fourrage lors des périodes de pénurie.



Quelquefois, le bois pour le feu est simplement collecté dans des zones inondables où il s’accumule après une crue : |de|P, par exemple dans les oueds sahariens (photo 13-4).

Le feu peut être allumé par la rotation rapide grâce à un archet d’une tige verticale sur une pièce de bois tendre susceptible de s’enflammer, l’énergie cinétique de rotation étant alors transformée en énergie calorifique.
La carbonisation (c) lente de bois dans des meules par pyrolyse et à l’abri de l’air, sans inflammation, aboutit à la formation du charbon de bois (photo 13-5). De telles meules se rencontraient au moins jusque dans les années 1960 dans les forêts françaises (photo 13-6 ; voir aussi Lieutaghi, 1998 : 32-33) et encore actuellement aux Antilles françaises (photo 13-7) ou dans des pays du Sud (photo 13-8 ; voir aussi Cacciari et al., 2003 : 165). Souvent localisée en forêt, près de la ressource ligneuse et donc assez loin des habitats permanents, cette activité s’est accompagnée d’abris forestiers temporairement occupés, par exemple le carcahoux de certaines forêts de Haute-Normandie, qui a fait l’objet d’une monographie e.botanique (de Foucault & Legrand, 1994 ; photo 7-7) analogue à celle de la cabana des feuillardiers (voir § 7), les abris forestiers ardennais, quoique surtout destinés aux sabotiers (Calozet, 1945), ou du plateau de Langres (Fédération des conservatoires d’espaces naturels, 2019 : 83)…




Les « roches organiques » que sont la tourbe (photo 13-9), le lignite, la houille et l’anthracite offrent aussi une source d’énergie et leur utilisation relève autant de l’e.botanique que de l’e.minéralogie. Le boghead est une houille riche en matières volatiles, intermédiaire entre les charbons et les schistes bitumeux, issue de dépôts d’algues. Si on considère l’e.minéralogie, la CO est assez simple (1) ; par contre, dans le cas de l’e.botanique, la CO est plus complexe et fait intervenir des relations indirectes : |pe| où p résume l’ensemble des transformations menant des débris végétaux initiaux au matériau final (tourbication, carbonification encore appelée houillification, avec augmentation de la température et de la pression). Du bois initial et de la tourbe à l’anthracite, on assiste à une augmentation du taux de carbone (50 % dans le bois, 92 à 98 % dans l’anthracite), à une diminution des matières volatiles et finalement à une élévation du pouvoir calorifique. Les « roches organiques liquides » comme le pétrole renvoient à la même CO |pe| mais en conditions sous-marines à partir du plancton. Toutes ces matériaux d’origine végétale se rangent dans la catégorie « énergies fossiles non renouvelables » associée au gaspillage des ressources naturelles, à l’élévation du dioxyde de carbone et au dérèglement climatique actuels.

Dans les régions plus ou moins steppiques où la matière ligneuse manque ou est très limitée, les déjections des troupeaux du Saltus (cf. § 15.2) soigneusement mises de côté et séchées au soleil (e.astronomie) offrent une source d’énergie pour l’hiver (d|i| ; photo 13-10).

Il faudrait encore citer les matériaux d’origine végétale utilisés pour l’éclairage (source d’énergie lumineuse) : huiles, gaz, dérivés du pétrole… (photo 13-11). Plusieurs huiles lampantes issues d’olives ou de noix de coco sont utilisées dans des rituels religieux (photo 13-12).
Le biométhane est un biogaz renouvelable résultant de la fermentation suivie d’une épuration de déchets organiques, notamment végétaux, utilisé comme carburant ou en complément des gaz d’origine fossile.


13.2. L’énergie d’origine animale
Aux hommes, les animaux offrent sutout leur force et leur puissance, souvent à la suite d’une domestication plus ou moins ancienne (animaux de trait) : chevaux, bœufs, buffles (Bubalus bubalis), rennes (Rangifer tarandus), yaks (ou yacks, Bos grunniens ; photo 13-13), ânes, chiens (de traîneau : race malamute de l’Alaska, husky sibérien… ; voir aussi l’carette à quiens de la Flandre et de la Thiérache, attelage d’un chien de trait à une petite charrette pour le transport du lait, pratique aujourd’hui délaissée ; photo 13-14), zébus (Bos taurus indicus ; photo 13-15), Camélidés divers : dromadaires d’Arabie et du Sahara (Camelus dromedarius ; photo 13-16), chameau de Bactriane d’Asie centrale (C. bactrianus), lamas (Lama glama ; photo 13-17) et alpagas (Vicugna pacos) des montagnes andines (les guanacos, Lama glama guanicoe, et les vigognes, Vicugna vicugna, ne sont pas domestiqués), éléphants d’Asie (Elephas maximus ; photo 13-18 ; l’éléphant d’Afrique, Loxodonta africana, n’est pas domestiqué).
Comme le règne végétal (cf. 13.1), le règne animal peut offrir des huiles lampantes ; c’est le cas du beurre de yak pour les cérémonies du bouddhisme tibétain, dans l’Himalaya.






13.3. L’énergie d’origine minérale
On a évoqué un peu plus haut les roches organiques dont les usages peuvent relever tant de l’e.botanique que de l’e.minéralogie. En dehors de ce cas, fort peu de minéraux offrent une source d’énergie. On peut penser à la géothermie, qui utilise la température naturelle des roches, mais il faut surtout citer le cas de l’eau, minéral liquide à la température ambiante. Les rames, pagaies, avirons, hélices, godilles… mues dans l’eau engendrent une force de propulsion entraînant la navigation sur un cours d’eau (photo 13-19). L’hydroélectricité transforme l’énergie potentielle d’un volume d’eau contenue dans un bassin de retenue avec barrage en énergie mécanique pour entraîner des turbines qui transformeront celle-ci en énergie électrique. De ce point de vue, on peut clairement parler de domestication de l’eau (DEau), à l’image de la domestication des plantes et animaux en vue de services rendus aux hommes. Parfois, c’est l’énergie cinétique de cette eau qui est utilisée, par exemple dans les moulins à céréales, à prières (photos 13-20 et 13-21), dans les usines marémotrices…



La domestication de l’eau s’est souvent accompagnée d’agressions sur les écosystèmes aquatiques : altération de l’oxygénation et du pouvoir épurateur du cours, régulation des crues et des lits mineurs, effets sur la vie de ses hôtes animaux (notamment remontée des Salmonidés vers le crénon et le rhitron par des barrages construits sur le cours d’eau…). Devant ces problèmes, on tend actuellement vers une libération, une dédomestication (D-1), de l’eau, tout à fait homologue du retour à l’état sauvage des végétaux et des animaux, par exemple par la construction de « passes à poissons » (photo 13-22).
L’allumage du feu peut passer par la percussion d’un silex sur une pyrite ou autre roche, l’énergie mécanique étant alors transformée en énergie calorifique susceptible d’enflammer un substrat tel que l’amadou (issu du champignon Fomes fomentarius, Polyporaceae ; e.mycologie), un bouquet d’herbes bien sèches…

13.4. L’énergie d’origine météorologique et astronomique
La découverte du feu par les hommes a certainement été contemporaine d’un embrasement par la foudre, mais ce n’est pas un moyen susceptible d’être volontairement utilisé par les hommes, il n’est donc pas de nature e.météorogique.
Grâce aux éoliennes, l’énergie cinétique du vent (1) peut être transformée en énergie électrique (éoliennes ; photo 13-23) ou mécanique (moulins à vent ; photo 13-24) ; le vent attise le feu et gonfle aussi les voiles d’un navire pour l’entraîner sur l’eau. Au contraire des énergies fossiles, celle-ci est renouvelable. Le rapprochement de plusieurs éoliennes dans un « champ » évoque parfois une forêt artificielle où l’énergie solaire utilisée dans la photosynthèse serait remplacée par l’énergie cinétique du vent.


L’énergie solaire utilise une source d’origine astronomique pour produire de la chaleur (énergie thermique pour le chauffage domestique ou la production d’eau chaude sanitaire) ou de l’électricté (énergie photovoltaïque grâce à des cellules intégrées à des panneaux qui peuvent être installés sur des bâtiments ou posés sur le sol ; photo 13-25).

14. Êtres naturels et mythologie
14.1. Généralités sur la mythologie
Au-delà de la classification et de la nomenclature des E, leur apparition peut être expliquée par une démarche scientifique faisant notamment intervenir la génétique et la théorie de l’évolution pour les êtres vivants. En dehors de ce domaine, de nombreux groupes humains expliquent l’origine du monde et de ces êtres par le biais de récits plus ou moins légendaires qu’on appellera ici globalement des « mythes » : « Ne pouvant élaborer des théories conceptuelles fondées sur des réalités vraies, il (l’Homme) légitimise alors ses propres réalités et les transpose sous forme de mythes » (Gonthier, 1990 : 1418). On reprendra ici les grandes idées d’une publication antérieure (de Foucault, 1992c).
Le mythe est reconnu pour véridique par les sociétés qui les narrent (en cela, il se distingue du conte, de la fable), mais pour invraisemblable par les observateurs extérieurs (il se distingue alors du récit historique). Il renvoie à une époque ancienne, éloignée, primodiale, une époque des commencements. Il explique comment une réalité est venue à l’existence, c’est le récit d’une création, quoique pas toujours distinct du conte récréatif qui n’explique rien. Il possède un caractère sacré, faisant souvent intervenir des êtres surnaturels (dieux, héros, fées…), parfois physiquement et moralement anormaux par rapport aux hommes réels (ogres, géants, lutins). À vrai dire, à la suite de Malinovski, Kerenyi (1968) rappelle que le mythe n’est pas tout à fait une explication au sens scientifique (un pourquoi ?), mais serait plutôt une clarification (un à la suite de quoi ?). D’une manière générale, la mythologie fournit des cosmogonies, explications globales de l’apparition de l’univers et de ses diverses composantes… On peut retenir le terme de mythification pour l’association d’un E à son mythe étiologique ; on ne peut évidemment pas parler ici de transformation de l’un en l’autre.
14.2. L’origine des taxons
L’analyse de divers mythes associés aux P permet de dégager plusieurs mécanismes causaux (on renvoie à la publication de 1992c pour de nombreux exemples) qui peuvent être étendus aux autres E :
- le don d’un être surnaturel aux hommes, avec la distinction manichéenne plantes « positives » (belles, utiles…) données par Dieu et plantes « négatives » (piquantes, toxiques, parasites…) apportées par l’anti-Dieu, Satan ;
- la métamorphose ou transformation majeure d’un être en un autre de règnes différents, surtout si l’on considère que du point de vue mythique les hommes n’appartiennent pas au règne animal ; si le résultat de cette transformation est un végétal, elle prend la forme d’une végétalisation, mais on peut aussi reconnaître une animalisation;
- la dérivation ou transformation mineure d’un être en un autre voisin du même règne ; ainsi à Gortyne (Crète) un platane (Platanus orientalis, normalement caducifolié) aurait acquis des feuilles persistantes en souvenir des noces divines d’Europe et de Zeus à l’ombre de cet arbre (photo 14-1) ;
- la naissance spontanée, apparemment plus rare que les mécanismes précédents ; mais si le don est lié à des divinités initiales, celles-ci ont bien dû apparaître spontanément parce qu’encore avant régnait le néant.
En ce qui concerne l’origine des sols S, il est intéressant de faire référence à l’étude de Feller et al. (1986) qui compare un discours paysan antillais au discours scientifique sur la genèse de ces formes : le pédologue sait que la roche s’altère progressivement pour engendrer un sol lors de la pédogénèse ; le paysan antillais pose quant à lui que le sol est la « terre » et que la roche naît à partir de celle-ci, qu’elle « profite de la terre » : au lieu d’une pédogénèse, on a donc une lithogénèse engendrant un R.

14.3. Êtres mythiques
Qui plus est, la mythologie ressent parfois le besoin d’introduire des P irréelles, mythiques. On n’en connaît souvent que le nom, parfois quelques traits morphologiques ou écologiques. Ce besoin est lié au fait qu’elles remplissent une fonction dans certains mythes.
Il existe aussi des Z mythiques, connus le plus souvent par leur représentation stylisée, au contraire des P mythiques guère représentées. Ces êtres fabuleux sont forgés de toutes pièces (dragon, vampire) ou formés par recombinaison d’au moins deux animaux réels (satyre, centaure, minotaure, sphinx, griffon… ; photo 14-2), soit ΣieZi ; ici, il y a donc une certaine structure matérielle, quoiqu’irréelle. La Tapisserie de Bayeux (s. d.) en rapporte de nombreuses illustrations dans les bandes encadrant les scènes historiques (voir par exemple le haut de la photo 15-48).

Une extension est possible aussi chez les champignons et surtout dans le règne minéral. À la suite de Flamand (1981), Gonthier (1990 : 1434) évoque des pierres imaginaires (antipathès, atizoé, brontée…). De son côté, la pétrification explique l’apparition de pierres R ou reliefs G anthropomorphes ou zoomorphes. Il est d’ailleurs intéressant d’analyser le mécanisme intellectuel, d’essence mythique, qui fait passer de ressemblances morphologiques entre deux formes à des parentés génétiques, expliquant par des transformations considérées comme réelles les passages d’une forme à l’autre. On peut illustrer ce phénomène par le beau mythe mexicain associé à deux massifs montagneux du sud de Mexico City, le Popocatépetl et l’Iztacchihuatl : Popo était un guerrier épris de la fille d’un empereur, Iztacchihuatl, laquelle mourut de chagrin lorsqu’il partit en guerre ; à son retour, il bâtit les deux montagnes, allongea le corps de sa bien-aimée sur l’une et resta sur l’autre avec sa torche funéraire ; de fait, la croupe supérieure de l’Iztacchihuatl évoque bien un corps couché, le Popocatépetl, volcan actif, évoquant la torche funéraire (photo 14-3). Au Sahara algérien, selon un mythe des Touareg, l’Akar-Akar serait le résultat de la pétrification d’un géant (photo 14-4).On connaît aussi la mythique Méduse, un des trois Gorgones, qui pétrifiait tout mortel croisant son regard.


Il existe aussi des éléments géomorphologiques mythiques, le plus connu étant sans doute le mont Meru (à ne pas confondre avec des monts Meru réels, tel celui du parc d’Arusha en Tanzanie) des hindouistes, qui constitue un axe central de la cosmogonie correspondante ; dans les temples actuels, il est représenté par de hautes constructions appelées gopuram (photo 14-5). D’autres, non mythiques, sont censés abriter des dieux ; le plus célèbre est sans doute le mont Olympe, en Thessalie (Grèce ; photo 14-6).


Ces objets posent un intéressant problème à l’e.botanique formelle. Si on se rappelle de la dualité fondamentale structure/fonction (cf. § 2), les faits e.botaniques liés à ces objets correspondent à la formule (0, F’), où 0 désigne une structure nulle, l’absence de structure matérielle suite à leur irréalité.
Évidemment, la science explique l’apparition des êtres naturels par des approches bien différentes. Ainsi, la géologie a éclairci l’origine des roches, des fossiles, des reliefs, du volcanisme. Les progrès de la climatologie ont exorcisé l’intervention des dieux et des héros, les remplaçant par le jeu des hautes et basses pressions. Pour les êtres vivants, on sait que la génération spontanée a été combattue au moyen d’expériences rigoureuses menées par Redi, puis Spallanzani et surtout Pasteur. L’apparition des espèces est sous-tendue d’implications génétiques et de mutations, favorisant les espèces les mieux adaptées. Des quatre mécanismes reconnus dans la mythologie, il ne reste donc que la dérivation à laquelle il faut cependant ajouter le hasard, avec une étape intermédiaire pré-scientique (voire encore actuelle dans certains pays) où le don reste encore bien présent (créationnisme) ainsi que la naissance spontanée.
Le long de l’axe historique, il y a augmentation de l’universalité explicative : à la grande diversité des mythes expliquant un être particulier, se substitue une théorie unique, globale, économique, expliquant un maximum de phénomènes par un minimum de mécanismes. Il y a diminution de l’implication des hommes, encore bien présents en mythologie, diminution de l’intervention du surnaturel (dieux, antidieux), de l’intervention de causes extérieures aux êtres naturels. La vitesse d’évolution diminue le long de cet axe : on peut considérer que le don, la métamorphose et la naissance spontanée se produisent quasi instantanément (vitesse infinie), alors que l’évolution des espèces est très progressive (vitesse très lente ; c’est probablement cette durée inaccessible aux humains qui a produit la théorie fixiste des espèces). Il reste quand même encore le hasard ; en définitive cette variable pourrait constituer la dernière influence du surnaturel dans la science moderne.
15. L’ethnophytosociologie
L’e.phytosociologie, e.science naturelle parmi les plus récemment apparues, a été incidemment évoquée par Kuhnholtz-Lordat dans son œuvre, puis par moi-même (1984 : 2). Mais c’est surtout en 1990 que je l’ai vraiment fondée et formalisée. J’y ai notamment proposé d’élargir le contenu de certains opérateurs classiques des e.sciences naturelles pour les utiliser en e.phytosociologie, par exemple e pour le fauchage et le faucardage, d pour le drainage, g pour l’eutrophisation. Elle a parfois été incidemment illustrée dans les paragraphes précédents, mais de nouveaux développements justifient que lui soit dédiée une grande partie au sein de ce panorama.
Cet article étant surtout dédié à la formalisation et à l’illustration des e.sciences naturelles, on n’abordera pas ici la phytosociologie populaire (de Foucault, 1995b), car ce serait une simple redite de cette publication. On renvoie donc à celle-ci, ainsi qu’à Baumer (1965), Meilleur (1984, 1987) et Scarpa & Arenas (2004) pour les détails et des compléments.
Renvoyant à la publication de 1990a pour d’autres développements, on va en premier lieu aborder ici la trilogie Silva – Saltus – Ager (incluant l’Hortus), déjà connue des Latins, mais qui fut particulièrement développée par Kuhnholtz-Lordat. S’il est pratique de distinguer ces stades pour les décrire et en dégager les caractéristiques, il faut bien reconnaître qu’un groupe humain participe souvent des trois. Par ailleurs on trouvera dans le bel ouvrage de Jean-Brunhes Delamare (1985) de nombreuses illustrations sur le Saltus et l’Ager.
15.1. La Silva
Ce premier type de civilisation se caractérise par l’invariant {1V, assurer la subsistance}. C’est par excellence la civilisation du prélèvement nécessaire à la survie ; il n’y a pas de transformation qualitative de V (d’où 1), sauf dans le passage vers le Saltus et l’Ager et dans les périodes modernes. En revanche, naturellement, des faits e.botaniques, e.zoologiques ou autres viennent s’y infiltrer, par exemple par la cueillette (Tubiana, 1969 ; photo 15-1), la chasse, la pêche. Parfois la récolte des produits alimentaires se fait sans coupe préalable, simplement par égrenage au-dessus d’un réceptacle ; ainsi des Indiens nord-américains récoltent un « riz » sauvage américain (en fait Zizania aquatica ou Z. palustris) en s’approchant de la végétation, en inclinant les inflorescences mûres vers l’embarcation et en les battant pour recevoir les caryopses (Jean-Brunhes Delamare, 1985 : 71).

On peut rattacher à cette première civilisation les Aïnous d’Hokkaidō (nord du Japon), les Vedda de Sri Lanka, les Pygmées de la forêt équatoriale africaine, les San du sud de l’Afrique, les Aborigènes d’Australie, les Semang de la presqu’île de Malacca, les Punans de Bornéo, les Amérindiens de la taïga subarctique, jadis diverses ethnies indiennes des USA actuels et de Patagonie.
Cette civilisation est surtout à l’œuvre dans la forêt, système naturel pluristratifié de haute complexité et de grande richesse biologique. Les H s’abritent dans des cavernes ou des abris rocheux naturels (1G), parfois seulement partiellement (photo 15-3), voire des dunes de sable (côte désertique de l’Angola), des huttes (photo 15-2), parfois temporairement des broussailles ; Jelínek (1989 : 285) évoque les abris aborigènes australiens en écorce pour la protection contre le soleil {ΣeP, -1Soleil}. Les zemlianka du Kamchatka et du Japon septentrional sont des habitations semi-enterrées (tG) pour se protéger (Jelínek, 1989 : 136), notamment des vents violents. Pour leur subsistance, ils chassent, surtout les grands mammifères placentaires Euthériens, parfois des oiseaux, des larves… En Australie où les Euthériens n’existent pas à l’état naturel, leur place est prise par les Marsupiaux ou Métathériens (kangourous notamment). Cette chasse fait parfois appel à des déguisements pour mieux approcher le gibier, ce que l’on peut tenter de formaliser en oH ≡ seZ, un H se déguisant pour mimer une partie d’un animal convoité et ainsi l’approcher au plus près.


Les animaux peuvent être attirés par des appâts, des appeaux ou des trompes (par exemple de Betula, photo 15-4). L’acquisition du gibier passe par des pièges (voir notamment Mérite, 1942) : la photo 15-5 montre l’un d’eux élaboré par les Indiens Guarani d’Amazonie, creusé dans le sol, le plancher du trou obtenu étant alors hérissé de pointes ligneuses dressées, soit Σ(tG, ΛteP), P non déterminée. De son côté, Gillet (1974) décrit un terrible piège à girafe chez les Tamas du Tchad oriental (on renvoie à cette publication pour les détails du système). Elle passe aussi par des armes diverses : traditionnellement sarbacanes et flèches, plus récemment fusils. Les flèches peuvent être empoisonnées par des produits divers (Yongo et al., 2014) d’origine végétale (curare…) et animale (venins) (photo 15-6). La pêche peut se faire par le biais de narcotiques qui permettent d’attraper plus facilement les poissons ainsi anesthésiés (Leblic, 1995). La conservation de la viande est facilitée par le fumage (synonyme de boucanage), une cuisson légère (cZ) par la fumée issue d’un feu.



Le fonctionnement du Précipice à bisons Head-Smashed-In, au Canada occidental, labellisé Patrimoine mondial de l’humanité par l’Unesco, consiste à pousser des troupeaux de bisons vers de hautes falaises {1G, F’ = mZ} ; ce type de piège est évoqué par Jelínek (1989 : 78) et se rapproche beaucoup de celui qui aurait été utilisé par les Néandertaliens pour les bouquetins en Asie centrale évoqué ailleurs par Jelínek (1989 : 49).
On peut rapprocher de ces faits la collecte de l’eau à la base des arbres recevant des précipitations occultes venant du large marin (photo 15-7).

La cueillette touche bien sûr les P (racines, tubercules, feuilles, fruits…), mais aussi des animaux ou des produits animaux fixés (œufs, miel), des mollusques saxicoles…
Les vêtements sont fabriqués à partir d’éléments végétaux (écorces : {ΣeP, vêtir}) ou animaux (peaux cousues, au moyen de nerfs notamment : {ΣemZ, vêtir}). On remarquera que les tissus externes protecteurs des végétaux et animaux deviennent, après diverses transformations, éléments protecteurs externes des organismes humains dans des régions auxquelles H est foncièrement inadapté (dynamique à F’ invariant). On peut en rapprocher l’usage de la graisse de phoque utilisée par les Amérindiens de Terre de Feu, aujourd’hui disparus, pour se protéger du vent et du froid.
Le sacré touche aussi l’e.phytosociologie à travers les bosquets et les forêts sacrés (de Foucault, 1993 ; photo 15-8). Ainsi, à Rome, les sept collines étaient couvertes de bois de chênes consacrés à Jupiter, donc à valeur de sanctuaires. Le caractère sacré est surtout marqué négativement, obéissant à des prohibitions : pas de récolte de bois, ni pâturage animal ni chasse, pas de pénétration par des personnes autres que quelques initiés ou des adolescents en initiation ; toutefois ils pouvaient accueillir des sépultures. Ce caractère sacré a des incidences environnementales non négligeables : préservation de la biodiversité, protection des sols, des sources et des rivières pour la ressource en eau en vue de l’alimentation de l’Ager (cas des rizières en Asie ; Aumeeruddy & Bakels, 1994).

