
Ranunculus trilobus Desf., synthèse sur la répartition et l’écologie d’une espèce longtemps négligée en Occitanie
Title
Ranunculus trilobus Desf., summary of the distribution and ecology of a long-neglected species in Occitanie region (France)
Résumé
Ranunculus trilobus Desf. est une espèce ouest-méditerranéenne thermophile qui a probablement toujours été rare en France, et notamment sur l’arc méditerranéen continental. Après sa description en 1798 à partir d’échantillons du Maghreb et sa mise en évidence au xixe siècle sur le littoral atlantique, les Pyrénées-Orientales, le Var et la Corse, l’espèce est progressivement sortie de la flore supposée indigène de l’Occitanie, au point qu’en 2007, lorsque nous l’y rencontrons, aucune station n’y semble alors répertoriée. Les flores de référence supposent l’espèce absente ou occasionnelle au sein de l’ancienne région Languedoc-Roussillon et évoquent des confusions avec des formes à petites fleurs de Ranunculus sardous Crantz. Depuis, l’espèce a été répertoriée au sein de trois départements de la région Occitanie (Hérault, Aude, Pyrénées-Orientales), région au sein de laquelle un statut d’autochtonie est estimé très vraisemblable compte-tenu de l’écologie spécialisée de l’espèce et de l’aire étendue occupée. Cette note propose un état des connaissances sur l’écologie et la répartition de l’espèce en Occitanie.
Abstract
Ranunculus trilobus Desf. is a thermophilic western Mediterranean species which has probably always been rare in France, and in particular on the continental Mediterranean arc. After its description in 1798 from samples from the Maghreb and its discovery in the 19th century on the Atlantic coast, the Pyrénées-Orientales, the Var and Corsica, the species gradually left the supposedly native flora of the region. Occitanie, to the point that in 2007, when we met it there, no station seemed to be listed there. The reference floras assume the species to be absent or occasional within the former Languedoc-Roussillon region and evoke confusion with small-flowered forms of Ranunculus sardous Crantz. Since then, the species has been listed within three departments of the Occitanie region (Hérault, Aude, Pyrénées-Orientales), region within which a status of autochthony is considered very likely given the specialized ecology of the species and the extensive area occupied. This note proposes a state of knowledge on the ecology and distribution of the species in Occitanie.
1. Redécouverte de Ranunculus trilobus en Languedoc-Roussillon
Herborisant au nord de Sigean (littoral de l’Aude) en avril 2007 près du Grand Salin, c’est au niveau d’une vigne bien enherbée que nous rencontrons pour la première fois cette espèce. Nous ne l’identifions pas sur le moment mais avons déjà le sentiment que l’unique individu rencontré ne correspond pas vraiment à Ranunculus sardous. Une recherche, peut-être un peu trop succincte à l’époque, ne nous aura pas permis d’attribuer avec confiance ce spécimen à une espèce ; le seul témoignage de cette rencontre fut alors matérialisé, durant six ans, par l’unique photo prise ce jour-là (photo 1) et rangée dans nos archives sous la bannière potentielle R. trilobus. Ce binôme fut attribué sans conviction à partir de Flora Iberica (López González et.al., 1986) et de Flora Europaea (Tutin et al., 1964) car l’espèce semblait douteuse dans la région à l’époque ; on parlait de confusions probables avec des individus de R. sardous déviant des canons de son habitus normal. Cette photo, nous le savons désormais, expose un individu plutôt mal développé de l’espèce ; moins de 10 cm, deux inflorescences en fruit, peu de feuilles.

Le temps a passé jusqu’au printemps 2013 durant lequel nous herborisions sur les zones humides littorales de l’Aude et des Pyrénées-Orientales et où nous confirmions alors la présence de R. trilobus sur au moins cinq secteurs distants.