Avec leurs troncs souvent dressés vers le ciel, les forêts sacrées ont peut-être servi de modèle aux cathédrales qui les ont suivies comme l’ont remarqué Chateaubriand (Le génie du christianisme, 3e partie, livre I, chapitre VIII, Des églises gothiques), puis Huysmans (1898, qui parle de futaie de cette cathédrale), Crampon (1936), Mercier (1990), Barrier (1991), de Foucault (1993a). Les troncs y deviennent colonnes (photo 15-9), les houppiers des voûtes avec éventuellement des sculptures de feuillage… ; des oiseaux (Colombidées, Corvidés) adoptent les tours pour y installer leurs nids ; ajourées elles les laissent même y pénétrer. Des prêtres, des évêques, voire des chefs civils ont été enterrés dans leur église comme d’autres l’ont été en forêt sacrée.

L’animisme, c’est-à-dire la croyance en des êtres spirituels, est encore bien présent dans ces groupes humains (photo 15-10), associé à la culture des morts et des ancêtres, étendus aux P et Z, voire même aux objets inanimés ; c’est donc globalement un culte à la nature.

Le tronc d’arbres isolés peut aussi constituer un tombeau pour des personnalités, comme des griots dans des baobabs en Afrique tropicale. On peut en rapprocher les falaises abruptes creusées (tG) pour constituer des nécropoles à l’abri des violateurs de sépulture ; ainsi les Toraja de Sulawesi (Indonésie) utilisent certains reliefs abrupts, plaçant en outre des effigies en bois (tau tau) à l’image des défunts en façade extérieure, comme pour tisser un lien entre eux et les vivants (photo 15-11). Ajoutons que des enfants toraja décédés avant l’apparition des dents peuvent être inhumés dans des troncs (tP ; photo 15-12) ; les tissus de l’arbre se reformant, ils absorbent le petit corps mort ainsi confié à un corps vivant.


Quelquefois, la forêt sacrée pourrait se réduire à quelques arbres seulement comme le montrent certains oratoires de campagne surmontés d’une petite canopée (photo 15-13). D’ailleurs certains arbres de bord de route, devenus involontairement meurtriers {|m|H, 0}, peuvent être convertis en petit édifice de mémoire (photos 15-14 et 15-15). On retrouve un fait comparable lorsque les racines aériennes simulent de petits troncs dressés, comme c’est le cas sur le figuier sacré (Ficus religiosa), au milieu desquelles sont installés de petits oratoires (photo 15-16).




D’une manière générale, ce mode de vie est caractérisé par une extrême extensivité et donc d’importants déplacements. Lorsque les saisons ne sont pas trop défavorables, les H se déplacent à la recherche du gibier ou plus généralement des êtres naturels (pirogue monoxyle teP ou plus complexe ; photo 15-17). Lorsque la saison froide est particulièrement contraignante, les groupes se rapprochent des rivages maritimes, plus tempérés, et regagnent l’intérieur des continents en été (cas des Ona de Patagonie).

Le mot clé caractéristique de la Silva, le prélèvement, peut s’étendre à des civilisations non liées à des forêts pour la simple raison qu’elles vivent dans des régions où la forêt ne peut plus croître à cause des contraintes climatiques. C’est le cas des Inuits qui prélèvent dans leur milieu plutôt hostile des animaux (phoque, caribou, renne, bœuf musqué, des Placentaires) qui offrent des vêtements, des matériaux pour s’abriter, les kayaks pour les déplacements (voir par exemple Le Mouël, 1969) ; ils se déplacent entre l’été (vie sous tente, pêche aux Salmonidés, chasse, un peu de cueillette) et l’hiver sur la côte plus tempérée (chasse aux Mammifères marins ; construction d’igloos : {ΣΔEau gelée, abriter}). Certains igloos des territoires inuits ont une charpente d’os de baleine supportant des briques de glace empilés (Molet, 1990b : 279), soit Σ(dR, ΣeZ).
Au-delà des ses variations dans l’espace, la Silva a pu varier dans le temps : en remontant à des périodes historiques anciennes, elle a existé en Europe, notamment à côté de l’Ager. Selon les saisons, la cueillette dans la nature apportait de nombreux produits animaux (gibier, miel servant de sucre avant l’amélioration et la culture de la betterave puis la découverte de la canne à sucre, charbon de bois et les meules ou les abris associés ; cf. photos 13-5 à 13-8), végétaux ou fongiques (fruits, sève, champignons, remèdes et poisons, psychotropes ; photo 15-18).

Puisque la Silva caractérise un stade primitif de l’ethnoévolution, on comprend qu’elle soit pratiquement disparue de nos civilisations modernes. Pourtant, cet effacement n’est pas complet : H a toujours besoin de la forêt, plus gééralement de la nature, mais la fonction secondaire F’ n’est plus la subsistance, devenant plutôt loisir, ressourcement… Les faits e.scientifiques peuvent toutefois s’y retrouver : si la chasse (safari, gibier d’eau ; pièges artisanaux), la cueillette et la pêche relèvent des loisirs, les produits prélevés sont encore souvent consommés, même si la survie de H n’en dépend plus fondamentalement. Certains prélèvements se pratiquent toutefois à grande échelle : pêche industrielle, coupes massives de bois en forêts naturelles.
Parmi les pièges artisanaux évoqués au niveau de la chasse, on peut décrire la « tenderie aux grives » (et autres oiseaux, car non sélective), propre à certaines communes des Ardennes franco-wallones (Calozet, 1945 ; Clerc, 1987 ; de Foucault, 1988a). Placé à hauteur d’homme ou parfois au ras du sol, ce piège est fixé à un bouleau (1Betula pendula ; photo 15-19a : 1) et est constitué d’un rameau de sorbier (« ployette ») plié et fiché dans le tronc du bouleau (eSorbus aucuparia ; 2). Ce rameau porte d’une part deux filins en crin de cheval (vpeCheval ; 3, à fonction d’étranglement) et d’autre part un fragment d’infrutescence mûre du même sorbier (eSorbus aucuparia ; 4, à fonction d’appât). Ce piège (photos 15-19b et 15-19c) assez barbare est très décrié de nos jours par les mouvements de protection de la biodiversité, à juste raison, mais continue à être utilisé en tant que « patrimoine cynégétique local » (par cinquante-deux chasseurs au 20 décembre 2021 selon L’Obs n° 2982 : 39). Un piège un peu équivalent est décrit de Sardaigne par Murru Corriga (1982).
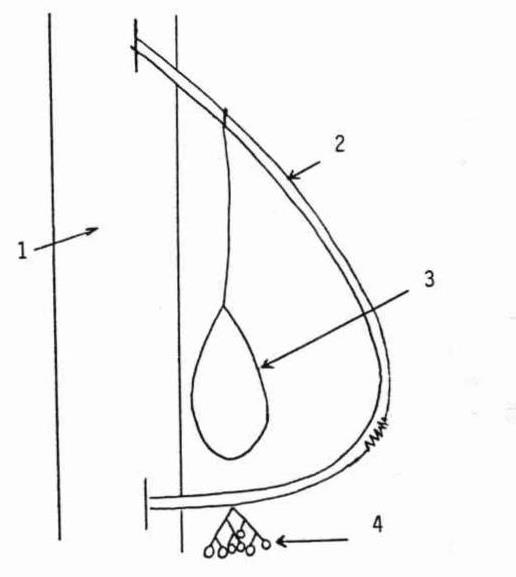


On pourrait encore discuter du rôle des bois et forêts (V) dans les guerres (de Foucault & Amat, 1996). Dans beaucoup de cas, V est utilisé tel quel sans transformation, pour des F’ diverses : abriter les combattants, protéger le repli des armées, constituer le champ d’essai d’armes nouvelles et des techniques de défense, se protéger de l’aviation…). Ces formations sont utilisées par les unités combattantes comme des points d’appui et leurs lisières comme des ados dans le système défensif. Ainsi la réflexion tactique intégra la forêt à tous les échelons de la hiérarchie. Les massifs forestiers n’ont toutefois pas que des avantages : obstacle à la perception du champ de bataille et à la progression des troupes, inefficacité des tirs tendus en l’absence de vue dégagée… Parfois V doit être aménagé pour mieux remplir sa future fonction ; c’est le domaine du génie militaire : tV.
Les arbres isolés P peuvent être utilisés lors des guerres (avec surtout l’opérateur 1) comme supports d’objets divers (câbles et isolateurs électriques, barbelés…), observatoires (photo 15-20), repères pour les pointages d’artillerie ou la marche des fantassins ; des observatoires mobiles ont utilisé des arbres postiches (sP). On peut rapprcher de ces observatoires les postes de surveillance présents au bord de certaines rizières de Sri Lanka pour éloigner les prédateurs (photo 15-21).


Actuellement, la Silva paraît encore plus présente chez l’enfant que chez l’adulte moderne comme si, dans son développement « ethno-ontogénique », l’espèce humaine devait repasser par quelques étapes de son développement « ethnophylogénique ». F’ est alors ici plutôt amuser, distraire. On jugera de la valeur de cette assertion à la nature des jeux des enfants qui rappellent assez les invariants cités pour la Silva : fascination des grottes (plusieurs des grottes ornées aujourd’hui les plus célèbres ont été découvertes par des adolescents), fabrication de cabanes ou huttes (photo 15-22), cueillette en grapillage, dénichage des oiseaux, armes enfantines comme sarbacanes, frondes et arbalètes (photo 15-23), flèches de nerprun (cpeRhamnus cathartica) et arc de noisetier (Valcke, 1989 : 154, 156), petits pièges et collets, « cigarettes » de Clematis vitalba ou de moelle de Sambucus nigra (Valcke, 1989 : 184). Les mouvements d’adolescents (scoutisme) vont aussi dans le même sens avec feux de camp, initiation à la survie, perception des indices laissés par la nature, utilisation des astres pour s’orienter, jeux de piste… D’ailleurs, d’une manière générale, l’e.botanique enfantine est d’une grande richesse (voir notamment Durand-Tullou, 1972 : 244 ; Valcke, 1989 ; Armengaud, 2009). Selon Poirier (1990 : 958), cette dynamique est un cas particulier de la transposition fonctionnelle déjà citée : « transposition ludique d’un objet abandonné : ainsi, à Madagascar, l’arc et les flèches disparus depuis longtemps survivent comme jeu chez les Mahafaly ».


15.2. Le Saltus
La civilisation du Saltus est bien différente de celle de la Silva, bien qu’elle porte encore sur des végétations spontanées. H ne prélève plus directement dans la nature pour subsister, mais survit grâce à des animaux plus ou moins domestiqués qui lui fournissent ce dont il a besoin : Saltus → Dherbivores → H (photo 15-24). Dans cette chaîne pourront intervenir des faits zoophytosociologiques associant Z et saltus parcourus {eV par Z, nourrir Z}, des faits e.phytosociologiques {aV, nourrir le troupeau}, des faits e.zoologiques {aZ, F’}.

Les faits e.phytosociologiques
Les faits de nature trophique
Dans ces premiers faits, on est amené à distinguer plusieurs types de Saltus. Le Saltus est dit « primaire » (Saltus I) quand V ne dérive pas de la régression d’une forêt et qu’elle est utilisée sans transformation, soit simplement 1V. À cette catégorie se rattache l’utilisation pastorale de végétations primaires relevant en Europe des pelouses alpines (Caricetea curvulae, Salicetea herbaceae, Elyno-Seslerietea caeruleae… ; photo 15-25), des pelouses primaires non alpines (Sedo-Scleranthetea biennis), des sansouires (Sarcocornietea fruticosae, Thero-Suaedetea splendentis, Juncetea maritimi…), des steppes et savanes primaires pour des raisons climatiques (de Foucault, 2013 ; photos
13-16 et 15-26), ainsi que des communautés phytoplanctoniques, bases de l’alimentation des huîtres et moules. Kuhnholtz-Lordat (1958) cite encore le cas des arbres abattus pour que les plantes parasites et épiphytes soient mises à la disposition du bétail.


La formule 1V du Saltus I pour Z rappelle beaucoup la formule 1V de la Silva pour H ; cela signifie que le Saltus relève de la civilisation de la Silva pour Z. On peut par ailleurs concevoir une évolution menant de cette dernière au premier dans laquelle la forêt ne serait plus utilisée directement par H pour sa subsistance, mais utilisée pour nourrir un troupeau. Ce fut le cas notamment au Moyen Âge, lorsque les forêts étaient parcourues par des troupeaux plus ou moins domestiqués (par exemple panage des porcs). Dans certaines régions, la strate arborescente des forêts claires peut être utilisée par Z (notion de pâturage aérien ; Piot, 1970 ; Boutrais, 1980 ; photo 15-27). Le pâturage intensif des arbres induit, on le sait, des modifications de ceux-ci connues sous le nom de biomorphoses (photo 15-28).


Parfois l’affouragement des Z en rameaux fins et feuilles d’une strate arborescente supplée le manque lié à une saison sèche : {eV, nourrir Z} (photo 15-29). Au Népal, trop intensivement appliquée, cette pratique est très nocive aux arbres qui passent par un stade « squelettique » avant finalement de mourir (Bergeret & Petit, 1986 : 132).

Le Saltus est dit « secondaire » (Saltus II) quand V dérive d’une V initiale non valorisable en tant que fourrage. Par la pratique de la pastoralisation (L.A. Fabre, in Kuhnholtz-Lordat, 1958), celle-ci est détruite, voire brûlée (b) pour libérer du terrain en vue d’étendre les surfaces pastorales, plus rarement les arbres sont mis à mort (m par dessévage ou annélation ; Fournier & Sasson, 1983). Sur le plan strictement phytosociologique, de nombreuses classes de végétation secondaire peuvent être soumises au pâturage (Rosmarinetea officinalis, Cisto-Lavanduletea stoechadis, Festuco-Brometea erecti, Agrostietea stoloniferae, Arrhenatheretea elatioris, Nardetea strictae, Stipo-Trachynietea distachyae… pour se contenter de l’Europe). Évidemment, le Saltus II est plus souvent un mélange de ces V secondaires, un paysage, qu’un individu d’association homogène auquel Z ne saurait se limiter ; on retrouve ici l’opposition entre les agronomes, pour lesquels la parcelle est une unité expérimentale, et les phytosociologues, pour lesquels c’est un paysage (de Foucault, 1981 : 25).
Les transformations d’origine anthropique peuvent s’accompagner naturellement, c’est-à-dire sans intervention directe de H, de transformations édaphiques comme l’oligotrophisation en système phytosociologique hygrophile (apparition de bas-marais à partir de prairies hygrophiles), l’humidification en système mésophile (remontée de la nappe d’eau temporaire suite à la déforestation, avec développement d’un horizon de pseudogley)…
Dans les agrosystèmes axés sur l’élevage intensif, la ressource des saltus initiaux est souvent insuffisante pour la production des troupeaux. L’éleveur doit améliorer cette ressource par des transformations trophiques, dont l’eutrophisation (g), l’amendement calcique par le chaulage, le drainage ou au contraire l’irrigation dans les régions ou l’eau est facteur limitant (photo 15-30) ; l’endiguement permet de remplacer des prés halophiles en prés seulement oligohalophiles par déchloruration progressive des sols. En Auvergne, l’ayguade est le saltus non transformé, s’opposant à la fumade fertilisée par le parcage de nuit, le parc (bargueriro) étant périodiquement déplacé. Ces transformations peuvent combiner leurs effets. Par exemple, le laquage est une pratique traditionnelle d’amélioration des alpages de moyenne Tarentaise par fertilisation et irrigation (Vertès, 1983). Parfois l’incendie (b) est à considérer comme opérateur d’amélioration car, appliqué périodiquement, il permet le maintien d’une V plus appétente, par les ovins notamment, que la V ligneuse (entretien des landes d’Écosse par exemple).

L’eutrophisation qui vient d’être évoquée n’est parfois pas de nature e.phytosociologique. C’est le cas lorsque le troupeau est parqué plus ou moins massivement sur un saltus originel qui dérive alors vers un saltus eutrophile où l’on peut observer des végétations du type Onopordetea acanthii ou Rumicion alpini.
D’autres cas d’amélioration d’un saltus font appel à l’épandage de goémon, ce dernier constitué d’algues détachées d’une Silva sous-marine et récoltées sur les plages à marée basse (photo 15-31).

Le Saltus amélioré étant lié à une sédentarisation de l’élevage, dans les pays à saisons contrastés dont l’une est défavorable à V (saison froide, sèche), l’éleveur doit accumuler des réserves de fourrage pour nourrir le troupeau lors de cette saison. Intervient alors le fauchage (eV) dont le produit est conservé par voie physique (dessiccation d en foin ; photo 15-32) ou biologique avec préparation préalable (ensilage, p). Parfois les gerbes de foin sont retenues par un brin torsadé de ce foin, soit {veV, tenir la gerbe}.

Les faits d’autre nature
D’autres saltus n’ont pas une fonction trophique pour le troupeau, étant plutôt utilisés comme prés à litière fauchés ou faucardés, puis séchage du produit coupé {deV, pailler}, là où les céréales sont trop peu répandues pour que leur paille assure cette fonction (voir par exemple Valcke, 1994). En Europe centrale, V est représenté par des végétations peu nutritives : roselières, mégaphorbiaies, prairies oligotrophiles du Molinion caeruleae. Au Pays basque, c’est la touya (photo 15-33), complexe phytosociologique régressif de la forêt initiale à Quercus pyrenaica où se mêlent des ourlets acidiphiles à Euphorbia angulata, des pelouses oligotrophiles à Pseudarrhenatherum longifolium, des moliniaies à Cirsium filipendulum. Dans les monts d’Arrée, en Bretagne, des landes à Ulex gallii et des pelouses à Agrostis curtisii étaient aussi utilisées à cette fin.
Dans l’étrépage, c’est tout l’ensemble de V qui est utilisé {1V, pailler}. Le foin peut être aussi destiné à la conservation des fruits, au bourrage des matelas (Guelliot, 1975 : 119, avec un Eriophorum, des fougères)…

Les faits zoophytosociologiques
En général, Z utilise V sans transformation autre que e. Pourtant la présence du troupeau élimine des espèces sensibles, sélectionne les espèces les plus résistantes, eutrophise le milieu en modifiant la flore, mais ce sont là des relations indirectes du type |g|. La relation Z-V est aussi indirecte dans le cas du foin et de l’ensilage, alors du type |de| ou |pe|.
Les faits e.zoologiques
Ces faits sont très nombreux, il convient de les ordonner pour mieux les saisir. Tous renvoient d’abord à la domestication (§ 4.1), DZ
Gardiens, chiens et troupeaux
Sauf cas d’élevages très extensifs, les troupeaux sont gardés par des pasteurs, des bergers… Selon les régions du monde, divers termes s’y rapportent : marcaire des Vosges, gardian de Camargue, tchiko de Hongrie, cow boy d’Amérique du Nord, boiadero ou vaqueiro du Brésil, gaucho d’Argentine, ovejero de Patagonie. Ces hommes peuvent être assistés par d’autres animaux, surtout des chiens, différentes races de ceux-ci étant associées au pastoralisme (patou pyrénéen, bergers divers… ; photo 15-34). Souvent initialement, ces chiens étaient surtout destinés à écarter les voleurs et prédateurs des troupeaux (molosses de défense ; cas observé chez les Kurdes de Turquie orientale contre les loups et les ours) ; actuellement ils ont plutôt pour fonction d’assister les gardiens et d’ailleurs aussi de rompre leur solitude pendant la saison de pâturage.

On peut en rapprocher quelques opérations menées sur Z, notamment le marquage pour faciliter la reconnaissance par les propriétaires, opération assimilée à tZ, comme l’entaille des oreilles des rennes chez les Saami, des moutons à Ouessant (Sélection du Reader’sDigest, 1978 : 222) ; la ferrade correspond plutôt à bZ, où b est assuré par le fer chauffé au rouge, en Camargue, en Amérique du Nord et du Sud. D’une manière plus générale, van Gennep (1905, 1906) pense que les marques de propriété ont pu être à l’origine de l’écriture via les alphabets runiques et runiformes.
La dispersion des troupeaux sur les saltus extensifs exige parfois des instruments sonores pour les retrouver aisément, notamment des cloches métalliques ou de bois, à battant(s) interne (photo 15-35) ou externes (photo 15-36).