C’est une visite au nord de la commune de Villeneuve-de-la-Raho (Pyrénées-Orientales) qui en fut, en quelque sorte, le « point d’orgue », au niveau du lieu-dit Els Estanyots, connu déjà à l’époque pour abriter quelques espèces typiques de l’habitat singulier et patrimonial de « mare temporaire méditerranéenne » (code Eur 27 3170, pour les initiés) sous son faciès plus eutrophe. Y était alors signalée la présence de deux espèces protégées : Pulicaria vulgaris et Lythrum tribracteatum. Là, c’est la confirmation définitive devant un peuplement de renoncules annuelles mêlant sans ambiguïtés deux populations importantes appartenant à deux espèces différentes, R. sardous et R. trilobus. Les individus des deux espèces y sont très bien développés et y font à peu près la même taille (20 à 50 cm). Ils présentent un port assez similaire, mais R. trilobus semble plus floribond et ramifié à conditions égales. Des différences constantes et évidentes, grâce à cette syntopie ad hoc, sont alors aisément notées (photos 2 et 3) ; R. trilobus possède des fleurs bien plus petites, des feuilles caulinaires bipennatiséquées et des akènes à tubercules plus prononcés. Aucun individu intermédiaire n’est observé. Ces différences, nous le verrons par la suite, sont déjà énoncées dans la description de l’espèce en 1798 par René-Louiche Desfontaines dans son Flora Atlantica, ouvrage qui couvre les régions atlasiques algériennes et tunisiennes.
Au printemps 2013, probablement une année favorable à l’espèce (précipitations hivernales abondantes), nous répertorierons ainsi cinq autres stations s’échelonnant de Sérignan à Villeneuve-de-la Raho, sans rencontrer aucune difficulté d’identification. La même année, nous faisons part au CBN med de notre observation (J. Molina) qui confirmera bien l’identification sur l’une des stations visitée (Sérignan, La Garenque).
A la sortie, en 2014, des deux ouvrages majeurs de la flore de France, Flora Gallica (Tison & de Foucault, 2014) et la « Flore med » (Tison et al., 2014), nous avons été confortés dans l’idée que cette espèce avait été longtemps négligée par le traitement qui en était fait. Cette espèce était, en effet, répertoriée comme occasionnelle sur l’arc méditerranéen français. Nous pensions, au contraire, par son lien avec les anciens étangs/paluds naturels et la proximité de son aire d’indigénat (Catalogne espagnole), que son autochtonie apparaissait probable en Languedoc-Roussillon. La découverte d’autres stations depuis (cinq autres) appuierait également, selon nous, cette hypothèse bien que l’espèce demeure rare et localisée.

2. Présentation de la renoncule à trois lobes Ranunculus trilobus
Ranunculus trilobus Desf. (R. trilobus) est une espèce ouest-méditerranéenne annuelle de la famille des Ranunculaceae, généralement ramifiée dès la base et formant une touffe assez lâche de tiges de longueur moyenne (10 à 50 cm). La floraison débute en avril et se prolonge en mai en Occitanie (dates extrêmes personnellement observées : 6 avril – 1er juin). Elle peut vraisemblablement varier suivant les températures et les dates d’assèchement du substrat. La phénologie de floraison renseignée dans la flore ibérique – de janvier à juillet – est ainsi très large (López González et. al., 1986).
Elle est facilement identifiable et bien caractérisée d’une part par la taille de ses pétales, pratiquement sans recoupement avec R. sardous, critère mis en avant par les auteurs ibériques (Lopez Gonzales et al, 1986), et d’autre part par l’ornementation des akènes, à tubercules décelables à l’œil nu, contrairement à R. sardous. Ces différences sont très bien renseignées dans la plupart des flores récentes et notamment dans Flora Gallica (Tison & de Foucault, 2014). Le critère des feuilles caulinaires bipennatiséquées, d’abord plus délicat, est seulement patent sur des exemplaires bien développés (photo 3).