Les mouvements des troupeaux
La dépendance du troupeau vis-à-vis du Saltus étant très étroite, sa survie est conditionnée par son état ; il doit alors se déplacer vers des zones plus favorables lorsque la ressource s’épuise, d’où des déplacements qui peuvent avoir des amplitudes variables.
Le nomadisme est un déplacement de grande amplitude, tant géographique que sociale : tout le groupe humain accompagne le troupeau à la recherche de nouveaux pâturages (bouviers masaï et peuls, chameliers sahariens…). La transhumance est un mouvement plus localisé, ne concernant que le troupeau avec gardiens et chiens, non le groupe humain qui reste sédentarisé ; elle touche surtout les ovins et caprins, parfois les ruches à la poursuite des floraisons ; en dehors de la France où il est classique quoique modifié à l’ère moderne, il a par exemple été noté en Crète (de Foucault, 1993c) et en Afrique du Nord. L’estivage permet d’alterner entre des pâturages d’hiver et des pâturages d’été de plus haute altitude.
Ces déplacements sont marqués par la recherche de la chaleur et de l’eau, parfois du sel lorsque qu’il est nécessaire à la physiologie des animaux. On peut distinguer
- les mouvements horizontaux, le long de gradients latitudinaux ou longitudinaux, surtout associés au nomadisme : les rennes domestiqués des Saami entre la toundra en été et la taïga en hiver, ceux des Tchouktches entre les zones continentales et les zones côtières ; les nomades sahariens se déplacent entre les zones subtropicales en été et les zones méditerranéennes plus septentrionales pour bénéficier des pluies d’hiver ravivant la végétation ;
- les mouvements verticaux, le long de gradients altitudinaux, plutôt associés à la transhumance et l’estivage (il existe cependant un nomadisme vertical, en Anatolie, en Iran, en Afghanistan), là où des reliefs fortement marqués déterminent des contrastes climatiques ; ainsi, autour du bassin méditerranéen, les saisons sèches de plaine coïncident avec les saisons végétatives d’altitude, alors que les hivers altitudinaux correspondent à des saisons humides en plaine ; selon l’emplacement de l’habitat permanent, on sépare la transhumance directe ou normale, lorsque cet habitat est plutôt localisé à basse altitude et le troupeau se déplace vers le haut en fin de printemps (pour les détails, voir de Foucault, 1990a, tableau 1), et la transhumance inverse, lorsque cet habitat est plutôt localisé en altitude et le troupeau se déplace vers le bas en fin d’automne (pour les détails, voir de Foucault, 1990a, tableau 2 ; voir aussi Lachowski, 1988, pour une transhumance entre Champsaur et forêts du Var).
Lorsque l’habitat est localisé à moyenne altitude, on rencontre la transhumance double, vers le haut et vers le bas par exemple en Corse, en Afrique du Nord.
Ces déplacements se faisaient le long de chemins souvent très anciens dont la nomenclature varie selon les régions : drailles du Languedoc, calades des Cévennes (photo 15-37), carraïres de Provence, sentiers pavés des Pyrénées (nom local indéterminé, voir cependant encadré ci-dessous ; photo 15-38)…


Toutefois, la problématique de ces chemins pastoraux n’est peut-être pas aussi simple comme en témoignent ces éléments rassemblés par Catherine Brau-Nogué, Béatrice Morisson et Gérard Largier (Conservatoire botanique national des Pyrénées et de Midi-Pyrénées). Une dralha en occitan, au moins dans les dialectes languedocien, vivaro-alpin, gascon, provençal (on n’ a pas vérifié pour auvergnat et limousin), est tout simplement un chemin de campagne, voire un sentier ou une trace. C’est le terme qui a été retenu en français, francisé en draille, pour les chemins de transhumance, mais en réalité c’est un mot générique. En Provence, on parle de carrairas, francisé en carraires (Mistral dans le Trésor du félibrige : « carrairos : chemin ou passage lié aux troupeaux transhumants, chemin rural, ancienne voie »), mais, malgré le premier sens donné par Mistral, c’est plutôt générique également, car l’étymologie est la même que pour la rue (carrèra/charrèra en occitan). En Pyrénées catalanes, on parle des camins ramaders. Or en catalan, ramader signifie éleveur et ramaderia troupeau ; cela fait un terme non générique en langue originale par rapport à dralha. À voir s’il y a un terme dans l’Aude pyrénéenne, en domaine occitan. C. Brau-Nogué note que Cavaillès (voir infra) semble dire que, si la grande transhumance existait bien alors de et vers le grand Sud-Ouest, il n’y avait pas de chemins particulièrement dédiés aux troupeaux. Donc de tels termes pour désigner ces traces n’existent peut-être pas en Gascogne ou se sont effacés depuis longtemps ; elle pense qu’on trouve encore en Espagne (delta de l’Ebre en direction des Pyrénées ?) des chemins avec de grands troupeaux qui transhument toujours à pied. Ces chemins ont peut-être un nom.
Voici par ailleurs ce qu’écrit Jean-François Le Nail, historien, ancien directeur des Archives départementales des Hautes-Pyrénées, occitaniste et locuteur occitan :
« En ce qui concerne le lexique, le plus riche répertoire sur le mot dralha et ses frères et soeurs est donné par le Französiches Etymologisches Wörterbuch, t. 13 (2), 173-177, accessible par internet. Les dictionnaires gascons connaissent le mot sous la forme tralh, tralha, avec la signification principale de ‘trace’ (au sens de ‘marque de pas, piste’), d’où ‘sentier, chemin’ et même, sous les formes tragno, tragnère, probablement citée par Rondou à Palay, ‘sentier de vidange en montagne’ (couloir de vidange, glissoire). Le mot n’est pas connu du récent Dictionnaire gascon de la Haute-Bigorre (J.-L. Massourre, 2022). Quant aux chemins empruntés par la transhumance, estivale ou hivernale, reliant Pyrénées, Gascogne et pays d’Outre-Garonne, s’ils ont évidemment existé et s’ils sont connus par l’intermédiaire des péages, il ne semble pas qu’ils aient donné lieu à des définitions (techniques et juridiques) ni à des appellations spécifiques, comme les cabañeras aragonaises, longuement décrites et racontées dans le magnifique ouvrage de Severino Pallaruelo, Pastores del Pirineo (Madrid, 1988, p. 52-56, 74-89, 229). C’est en tout cas ce qui ressort de la thèse secondaire d’Henri Cavaillès (La transhumance pyrénéenne et la circulation des troupeaux dans les plaines de Gascogne, Paris, Armand Colin, 1931), travail qui n’a pas encore été remplacé.
Quant à moi, je ne crois pas avoir rencontré dans ma fréquentation des archives des xive-xvie siècles (de Campan, du Lavedan et de Barège) de terme spécifique pour désigner les chemins d’accès aux estives. Dans les actes de Barège fixant ces conditions pour les troupeaux aragonais arrivant par Gavarnie, ces itinéraires d’accès et de retour ne sont décrits qu’à travers la mention des aires de repos, des lieux de franchissement du Gave, de points caractéristiques du terrain, cols, carrefours, bois, etc. Et dans le Livre vert de Bénac (Cartulaire des vicomtes de Lavedan), la charte dite des Hauts-Pâturages (1384) ne mentionne les chemins des montagnes qu’à titre de limite à ne pas franchir (podge, 18, p. 113 ; escale deu Bazus, sober lo camin, 21, p. 113 ; darer la bia, dauant la bia, 24, p. 114 ; deu semder deu Bedot, 28, p. 115). Le sujet mériterait d’être approfondi, ainsi qu’une comparaison avec les usages des Cévennes, de la Provence, des Pyrénées orientales et du bas Languedoc, des Pyrénées ibériques… ».
En définitive, on observe une symétrie dans les déplacements entre la dimension horizontale (latitudinale et longitudinale) et la dimension verticale (altitudinale) comparable à celle que la phytogéographie a notée entre la zonation latitudinale et l’étagement de la végétation, nouvel exemple d’isomorphisme de variables (de Foucault, 2014a).
Les habitats humains temporaires
Ces multiples déplacements obligent les hommes à occuper des habitats originaux, soit dans leur structure, soit dans leur occupation. Dans le cas du nomadisme, ces habitations sont mobiles, faciles à démonter, à transporter, à remonter ; c’est la tente traditionnelle (photo 15-39), dont la yourte d’Asie centrale.

Dans le cas de la transhumance et de l’estivage, les habitats temporaires sont plutôt fixes et seulement temporairement occupés, à la saison du pâturage. Ils sont de petite taille, car seules quelques personnes accompagnent le troupeau. Leur diversité et leur nomenclature sont grandes ; on en a déjà évoqué en 10.3, auxquels on peut encore ajouter les burons d’Auvergne, les chalets (au sens initial du terme) des Alpes, les cayolar basques. Ils utilisent des matériaux locaux, surtout la pierre calcaire ou volcanique, le bois. Plus rarement, l’habitat est mobile quoique hors nomadisme (photo 15-40 ; voir aussi pour le Népal Toffin et al., 1986 : 116). Fonctionnellement parlant, un ensemble d’habitats dispersés dans l’espace est équivalent à un habitat se déplaçant au cours du temps : « Le déplacement d’un corps est équivalent à une pluralité de corps diversement placés dans un espace. » (Valéry, Cahiers I : 550). Jean-Brunhes Delamare (1985 : 162) rapporte plusieurs photographies de tels habitats.

Parfois les animaux élevés sont abrités dans des habitats pour les protéger des prédateurs. La photo 15-41 montre le cas d’un abri sous roche dont l’ouverture externe est partiellement obturée par un muret de pierres : Σ(G, R).

Les produits animaux
Les animaux domestiqués (DZ noté ci-dessous Z1) sont élevés pour apporter aux H des produits divers. Pour ordonner cette diversité, on peut s’appuyer sur les opérateurs relationnels.
Les faits du type {1Z1, F’} concernent les animaux comme source d’énergie (voir § 13.2) ; on a aussi le cas des loisirs, comme les courses de dromadaires.
Les faits du type {eZ1, F’} concernent des parties de l’animal vivant pour les transformer. C’est le cas de la laine qui est recueillie, préparée puis tissée, soit {vpeZ1, F’}, pour créer des vêtements (la laine des moutons de race manech permet de fabriquer le béret basque), des abris… (photo 15-42). La corne peut servir à élaborer des instruments de musique (photos 11-15 et 11-16).

Les faits du type {iZ1, F’} concernent des produits extraits de l’animal vivant en vue de l’alimentation humaine. Le plus important au monde est sans conteste le lait dont la fonction primaire est de permettre la croissance du petit animal mis-bas (veau, chevreau, poulain…). L’opérateur i est mis en œuvre par la traite (photo 15-43) ; chez certains pasteurs, le contact de ce produit avec le métal ou l’argile est prohibé, d’où des faits e.botaniques pour des récipients végétaux. Le lait peut être bu directement (iZ1) ou chauffé (ciZ1). Il peut parfois être conservé dans de petites constructions minérales à moitié enfouies (la photo 15-44 illustre la gratte des Alpes), mais c’est un produit fragile qui s’altère rapidement ; pour le stabiliser, on doit le transformer en produits dérivés qui pourront être conservés plus longtemps. C’est un produit biochimiquement complexe et on peut distinguer deux grands types de produits selon l’invariant laissé par la transformation :
- en conservant la partie lipidique, on prépare la crème (obtenue par concentration des globules gras), puis le beurre (obtenu par barattage de la crème, éventuellement dans une baratte entraînée par énergie hydraulique ; Sélection du Reader’s Digest, 1978 : 568) ; leur F’ est surtout alimentaire, mais des ethnies d’Afrique du Sud et de Namibie en font un onguent et un produit protecteur ;
- en conservant la partie protéique (caséine), qui coagule par acidification (fermentation lactique transformant le lactose en acide lactique) ou par intervention enzymatique (présure formée de chymosine et pepsine, produit extrait de la caillette du veau, autre fait e.zoologique), donnant le caillé; on peut ensuite préparer les yoghourts, des fromages mous ou durs, le kéfyr issu d’une fermentation lactique et alcoolique… (photo 15-45) ; dans les régions de transhumance et estivage, cette activité est un des travaux importants des gardiens de troupeau, de là la diversité des types de fromages selon les grandes régions de saltus ; le complément du caillé dans cette préparation est le petit-lait, parfois utilisé pour nourrir des animaux autres que ceux du Saltus (par exemple porcs associés aux burons d’Auvergne).



Plus rarement (Masaï, Bantous…), c’est le sang de Z1 qui est consommé, extrait au niveau du cou de l’animal. Des produits animaux moins nobles sont diversement utilisés (déjections, photo 13-10 ; matériau pour les habitations, photos 11-11 et 11-12 ; urine pour oindre les cheveux chez les Masaï, qui l’utilisent aussi pour laver la vaisselle devant contenir le lait, car ce produit noble ne doit pas être souillé par un contact avec l’eau). L’hydromel est une boisson issue du miel des abeilles.
Dans les faits du type {mZ1, F’}, les animaux sont abattus pour que certaines de leurs parties soient associées à des F’. Ainsi, les peaux peuvent servir de vêtements, d’abris. Elles peuvent entrer dans la préparation de gourdes (en Pays basque, la gourde navarraise est en peau de mouton manech) ou d’outres (photo 15-46). Mais c’est évidemment surtout la viande qui est recherchée pour la nourriture après préparation (bovins, ovins, rennes). Parfois l’animal tué a une fonction sacrée (chèvre au Népal) ou ludique (le bozkachi est un jeu au cours duquel des cavaliers cherchent à s’emparer d’une chèvre décapitée en Afghanistan ; photo 15-47).


La dynamique du Saltus
Tout ce qui vient d’être écrit correspond à la civilisation traditionnelle du Saltus. De nos jours, plusieurs de ces faits sont soumis à des transformations diverses, agents de dynamique aux effets plus ou moins heureux.
Les faits e.phytosociologiques
Il faut d’abord se rappeler que, au contraire du Saltus I, le Saltus II dérive de la régression d’une forêt ; c’est donc un stade instable que la dynamique progressive peut ramener au stade forestier dès que la pression biotique tend à diminuer ; l’éleveur cherche donc à stabiliser ce stade, notamment en éliminant les arbustes et les jeunes arbres qui préparent le retour de la forêt. En cas de déprise pastorale, des organismes tels que les conservatoires d’espaces naturels engagent des mesures de gestion pour stabiliser les stades secondaires souvent de plus grande richesse biologique que les stades forestiers, notamment en mettant en œuvre les méthodes du génie écologique (de Foucault, 1988c), associant éventuellement des races animales rustiques (Lecomte et al., 1981 ; Maubert, 1988 ; Manneville, 1988).
Pour valoriser autrement les saltus délaissé, H peut planter des arbres ; c’est un retour à une forme de Silva selon une dynamique progressive provoquée et plus ou moins accélérée. Une silva est recréée si les essences plantées sont choisies de façon à reformer une strate arborescente représentative de la forêt locale, selon la végétation potentielle naturelle ; c’est plutôt un ager ligneux qui est recréé en cas de monoculture d’une essence étrangère à la végétation potentielle locale (photo 15-50).

Les faits e.zoologiques
Au niveau e.zoologique, on a déjà évoqué la dynamique du chien de troupeau devenant assistant du gardien, puis animal de compagnie : {1Z1, protéger} → {1Z1, aider} → {1Z1, accompagner}, dynamique à 1Z1 invariant (photo 15-34). Au niveau des déplacements de troupeau, la sédentarisation réduit le nomadisme à la transhumance. Les sentiers de déplacements des troupeaux (drailles, calades…) sont souvent repris actuellement comme sentiers de randonnée.
Suite à la déprise de la transhumance et de l’estivage, les habitats humains temporairement occupés perdent leur F’. S’ils ne sont pas réaffectés à une autre fonction, ils se dégradent progressivement jusqu’à la ruine complète : dynamique fonctionnelle F’ → 0 traduisant la désuétude. Certains heureusement sont conservés ou légèrement restaurés pour une autre F’ (donc structure architecturale plus ou moins invariante) : habitations secondaires pour vacanciers (chalets des Alpes), éléments de musées de la vie rurale traditionnelle (ethnomusées) pour témoigner de la vie passée (burons d’Auvergne), auberges et refuges de montagne.
Cette dynamique peut toucher enfin la fonction des Z1 élevés. Ainsi le cheval, initialement chassé pour sa viande, a pu être domestiqué pour le trait ou le combat et est aujourd’hui associé aux loisirs et la compagnie. Le Saltus actuel est moins d’autoconsommation que de rémunération pour l’éleveur, directe (circuits courts) ou via des intermédiaires qui transforment ses produits en dérivés pour des consommateurs sans lien direct avec les troupeaux. Ainsi les fromages fabriqués artisanalement à l’estive sont peu à peu remplacés par des fromages fabriqués industriellement (gruyère, beaufort…).
14.3. L’Ager
Troisième volet de la trilogie après Silva et Saltus, l’Ager désigne un peuplement végétal artificiel créé à la place d’une V spontanée à laquelle il se substitue ; l’agriculteur y remplace le chasseur-cueilleur-pêcheur et le pasteur.
Création de l’Ager
Dans cette première étape, il s’agit d’une part de débarrasser la terre convoitée de la V spontanée qui l’occupe, d’autre part d’enrichir cette terre en éléments nutritifs pour les futures plantations.
La première intervention est assurée par le défrichement de V (forêts, landes, pelouses) puis le retournement de la terre (tG) jusqu’au stade de substrat quasi dénudé. S’y rattache l’essartage, les clairières acquises étant dénomées « essarts », « esserts », « sarts », termes qui ont laissé des traces dans la toponymie actuelle ; en Guyane française, on parle plutôt d’abattis (Gély, 1986).
Une double opération permet de débarasser le sol et d’apporter des substances nutritives ; elle met en jeu un agent écologique hautement énergétique, le feu, qui passe sur la végétation abattue et séchée, et minéralise la matière organique végétale, permettant d’établir des cultures sur brûlis (figure 18 et photo 15-48), les cendres résiduelles fertilisant la terre ainsi débarrassée : {bdeV, permettre la culture}. Parfois, des arbres sont épargnés, soit pour des raisons culturelles, soit pour des raisons culturales, par exemple comme futurs supports de plantes alimentaires grimpantes (Dioscorea ou ignames, Piper ou poivriers) ou pour protéger du soleil des plantes cultivées sensibles (cacaoyères et caféières ; Portères, 1949). Dans certains systèmes agraires, on n’abat pas avant le brûlis, mais on laisse courir le feu, surtout pour déblayer rapidement le terrain, soit {bV, permettre de cultiver} ; c’était notamment le cas en Tarentaise dans le système dit de Montvalesan, où le feu est difficile à maîtriser : sous l’action du vent, il peut sortir des limites désirées et la végétation incendiée en dehors des limites forme l’usclade. De telles parcelles cultivées après brûlis sont fréquentes dans les agricultures tropicales où elles sont diversement dénommées : ray, ladang, jhum, caingin… en Asie (Hubert-Schoumann, 1974), milpa, conuco, chacra en Amérique, lougan, tavy en Afrique occidentale ; von Verschuer (1995) rapporte de nombreux termes pour les cultures sur brûlis japonaises.


Une fois ces opérations réalisées, l’agriculteur peut entretenir sa parcelle (semis, hersage… ; photo 15-49).

Amélioration de l’Ager
Lorsque les éléments minéraux apportés par le brûlis sont insuffisants, une fertilisation complémentaire (g) peut être délivrée, reliant parfois Saltus et Ager. Ainsi, la litière acquise dans l’exploitation des saltus et enrichie par les déjections des Z peut être épandue sur les agers. L’écobuage consister à peler la terre (écobuer), à sécher puis brûler le produit ainsi recueilli, c’est donc une minéralisation accélérée (Portères, 1972). Le soutrage est une pratique voisine propre au sud-ouest de la France qui consiste à enlever la terre de bruyère et les plantes qu’elle porte pour récupérer le fumier qui y était formé par les troupeaux ovins. La pratique du mafuku en Afrique s’y rattache aussi (Kuhnholtz-Lordat, 1958 : 129). D’autres éléments fertilisants peuvent être acquis par des pratiques e.phytosociologiques (brûlage de branches issues de haies) ou e.zoologiques (parcours des troupeaux sur les agers qui vont bénéficier des déjections, colombine des pigeonniers du Périgord, guano).
L’élévation du pH des terres acides fait appel à l’e.minéralogie, à travers le falunage (en Touraine) et le chaulage (cCalcaire dans des fours à chaux), et l’e.phytosociologie à nouveau (maerl issu de communautés algales incrustantes).
Physiologie artificielle de l’Ager
Pour une croissance optimale des végétaux cultivés, il faut parfois que l’agriculteur recrée des conditions culturales rappelant leur écologie naturelle.
L’ensemble des facteurs écologiques naturels inclut d’abord des données climatiques, notamment des températures suffisantes, qui peuvent être acquises par des techniques artificielles : serres, châssis et cloches, lutte contre le gel chez les fruitiers et la vigne au moyen de brûleurs, de nuages artificiels pour diminuer le rayonnement nocturne terrestre… ; inversement, le gel lève la dormance des bourgeons et de certaines semences.
Pour les facteurs hydriques, les conditions naturelles limitantes sont suppléées par le drainage pour évacuer l’excès d’eau dans la parcelle cultivée (culture sur planches surélevées), l’irrigation dans les régions arides (oasis ;
photos 15-50 et 15-51) ou à saison sèche marquée, l’arrosage, qui peut être associé à un puits (photo 15-52). On connaît bien le cas du riz qui est parfois planté en terrasses complexes irriguées dans des reliefs qui ont été en quelque sorte comme sculptés par les riziculteurs ({tG} ; photo 15-53). Certains systèmes d’irrigation ont été labellisés Patrimoine mondial de l’humanité par l’Unesco (dont les cinq systèmes aflaj d’Oman).



La protection contre le soleil direct (facteur photique) peut être assurée par l’installation de claies protectrices végétales supérieures, donc plutôt arbustives, voire arborescentes ; c’est nécessaire pour la culture des Anthurium scherzerianum en Guadeloupe, souvent sous bananiers (photo 15-54), des Theobroma ou Coffea.

Dans les régions soumises à des vents intenses (surtout le mistral), des coupe-vent en réduisent les effets, souvent réalisés en Cupressus (photo 15-55).

La physiologie artificielle inclut aussi l’usage de la multiplication végétative, de la pollinisation artificielle, de moyens empiriques comme l’éthologie artificielle (tuteurs, supports biologiques), des rituels a priori irrationnels : périodes d’intervention (calendrier lunaire, e.astronomie), protection contre le mauvais œil (photo 15-56)… Le conjurador est un petit bâtiment où l’on lisait les Évangiles aux quatre vents pour écarter des nuages inquiétants amenant la grêle dans le Roussillon (Sélection du Reader’s Digest, 1978 : 545). On peut en rapprocher les rogations, cérémonies chrétiennes du printemps ayant pour but d’attirer les bénédictions divines sur les champs et les récoltes à venir.

La qualité édaphique des agers est assurée par une modification des caractères physico-chimiques du sol : aération, épierrement, amendements minéraux ou organiques (terre de bruyère pour les végétaux qui ne supportent pas le calcaire), structure et texture (apport de sable ou au contraire d’argile), fertilisation, poldérisation. Le labour permet de remonter à la surface des horizons enfouis et d’aérer le sol ; à cela se rattachent les araires (photo 15-57) et surtout les charrues qui ont fait l’objet d’une vaste monographie d’envergure planétaire (Haudricourt & Jean-Bruhnes-Delamare, 1955 réédité 1986) ; pour les araires du Népal, voir Blamont (1986 : 149).
Les facteurs biotiques sont représentés par la protection des cultures, en luttant contre les « nuisibles » (parasites, prédateurs, « mauvaises herbes ») : dans une étude antérieure (de Foucault, 1997c, avec formalisation par les opérateurs e.scienifiques et nombreux exemples), on a distingué la lutte mécanique, la lutte physique, la lutte chimique, la lutte biochimique, la lutte culturale, la lutte éthologique, la lutte génétique, la lutte biologique, la lutte intégrée. Dans une émission radiophonique récente, un agriculteur a indiqué une méthode de lutte contre les dégats des sangliers : déposer des poils de chien sur les cultures pour éloigner les suidés : {ΣepoilsChien, -1Sanglier}.

Espèces cultivées et exploitation de l’Ager
Les cultures installées sur l’Ager ne sont pas associées à des faits e.phytosiociologiques, mais seulement e.botaniques où F’ est nourrir H (cultures vivrières), des Z domestiqués (cultures fourragères) ou non (cultures pour la chasse) ou autres (cutures industrielles). Ces taxons cultivés sont des espèces spontanés autochtones, toutefois sorties de leur association végétale naturelle, ou plus souvent des espèces transformées par la culture ou l’amélioration génétique (gP) pour une plus grande production, un meilleur développement dans des conditions écologiques inhabituelles, une meilleure résistance aux nuisibles, une meilleure utilisation. Dans certains cas, leur ancêtre sauvage est inconnu ; il semble bien que des mauvaises herbes annuelles commensales de culture (Avena sativa, Secale cereale dérivé du vivace S. montanum) ont pu être domestiquées en céréales alimentaires : {1P, 0} → {DP, F’} (Haudricourt, 1939). Il n’est pas utile d’insister ici sur la diversité des végétaux cultivés tant elle est en grande partie connue de tous. Plusieurs publications traitent de l’origine des végétaux cultivés depuis au moins A. de Candolle (de Candolle, 1883 ; Haudricourt & Hédin, 1943…).
Les dehesas (photo 15-58), habitats surtout ibériques inscrits à l’annexe 1 de la directive européenne Habitats-Faune-Flore (Eur 27 6310), associent des canopées de chênes sempervirents méditerranéens, des pâturages (Saltus) et des cutures (Ager) ; c’est le seul habitat à connotation nettement e.phytosociologique d’intérêt communautaire.

Aux agers ligneux se rattachent notamment les vergers fruitiers (photos 15-59 et 15-60), les plantations de thé (Camellia sinensis ; photo 15-61), de kat (Catha edulis), de maté (Ilex paraguariensis ; photo 15-62), les houblonnières (photo 15-63), les chènevières ou cannebières, les vignobles (photo 15-64), les pépinières (agers de jeunes plants en vue d’un reboisement ornemental ou productif)… et surtout les agroforêts.






Les agroforêts (photo 15-65) consistent en général en des systèmes forestiers complexes conduits par les hommes, avec une structure multistrate de la végétation (Gautier, 1994 ; Pasquis, 1998), un grand nombre de composantes végétales (arbres, arbustes, lianes, herbacées) diversement utiles (plantes alimentaires, épices, bois d’œuvre…), avec un fonctionnement écologique similaire à celui des forêts naturelles. De par leur structure multi-étagée, leur faciès forestier et leur composition floristique, les agroforêts détiennent une grande diversité biologique. Décrivant des agroforêts destinées à la récolte de résine de damar (Shorea javanica, Dipterocarpaceae), Michon et al. (1995) leur reconnaissent plusieurs fonctions essentielles : un « …mode de conduite particulier de l’agroforêt, qui tire parti des dynamiques naturelles de production et de reproduction biologiques afin de réduire la contribution du facteur économique le plus rare, le travail » (p. 213-214), « conservation […] des processus biologiques déterminants pour la survie à long terme de l’agroforêt. Ce sont ces relations qui se nouent entre sol et plantes, entre plantes et animaux qui permettent le maintien à peu de frais de l’agroforêt » (p. 214-215), « grâce à la reconstitution d’un niveau élevé de biodiversité permettant de reproduire la ressource forêt dans son ensemble, l’agroforêt a pu reprendre le rôle traditionnellement dévolu aux forêts naturelles, celui d’un espace à utilisation multiple, ouvert pour la cueillette et pour la collecte commerciale » (p. 215-216), « Cette stratégie d’appropriation ne permet pas de conserver l’intégralité de l’écosystème naturel : la forêt climacique est largement détruite et l’agroforêt contribue pour une grande part à cette destruction. Mais elle permet la restauration de la plupart des ressources de l’écosystème de départ au sein d’une structure artificielle mais biologiquement forestière. » (p. 218).

Les produits de l’Ager
Beaucoup des productions de l’Ager sont des plantes à destinée alimentaire. Notamment, une fois mûres, les céréales sont coupées et le grain (plus précisément le caryopse chez les Poaceae) est détaché (ΔP) de la balle (reste des glumes et glumelles desséchées chez les Poaceae) par des Z qui piétinent la récolte, par un tribulum (planche de bois armée de silex taillés, soit {Σ(teP, tR)} ; photo 15-66 ; le trillo en est l’équivalent espagnol) tracté, des fléaux (dépiquage, chaubage ou battage). L’ensemble grain + balle est ensuite séparé en ses deux composantes par le vannage, qui consiste traditionnellement à le jeter dans un courant d’air (en pratique le vent, e.météorologie) écartant la balle plus légère, qui pourra être récupérée pour d’autres F’ (comme le bourrage de matelas), laissant retomber à terre le grain plus lourd (photo 15-67 ; voir aussi Baumann, 1984 :143) ; des vans mécaniques (tarares, batteuses ; photo 15-69) peuvent faciliter cette opération assez fastidieuse, alors que les moissonneuses-batteuses modernes regroupent les diverses étapes de la moisson, de la coupe au vannage. Des cas de silicose ont été récemment mis en évidence en lien avec de telles pratiques, analogue à la silicose des mineurs, la balle étant composée presque uniquement de silice quand elle est sèche (c’est le cas par exemple en Camargue, sur des ouvriers des rizières ; P. Coulot, courriel en date du 15 décembre 2022).



Des charettes de diverses formes facilitent le transport des produits des champs ; Jean-Brunhes Delamare (1985) en présente plusieurs modèles ; certaines sont hautement ornementées (Sicile, carretas du Costa Rica ; photo 15-68).

Si les produits moissonnés utiles ne sont pas consommés aussitôt, il faut prévoir leur conservation à l’abri des prédateurs (insectes, rongeurs), mais aussi parfois des voleurs. Elle est assurée par des greniers, des silos, des granges (qui peuvent accueillir aussi du foin, produit du Saltus ; photos 15-70 et 15-71), celles-ci parfois réduites à un arbre isolé protecteur (« arbre grange » ; photo 15-72)… ; en Tunisie méridionale, on peut encore observer des ksour (au singulier ksar), ensembles de greniers ou ghorfa plus ou moins fortifiés qui ont eu cette fonction, quelque peu abandonnés de nos jours (de Foucault & Claisse, 1994 ; photo 15-73). Au Pérou, des agriculteurs des environs de Cuzco ont remis en honneur des greniers incas appelés qollqas formés de murets de pierre et recouverts d’une toiture de q’olla (chaume d’une graminée locale, peut-être Stipa ichu) (Liendo Tagle, 2021).




On peut rapprocher de ces greniers quelques abris végétaux associés aux agers, par exemple aux rizières en Asie tropicale (photo 15-74).

Pour pouvoir être préparés en vue d’être consommés, ces produits ont souvent besoin d’être coupés en petits morceaux ou écrasés (ΔP) ; cette destructuration passe par exemple par des meules artisanales (photo 15-75), des moulins à vent (e.météorologie ; photo 13-24) ou à eau (e.minéralogie ; photo 13-20). Avant l’extraction de l’huile ou du jus sucré, les olives et les pommes sont écrasées dans un pressoir (ou gattage dans le cas des pommes en Normandie) ; pour les olives en Afrique du Nord, traditionnellement, ce sont les dromadaires qui apportent l’énergie nécessaire à cette opération (photo 15-76) et les olives écrasées sont recueillies dans des vanneries d’alfa (veMacrochloa tenacissima) en vue du pressage.


On ne rentrera évidemment pas ici dans le détail de la préparation des aliments par la cuisine, que l’on illustrera simplement par la taguella (ou taghella) du Hoggar, une galette préparé sans levain à partir de farine et d’eau, cuite dans la cendre chaude du sable désertique (e. minéralogie) puis rincée à l’eau (pour éviter les éventuels grains de sable qui pourraient gêner la consommation) avant d’être partagée entre les convives (photos 15-77 et 15-78).


Les déplacements de l’Ager
Les espèces cultivées, plus ou moins exigeantes au plan trophique, absorbent progressivement les réserves minérales du sol et l’exploitation n’en restitue qu’une faible partie. Le sol agricole finit donc par s’épuiser et n’être plus à même d’assurer la nutrition de nouvelles cultures. L’agriculteur est alors obligé d’abandonner la parcelle épuisée, laissée en jachère, pour recréer un nouvel ager à quelque distance.
Délaissée, la parcelle est reprise par la dynamique végétale progressive menant jusqu’à une V plus évoluée qui peut reconstituer le stock nutritif du sol après un laps de temps suffisant. Dans la publication de 1990a, j’ai décrit plusieurs cycles culturaux. On a notamment mis en évidence un isomorphisme entre un cycle associant un saltus dérivant vers un ager par la culture, lequel y retourne après abandon de la parcelle, et un cycle associé à la gestion des étangs en Brenne, Woevre, Bresse comtoise, le saltus correspondant en un élevage de poissons, l’ager correspondant à la mise en assec de l’étang pour y semer des céréales, avant la remise en eau.
La dynamique de l’Ager
Le principal aspect dynamique de l’Ager est lié à la sédentarisation et à la fixation du champ par suppression de la jachère. Ceci n’est possible que si l’agriculteur apporte à la parcelle des éléments nutritifs et, localement, l’eau : « c’est l’engrais qui fixe le champ » (Lebeau, 1979).
Comme pour le Saltus, l’Ager d’autoconsommation peut être remplacé par l’Ager rémunérateur, avec développement d’une chaîne de transformations aboutissant à un consommateur.
L’Hortus
L’Hortus (photo 15-79) est un ager plus domestique, entourant souvent la maison (ou domus), géré par la main d’œuvre familiale et réservé à l’autoconsommation. Il prend la forme du jardin, du jardin de case (Tchatat et al., 1995), du chinampa (cultures sur radeau) des Indiens du Mexique (photo 15-80), du pekarangan à Java (Christanty, 1981). Les P y possèdent des fonctions diverses : potagères, condimentaires, fruitières, médicinales, ornementales, protectrices des cultures (par exemple contre les nématodes ; Bourdy et al., 1995), à boisson (pour le vin de palme par exemple, pieP ou piP selon que P est abattue ou incisée sur pied ; Tchatat et al., 1995 ; photo 15-81), à usages divers. Il peut être enrichi par les détritus issus des activités de la domus : cendres, compost…



On doit mettre un peu à part certains jardins chinois et japonais. Ces derniers (photo 15-82) sont formés par des assemblages complexes de végétaux ligneux à feuillage persistant, de pierres (montagnes artificielles en réduction,
ΣR = hG), de petits cours d’eau, le tout formant un abrégé du monde, mais pas tout à fait une copie de la nature, car il tente d’en donner les sensations et une interprétation subjective, d’offrir un cadre propice à la méditation religieuse et philosophique ; ils sont souvent associés à la cérémonie ritualisée du thé. Les jardins de lettrés chinois (Métailié, 1995) assemblent eux-mêmes de telles formes, mais associées à une salle de lecture, une bibliothèque, matérialisant la coupure avec le monde public, ce qui n’empêche pas de tirer un profit économique des plantes qui y sont cultivées. La miniaturisation du monde va de pair avec celle de certains végétaux dans la réalisation des bonsaï japonais (photo 15-83) ou des penjing chinois.


On peut considérer l’ensemble des petits élevages de basse-cour (poules, lapins, canards, pintades, oies, pigeons… ; photo 15-84) comme un hortus animal, dont les produits sont autoconsommés, pouvant être eux-mêmes nourris par les restes issus des activités de la domus. Parfois les petits animaux sont rassemblés par catégorie dans des espaces dédiés : poulaillers, clapiers, pigeonniers (photo 15-87). Cet hortus peut d’ailleurs parfois entrer en conflit avec l’hortus végétal lorsque les animaux s’en échappent…


Comme l’Ager, l’Hortus d’autoconsommation peut évoluer vers l’Hortus rémunérateur, ainsi les hortillonnages de cultures maraîchères (région d’Amiens ; Marais audomarois autour de Saint-Omer, Pas-de-Calais ; photos 15-85 et 15-86), les huertas méridionales, les cultures de plantes médicinales en grand (en France, Chemillé, Milly-la-Forêt). De même l’Hortus animal peut évoluer vers des élevages en grand, voire industriels, cuniculicoles, aviaires…


On peut encore aller au-delà de l’Hortus en définissant l’ensemble des E (végétaux et animaux) ornant l’intérieur de la domus ou accompagnant ses habitants : P d’orgine tropicale ou australe (Orchidaceae, Bromeliaceae, Arecaceae, Araceae, Gesneriaceae, Acanthaceae, Commelinaceae, Urticaceae, Begonia, Schefflera, Monstera et Philodendron, Impatiens, Ficus, Cyperus alternifolius…) pour les végétaux, chien (Canis lupus familiaris), chat (Felis silvestris catus), poissons (souvent le poisson rouge Carassius auratus), tortue (souvent de Floride, Trachemys scripta elegans), oiseaux (comme le serin Serinus canaria), petits mammifères (cochon d’Inde Cavia porcellus, voisin du cobaye C. aperea des Andes, ce dernier relevant souvent plutôt de l’hortus animal d’autoconsommation). De petits terrariums, vivariums ou aquariums contribuent à la culture des plantes ou à l’élevage de certains petits animaux. L’hygrométrie atmosphérique nécessaire en appartement pour des végétaux originaires de forêts ombrophiles (rain forests) ou néphéliphiles (cloud forests) peut être apportée par la brumisation, le bassinage des feuilles ou encore leur placement en fonction des microclimats de la domus (les plantes aérohygrophiles seront bien placées dans une salle de bain, les fougères dans des parties où la lumière est plus limitée). L’association d’Orchidaceae et d’Apocynaceae d’un côté, de Bromeliaceae d’un autre vise inconsciemment à créer des parties des structures végétales systématiques définies par ailleurs, les S(Orchidaceae-Bromeliaceae) ou les S(Apocynaceae-Orchidaceae) [de Foucault, 1994b, les secondes sub S(Asclepiadaceae-Orchidaceae), mais la première famille éponyme est maintenant incluse dans les Apocynaceae]. D’une manière générale, ces assemblages végétaux reconstituent dans la sphère domestique des associations végétales des sous-bois tropicaux telles que celles analysées par Blanc (2002). À défaut de nom consacré antérieur et en attendant mieux, on peut proposer de dénommer intradomus ces agers domestiques très particuliers.
Les P des intradomus font l’objet de soins intensifs, individualisés au pied (arrosage, fertilisation), qui se matérialisent au niveau de pots de fleur souvent en poterie (photo 15-88), quelquefois en plus entourés d’un cache-pot pour en améliorer l’esthétique, mais parfois aussi d’origine végétale (ΔeCocos nucifera en photos 15-89 et 15-90 ; aux Antilles, on peut trouver des pots réalisés à partir de stipes de fougères arborescentes creusés, teFougère) ; évidemment, on laisse ici de côté les pots en plastique. Ces pots facilitent en outre les déplacements de ces végétaux selon les saisons. Les sols y sont simplifiés, confinés, non structurés, et peuvent être rattachés aux anthroposols (Baize & Girard, 1992). En Asie du Sud-est, on observe assez souvent des plantes aquatiques en pot (photo 15-91), les sols étant alors submergés et se rapprochant des gyttja.




Dans les régions tempérées, le microclimat de la domus est de type tempéré chaud à subtropical ; il rappelle que l’origine des hommes se situe en Afrique tropicale et que les Homo ont dû développer le chauffage pour coloniser des régions plus hostiles, traditionnellement le feu. D’ailleurs, dans les régions tropicales, on observe que l’intradomus sort de la maison pour s’étendre sur les terrasses adjacentes ou en limite de la domus (photo 15-92).
La participation des chiens à l’intradomus animal semble assez limitée aux régions tempérées ; dans les régions plus chaudes, les chiens apparaissent plutôt comme commensaux des hommes que véritables compagnons ; ils y sont même parfois consommés, ce qui est impensable dans les pays du Nord.

15.4. La phytosociologie des associations végétales artificielles
Alors que Silva et Saltus portent sur des V spontanées (communautés végétales spontanées, CVS), l’Ager concerne des espaces cultivés, où sont introduites et soignées des espèces souvent étrangères à la région de culture. Pourtant, associées, elles semblent constituter des V, mais alors non spontanées (communautés végétales artificielles, CVA). Jusqu’à quel point ces CVA sont-elles comparables dans leurs propriétés, description, classification, nomenclature et dynamique éventuelles aux CVS ? En 1996, j’ai consacré une publication à ce sujet dont les grandes lignes sont reprises ici, en parallèle complet avec la phytosociologie des CVS. Wattez et de Foucault (2018) ont proposé d’utiliser cette méthode pour décrire la végétation ornementale des cimetières militaires issus de la Première Guerre mondiale, en particulier britanniques, les plus fleuris.
La flore cultivée des agers
Les CVA rassemblent des plantes généralement étrangères à la zone de culture, parfois génétiquement améliorées (variétés, hybrides, cultivars) ; à la base de cette réflexion, il y a donc la notion de taxon cultivé (TC) à double connotation, biologique et ethnologique, étant doté d’une fonction secondaire par H, taxon donc généralement associé à des faits e-botaniques. Ainsi, un récolteur de plantes P dans une communauté végétale V présente une relation e-phytosociologique avec V, puis, au sein de V, une relation e-botanique avec la ou les P récoltées pour des opérateurs et une fonction secondaire à préciser selon le récolteur, soit le passage
{1 ou eV, accueillir P} → {aP, F’}.
Pour les jardiniers et les agriculteurs, ces TC entrent dans des catégories souvent assez éloignées de celles de la taxonomie scientifique ; on peut ainsi distinguer les plantes ornementales, les arbres fruitiers, les céréales (des Poaceae à caryopse alimentaire, mais aussi le sarrasin, Fagopyrum esculentum, le quinoa, Chenopodium quinoa).
Notion d’individu d’association artificielle (IAA)
En associant délibérement divers TC, les acteurs de l’Ager créent donc ce qu’on peut appeler un individu d’association artificielle (IAA) par opposition à un individu d’association spontanée (IAS), objet de la phytosociologie classique. D’un point de vue formel, le passage de P isolés à un IAA peut s’exprimer par une formule exprimant le passage de l’e-botanique à l’e-phytosociologie : {ΣiPi, F’} → {aV, F’}.
Quels peuvent être les critères d’homogénéité d’un IAA ? Sans aucun doute une certaine isotropie ou homogénéité floristique, encore que, dans ce cas, cette notion soit bien subtile (cf. infra, § suivant). L’introduction de F’ dans les formules montre que l’IAA doit être aussi homogène du point de vue fonctionnel, donc posséder une fonction à peu près uniforme pour les H : ornementale, médicinale, sylvicole… On retrouve ici le concept de cellule paysagère isofonctionnelle explicité notamment par Géhu (1991), un élément visuellement marquant et caractéristique d’un paysage, tel un openfield labouré indépendamment de la rotation des cultures, élément par ailleurs caractérisé par sa F’ socio-économique.
À la différence des IAS, qui s’avèrent la plupart du temps des ensembles aux limites floues (de Foucault, 1981), les limites des IAA sont souvent nettes (presque comme « tirées au cordeau »), ils sont dotés de formes géométriques plus ou moins régulières (rectangulaires ou courbes) ; Bruneton-Governatori (1986) remarque que la géométrie de jardins potagers pyrénéens exprime consciemment ou non une notion d’ordre. La géométrie des jardins d’ornement permet de distinguer des styles dits à la française, à l’anglaise, à l’italienne. On note encore une sociabilité élevée des TC présents, ce qui peut compliquer l’appréciation du critère d’homogénéité floristique. La structure (pattern) montre un certain alignement des TC ; c’est là une des différences relativement aux IAS qui permettent souvent de distinguer ceux-ci dans les paysages. Elle est liée à une meilleure efficacité de la culture et de son exploitation, une plus grande efficacité du travail agricole (semis, nettoyage, traitements, récolte).
Comme dans les IAS, les IAA peuvent présenter une stratification, faisant de ceux-ci des phytocénoses artificielles, par exemple les agroforêts (photo 15-65), qui permettent de conserver la grande diversité biologique, la stratification des forêts spontanées et, comme ces dernières, de protéger les sols, les climats, la ressource en eau, le patrimoine génétique : « l’agroforêt doit être considérée comme la manifestation la plus achevée d’une tendance générale, typiquement tropicale, qui consiste à associer entre elles des plantes utiles de dimensions diverses » (Hallé, 1993).
Par ailleurs, les IAA peuvent entrer en contact avec les IAS, par exemple IAS des synusies inférieures pour des agers arborescents, IAA cultivés et IAS formés des taxons commensaux de l’ager (céréales et messicoles).
Comme les IAS, la structure des IAA est aussi déterminée par la représentation des formes biologiques élémentaires : arbres, arbustes, herbes vivaces, herbes annuelles notamment. On peut donc transposer à l’e-phytosociologie les formations végétales en y associant leur fonction ethnologique et parler, par exemple, de mégaphorbiaie fruitière pour une bananeraie. À l’appui de cette interprétation, on peut évoquer la description d’une mégaphorbiaie tropicale au Togo (de Foucault et al., 2011), qui a donné lieu à des comparaisons ayant mis en évidence le rôle des Monocotylédones zoogames dans diverses mégaphorbiaies tropicales (Marantaceae, Commelinaceae, Musaceae, Zingiberaceae, Costaceae, Heliconiaceae… ; photo 15-93).
En parallèle de la phytosociologie moderne des CVS, dite synusiale, il paraît aussi utile de considérer que l’IAA soit biologiquement homogène, donc distinguer des cultures vivaces et des cultures annuelles.
Enfin, comme en phytosociologie habituelle, il faut considérer la répétitivité de la composition en TC des IAA pour les décrire. Ceci a notamment été démontré localement par Brun-Hool (1980) qui a mis en évidence des types de jardins selon la flore cutivée.
Toujours au niveau de la structure des IAA, en reconstituant artificiellement des paysages, il serait souhaitable que celle-ci soit comparable à la structure des IAS pour une meilleure intégration au paysage naturel riverain. Pour cela, il faudrait établir les lois statistiques présidant à la cohabitation des P dans les IAS, sans doute intermédiaires entre une distribution au hasard (loi dite en 1/f0) et une distribution fortement corrélée (en 1/f2), soit une distribution en 1/f comme en musique et dans beaucoup de phénomènes naturels (Gardner, 1978).

Le relevé de l’IAA
La suite de la démarche descriptive des IAA est calquée sur celle des IAS et passe surtout par le relevé floristique. Plusieurs exemples sont donnés dans la publication de 1996 selon les régions du monde et leurs fonctions. Voici un exemple inédit, un boisement sylvicole de teck à la Lama au Bénin :
Tectona grandis 5 (avec présence d’une Loranthacée hémiparasite), Senna siamea +, Acacia auriculiformis +, Khaya senegalensis +, Gmelina arborea +.
Les syntaxons élémentaires artificiels et les associations végétales artificielles
Rapprochés par leur composition floristique, de nombreux relevés permettent de définir des catégories e.phytosociologiques homotones, des syntaxons élémentaires artificiels (Sy E A), sans doute de plus grande variabilité que les Sy E spontanés. Toutes les propriétés et descriptions de ceux-ci s’étendent sans difficultés aux Sy E A.
Ultérieurement, des Sy E A affines par leur composition en TC peuvent être rapprochés dans des associations végétales artificielles (AVA). Elles sont dotés d’une dimension chorologique ; d’ailleurs comme on l’a vu dès le début de ce panorama, cette dimension est inscrite d’emblée dans la définition du fait e-scientifique, donc aussi du fait e-phytosociologique.
Synécologie des AVA
On ne peut tout cultiver n’importe où, sauf à des coûts très élevés incompatibles avec la vie quotidienne. On peut donc associer à chaque Sy E A ou AVA une synécologie faite de plusieurs facteurs. Ce sont en grande partie ceux qui gouvernent la physiologie artificielle de l’Ager (§ 15.3). En particulier, l’intervention des facteurs climatiques peut se traduire par un étagement des AVA selon un gradient altitudinal ou latitudinal ; ainsi en Indonésie (Java, Bali), dans les plaines et collines, on observe une association à Manihot utilissima-Musa sapientium-Oryza sativa ; à partir de l’altitude de 900 m apparaissent plutôt des cultures de conditions plus tempérées (Daucus carota, Solanum tuberosum, Allium porrum, Brassica oleracea…).
Il faut y ajouter des facteurs ethnoécologiques et historiques induits par H : déplacements et remplacements de plantes ; choix, goûts, modes et savoir-faire des groupes ethniques, voire même psychologiques (l’ordre et la propreté, le désordre est combattu par la taille, la tonte, l’élagage ; Clément, 1990).
Dynamique des AVA
Les AVA possèdent une double dynamique. Comme pour les AVS, on peut distinguer d’abord une dynamique naturelle : les AVA sont stabilisées par les pratiques humaines ; abandonnées, elles sont progressivement remplacées par des AVS (cas des cultures sur brûlis après épuisement du sol). Lorsqu’une espèce est cultivée dans son aire spontanée, l’AVA peut parfois accueillir des espèces spontanées de l’AVS à laquelle cette espèce appartient (cas des cressonnières, AVA de Nasturtium officinale, qui s’enrichissent spontanément en Veronica beccabunga ou V. anagallis-aquatica ; photo 15-94).
Il existe aussi une dynamique provoquée : AVA1 → AVA2 par remplacement et modification des facteurs socio-économiques, disparition de plusieurs espèces anciennement cultivées. En 1848-49, de Mélicocq a dressé un inventaire des plantes spontanées et cultivées d’une petite région du nord de la France, d’un certain intérêt dans une perspective d’histoire rurale de nos terroirs ; à cette époque étaient donc cultivés (nomenclature actualisée) : Cameline (Camelina sativa), Oeillette (Papaver somniferum var. nigrum), Colza (Brassica napus var. napus), Lin (Linum usitatissimum), Trèfle (Trifolium pratense), Sainfoin (Onobrychis viciifolia), Vicia lens, Vesce (V. sativa), Fève (V. faba), Pois (Lathyrus oleraceus), L. sativus, Laitue (Lactuca sativa), Pomme de terre (Solanum tuberosum), Tabac (Nicotiana tabacum), Betterave (Beta vulgaris subsp. vulgaris), Sarrasin ou Boquis (Fagopyrum esculentum), Houblon (Humulus lupulus), Avoine (Avena sp.), Blé (Triticum sp.), Seigle (Secale cereale), Orge (Hordeum sp.). Plusieurs de ces taxons sont totalement éteints de nos jours dans cette région (Cameline, Oeillette, Sainfoin…), alors que d’autres cultivés actuellement y manquent, dont surtout le Maïs. D’un point de vue historique, on peut aussi se demander quelles étaient les espèces cultivées avec celles que l’on observe actuellement, lorsque celles-ci proviennent de régions assez récemment découvertes. C’est la question que se posent notamment Chevalier (1949) pour la zone soudanienne, Haudricourt & Hédin (1987) pour des espèces cultivées en Afrique après la découverte du Nouveau Monde. Le tableau 2 liste les espèces observées par ces auteurs et précise celles d’origine africaine qui auraient pu précéder les cultures issues du Nouveau Monde.