Elle appartient à la section Echinella DC. qui regroupe des espèces partageant un type biologique annuel et des fruits comprimés et ornementés d’appendices censés les adapter à une dispersion ectozoochore. Ce groupe inclut notamment, pour le territoire français, R. arvensis, R. muricatus, R. parviflorus, R. chius entre autres. En fait, de récentes études phylogénétiques montrent au moins, même si tous les liens ne sont pas résolus, une paraphylie probable de cette section avec notamment R. parviflorus et R. chius, qui se rangeraient plutôt avec R. acris dans une section Ranunculus, et R. arvensis qui serait largement divergent avec une section pratiquement monospécifique (Hörandl et al., 2005). Ces mêmes études montrent en fait que R. trilobus et R. sardous seraient assez éloignées avec, en considérant seulement les espèces françaises, la première plus proche de R. repens et la seconde de R. bulbosus et R. muricatus. Des études cytotaxonomiques (Diosdado & Diaz, 1993) montrent également des différences notables avec R. sardous seule diploïde du groupe (2n = 16) et R. trilobus qui serait en majorité hexaploïde par rapport au nombre de base (n = 8). Au niveau mondial, R. trilobus est clairement un proche parent de deux espèces est-méditerranéo-touraniennes, R. cornutus et R. marginatus. Un phénomène de vicariance géographique est même tout à fait probable lorsque l’on observe les répartitions potentielles des deux espèces R. marginatus et R. trilobus (GBIF Secrétariat, 2022). Il serait alors intéressant d’étudier plus finement les relations génétiques (existence d’hybridation) entre ces espèces afin de les traiter au rang taxonomique pertinent. Ainsi, si R. trilobus devait être mis au rang subspécifique, ce qui a été le cas sous la bannière de R. sardous jadis, ce serait plutôt sous son propre binôme R. trilobus subsp. trilobus, aux côtés des taxons cornutus ou marginatus. Par ailleurs, une espèce répandue en Afrique, R. multifidus, ferait office de groupe-frère du groupe de R. trilobus. En examinant rapidement quelques photos de cette plante, qui nous est inconnue, glanées sur internet, la ressemblance apparaît frappante, renforçant, s’il en était besoin, l’improbabilité de rapprochement de R. trilobus et R. sardous. Les analyses phylogénétiques récentes consacrées aux Ranunculus au niveau mondial (Hörandl et al., 2005) font l’hypothèse intéressante d’une apparition convergente du caractère annuel chez R. trilobus et R. sardous à partir de deux ancêtres distincts et vivaces : un ancêtre partagé avec R. multifidus pour le groupe de R. trilobus (R. trilobus, R. marginatus et R. cornutus) et un ancêtre partagé avec R. bulbosus pour R. sardous.
3. Traitement diachronique du taxon par les flores et herbiers
Comme évoqué, la description valide de ce taxon au rang spécifique a été effectuée par René Louiche Desfontaines, dans sa Flora Atlantica parue en 1798, à partir de plantes provenant d’Algérie (Desfontaines, 1798). Cette description est retranscrite ici :
« Renoncule à tiges dressées, feuilles glabres, les caulinaires trilobées, pédoncules striés, petites fleurs, graines comprimées et tuberculées. Affine de Ranunculus parviflorus, en diffère par ses tiges dressées. Habite les champs humides, près de Mayane ».
Cette localité, aujourd’hui appelée Miliana, se situe en Algérie, à une centaine de kilomètres au sud-ouest d’Alger. Coincée entre deux montagnes, la ville domine effectivement une vaste plaine céréalière (vallée du Chélif) dont on peut supposer le potentiel inondable, avec formations de mouillères en saison froide.
Cette description de la plante, concise et efficace, appelle selon nous une seule remarque : la comparaison à R. parviflorus, auquel elle ressemble plutôt vaguement et par des caractères convergents (espèce éloignée phylogénétiquement).