De même Lieutaghi (1992) précise que les doliques de l’Ancien Monde (Lablab purpureus, Vigna unguiculata) furent remplacés par des Phaseolus après la découverte de l’Amérique.

Hiérarchie et synsystème, nomenclature
Comme pour les AVS, il existe une possibilité de hiérarchiser les AVA en un système homologue du synsystème de la phytosociologie classique. D’ailleurs Brun-Hool (1980) a dressé une ordination hiérarchisée des AVA qu’il a décrites avec la « classe des Callistepho-Violetea », subdivisée en différentes AVA.
Pour faciliter le dialogue et la transmission du savoir acquis, on peut enfin envisager une nomenclature calquée sur celle des AVS en modifiant légèrement les radicaux caractéristiques des niveaux hiérarchiques : par exemple –elum pour l’AVA au lieu de –etum. Ainsi la classe de Brun-Hool pourrait être plutôt dénommée « Callistepho-Violelea » ; le boisement à Teck évoqué plus haut peut se rattacher à un Senno siameae-Tectonelum grandis ; la photo 15-95 illustre une AVA de balconnière fleurie assez répandue, le Petunio punctatae-Pelargonielum peltati ; le fourré fruitier sous dattiers de la photo 15-96 peut être dénommé Punico granati-Oleelum europaeae. Plusieurs AVA tempérées culturales ont par ailleurs été définies dans la publication de 1996 : Beto vulgaris-Triticelum aestivi, Dauco sativi-Brassicelum oleraceae, Brassico oleraceae-Betelum crassae, Zeo maydis-Vitielum viniferae, Aveno sativae-Triticelum aestivi, Solano tuberosi-Triticelum aestivi.
En Guyane française, Gély (1986) reconnaît des agers fruitiers à cacaoyers (Theobroma cacao), orangers et citronniers (Citrus div. sp.), avocatiers (Persea americana), manguiers (Mangifera indica), mombins (Spondias mombin), sapotillers (Achras sapota), corossoliers (Annona muricata) ; l’auteur décrit ensuite diverses parcelles de cultures herbacées qui pourraient être décrites en termes de relevés.
En conclusion, il s’avère que les CVA peuvent être étudiées d’une manière tout à fait similaire aux CVS dans le cadre de l’e.phytosociologie. Cette approche permettrait aussi de symétriser les relevés de paysages naturels en abordant en parallèle les éléments spontanés et les éléments artificiels dans le cadre de la symphytosociologie.


15.5. Murets, clôtures, haies et barrières
Tant dans le Saltus que dans l’Ager, il est parfois important de disposer des limites à la divagation des animaux, pour les empêcher soit de partir trop loin, soit de détruire les récoltes. Murets et clôtures sont nécessaires aussi pour que les H se défendent ou se dérobent aux regards extérieurs, pour réduire la vitesse du vent (photo 15-55).
Les murets font appel à la structuration de pierres (Lewillon, 1991), soit {ΣR} (photo 15-97). Ces pierres peuvent avoir été accumulées en murger (photo 15-98) lors de l’épierrage des agers pour en faciliter la culture et alors reprises pour l’enclôture (cas de dynamique {ΣR, 0} → {ΣR, enclore}), mais elles peuvent aussi avoir été extraites de carrières. Il peut y avoir empilement de pierres jusqu’à une hauteur convenable ou plantation dans le sol de dalles plates dressées juxtaposées (opérateur Λ ; photo 15-99). Les murets servent aussi à retenir la terre dans des zones aux reliefs accusés (photo 15-100) ; dans les régions en déprise humaine, ils demeurent, plus ou moins éboulés, pour témoigner d’un passé agricole révolu (photo 15-101).





Les haies sont plutôt construites à partir de matériaux végétaux. D’une manière générale, les végétaux qui les structurent sont armés, de façon à bien remplir la fonction qui leur sera dévolue ; divers exemples en seront cités ci-après. À la suite de Diguet (1928, chapitre XII) sur des clôtures de Cactaceae mexicaines, on doit à Aubert de la Rüe (1968) une remarquable étude sur les clôtures à travers le monde (bien plus large que le simple mot Remarques qui commence le titre de cette publication), bien que la détermination botanique des taxons reste souvent peu précise ; il n’évoque toutefois pas celles à Alluaudia ou Didierea madagascariensis (Didiereaceae, famille vicariante des Cactaceae) pour le sud de Madagascar (photos 15-102 et 15-103). Il est intéressant de remarquer que cette vicariance e.scientifique s’accompagne d’une proximité systématique puisque ces deux familles se rangent dans le même ordre des Caryophyllales.


On peut d’abord distinguer les « haies mortes » structurées à partir de matériaux secs : {ΛΣde ou |de| P} ou {vde ou |de| P} selon que la structuration s’apparente à une sorte de vannerie dressée (cas de palissades à base de Phoenix dactylifera ou Arundo donax au Maghreb ; photo 15-104) ou non (photo 15-105) et selon que le matériau utilisé fait intervenir une relation directe ou indirecte. Viennent ensuite les « haies vives » où les végétaux sont encore vivants ; c’est le cas le plus fréquent dans les pays de bocage où elles peuvent faire l’objet de soins divers : taille pour l’entretien (t ; photo 15-106), plessage (ou tressage, c’est-à-dire à rameaux entrelacés augmentant sa fonction défensive, opération assimilable à v ; photo 15-107). On peut étudier les haies vives semi-naturelles, ensembles d’arbustes réunis en unités physionomiquement homogènes par la méthode phytosociologique classique (voir par exemple Delelis-Dusollier, 1973), mais la description des haies vives artificielles nécessite l’utilisation de la méthode des AVA. Ainsi, la photo 15-108 illustre un type de haie vive bien armée à Opuntia ficus-indica et Agave americana interprétable en AVA sous le nom d’Opuntio fici-indicae-Agavelum americanae (de Foucault, 1996, 2013). Diguet (1928 : 355) rapporte la photographie d’une association artificielle à Pachycereus marginatus, Opuntia auberi et Pereskiopsis rotundifolia (nomenclature actualisée selon Catalogue of life), alors que Aubert de la Rüe (1968 : 226) évoque une haie à Agave et Cereus sepium (actuellement Borzicactus sepium).





La distinction entre haie morte et haie vive n’est pas toujours nette, notamment quand des végétaux a priori morts sont plantés pour former une haie morte et se remettent à bourgeonner et se développer (cas des saules ; Aubert de la Rüe, 1968 : 235, évoque encore le cas de Jatropha curcas).
Il existe aussi des murets mixtes (« minéro-végétaux ») plus complexes issus de la structuration d’un ensemble de pierres et de rameaux végétaux secs armés afin d’augmenter la fonction défensive (photos 15-109 et 15-110) : {Σde ou |de| P, R}. Ces photos illustrent le cas de murets où les deux composantes sont superposées, mais il existe des cas où elles sont juxtaposées de façon à constituer une forme plus large. Un tel cas a été illustré par Seignobos (1980) en zone soudano-sahélienne (Nord-Cameroun et Tchad). Des figures 15 et 18 de cet auteur, on peut décrire par la méthode des AVA (§ 15-4) huit IAA combinant un mur de pierres (matériau non précisé) et des plantes armées (tableau 3). On a réuni Euphorbia unispina et E. desmondii car le symbolisme utilisé sur ces figures (× pour le premier, + pour le second) ne permet pas d’y distinguer clairement les deux taxons quand ces signes sont inclinés. Ces huit relevés montrent la répétitivité de cette combinaison de formes naturelles et la possibilité de décrire une AVA pouvant être dénommée Euphorbio unispinae-Commiphorelum africanae. La figure 17 du même auteur donne une coupe transversale de cette AVA.



Aubert de la Rüe (1968, planche IV, bas) illustre un cas de muret mixte du Brésil à composantes minéro-végétales superposées, soit Σ(ΣR, veP). Dans sa planche IX (toujours en bas), il illustre un cas plus complexe à composantes juxtaposées : une composante vive à Euphorbia tirucalli (E. t.) dense vers le haut mais dénudée à la base, ce qui nécessite d’y placer devant une composante mixte où P n’est pas précisé : Σ[ΣE. t., (ΣR, veP)]. Dans sa planche X (haut), il décrit une palissade de Bambou fendu et tressé : vΔeP.
Si la défense est la fonction principale de la haie, elle peut être aussi une source d’aliments (surtout petits fruits) et de fourrage lors des saisons défavorables à la pousse en prairie, de plantes médicinales, de plantes toxiques pour la chasse ou la lutte contre les prédateurs, d’énergie pour le chauffage et de matériaux, de refuge (voir l’histoire de la chouannerie dans l’Ouest et le Nord-Ouest). Les haies de Cactaceae et Agave d’Amérique tropicale offrent ainsi leurs fruits et un produit permettant de préparer le pulque, voire la tequila. La fonction symbolique et magique, en protégeant l’occupant de la parcelle enclose des esprits malfaisants, s’est quelque peu perdue de nos jours (Portères, 1965 ; Vivier, 1970), quoique le transfert des maladies humaines aux arbres et arbustes, via des rituels ou des vêtements ayant touché le malade (« loques »), soit encore actif dans certaines régions de France et de Belgique (Saintyves, 1918 ; photo 15-111).

Ces haies semi-naturelles ou artificielles font l’objet d’une dynamique vers des clôtures non naturelles aussi armées, mais alors en fil de fer barbelé, voire plus récemment vers des clôtures électriques.
L’enclos ne peut pas être continu afin de permettre les mouvements des troupeaux et des hommes ; des passages susceptibles d’être ouverts puis fermés doivent être ménagés, ce sont les barrières. Les barrières de prairie ont fait l’objet d’une large monographie (de Foucault, 1992a) à laquelle on renvoie pour les détails.
Après la description de trente-et-un types de barrière, diversité associée à une fonction unique (comme en anatomie humaine : « à une grande fonction correspond une mutiplicité de structures », Piaget, 1967 : 202), accompagnée de cartes de dispersion géographique (parfois deux types sont observables dans la même région ; ils sont distingués ici par les lettres a et b), une synthèse a permis de les hiérarchiser selon un système (tableau 4).







Dans un autre chapitre, on a abordé la dynamique de ces objets, en reconnaissant trois grandes voies d’évolution (figure 19).
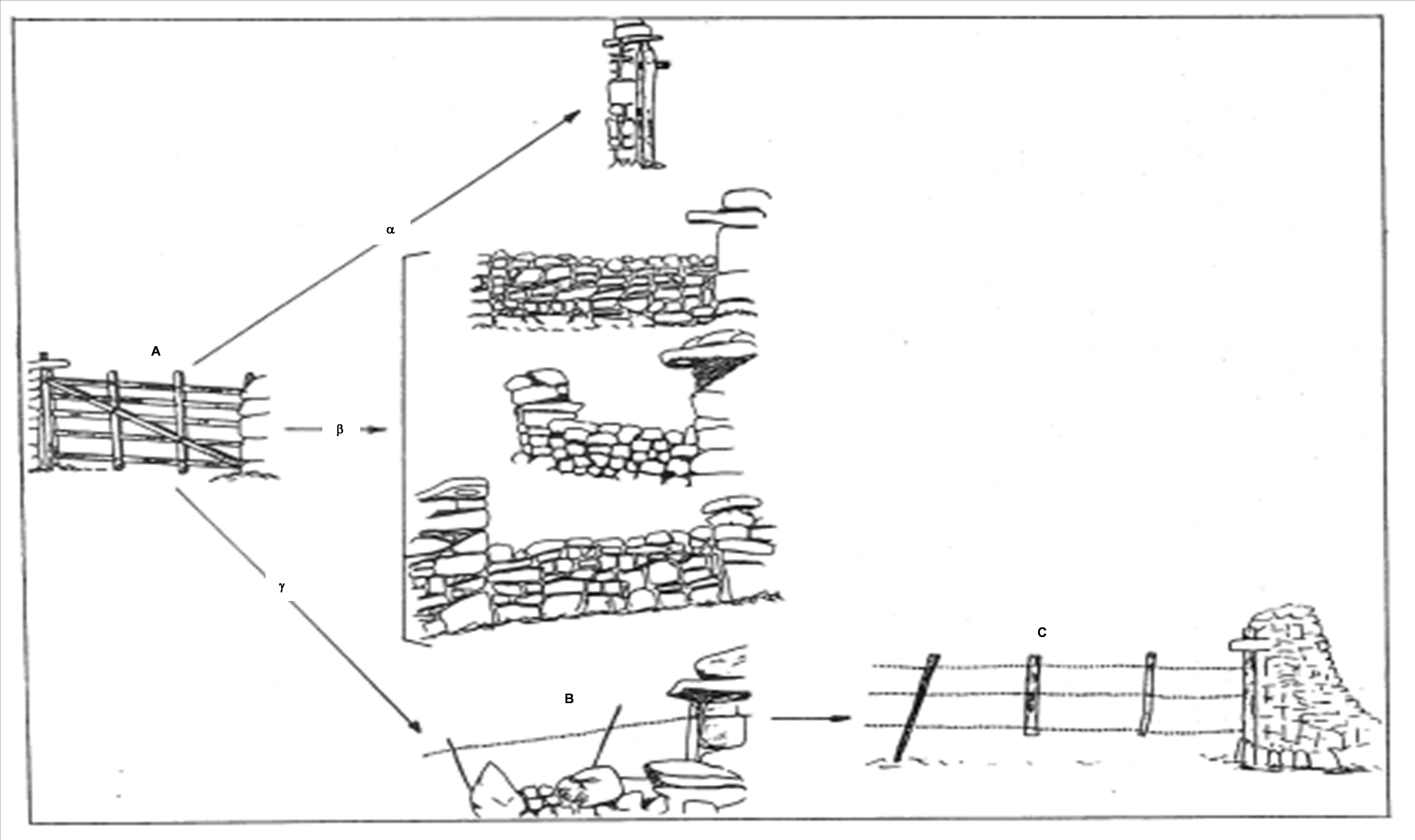
La voie dite α est un abandon pur et simple de la barrière ; oubliée de tous, celle-ci n’est plus utilisée pour sa fonction, elle perd progressivement ses éléments structurants. Cette dynamique est souvent associée à un changement de fonction de la parcelle riveraine. Cette voie se retrouve dans l’évolution d’autres formes dépourvues de fonction : maisons, tours, donjons… (photo 15-118). Pour lutter contre cette évolution, il faut injecter de l’énergie et reconstruire, soit par un travail direct, soit en passant par des artisans rémunérés ; on retrouve ici l’équivalence monnaie/énergie déjà évoquée en conclusion d’une autre publication (de Foucault, 2014b : 437).

La voie ß décrit une dynamique où la barrière est enlevée et l’espace ainsi libéré fermé par un muret de pierre ou une haie, formes qui prolongent donc les lignes clôturantes interrompues par la barrière initiale, rétablissant une continuité. Les annexes de la barrière qui assuraient son maintien peuvent se maintenir en tant qu’élément relictuel invariant (dans la figure 19 : une pierre percée), mais de fonction désormais nulle. Cette voie se retrouve dans la dynamique de certaines ouvertures (portes, fenêtres) murées (photo 15-119).

La voie χ, enfin, est la plus fréquente, c’est la substitution progressive de la forme initiale par une barrière finale différente (à fil barbelé ou électrique) ; ici, la fonction globale de la forme est invariante, de même que celle de la parcelle riveraine le plus souvent. Il peut aussi rester des éléments relictuels de la barrière initiale, de fonction variante ou invariante. Cette voie est très générale car elle se retrouve aussi dans l’évolution de structures architecturales comme des bâtiments (remplacement de portes anciennes, restauration…). Mais parfois elle aboutit aussi à une banalisation des barrières : aux types régionaux localisés géographiquement, se substitue alors un type unique d’aire généralement bien plus vaste (de Foucault, 1995a).
La présence d’éléments de fonction nulle amène à une autre réflexion : ces éléments inutiles ne peuvent être interprétés que comme relictuels d’une barrière initiale dans laquelle ils avaient quelque fonction non nulle. Là encore, un rapprochement peut être fait avec l’évolution des êtres vivants : « on connaît aujourd’hui chez la plupart des êtres vivants de pareilles structures totalement dénuées d’utilité actuelle et qui ne sont que d’énigmatiques absurdité si l’on se refuse à les considérer comme des vestiges de structures jadis fonctionnelles » (Rostand, 1931).
Sous cette hypothèse, alors ces éléments de fonction nulle peuvent apporter des informations sur la forme initiale disparue dans laquelle ces éléments possédaient une fonction : l’inutile fonctionnel se transforme en utile heuristique (de Foucault, 1991b : 91) ; n’a-t-on pas là toute la philosophie de la recherche archéologique et paléontologique sur des objets du passé de fonction actuelle nulle ?
16. Mécanismes nomenclaturaux
La nomenclature c’est la démarche de mettre un nom sur un objet. Elle paraît non forcément nécessaire : c’est un acte social, nommer ne devient nécessaire que pour communiquer à l’intérieur d’un groupe humain, quelle que soit la nature de celui-ci.
En termes sémiologiques, on a donc un sème *signifiant N(E)/signifié = taxon E* (cf. § 2.1). On en déduit la notion de synonyme (plusieurs N distincts nomment le même E) et d’homonyme (plusieurs E distincts portent le même N). Les homonymes étant source de confusion, le groupe social cherchera à en minimiser le nombre. Les botanistes ou phytosociologues scientifiques n’échappent pas à cette règle et le traitement des homonymes et des synonymes est régi par les codes internationaux de nomenclature botanique ou phytosociologique. Quand on change de règne, l’homonymie peut cependant réapparaître ; ainsi Oenanthe et Pieris désignent des genres de P, mais aussi d’oiseau pour le premier, d’insecte pour le second.
16.1. Nomenclature des taxons scientifiques de rang supérieur au genre
Pour permettre le dialogue entre scientifiques, il est utile de nommer les catégories systématiques, quel que soit leur rang hiérarchique. Cette nomenclature peut être aussi arbitraire qu’on le désire ; mais, afin de faciliter la mémorisation des rangs hiérarchiques de ces catégories, il sera commode de forger les étiquettes nomenclaturales à partir de radicaux linguistiques invariants, signifiants universels des divers rangs, donc de définir des sèmes *rang hiérarchique / radical signifiant* ; en botanique :
- -psida/classe
- -ales/ordre
- -aceae/famille
- -oideae/sous-famille.
Ces radicaux s’appliquent à un nom représentant de la catégorie nommée : Magnoliopsida, Rosales, Ericaceae, Vaccinioideae.
16.2. Nomenclature des taxons de rang inferieur
Le mécanisme nomenclatural est le passage E → N(E). Deux grands types de nomenclature au rang inférieur existent :
- la nomenclature scientifique (nom. sci.) : binominale, genre et espèce, dont on peut étudier la genèse indépendamment ; elle possède une valeur internationale, permettant aux scientifiques de se comprendre, de parler de la même chose ; dans ce domaine, il ne faut pas prendre les noms scientifique trop à la lettre, il s’agit avant tout d’une étiquette ; par exemple, Paris quadrifolia peut posséder cinq feuilles, Athamanta cretensis n’existe pas en Crète, Petasites hybridus n’est pas un hybride, Melampyrum pratense ne vit pas vraiment dans les prés ;
- la nomenclature vernaculaire (nom. vern.) : non fixée et à valeur purement locale, donc riche en synonymes ; le recueil des noms populaires d’un groupe social donné est souvent réalisé, mais se réduit fréquemment à une mise en correspondance (traduction) avec les équivalents scientifiques objectifs ; dans de telles enquêtes, il paraît essentiel non seulement bien sûr de recueillir cette diversité, ce patrimoine linguistique, mais encore de tenter d’en expliciter la signification : « Dresser la liste des noms vernaculaires est utile, mais une meilleure connaissance de leurs significations semble nécessaire. À ce sujet une étude plus minutieuse des manières employées aujourd’hui dans les îles pour désigner des plantes récemment introduites pourrait nous aider à comprendre la signification et l’origine de quelques-uns de ces noms anciens » (Barrau, 1964 : 208).
L’étude de nombreux mécanismes nomenclaturaux botaniques, tant scientifiques que vernaculaires, a révélé une vingtaine de grands types, dits aussi structures. Beaucoup d’entre eux peuvent s’étendre à la myconymie, à la phytosocionymie… (de Foucault, 1995b ; de Foucault & Claisse 1995 ; de Foucault & Amat, 1996 ; Claisse et al., 2000)… ; on se placera donc ici dans le cadre général des E. Exceptionnellement, on écrira en italique les termes illustrant les structures ; entre parenthèses on précisera le nom scientifique.
Les structures élémentaires
Les structures les plus simples, élémentaires, ont donné lieu à la systématique qui suit (de Foucault, 1993b).
- 0 : c’est le zéro nomenclatural, « appliqué » aux E rares ou sans rapport avec les H
- 1 à 13 : structures « phytologiques », attachées de près à E
- 1 à 4 : au sens strict, le mécanisme nomenclatural ne faisant appel qu’à des particularités de E
- 1 – sans comparaison : utilisation des organes, de caractères divers (forme, couleur, comportement)… traduits par des radicaux signifiants
- 1a – morphologiques, perçus par les sens (vue, toucher, odorat, goût) : Pterocarya, gratteron (pour Galium aparine), petit-gris (pour Tricholoma div. sp.), rouge-gorge, phonolite, henza (la puante pour Cleome arabica au Maroc), bois Carré et bois en T
- 1b – éthologiques : Epifagus, perce-neige (pour Galanthus nivalis), jouet du vent (pour Apera spica-venti), végétation nomade, cincle plongeur, sable mouvant
- 1c – phénologiques : Eranthis, herbe de la Saint-Jean (pour Hypericum perforatum et d’autres P), aoûtat
- 2 – par comparaison à autre E qui est un P : Daphniphyllum, chou-fleur (pour Sparassis crispa), anémone de mer (pour Actinaria sp.)
- 3 – par comparaison à autre E qui est un Z : Cynosurus, pied d’oiseau (pour Ornithopus), pied de mouton (pour Hydnum repandum)
- 4 – par comparaison à objet, liquide… : Sagittaria, bourse à pasteur (pour Capsella bursa-pastoris), ġaïta (la petite trompette au Maroc pour Datura stramonium), mitre d’évêque (pour Helvella sulcata), manteau et ourlet, orgues basaltiques (photo 16-1), bois en Accent circonflexe, bois en Fer de lance
- 1 – sans comparaison : utilisation des organes, de caractères divers (forme, couleur, comportement)… traduits par des radicaux signifiants
- 1 à 4 : au sens strict, le mécanisme nomenclatural ne faisant appel qu’à des particularités de E

-
- 5 à 8 : structures sémiques : utilisation de sèmes dans lesquels les E sont rapprochés de critères d’autre nature
- 5 – autoécologique : Ammophila, anémone sylvie (pour Anemone nemorosa), mouillère, cryosol
- 6 – chorologique : Kalaharia (genre africain de Lamiaceae), champignon de Paris (pour Agaricus bisporus), l’india (l’indien pour Opuntia ficus-indica au Maroc)
- 7 – e.scientifique : Sambucus, herbe aux verrues (pour Chelidonium majus), fasuh (celle qui dénoue [les sortilèges] pour Ferula communis au Maroc), anthroposol, calcaire lithographique, bois des Mitrailleuses
- 8 – zoobotanique : Myrmecodia, arbre aux papillons (pour Buddleja davidii), cabaret des oiseaux (pour Dipsacus fullonum), bois des Lapins, bois des Geais
- 9 à 13 : « phyto-linguistiques » : des ressemblances purement liées à E sont traduites par une transformation linguistique
- 9 – sans transformation ou presque, utilisé dans les structures complexes ci-après
- 10 – par dérivation au moyen d’affixes
- 10a – d’un suffixe : Limoniastrum, phalloïde (Amanita phalloides), rizière, roseraie, landine
- 10b – d’un préfixe (ou d’un adjectif placé devant) : faux-fraisier (pour Potentilla sterilis), fausse girolle (pour Hygrophoropsis aurantiaca), pseudomaquis, pseudogley
- 10c – on peut placer ici des termes animaux à signifiant péjoratif apparaissant en affixe, par exemple des noms en –seris (Lagoseris, Hyoseris), cynocrambe, Hyoscyamus ; Frérot (2011 : 77) cite des noms d’espèces sahariennes sauvages non comestibles affublés d’affixes tels que « d’âne, de rat, de chacal, de lièvre »
- 11 – par anagrammatisation : Ifloga, Ofliga et Logfia (anagrammes de Filago, genre proche) ; absent de la nom. vern.
- 12 – par homophonie : Aremonia (homophone approximatif de Agrimonia) ; absent de la nom. vern.
- 13 – par emprunt littéral : Lonas (formé de lettres de Santolina) ; absent de la nom. vern.
- 14 à 17 : structures purement linguistiques utilisant des jeux sur les mots et les lettres
- 14 – sans transformation, N(E) étant conservé d’un nom usité en période prélinnéenne ou préscientifique : Yucca, harmal repris dans Peganum harmala et rtem dans Retama raetam au Maroc, taïga, igapo, erg ; en nom. vern., on peut illustrer par l’usage du nom scientifique en l’absence de nom populaire (Dahlia) ; cette structure participe à la conservation de noms vernaculaires, du patrimoine phytolinguistique populaire dans le domaine scientifique, qui a par exemple été bien mise en avant par des botanistes comme F. Aublet pour le patrimoine guyanais ;
- 15 – par transfert, N(E) est issu du transfert du nom d’un E bien différent, parfois indéterminé, du nom d’un personnage, d’un dieu : Paris, Daphne ; en nom. vern., cas d’utilisation du nom scientifique d’un genre proche comme geranium (pour Pelargonium), mimosa (pour Acacia)
- 16 – artificielle, N(E) créé de toutes pièces à partir de quelques lettres : Calepina par Adanson, Rorippa par Scopoli ; absent de la nom. vern.
- 17 – dédicace à un personnage dont on a utilisé le nom : Woodsia, Sesleria, Russula queletii ; en nom. vern., nom offert par le créateur d’une variété nouvelle (par exemple de Rosa), mais aussi reine-marguerite (pour Callistephus chinensis), modèles de Holttum, de Corner… (Hallé & Oldeman, 1970), napoléonite, cratère Copernicus (de la Lune), einsteinium, pic Laperrine (au Sahara), bois Cristofari ; peut être étendu à des objets mathématiques (théorème de Pythagore, constante d’Euler, postulat d’Euclide, droite de Simson), des lois de chimie et physique (loi d’Ohm, équation de Schrödinger, point de Curie…)…
- 18 à 21 : autres, la plupart n’existant qu’en nom. vern.
- 18 – origine surnaturelle : Dianthus, mors du diable (pour Succisa pratensis), champignon des fées (pour Marasmius oreades), bête à Bon Dieu
- 19 – personnification : bon Henri (pour Blitum bonus-henricus), demoiselle coiffée (photo 16-2)
- 5 à 8 : structures sémiques : utilisation de sèmes dans lesquels les E sont rapprochés de critères d’autre nature

-
-
- 20 – animalisation : grasse poulette (pour Chenopodium album)
- 21 – noms sans transparence, lorsque l’attribution du nom n’est pas explicitée ou reconnaissable
-
Les structures complexes
Assez souvent, les noms sont plus complexes que ceux que l’on vient de citer ; les structures élémentaires peuvent s’y combiner en sommes ou produits pour engendrer des noms plus complexes.
Les sommes
Le mécanisme est de la forme

par exemple (1a,5,+) : Agrostemma, jaunet d’eau (pour Nuphar lutea) ; (4,18,+) : sabot de Vénus (pour Cypripedium calceolus), bâton de Moïse (pour Nerium oleander au Maroc), cheminée de fées (photo 16-2), marmite de géants (photo 16-3), rond de sorcière (photo 16-4) ; (1d,10,+) : chênaie à molinie ; (5,10,+) : chênaie calcicole.


Les structures (X,9,+) utilisent un terme de base (9) associé à un déterminant (X) permettant de distinguer les divers E ayant le même terme de base ; (5,9,+) : (Asplenium) ruta-muraria, artichaut de muraille (pour Sempervivum sp.), muguet des pampas (pour Salpichroa origanifolia), ananas montagne et thym montagne (pour Pitcairnia bifrons et Tibouchina ornata en Guadeloupe), psalliote des forêts (pour Agaricus sylvicola) ; (9,17,+) : Inocybe patouillardi, Cortinarius bulliardii.
Les produits
Le mécanisme est ici de la forme E –X→ N1 –Y→ N2 → N(E) qui s’écrit (X,Y,×), X (ou Y) pouvant être une association d’idées A, par exemple (5,A,×) pour Ranunculus et (Juncus) bufonius, (6,A,×) pour Romulea, (6,11,×) pour Jacmaia.
Les opérations mixtes
Plus rarement une somme et un produit peuvent se combiner


Certaines structures n’existent qu’en nom. sci., d’autres qu’en vern. On peut approcher par là même la dynamique nomenclaturale dans la transformation intellectuelle
populaire → scientifique, avec
- disparition des structures 0 traduisant les progrès de la science qui tend à minimiser le nombre d’E inconnus ou ignorés, pouvant induire l’utilisation de termes spécifiques (structure 7) tels que praetermissa (pour ignoré dans Dactylorhiza praetermissa), neglectus (pour négligé des botanistes dans Galium neglectum), 19 et 20 (diminution puis disparition de la relation affective de E à H ; le savant vise l’objectivité), 18 (rare en nom. sci. ; le progrès des connaissances abolit le surnaturel et la mythologie) ;
- apparition des structures linguistiques 11 à 13, 16, traduisant des opérations plutôt intellectuelles, voire des jeux sur les mots.
17. Aspects heuristiques dans les e.sciences
Développer des approches heuristiques au sein des e.sciences devient nécessaire lorsque les informateurs font défaut pour aider l’ethnologue ; elles doivent permettre de poser des hypothèses à valider ou abandonner, faisant ainsi avancer la science. Ces informateurs sont soit des personnes qu’on ne comprend pas par quelque langage que ce soit (barrière linguistique dans l’espace), soit des personnes absentes ou qui n’existent plus (barrière temporelle).
De la formule initiale {E’ = aE, F’}, on peut déduire des problèmes heuristiques à une inconnue x ou X de la forme :
A. {E’ = xE, F’}, reconstitution des chaînes opératoires (Pélegrin et al., 1988) ;
B. {E’ = aX, F’}, reconnaissance des E entrant à l’origine d’un fait e.scientifique ;
C. {E’ = aE, X’}, reconnaissance de la fonction d’un fait observé.
Nous avons déjà plus ou moins abordé les problèmes du type A, passons alors aux types B et C.
17.1. Problème du type B
Le problème du type B revient à acquérir des informations sur le E inconnu impliqué dans un fait e.scientifique. Il a été posé par moi-même (1997a) pour connaître la plante à l’origine de balais rencontrés lors d’enquêtes. En présence d’informateurs, l’ethnologue pourra avoir accès à une partie de la réponse, se faire montrer E ou recueillir son nom vernaculaire N(E) qu’il pourra transformer éventuellement en nom scientifique. Mais que faire lorsque l’informateur fait défaut, lorsque l’ethnologue est seul devant E’ ? Quel degré de connaissance pourra-t-il acquérir ? Dans la formule
{aE = E’}, le problème revient à acquérir des informations sur E connaissant E’. Celles-ci ne peuvent être recherchées qu’au niveau des invariants de E dans E’ par a.
On va d’abord explorer ces invariants possibles selon les divers a élémentaires : 1, -1, v, Σ, d, h, g, m laissent une invariance totale ; d’autres transforment suffisamment E pour ne laisser qu’une invariance partielle : e, i, t, d, p, b, s, Δ. Pour les invariants par les chaînes opératoires plus complexes, on pourra alors réduire ces CO aux a de la seconde liste.
Notre problème de reconnaissance peut s’écrire comme une équation à une inconnue E’ = aX. Si on a pu reconnaître en E’ un invariant J par a, avec E’ = J ∪ E’, on pourra écrire X = J ∪ X puisque J est commun à E’ et X, X étant un résidu inaccessible par cette approche. On n’a donc pas forcément accès à l’intégralité de E mais seulement à sa partie invariante par a et donc à un ensemble CJ de E tel que [E / J ⊂ E] caractérisé par J. Si les êtres du type de E sont hiérarchisés dans un système (botanique pour P, zoologie pour Z…), selon l’intensité de l’invariance notée, CJ pourra être une unité systématique plus ou moins fine.
Premiers exemples à partir de quelques a
Pour e, la résolution dépend de la partie empruntée et des connaissances de l’ethnologue en systématique si l’emprunt appartient à la partie reproductive de P ; ainsi l’ethnologue pourra descendre assez bas dans la hiérarchie botanique justement surtout basée sur l’appareil reproducteur. Grâce à ses compétences taxonomiques, l’éminent bryologue P. Allorge (1937) a pu étudier la flore muscinale utilisée dans des matelas des Açores. Dans la publication de 1997b, on a reconnu la présence d’ombelles doubles dans un balai utilisé par des éleveurs nomades kurdes (région de Doğubayazıt, Turquie orientale ; photo 17-1), orientant évidemment vers les Apiaceae. L’examen d’un balai végétal rencontré sur des places de Venise révélait des feuilles aciculaires opposées (ou originellement verticillées) accompagnées de fleurs séchées et de capsules, orientant vers un Erica, plus précisément E. scoparia (à anthères dépourvues d’appendices). Les bois d’ébénisterie pourront être reconnus grâce à des ouvrages ou des clés spécialisés (Jacquiot, 1955 ; Jacquiot & Traynard-Durol, 1973).

Pour be, il s’agit de reconnaître E à partir de restes calcinés, c’est le domaine de l’anthracologie.
Le problème de la reconnaissance pour cpe est parfois posé par une maîtresse de maison interrogeant ses invités : Qui saura me dire quels sont les ingrédients entrant dans la préparation de ce plat ?.
La résolution pour se est reliée à la fidélité de la représentation artistique. À propos de l’utilisation des plantes dans l’art grec, Baumann (1984) rappelle la reconnaissance de Platanus orientalis sur des pièces de monnaie, du Lis blanc (Lilium candidum) ou de Pancratium maritimum sur des fresques ou des peintures. Mais lorsque la stylisation est extrême, aux fleurs, on peut tout au plus reconnaître une Dicotylédone, voire une Angiosperme (voir photo 12-28 pour le logo de la Thai Airways International : qui aurait reconnu une orchidée ici ?). Godin et al. (2008) tentent la même démarche d’analyse dans des tableaux de la peinture flamande en cherchant à reconnaître des plantes, des oiseaux, et même des associations végétales (Hordeetum murini par exemple).
Démarches complémentaires
Face à cette limite des invariants par a, on peut se demander si des démarches complémentaires ne pourraient pas réduire la part d’inconnu. On peut se baser sur la valeur heuristique de quelques lois.
On peut définir d’abord définir des lois chorologiques E/A(E) qui associent les E à leur aire géographique A(E). Au voisinage d’un petit territoire y de l’espace d’enquête, sous l’hypothèse de spontanéité de E en y, on peut définir un ensemble Cy = [E / y ⊂ A(E)]. L’hypothèse de spontanéité est importante ; par exemple beaucoup de P utiles sont étrangères à leur aire d’usage.
On peut définir ensuite des lois d’efficacité : n’importe quel E n’est pas utilisable pour remplir une fonction F’ donnée ; seuls ceux qui appartiennent à certains ensembles seront efficaces. On peut alors distinguer deux lois d’efficacité :
- l’une (loi d’efficacité I) qui associe à chaque F’ l’ensemble des E susceptibles de la remplir (quel que soit a) :
CF’ = [E / aE assure F’] ; - une seconde (loi d’efficacité II) associe à un E l’ensemble des F’ qu’il est capable de remplir : CE = [F’ / F’ assuré par aE].
Si on retient la loi d’efficacité I, on peut alors rapprocher nos démarches pour faire entrer l’inconnue X dans trois ensembles indépendants, de sorte que X appartient à l’intersection ensembliste CJ ∩ Cy ∩ CF’, ce qui limite donc encore l’imprécision sur X.
Ainsi, dans le cas du balai des steppes kurdes (photo 17-1), comme on l’a vu, la présence d’ombelles doubles renvoie vers une Apiacée, probablement spontanée dans ces steppes fort peu cultivées. L’étape ultérieure serait d’utiliser la flore de Turquie coordonnée par P.H. Davis pour éliminer les Apiacées non ligneuses et étrangères à la flore spontanée de cette région de Turquie orientale. La CO complète attachée à cet objet de la vie quotidienne du groupe kurde s’écrit {ΣeApiaceae, rassembler} et {veChèvre, assurer Σ}, cet objet servant à rassembler les déjections du troupeau de moutons et chèvres utilisées pour alimenter le feu familial dans cette région steppique où le bois est rare – {diZ, combustible} –, sa structuration étant assurée par un cordon de poils de chèvre tressés (d’où ve).
Un autre exemple porte sur un balai rencontré dans le Bugey, montrant des rameaux ligneux avec des cicatrices de rameaux secondaires opposés ; ce caractère morphologique et la loi d’efficacité I orientent vers un P phanérophyte (= arbre) ou nanophanérophyte (= arbuste, arbrisseau), en associant les cicatrices de rameaux secondaires opposés et la loi chorologique, soit les Buxus, Euonymus, Acer, Ligustrum, Fraxinus, Staphylea, Cornus, Viburnum, Sambucus et Lonicera arbustifs. Pour aller plus loin, il faudrait comparer des rameaux de ces plantes à ceux du balai ; il semble en définitive qu’il s’agissait de Lonicera xylosteum, le camerisier.
La balayette de Guadeloupe (Antilles françaises) de la photo 17-2 montre un limbe foliaire flabellé orientant vers un palmier. Ce pourrait être un taxon introduit, la flore de Fournet (2002) listant une dizaine de tels genres. Toutefois ici, une nouvelle loi, phytonymique celle-ci, pourrait rendre quelque service en limitant cette liste à deux genres seulement : deux Thrinax et un Coccothrinax localement dénommés « palmier à balai », d’où l’intérêt de collecter les noms vernaculaires des végétaux N(P). Cette nouvelle loi permet de définir la catégorie CN = [P / N(P) relié à F’].

17.2. Problème du type C
Il s’agit maintenant de déduire de la forme observée E’ sa fonction F’. Ce problème est bien plus difficile que le précédent et pourtant très important, en archéologie notamment. Leroi-Gourhan (1964 : 2) l’exprime ainsi : « Comment passerait-il (le préhistorien) de la superficie décevante des représentations à la profondeur mystique des concepts ? ». En termes sémiologiques, il s’agit globalement de rechercher le signifié d’un signifiant donné (cf. § 3). On perçoit encore cette difficulté dans un cahier consacré aux origines du sacré (Collectif, 2011), qui reconnaît que certains chercheurs vont au-delà des faits connus : « l’intime conviction du chercheur outrepasse souvent les limites qu’impose le témoignage matériel. » (p. 11, de J. Guilaine) ; de leur côté, Cahen et Karlin (19880) écrivent que « Plusieurs générations de préhistoriens se sont heurtées au problème de la fonction des outils sans parvenir à le résoudre efficacement. » et Dauvois (1989) discute la fonction d’une pièce allongée, mince, percée à une extrémité : s’agit-il d’un rhombe ou d’une pendeloque ?
Par approche analogique, on a longtemps cru que la comparaison avec des E’ se ressemblant morphologiquement et techniquement et de F’ connus pourrait apporter une solution ou au moins des hypothèses, en vain. On a ensuite recherché sur E’ des microtraces d’utilisation spécifiques de F’ et de la chaîne opératoire (Vaughan, 1983), comme par exemple un outil de silex ayant travaillé sur divers matériaux, de la même manière qu’en sédimentologie la microstructure des matériaux donne des informations sur leur origine et leur transport. Cette analyse fonctionnelle est l’objet d’une discipline archéologique appelée tracéologie, qui identifie les matériaux travaillés, le type d’action, voire finalement la fonction des objets analysés (Karlin et al., 1992). Cette possibilité est notamment liée au caractère impérissable de la pierre, qui conserve alors l’histoire des chaînes opératoires.
Pour remonter le temps, il est parfois possible de se déplacer dans l’espace pour enquêter auprès des personnes initiées qui connaissent encore les significations ; mais progressivement celles-ci disparaissent. Il y a donc urgence à collecter ces données.
Cas des gravures et peintures rupestres
Ainsi, en ce qui concerne la fonction des gravures et peintures rupestres (tG ou oG), s’agit-il d’ornements, d’un langage, d’un autre F’ ? Clottes (2000) y voit :
- l’affirmation d’une présence, une signature, un peu comme les graffiti qui en sont proches ;
- le témoignage, un message, donc une communication sur des évènements marquants, rituels, mythes, légendes, histoires ;
- une action sur le monde pour écarter le danger, participer à l’équilibre de la nature, en lien avec la sorcellerie ;
- la révélation d’un autre monde (chamanisme).
En revanche, Anati (1997) voit dans cet art visuel un langage afin que des peuples sans écriture, souvent non urbains et non lettrés, puissent communiquer ou mémoriser. Il disparaîtrait souvent par l’acquisition d’une écriture ; avec l’apparition de celle-ci, la syntaxe et l’essence des arts rupestres qui l’ont précédée viennent à disparaître, car les significations idéographiques sont remplacées par les inscriptions. De ce point de vue, l’art rupestre apparaît alors comme une bibliothèque d’archives. Le paléoethnologue italien y reconnaît
- des pictogrammes accompagnés de signes ou idéogrammes à valeur de relation ;
- des graphèmes, ensembles d’une grammaire, d’une syntaxe, des phrases ;
- des représentations accompagnées de symboles idéographiques pour former des phrases traduisant un langage visuel logique ;
- des signes sur une flèche qui pourraient signifier une action telle que « atteindre l’objectif ».
Il utilise donc des termes de lexicologie comme phrases, syntaxe, grammaire ; il rapproche certaines gravures des idéogrammes chinois actuels qui relèvent d’une véritable écriture. Des superpositions de figures pourraient correspondre à des palimpsestes sans effacement, lesquelles pourraient alors permettre d’établir une chronologie et d’entreprendre une tentative de classification stylistique.
Cas des géoglyphes
Les géoglyphes sont des figures formées en dégageant ou en retournant les pierres assombries par la patine du temps, laissant ainsi apparaître une couleur bien plus claire de façon à représenter des animaux : ΔG ≡ sZ, où Z est un lézard, des camélidés, un colibri, un condor, un jaguar, un singe… ; les plus célèbres sont celles de Nazca (ou Nasca) au Pérou méridional, mais qui se prolongent vers le Chili septentrional (photo 17-3 ; voir aussi Clottes, 2000 : photo 38). Le même problème se pose ici : quelle la fonction des ces formes ? Certains pensent à des fonctions rituelles liées à l’astronomie (Unesco, 2009 : 484) ; d’autres pensent qu’ils feraient partie d’un système hydraulique très sophistiqué.

Lors de la résolution des problèmes de type B, on a utilisé la loi d’efficacité I ; on peut alors se demander si, par symétrie, la loi d’efficacité II pourrait être utile pour résoudre les problèmes de type C. Malheureusement, on ne pourra aller très loin dans cette voie, tant la richesse des F’ possibles pour un E’ donné paraît élevée. Ce type de loi formalise l’observation triviale qu’un liquide et un solide ont des fonctions distinctes, que pour polir une roche il est nécessaire de posséder un abrasif situé plus haut que celle-ci dans une échelle de dureté croissante, type échelle de Mohs.
D’ailleurs, ces hypothèses et théories sur des fonctions inconnues relèvent-elles vraiment de la science ? Si on suit les idées de l’épistémologue Popper (1982), on peut estimer que non, qu’elles sont en dehors de la science car irréfutables, ne pouvant être vulnérables à l’expérience : « …je prétends que les théories scientifiques ne peuvent jamais être tout à fait justifiées ou vérifiées, mais elles peuvent néanmoins être soumises à des tests. Je dirai donc que l’objectivité des énoncés scientifiques réside dans le fait qu’ils peuvent être intersubjectivement soumis à des tests. » ; « Cela signifie que leur forme (celle des énoncés) doit être telle qu’il soit logiquement possible tant de les vérifier que de les falsifier » (Popper, 1982 : 41, 36 ; les termes soulignés ici sont mis en italique par Popper lui-même).
Il est possible par ailleurs que la reconstitution des paléopaysages (photo 17-4) que l’on trouve parfois dans les ouvrages de paléobotanique soient aussi infalsifiables, et donc ne relèvent pas vraiment de la science réfutable. On a montré cependant (de Foucault & Coquel, 1998) que de telles reconstitutions associent des structures architecturales et des adaptations aux biotopes, déduites de l’analyse morphofonctionnelle des êtres actuels : « dans une large mesure, le présent demeure la clé qui permet de comprendre le passé » (Gall, 1995 : 5).

18. Conclusion
Parvenus au terme de ce panorama, nous ne pouvons que remarquer la richesse des interactions entre les hommes et leur environnement naturel, terme pris dans toute sa diversité. Le montage des photos permet parfois d’observer les vicariances entre phénomènes comparables mais observés dans des régions ou lors de périodes éloignées (voir les ensembles de photos 2-1 et 2-2, 5-8 et 5-9, 11-1 et 11-2, 11-5 à 11-10, 11-11 et 11-12, 12-27 et 12-28, 13-6 à 13-8,
14-3 et 14-4, 15-14 et 15-15, 15-20 et 15-21, 15-25 et 15-26, 15-37 et 15-38, 15-116 et 15-117). Cette richesse des productions humaines apparaît aussi dans le Patrimoine mondial de l’humanité (dit improprement « Patrimoine mondial de l’Unesco », ce dernier n’étant que l’organisme qui délivre le label), tant matériel qu’immatériel ; le présent panorama insiste aussi sur cette dimension en apportant des illustrations de ce patrimoine [photos 2-1 (à ce jour candidat à l’inscription sur la liste du Patrimoine mondial) à 2-3, 5-12, 5-13, 5-15, 9-3, 12-2, 12-5, 13-19, 15-18, 15-37, 15-38, 15-53, 15-68, 15-80, 15-118]. La diversité des liens structurels entre les hommes et les éléments naturels est encadrée par un petit nombre de relations élémentaires (l’annexe 3 en liste vingt-quatre), qui peuvent se combiner pour décrire des relations plus complexes. Il y a clairement une symétrie méthodologique avec l’approche systémique de la végétation, où un petit nombre de relations écologiques spatio-temporelles relient des unités définies au préalable par les méthodes habituelles de la phytosociologie (de Foucault, 1984, 1993d), mais aussi avec la typologie génétique des jeux de mots, où un petit nombre de relations linguistiques suffit à rendre compte de leur diversité (de Foucault, 1988b).
À plusieurs reprises, on a mis en évidence un parallélisme entre le vocabulaire de la biodiversité et celui de l’e.diversité (de Foucault, 2001) ; Haudricourt émet des idées voisines quand il écrit : « L’étude ethnographique d’un outil peut se faire de la même manière que l’étude d’une espèce vivante » (1955). Il existe en effet entre ces problématiques un certain nombre de concepts, des « invariants intersciences », révélant une unité profonde entre diverses sciences a priori sans rapports, concepts que l’on pourrait étudier en eux-mêmes avant de les appliquer à de nouvelles investigations. Alors les sciences les plus avancées sur le plan formel pourraient aider les sciences moins avancées dans leurs progrès épistémologiques. De son côté, Lévi-Strauss évoque des isomorphimes entre anthropologie et linguistique : « Comme les phonèmes, les termes de parenté sont éléments de signification ; comme eux, ils n’acquièrent cette signification qu’à la condition de s’intégrer en systèmes […] ; enfin la récurrence, en des régions éloignées du monde et dans des sociétés profondément différentes, de formes de parenté, règles de mariage […] donne à croire que, dans un cas comme dans l’autre, les phénomènes observables résultent du jeu de lois générales, mais cachées […], les phénomènes de parenté sont du même type que les phénomènes linguistiques » (1958/1985, Anthropologie structurale : 47). L’intérêt de cette idée réside dans le fait que, très probablement, les formalisations de sciences isomorphes sont elles-mêmes isomorphes ; dans chaque catégorie, on pourra considérer un paradigme (au sens de modèle) de science et le formaliser, alors cette formalisation pourra être en partie étendue aux sciences isomorphes au paradigme. Ces idées ont été à nouveau expressément évoquées par Lévi-Strauss : « Sans doute, ces problèmes diffèrent-ils quand au contenu, mais nous avons à tort ou à raison le sentiment que nos propres problèmes pourraient en être rapprochés, à condition d’adopter le même type de formalisation. L’intérêt des recherches structurales est, précisément, qu’elles nous donnent l’espérance que des sciences plus avancées que nous sous ce rapport peuvent nous fournir des modèles de méthodes et de solutions » (1958/1985, Anthropologie structurale : 330). Finalement, au sein de la grande diversité des sciences, quelques-unes, les plus anciennes, devraient se dégager comme paradigmes pour les sciences plus jeunes à la recherche de leur maturité : « Quand on parle de structure sociale, on s’attache surtout aux aspects formels des phnomènes sociaux ; on sort donc du domaine de la description pour considérer des notions et des catégories qui n’appartiennent pas en propre à l’ethnologie, mais qu’elle voudrait utiliser, à l’instar d’autres disciplines scientifiques qui, depuis longtemps, traitent certains de leurs problèmes comme nous souhaiterions le faire des nôtres » (Lévi-Strauss, 1958/1985, Anthropologie structurale : 330). Manifestement, ce modèle mûr, achevé à beaucoup d’égards, étendard auquel se rallient les jeunes sciences envieuses, est représenté par la physique classique, cette discipline si ancienne, la première à avoir usé du langage mathématique, lequel en retour lui a permis d’accéder au statut de science. Lévi-Strauss ne cache pas son attirance du modèle physique : « À condition de pousser assez loin l’analyse structurale, les sciences humaines pourraient espérer, comme les sciences physiques, atteindre un plan sur lequel, dans des régions et à des époques différentes, les mêmes expériences se dérouleraient de la même façon. Ainsi serions-nous en mesure de contrôler et de vérifier nos hypothèses » (1968, L’origine des manières de table : 39), tout en reconnaissant que l’homologie est seulement formelle, le contenu étant différent : « Aussi les sciences humaines ne peuvent-elles prétendre qu’à une homologie formelle, non substantielle, avec l’étude du monde physique et de la nature vivante » (1971, L’Homme nu : 575).
En définitive, c’est une transdisciplinarité qu’il s’agit de fonder, à bien distinguer de la pluri- ou multidisciplinarité qui vise à enrichir l’étude d’un objet d’étude concret en faisant converger diverses disciplines complémentaires. Au contraire, la transdisciplinarité explicite des structures générales formelles qui peuvent ultérieurement diverger en se concrérisant dans diverses disciplines isomorphes de ce point de vue ; comme l’écrit Benoist (1980), « Transdisciplinaire sera l’attitude philosophique qui cherchera à analyser les récurrences dans l’histoire et dans d’autres domaines voisins ou déconnectés d’enjeux épistémologiques semblables ». En définitive, cette démarche vise essentiellement à jeter des ponts entre les diverses sciences, à abattre des barrières virtuelles fréquemment dressées par une spécialisation trop poussée : « Il est intéressant d’établir à ce niveau des rapprochements méthodologiques avec des domaines très distincts, par exemple les sciences de la nature. Il n’en reste pas moins qu’il existe aujourd’hui dans l’espace méthodologique une zone de convergence où se trouvent abolies certaines des cloisons traditionnelles » (Borillo, 1978, à propos des progrès en archéologie).
ANNEXES
Annexe 1 – Le structuralisme (pour compléments, voir de Foucault, 1993d)
Pour comprendre ce qu’est le structuralisme, le plus simple est de partir de la notion classique de système, un ensemble d’éléments en relations les uns avec les autres. En décrivant et en nommant les éléments, puis en reconnaissant les relations qui les unissent, on peut décrire un certain nombre de systèmes élémentaires. Alors, de même que l’on classe des êtres vivants en catégories abstraites nommées espèces, caractérisées par le plus grand nombre possible de traits morphologiques communs (des invariants), on peut chercher à classer les systèmes élémentaires décrits qui se ressemblent en catégories appelées structures formelles (F-structures ; adjectif structural). Celles-ci seront caractérisées par des invariants portant sur les éléments des systèmes (traits biologiques et écologiques), mais aussi et surtout par les relations, qui acquièrent ainsi une certaine universalité. Des systèmes élémentaires rangés dans une même F-structure sont dits isomorphes (ils fonctionnement de la même manière) ; ils sont encadrés par des invariants, ce qui n’empêche nullement leur originalité, leur diversité. Les éléments de deux systèmes isomorphes reliés par une même relation à leurs voisins sont qualifiés d’homologues.
La démarche structuraliste se retrouve en mathématiques (structures de groupe, d’anneau, de corps, d’espace vectoriel), dans les sciences humaines (voir surtout C. Lévi-Strauss, G. Dumézil), la linguistique (N. Chomski), et a été introduite en phytosociologie (de Foucault, 1984). Dans le corps même de ce panorama formalisé, elle apparaît aussi dans les § 4.1, 12.6, 16.2 et 18.
Annexe 2 – Complément au § 8 (Un second cas concret : les balayettes et balais traditionnels)
Dans un courriel en date du 15 décembre 2022 modifié le 19 du même mois, Pierre Coulot rapporte une ancienne pratique du Haut-Doubs, en Franche-Comté : les balais utilisés dans les fumoirs à salaisons ou tuyés (prononcer tu-é ; on trouve aussi les écritures thués et tués) étaient fabriqués avec des branches d’épicéa (Picea abies, P. a.) et, quand ils étaient usés, la base des branches était conservée pour faire la cheville qui ferme les saucisses de Morteau. Ce fait e.botanique peut d’abord s’écrire {ΣeP.a., nettoyer les tuyés}. Il présente alors une dynamique originale faisant intervenir une relation indirecte non encore considérée mais qui devrait se retrouver dans de futurs autres faits, l’usure |u| (ou encore l’obsolescence), accompagnée d’une déstructuration du balai initial pour n’en garder que quelques baguettes :
{ΣeP.a. = balai, nettoyer les tuyés} → {Δ|u|ΣeP.a. = cheviller la saucisse de Morteau}.
De nos jours, toutefois, ces chevilles sont plutôt fabriquées en bois de hêtre, nouvel exemple de dynamique à invariance fonctionnelle.
Annexe 3 – Abréviations et symboles
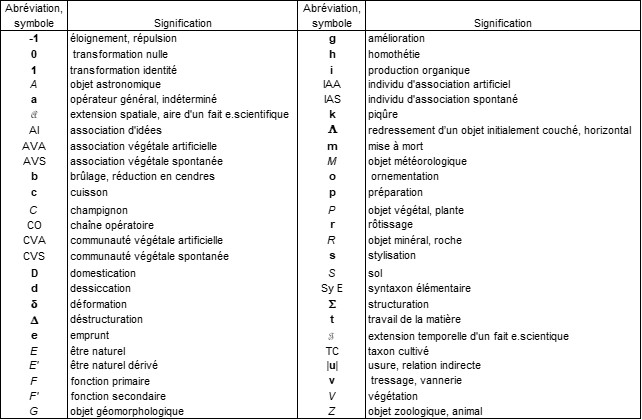
Bibliographie
Allorge P., 1937. Analyse bryologique de matelas. Revue bryologique et lichénologique 10 : 93.
Anati E., 1997. L’Art rupestre dans le monde, l’imaginaire de la préhistoire. Larousse, Paris, 422 p.
Anderson M., 1971. L’ethnobiologie. La Recherche 18 : 1029-1038.
Anonyme, 1974. L’arbre du Ténéré est mort. Bois et forêts des tropiques 153 : 61-65.
Armengaud Ch., 2009. Jouets de plantes, histoires et secrets de fabrications. Plume de carotte, Toulouse, 172 p.
Arnold N., 1985. Contribution à la connaissance ethnobotanique et médicinale de la flore de Chypre. Thèse de doctorat d’État, université de Paris V, 2 203 p.
Artielle G., 1986. Mycophilathélie. Bulletin de la Société mycologique du Nord 38, 39, 40.
Association des Stéphanois, 1987. Lous Esteves et las plantos. Saint-Étienne-de-Tinée, 50 p.
Aubert de la Rüe E., 1968. Remarques sur divers types de clôtures d’origine végétale. Journal d’agriculture traditionnelle et de botanique appliquée 15 (7-8) : 213-247.
Aumeeruddy Y. & Bakels J., 1994. Management of a sacred forest in the Kerinci valley, central Sumatra: an example of conservation of biological diversity and its cutural basis. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 36 (2) : 39-65.
Baize D. & Girard M.-C. (coord.), 1992. Référentiel pédologique, principaux sols d’Europe. INRA éditions, Paris, 221 p.
Barrau J., 1964. Notes sur la signification de quelques noms vernaculaires de plantes alimentaires dans les îles du Pacifique Sud. Journal d’agriculture traditionnelle et de botanique appliquée 11 (5-7) : 205-208.
Barrau J., 1970. Note sur le langage des plantes en Nouvelle-Calédonie mélanésienne. Journal d’agriculture traditionnelle et de botanique appliquée 17 (10-11) : 461-463.
Barrau J., 1971. L’ethnobotanique au carrefour des sciences naturelles et des sciences humaines. Bulletin de la Société botanique de France 118 : 237-248.
Barrau J., 1990. Ethnologie, II – Ethnosciences. Encyclopædia Universalis 8 : 1010-1012.
Barrier Ph., 1991. Forêt légendaire. Éditions Ch. de Bartillat, Paris, 254 p.
Baumann H., 1984. Le bouquet d’Athéna, les plantes dans la mythologie et l’art grecs. La Maison rustique, Flammarion, Paris, 250 p.
Baumer M., 1960. Quelques noms vernaculaires du Soudan nilotique utiles en écologie. Journal d’agriculture traditionnelle et de botanique appliquée 7 (6-8) : 299-315.
Baumer M., 1965. Quelques noms vernaculaires d’Arabie saoudite utiles en écologie. Journal d’agriculture traditionnelle et de botanique appliquée 12 (1-3) : 5-22.
Bellakhdar J., 2022. La jacinthe d’Orient (Hyacinthus orientalis L.) entre tradition et marchandisation, flânerie parfumée dans les allées de l’histoire sur la piste d’une fine fleur. Carnets botaniques 81 : 1-22.
Benítez Cruz G., Molero Mesa J. & González-Tejero García M.R., 2010. Floristic and ecological diversity of ethnobotanical resources used in western Granada (Spain) and their conservation. Acta Botanica Gallica 157 (4) : 769-786.
Benoist J.-M., 1980. La révolution structurale. Grasset, Paris, 345 p.
Bergeret P. & Petit M., 1986. Les systèmes agraires et la diversité des pratiques agricoles dans la zone des collines himalayennes. In J.-F. Dobremez (dir.), Les collines du Népal central, écosystèmes, structures sociales et systèmes agraires, I – Paysages et sociétés dans les collines du Népal, INRA, Paris : 119-139.
Blamont D., 1986. Facteurs de différenciation des systèmes agro-pastoraux des hauts pays du centre du Népal. In J.-F. Dobremez (dir.), Les collines du Népal central, écosystèmes, structures sociales et systèmes agraires, I – Paysages et sociétés dans les collines du Népal, INRA, Paris : 141-166.
Blanc P., 2002. Être plante à l’ombre des forêts tropicales. Nathan, Paris, 432 p.
Bonnet J., 1990. L’homme et le parfum. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 679-720.
Bourdy G., Cabalion P., Walter A. & Dijan-Caporalino C., 1995. Plantes magiques, plantes protectrices : quelques techniques d’horticulture traditionnelle à Vanuatu. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 37 (2) : 51-78.
Borillo M. (coord.), 1978. Archéologie et calcul. Collection 10-18, Editis, Paris, 246 p.
Boutrais J., 1980. L’arbre et le bœuf en zone soudano-guinéenne. Cahiers de l’ORSTOM, série Sciences humaines, XVII (3-4), L’Arbre en Afrique tropicale, la fonction et le signe : 235-246.
Braun-Blanquet J., 1936. La chênaie d’yeuse méditerranéenne (Quercion ilicis), monographie phytosociologique. Communications de la Station internationale de géobotanique méditerranéenne et alpine 45 : 1-150.
Brosse J., 1989. Mythologie des arbres. Plon, Paris, 360 p.
Brun-Hool J., 1980. Zur Pflanzengesellschaften schweizerischer Gärten. Phytocoenologia 7 : 73-99.
Bruneton-Governatori A., 1986. Un espace singulier : les jardins potagers en Pyrénées centrales. Terrain 6 : 34-46.
Cacciari I., Di Mattia E., Quatrini P., Moscatelli M.C., Grego S., Lippi D. & De Paolis M.R., 2003. Réponses adaptatives des isolats de Rhizobium aux stress. In M. Grouzis & É. Le Floc’h, Un arbre au désert, Acacia raddiana, IRD éditions, Marseille : 155-176.
Cahen D. & Karlin C., 1980. Les artisans de la préhistoire. La Recherche 116 : 1258-1268.
Calozet J., 1945. O payis dès sabotîs (Au pays des sabotiers), 2e édition. Nos Dialectes 1 : 53… (bilingue français-dialecte d’Awenne, Wallonie).
Candolle A. (de), 1883. Origine des plantes cultivées. G. Baillière, Paris.
Castetter E.F., 1944. The domain of ethnobiology. The American Naturalist 78, 775 : 158-170.
Chast F., 2010. La thériaque à l’époque moderne, chronique d’une fin annoncée du xvie au xixe siècle. Revue d’histoire de la pharmacie 368 : 493-510.
Chevalier A., 1949. L’agriculture coloniale. Que sais-je ? 62 : 1-125.
Chippaux C., 1990. Des mutilations, déformations, tatouages rituels et intentionnels chez l’homme. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 483-600.
Christanty L., 1981. An ecosystem analysis of West-Javanese home garden. Environment and Policy Institute, Honolulu (Hawaï), 15 p.
Claisse R., 1997. Environnement naturel et cultes féminins au Maroc. Al Biruniya (Revue marocaine de pharmacie) 13 (1) : 7-12.
Claisse R. & de Foucault B., 1999a. Banquets thérapeutiques et nourritures sacrificielles au Maroc. In A. Guerci (ed.), Il cibo culturale: dal cino alla cultura, dalla cultura al cibo, Erga ed., Genova : 117-126.
Claisse R. & de Foucault B., 1999b. Notes sur la « théorie des signatures ». Journal de botanique de la Société botanique de France 9 : 93-95.
Claisse R., de Foucault B. & Delelis A., 2000. Nommer les plantes et les formations végétales. L’Homme 153 : 173-182.
Claisse-Dauchy R. & de Foucault B., 2008. Espaces naturels, espaces sacrés : exemples africains, indiens et arabo-musulmans. In La perception de la nature de l’Antiquité à nos jours, Lille, 7 décembre 2007, 43-54.
Clapier-Valladon S., 1990. Les modes médicales. In J. Poirier, Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 775-830.
Clément G., 1990. Le jardin en mouvement. Pandora éditions, 101 p.
Clerc J., 1987. La tenderie aux grives, des mots, des procédés, des usages. Terres ardennaises 20 : 41-46.
Clottes J., 2000. Le musée des roches, l’art rupestre dans le monde. Seuil, Paris, 120 p.
Collectif, 2011. Aux origines du sacré et des dieux. Les Cahiers de Science & Vie 124 : 1-114.
Couderc J.-M., 1985. Les végétations anthropogènes et nitrophiles et la prospection archéologique. Colloques phytosociologiques XII, Les végétations nitrophiles et anthropogènes : 331-344.
Crampon M., 1936. Le culte de l’arbre et de la forêt en Picardie, esquisse sur le folklore picard. Mémoires de laSociété des antiquaires de Picardie 46 : 1-584.
Crosnier C., 1990. Savoirs en herbe, de la cueillette à l’usage. Exposition au Parc naturel régional du Morvan, Saint-Brisson.
Cuénot A., 1967. Les succédanés du Tabac (note préliminaire). Journal d’agriculture traditionnelle et de botanique appliquée 14 (6-7) : 191-252.
Dauvois M., 1989. Son et musique paléolithiques. Les Dossiers d’archéologie 142 : 2-11.
Decary R., 1968. L’emploi des succédanés végétaux à Madagascar pendant la dernière guerre. Journal d’agriculture traditionnelle et de botanique appliquée 15 (7-8) : 286-296.
Delelis-Dusollier A., 1973. Contribution à l’étude des haies, des fourrés préforestiers, des manteaux sylvatiques de France. Thèse de doctorat d’État en pharmacie, université de Lille, 146 p.
Diagram Group, 1978. Les instruments de musique du monde entier. Albin Michel, Paris, 320 p.
Diguet L. (revu par A. Guillaumin), 1928. Les Cactacées utiles du Mexique. Société nationale d’acclimatation de France, Archives d’histoire naturelle, Paris, 551 p., consultable à https://www.yumpu.com/fr/document/view/16564686/les-cactacees-utiles-du-mexique-bibliotheque-numerique.d-/402.
Diouri A., 1994. Lahlou : nourriture sacrificielle des Gnaoua du Maroc. In M. Marin & D. Waines, La alimentación en las culturas islámicas, Agencia española de cooperación internacional, Madrid : 169-216.
Dupont F. & de Foucault B., 1995. Ethnobotanique des bambous aux Célèbes. Bulletin de la Société de botanique du nord de la France 48 (1) : 47-52.
Durand-Tullou A., 1972. Rôle des végétaux dans la vie de l’homme au temps de la civilisation traditionnelle (étude ethnobotanique sur le Causse de Blandas, Gard). Journal d’agriculture traditionnelle et de botanique appliquée 19 (6-7) : 222-248.
Fédération des conservatoires d’espaces naturels, 2019. Conservatoires d’espaces naturels, à la découverte de sites remarquables. Glénat, Grenoble, 168 p.
Feller C., Étifier-Chalono E. & de Guiran E., 1986. Fragments d’un discours paysan antillais : « roche-pierre » et « sol-terre ». Journal d’agriculture traditionnelle et de botanique appliquée 33 : 131-142.
Flamand E.C., 1981. Les pierres magiques. Le Courrier du livre, Paris.
Foucault M., 1979. Les mots et les choses. Gallimard, Paris, 400 p.
Foucault B. (de), 1981. Les prairies permanentes du Bocage virois (Basse-Normandie, France) : typologie phytosociologique et essai de reconstitution des séries évolutives herbagères. Documents phytosociologiques, NS, V : 1-109.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’État, université de Rouen, 675 p.
Foucault B. (de), 1987a. Essai de formalisation de l’ethnobotanique. Journal d’agriculture traditionnelle et de botanique appliquée 34 : 31-45.
Foucault B. (de), 1987b. Nouvelles recherches sur les structures systématiques végétales : caractérisation, ordination, signification. Phytocoenologia 15 (2) : 159-199.
Foucault B. (de), 1988a. Compte-rendu de la session de la Société de botanique du nord de la France dans les Ardennes françaises (26-27 juin 1988). Bulletin de la Société de botanique du nord de la France 41 (3-4) : 3-6.
Foucault B. (de), 1988b. Les structures linguistiques de la genèse des jeux de mots. Sciences pour la communication 23 : 1-142.
Foucault B. (de), 1988c. Sur les bases épistémologiques du génie écologique, plus spécialement phyto-écologique. Colloques phytosociologiques XV, Phytosociologie et conservation de la nature : 77-91.
Foucault B. (de), 1990a. Introduction à une ethnophytosociologie. Lejeunia, NS, 134 : 1-60.
Foucault B. (de), 1990b. Ethnobotanique et invariance. Informatore Botanico Italiano 22 (1-2) : 34-41.
Foucault B. (de), 1991a. Approche sémiologique et structurale du langage des plantes. Bulletin de la Société botanique du Centre-Ouest, NS, 22 : 207-214.
Foucault B. (de), 1991b. Un chapitre de technologie rurale : la barrière traditionnelle de prairie, typologie et dynamique. Mémoires de la Société linnéenne Nord-Picardie II : 1-107.
Foucault B. (de), 1992a. Le domaine de l’ethnobotanique et ses extensions. Bulletin de la Société de botanique du nord de la France 45 : 57-64.
Foucault B. (de), 1992b. De l’ethnobotanique à l’ethnoscience naturelle. Bulletin de la Société linnéenne Nord-Picardie 10 : 15-34.
Foucault B. (de), 1992c. Mythologie et spéciation chez les plantes. Bulletin de la Société botanique du Centre-Ouest, NS, 23 : 139-156.
Foucault B. (de), 1993a. Contribution à une ethnobotanique de l’arbre vénéré. Revue forestière française 45 (5) : 581-590.
Foucault B. (de), 1993b. Les plantes et leurs noms : essai de phytonymie structurale. Dissertationes Botanicae 201 : 1-64.
Foucault B. (de), 1993c. II – Données phytosociologiques. In B. de Foucault & J.-P. Cornu, Itinéraire botanique d’un groupe de linnéens en Crète, 29 avril-13 mai 1991 (2e partie), Bulletin de la Société linnéenne Nord-Picardie 11 : 85-105.
Foucault B. (de), 1993d. Systémique qualitative et structuralisme en phytosociologie. Revue internationale de systémique 7 (4) : 363-384.
Foucault B. (de), 1994a. Monographie ethnobotanique de la cabana du feuillardier limousin. Annales scientifiques du Limousin 10 : 3-12.
Foucault B. (de), 1994b. Notes analytiques et comparatives sur la végétation tropicale observée en Asie du Sud-Est. Bulletin de la Société de botanique du nord de la France 47 (2) : 29-39.
Foucault B. (de), 1994c. Extension à la phytosociologie d’un concept botanique : la variation parallèle. Candollea 49 : 121-127.
Foucault B. (de), 1995a. Dynamique convergente et banalisation dans les paysages ruraux. Actes du colloque de géographie rurale, Les mutations dans le milieu rural, Caen 1992, Centre de recherches sur l’évolution de la vie rurale 18 : 253-264.
Foucault B. (de), 1995b. Sur la définition d’une phytosociologie populaire. Documents phytosociologiques, NS, XV : 141-151.
Foucault B. (de), 1996. La phytosociologie des associations végétales artificielles. Documents phytosociologiques, NS, XVI : 293-314.
Foucault B. (de), 1997a. Problèmes de reconnaissance de plantes en ethnobotanique : application aux balais et balayettes végétaux. Journal de botanique de la Société botanique de France 4 : 59-69.
Foucault B. (de), 1997b. La dualité structure-fonction en botanique : nouvelles réflexions. Adoxa 15-16 : 17-22.
Foucault B. (de), 1997c. La lutte contre les nuisibles de l’homme à la lumière de l’ethnobiologie formelle. Bulletin de la Société linnéenne Nord-Picardie, NS, 15 : 4-16.
Foucault B. (de), 2001. De la biodiversité à l’ethnodiversité. Journal de botanique de la Société botanique de France 16 : 45-59.
Foucault B. (de), 2010a. Ethnographie de quelques ruches traditionnelles. In L’Abeille et l’apiculture, CSENPC, Lille : 43-50.
Foucault B. (de), 2010b. Sur l’extension à la phytosociologie d’un concept de la physique : le phénomène d’hystérésis. Braun-Blanquetia 46 : 251-253.
Foucault B. (de), 2013. Les journées de l’Association tunisienne de physiologie et de biosurveillance de l’environnement (12-15 mai 2012) ; compte rendu phytosociologique des herborisations. Revue Écologie Environnement (Tiaret, Algérie) 9, non paginé (http://fsnv.univ-tiaret.dz/revues.php).
Foucault B. (de), 2014a. Les isomorphismes spatio-temporels : approche transdisciplinaire et valeur heuristique. Bulletin de la Société botanique du Centre-Ouest, NS, 45 : 438-445.
Foucault B. (de), 2014b. Un modèle énergétique pour la dynamique de la végétation. Acta Botanica Gallica: Botany Letters 161 (4) :429-439.
Foucault B. (de) & Bonnard A., 1992. Les champignons et leurs noms : note de myconymie structurale. Bulletin de la Société mycologique du Nord 51 (1) : 7-16.
Foucault B. (de) & Legrand J.-P., 1994. Monographie d’un fait ethnobotanique : le carcahoux de la forêt d’Eu (Seine-Maritime). Bulletin de la Société linnéenne Nord-Picardie 12 : 31-38.
Foucault B. (de) & Claisse R., 1994. Observations botaniques et ethnobotaniques en Afrique du Nord. Bulletin de la Société de botanique du nord de la France 47 (3) : 17-25.
Foucault B. (de) & Claisse R., 1995. Sur quelques noms vernaculaires de plantes de la pharmacopée traditionnelle marocaine (région de Rabat-Salé). Al Birunyia (Revue marocaine de pharmacie) 11 (1) : 45-50.
Foucault B. (de) & Amat J.-P., 1996. Bois et guerre : introduction à une analyse ethnophytosociologique. Mémoires de la Société linnéenne Nord-Picardie III, actes du colloque L’impact de la guerre sur la végétation et la régénération du tapis végétal, Albert, juin 1996 : 39-43.
Foucault B. (de) & Valcke M.-A., 1996. Coquelicots et bleuets après la guerre : analyse ethnobotanique. Mémoires de la Société linnéenne Nord-Picardie III, actes du colloque L’impact de la guerre sur la végétation et la régénération du tapis végétal, Albert, juin 1996 : 35-37.
Foucault B. (de) & Coquel R., 1998. Du fossile au paléo-paysage reconstitué. Bulletin de la Société de botanique du nord de la France 51 (1) : 43-49.
Foucault B. (de) & Claisse R., 1999. Le rite en tant qu’opérateur fonctionnel de l’ethnobotanique. In A. Guerci (éd.), Incontrita medicine (Meetings between medicines), Erga ed., Genova : 124-132.
Foucault B. (de) & Claisse R., 2000. La nomenclature des objets de la botanique et des sciences associées. In H. Vérine (ed.), Les sciences et leurs langages, éditions du CTHS, actes 119e et 120e congrès nationaux : 135-146.
Foucault B. (de), Batawila K., Wala K. & Akpagana K., 2011. Une formation végétale méconnue pour l’Afrique tropicale, la mégaphorbiaie ; le cas du Marantochloo–Anubiadetum giganteae du Togo. Acta Botanica Gallica 158 (3) : 409-424.
Fournet J., 2002. Flore illustrée des phanérogames de Guadeloupe et de Martinique. Cirad et Gondwana éditions, deux tomes, 2 538 p.
Fournier F. & Sasson A., 1983. Écosystèmes forestiers tropicaux d’Afrique. Recherches sur les ressources naturelles 19 : 1-473.
François M., 2014. Les baraques en sanills de l’étang de Salses. Chantiers 8 : 28-29 (www.culture.gouv.fr/regions/drac-occitanie/ressources-documentaires/publications/chantiers-la-lettre-du-pole-architecture-et-patrimoines).
Frérot A.-M., 2011. Imaginaires des Sahariens, habiter le paysage. Éditions du CTHS, Paris, 191 p.
Gall J.-C., 1997. Paléoécologie : paysages et environnements disparus. Masson, Paris, 239 p.
Gardner M., 1978. Musique blanche et musique brownienne, courbes fractales et musique en un sur f. Pour la Science 8 : 118-126.
Gautier D., 1994. La diversité des systèmes agroforestiers bamiléké et ses évolutions contemporaines. Journal d’agriculture tropicale et de botanique appliquée, NS, 36 (2) : 159-178.
Géhu J.-M., 1991. Sur la notion de cellules paysagères isofonctionnelles. Colloques phytosociologiques XVII, Phytosociologie et paysages : 189-193.
Gély A., 1986. Agriculture créole ; une étude de cas : Saül (Guyane française). Journal d’agriculture tropicale et de botanique appliquée 33 : 143-192.
Gennep A. (van), 1905. De l’héraldisation de la marque de propriété et des origines du blason. Bulletins et mémoires de la Société d’anthropologie de Paris 6 : 103-112.
Gennep A. (van), 1906. À propos de l’origine des runes. Revue des traditions populaires XXI (2) : 73-78.
Ghestem A., 2002. La flore des vestiges archéologiques en Limousin : proposition de synthèse. Travaux d’archéologie limousine 22 : 7-21.
Ghestem A., Botineau M., Froissard D. & Hourdin Ph., 2003. La voie romaine d’Orléans à Sens : analyse de son impact sur la flore forestière et comparaison avec la flore des vestiges gallo-romains limousins. Travaux d’archéologie limousine 23 : 17-28.
Gillet H., 1974. Découverte d’un piège à girafe. Journal d’agriculture tropicale et de botanique appliquée 21 (10-12) : 361-364.
Godin J., de Foucault B. & Petit D., 2008. Nature et paysages dans la peinture flamande : une approche naturaliste. In La perception de la nature de l’Antiquité à nos jours, Lille, 7 décembre 2007 : 71-78.
Gonthier E., 1990. L’homme et le minéral. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47 : 1389-1446.
Granet P. & Mathieu B., 1990. Cours d’égyptien hiéroglyphique, I. Khéops, Paris, 426 p.
Guelliot O., 1975. Géographie traditionnelle et populaire du département des Ardennes, 2e édition (la première en 1931). Librairie Guenegaud S.A., Paris, 410 p.
Guillaumet J.-L., 1972. Note sur la connaissance du milieu végétal par les nomades de la basse vallée du Wabi Shebellé (Éthiopie). Journal d’agriculture tropicale et de botanique appliquée 19 (4-5) : 73-84.
Hallé F., 1993. Un monde sans hiver. Le Seuil, Paris, 361 p.
Hallé F. & Oldeman R.A.A., 1970. Essai sur l’architecture et la dynamique de croissance des arbres tropicaux. Masson, Paris, 192 p.
Haudricourt A.-G., 1939. De l’origine de quelques céréales. Annales d’histoire sociale 1 : 180-182. Intégralement reproduit in Haudricourt, 1987 : 211-213.
Haudricourt A.-G., 1955. Biogéographie des araires et des charrues. Comptes-rendus des séances de la Société de biogéographie 280 : 77-83.
Haudricourt A.-G., 1968. L’ethnominéralogie. Encyclopédie de la Pléiade 24, Ethnologie générale : 1767-1771.
Haudricourt A.-G., 1987. La technologie, science humaine ; recherches d’histoire et d’ethnologie des techniques. Éditions de la Maison des sciences de l’homme, Paris, 343 p.
Haudricourt A.-G. & Hédin L., 1943 réédition 1987. L’homme et les plantes cultivées. A.M. Métailié, Paris, 281 p.
Haudricourt A.-G. & Jean-Brunhes Delamarre M., 1955, réédition 1986. L’homme et la charrue à travers le monde. L’homme et la nature, La Manufacture, Lyon, 410 p.
Hubert-Schoumann A., 1974. L’essartage ou culture sur brûlis en Thaïlande du Nord. Journal d’agriculture tropicale et de botanique appliquée 21 (7-9) : 251-255.
Huffman M.A., 1995. La pharmacopée des chimpanzés ; du bon usage des plantes chez les animaux, histoire d’une science en herbe. La Recherche 280 : 66-71.
Huysmans J.-K., 1898 (réédité 1930). La cathédrale. In Œuvres complètes, XIV, éditions G. Grès & Cie, 348 et 342 p.
Jackson D., 1982. Histoire de l’écriture. Denoël, Paris, 176 p.
Jacquiot C., 1955. Atlas anatomique des bois de conifères, 2 volumes. Centre technique du bois, Paris, 135 p. + planches.
Jacquiot C. & Traynard-Durol D., 1973. Atlas anatomique des bois des Angiospermes, 2 volumes. Centre technique du bois, Paris, 75 p. + planches.
Jean-Brunhes Delamare M., 1985. La vie agricole et pastorale dans le monde, techniques et outils traditionnels. Édition Joël Cuénot, 216 p.
Jelínek J., 1989. Sociétés de chasseurs, ces hommes qui vivent de la nature sauvage. Gründ, Paris, 208 p.
Karlin C., Pigeot N. & Ploux S., 1992. L’ethnologie préhistorique. La Recherche 247 : 1106-1116.
Kerenyi C., 1968. De l’origine et du fondement de la mythologie. In C.G. Jung & C. Kerenyi, L’essence de la mythologie, Petite bibliothèque Payot 124, Paris : 11-41.
Kuhnholtz-Lordat G., 1958. L’écran vert. Mémoires du Muséum national d’histoire naturelle, NS, Botanique, 9 : 1-276.
Lachowski F., 1988. Faites inverner vos génisses sur la côte. La France agricole 2238 : 37-38.
Lapeyre O. & Dodinet E., 1982. L’homme et la plante. Bulletin du Groupe de recherches historiques et archéologiques de la vallée de la Sumène 26-27 : 1-96.
Le Floc’h É. & Grouzis M., 2003. Acacia raddiana, un arbre des zones arides à usages multiples. In M. Grouzis & É. Le Floc’h (dir.), Un arbre au désert, Acacia raddiana, IRD éditions, Marseille : 17-49.
Le Mouël J.-F., 1969. Connaissance et utilisation des végétaux chez les Eskimos Naujâmiut (Groenland occidental). Journal d’agriculture tropicale et de botanique appliquée 16 (11-12) : 469-494.
Lebeau R., 1979. Les grands types de structures agraires dans le monde. Masson, Paris, 162 p.
Leblic I., 1995. La pêche au poison en Nouvelle-Calédonie. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 37 (1) : 217-235.
Lecomte Th., Le Neveu Ch. & Jauneau A., 1981. Restauration de biocénoses palustres par l’utilisation d’une race bovine ancienne (Highland Cattle) : cas de la réserve naturelle des Mannevilles (Marais Vernier, Eure). Bulletin d’écologie 12 : 225-247.
Ledrut R., 1990. L’homme et l’espace. In J. Poirier, Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 58-114.
Léon J. (de). Notes sur l’élevage en Israël du ver à soie nourri de feuilles de ricin. Journal d’agriculture traditionnelle et de botanique appliquée 10 (12) : 545-550.
Leroi-Gourhan A., 1964. Les religions de la Préhistoire (Paléolithique). Quadrige, Les Presses universitaires de France, Paris, 152 p.
Leroi-Gourhan A., 1992a. Milieu et technique. Albin Michel, collection Sciences d’aujourd’hui, Paris, 475 p.
Leroi-Gourhan A., 1992b. L’homme et la matière. Albin Michel, collection Sciences d’aujourd’hui, Paris, 348 p.
Lévi-Strauss C., 1958, réédition 1985. Anthropologie structurale. Agora, Plon, Paris, 480 p.
Lévi-Strauss C., 1968. Mythologiques, III – L’origine des manières de table. Plon, Paris, 478 p.
Lévi-Strauss C., 1971. Mythologiques, IV – L’Homme nu. Plon, Paris, 688 p.
Lewillon S., 1991. Les murs de pierre sèche en milieu rural. In J. Guilaine (dir.), Pour une archéologie agraire, Armand Colin, Paris : 193-221.
Liendo Tagle I., 2021. Le génie des Incas pour protéger les récoltes. Courrier international 1617 : 31-35.
Lieutaghi P., 1992. Jardin de savoir, jardin d’histoire. Les Alpes de lumière 110-111 : 1-148.
Lieutaghi P., 1998. La plante compagne ; pratique et imaginaire de la flore sauvage en Europe occidentale. Actes Sud, Aix-en-Provence, 299 p.
Lormée N., Caballion P. & Hnawia É. (éds), 2011. Hommes et plantes de Mahé, îles Loyauté, Nouvelle-Calédonie. IRD éditions, Marseille, 358 p.
Mélicocq A. (de Lafons de), 1848-49. Plantes croissant spontanément dans les environs de Béthune. Annuaire du Pas-de-Calais : 203-243.
Métailié G., 1995. Lettrés jardiniers en Chine ancienne. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 37 (1) : 31-44.
Manneville O., 1988. Bilan phytosociologique et cartographie de la végétation appliquée à l’évaluation de la valeur biologique et à la gestion des marais tourbeux eutrophes de basse altitude : cas des bordures du lac d’Annecy (Haute-Savoie) et du marais de Lavour (Ain). Colloques phytosociologiques XV, Phytosociologie et conservation de la nature : 453-468.
Martin M.-A., 1971. Introduction à l’ethnobotanique du Cambodge. Atlas ethnolinguistique, série II, CNRS, Paris, 257 p.
Maubert Ph., 1988. Un exemple d’application de la phytosociologie à la définition d’une gestion conservatoire de milieux prairiaux en Brenne : la réserve naturelle de Chérine (Indre). Colloques phytosociologiques XV, Phytosociologie et conservation de la nature : 391-415.
Mauss M., 1989 (première édition en 1947). Manuel d’ethnographie. Payot, Petite Bibliothèque Payot P13, Paris, 264 p.
Meilleur B., 1984. Une recherche ethnoécologique en Vanoise. Travaux scientifiques du Parc national de la Vanoise 14 : 123-133.
Meilleur B., 1987. Des ethnosciences à l’ethnoécologie ou du rôle des représentations écologiques populaires dans les sociétés traditionnelles. Écologie humaine 5 (2) : 3-23.
Mercier M., 1990. La nature et le sacré. Éditions Dangles, Saint-Jean-de-Braye, 328 p.
Mérite É., 1942 (réédité en 2011). Les pièges : histoire et techniques de piégeage à travers le monde. Montbel, Paris, 343 p.
Michon G., de Foresta H. & Levang P., 1995. Stratégies agroforestières paysannes et développement durable : les agroforêts à damar de Sumatra. Natures, Sciences, Sociétés 3 (3) : 207-221.
Molet L., 1990a. Histoire du comput et de quelques calendriers. In J. Poirier, Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 181-264.
Molet L., 1990b. L’année sacrale, la fête et les rythmes du temps. In J. Poirier, Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 269-412.
Murru Corriga G., 1982. La tenderie aux grives en Sardaigne. Études rurales 87-88 : 115-129.
Pasquis R., 1998. Agroforesterie et développement durable en Amazonie. Bois et Forêts des Tropiques 258 (4) : 19-36.
Pélegrin J., Karlin C. & Bodu P., 1988. « Chaînes opératoires » : un outil pour le préhistorien. In J. Tixier (éd.), Technologie préhistorique (Notes et monographies techniques 25), CNRS, Paris : 55-62.
Peyron L. 1960. Les fleurs dans la philathélie mondiale. Journal d’agriculture tropicale et de botanique appliquée 7 (4-5) : 145-168.
Piaget J., 1967. Biologie et connaissance. Gallimard, Paris, 510 p.
Piot J., 1970. Pâturage aérien au Cameroun : utilisation des ligneux par les bovins. Revue d’élevage et de médecine vétérinaire des pays tropicaux XXXIII (4) : 503-517.
Poirier J., 1990. L’homme, l’objet et la chose. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 901-960.
Popper K.R., 1982. La logique de la découverte scientifique (édition française). Payot, Bibliothèque scientifique Payot, Paris, 480 p.
Portères R., 1949. Les arbres, arbustes et arbrisseaux conservés comme ombrage naturel dans les plantations de caféiers indigènes de la région de Macenta (Guinée française) et leur signification. Revue internationale de botanique appliquée et d’agriculture tropicale 321-322 : 336-355.
Portères R., 1962. De l’écobuage comme un système mixte de culture et de production. Journal d’agriculture traditionnelle et de botanique appliquée 19 (6-7) : 151-207.
Portères R., 1965. Le caractère magique originel des haies vives et de leurs constituants (Europe et Afrique occidentale). Journal d’agriculture traditionnelle et de botanique appliquée 12 (4-5) : 133-152, 253-291.
Pujol R. & Carbone G., 1990. L’homme et l’animal. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 1307-1388.
Rostand J., 1931. L’état présent du transformisme. Librairie Stock, Paris, 187 p.
Roussel J.-C., 1993. Timbres-poste : nouveautés à sujet forestier. Revue forestière française 45 (6) : 689-694.
Saintyves P., 1918. Le transfert des maladies aux arbres et aux buissons. Bulletin de la Société préhistorique française 15 : 296-300.
Sannier J.-M., 1985. Les métiers de la forêt d’Eu, la construction du carcahoux. Linguistique picarde 25 (2) : 1-8.
Saussure F. (de), 1915, réédition 1968. Cours de linguistique générale. Payot, Bibliothèque scientifique Payot, Paris, 520 p.
Scarpa G.F. & Arenas P., 2004. Vegetation units of the Argentine semi-arid Chaco: the Toba-Pilaga´ perception. Phytocoenologia 34 (1) : 133-161.
Seignobos Ch., 1980. Des fortifications végétales dans la zone soudano-sahélienne (Tchad et Nord-Cameroun). Cahiers de l’ORSTOM, série Sciences humaines, XVII (3-4), L’Arbre en Afrique tropicale, la fonction et le signe : 191-222.
Sélection du Reader’s Digest, 1978. Les mille visages de la campagne française, guide encyclopédique illustré. Paris, 628 p.
Sigaut F., 1991. Un couteau ne sert pas à couper, mais en coupant. Structure, fonctionnement et fonction dans l’analyse des objets. Actes XIes rencontres internationales d’archéologie et d’histoire, Antibes 1990 : 21-34.
Stehlé H., 1963. Les stades précurseurs, les associations végétales des bananeraies et leur valeur indicatrice culturale aux Antilles françaises. Journal d’agriculture traditionnelle et de botanique appliquée 10 (6-7) : 168-176.
Tchatat M., Puig H. & Tiki Manga Th., 1995. Les jardins de case des provinces du centre et du sud du Cameroun : description et utilisations d’un système agroforestier traditionnel. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 37 (2) : 165-182.
Toffin G., Meyer F., Jest C. & de Garine I., 1986. Salme, ethnologie et démographie d’un village tamang. In J.-F. Dobremez (dir.), Les collines du Népal central, écosystèmes, structures sociales et systèmes agraires, II – Milieux et activités dans un village népalais, INRA, Paris : 55-119.
Tubiana M.-J., 1969. La pratique actuelle de la cueillette chez les Zaghawa du Tchad. Journal d’agriculture traditionnelle et de botanique appliquée 16 (2-5) : 55-83.
Unesco, 2009. Le Patrimoine mondial de l’Unesco, le guide complet des lieux les plus extraordinaires. Éditions Ouest-France, Rennes, 832 p. (dernière édition mise à jour en 2018, avec 960 p.).
Valcke M.-A., 1989. L’homme et les plantes dans la région de Bouchavesnes-Bergen (département de la Somme). Thèse de doctorat en pharmacie, Lille, 258 p.
Valcke M.-A., 1994. La fougeraie du Benou, vallée d’Ossau, Pyrénées-Atlantiques. Bulletin de la Société linnéenne Nord-Picardie 12 : 39-40.
Valéry P., 1976. Cahiers, I. Gallimard, Bibliothèque de la Pléiade, Paris, 1 491 p.
Vaughan P., 1983. La fonction des outils préhistoriques. La Recherche 148 : 1226-1234.
Verheggen F., Bogaert J. & Haubruge E. (coord.), 2013. Les vers à soie malgache, enjeux écologiques et socio-économiques. Les Presses universitaires de Gembloux, 326 p.
Verschuer C. (von), 1995. L’autre agriculture : les cultures sur brûlis dans le Japon ancien. Journal d’agriculture traditionnelle et de botanique appliquée, NS, 37 (2) : 129-164.
Vertès F., 1983. Contribution à l’étude phytosociologique et écologique des prairies et alpages en moyenne Tarentaise ; application à l’évaluation des potentialités fourragères de la vallée de Peisey-Nancroix. Thèse de docteur-ingénieur, Institut national agronomique Paris-Grignon, Paris, 167 p.
Ville de Bayeux, s. d. La Tapisserie de Bayeux (reproduction intégrale au 1/7e). Édition Artaud Frères, Carquefou, 58 volets.
Vivier M., 1970. Pour une définition ethnobotanique du bocage. Actes du colloque de Domfront, Chambre d’agriculture du Calvados, 20 p.
Walker A., 1952. Substances végétales utilisées comme appâts. Revue internationale de botanique appliquée et d’agriculture tropicale 32 : 395396.
Wattez J.-R. & de Foucault B., 2018. L’impact de la guerre de 1914-1918 sur la flore, la végétation et le fleurissement en Picardie. Journal de botanique de la Société botanique de France 83 : 75-82.
Yongo O.D., Ponibanda A.K. & de Foucault B., 2014. Les poisons de flèches utilisés par les Pygmées de la préfecture de la Lobaye (République centrafricaine). International Journal of Biological and Chemical Sciences 8 (3) : 1129-1139.
Zahan D., 1990. L’Homme et la couleur. In J. Poirier (dir.), Histoire des mœurs I, Encyclopédie de la Pléiade 47, Gallimard, Paris : 115-180.
Remerciements
à Catherine Brau-Nogué, Béatrice Morisson et Gérard Largier (Conservatoire botanique national des Pyrénées et de Midi-Pyrénées), ainsi qu’à Jean-François Le Nail (historien) pour leurs informations sur les chemins de troupeaux du Saltus ; à Pierre Coulot (président de la Société botanique d’Occitanie), qui a bien voulu se charger de la relecture de cet article assez éloigné de ses préoccupations professionnelles et associatives, l’une de ses remarques ayant alimenté l’annexe 2 (supra).