Dans son Histoire abrégée des plantes des Pyrénées, Picot de Lapeyrouse (1813) cite la plante pour la première fois officiellement comme présente sur le territoire national. L’auteur cite la présence de R. trilobus dans le secteur de Perpignan et la différencie de R. philonotis (i.e. R. sardous), également présente dans la dition concernée par l’ouvrage. Malheureusement, en 1818, dans les suppléments et addenda à cet ouvrage, il décrit pour la première fois le taxon xatartii au rang spécifique sous le binôme Ranunculus Xatardi. La description de ce nouveau taxon apparaît assez douteuse :
« R. seminibus tuberculato-punctatis ; foliis pinnatis, pinnis trilobis laciniatis acutis ; caule diffuso. Fleurs jaunes, grandes, brillantes. A Paulillas de Bagnols. Pyrén.-Orient. M. Xatard. 1815. Cette renoncule se rattache à celle à petites fleurs, elle a le même port, les mêmes formes, les mêmes proportions ; mais ses fleurs sont plus grandes, et elle n’a sur sa tige que quelques poils rares. Les différences les plus marquées se trouvent sur les feuilles. Elles sont pennées ; les pinnules sont brièvement pétiolées, et composées, ainsi que l’impaire, de folioles simples, trilobées, à déchirures aiguës. On y reconnaît le type de feuilles simples du R. parviflorus. »
En fait, la description pourrait bien correspondre à un petit exemplaire de R. trilobus qui a bien des fruits tuberculés et des fleurs que l’on peut considérer comme grandes, mais effectivement seulement par rapport à R. parviflorus. À défaut de visualiser cette part d’herbier, un doute subsistera sur l’identité de cette plante. Nous verrons que ce taxon xatartii aura surtout empoisonné, par la suite, la bonne appréhension des phénotypes propres aux deux espèces, trilobus et sardous. Quant à la comparaison, ou le lien, avec R. parviflorus, il ne semble plus avoir été repris par la suite, à juste titre.
Dans la Flore de Grenier & Godron (1847), R. trilobus apparaît encore au rang d’espèce, aux côtés de deux autres proches, R. philonotis (i.e. R. sardous) et R. cordigerus. Aucune variation notable ni sous-espèce n’y sontt évoquées. Le taxon R. xatartii y est synonymisé à raison avec R. trilobus. Il est assez amusant de constater que ce traitement global (systématique, description, répartition), vieux de près de deux siècles, concorde peu ou prou avec l’actuel. C’est, semble-t-il, à partir de la fin du xixe siècle que les choses se gâtent pour notre espèce…
Dans la Flore de Rouy & Foucaud (1893), R. trilobus apparaît comme une sous-espèce de R. sardous aux côtés de quatre autres, les subsp. sardous, xatartii, cordigerus et rhoeadifolius. Cette dernière est agrémentée du qualificatif « prospecies » qui invite les botanistes à la décrire officiellement. Considérant également un nombre important de variétés décrites dans cet ouvrage et d’autre part de probables confusions dans les localisations des stations, on imagine dès lors la perplexité du botaniste amateur de l’époque et surtout la probabilité importante pour qu’il n’attribue pas un échantillon donné à l’un des trois taxons aujourd’hui reconnus et bien délimités : sardous, cordiger ou trilobus.
Dans la Flore de Coste (1937), le traitement est digne des lumpers les plus zélés (i.e. ceux qui ont une conception large de la variabilité de l’espèce) avec l’évocation succincte de deux variétés de R. sardous, la var. trilobus et la var. cordiger. La négligence de R. trilobus commence à s’opérer en France continentale.
Dans la Flore d’Afrique du Nord de Maire (1967), le taxon est également traité au rang subspécifique, mais il y est seulement appairé à un autre taxon. Deux variétés (var. fontanesii et rhoeadifolius) sont décrites pour R. sardous trilobus et trois variétés, dont le fameux taxon xatartii, sous la bannière de R. sardous philonotis (i.e. R. sardous). La confusion devient patente et totale car la subsp. xatartii aurait dû « logiquement » (si elle existait réellement ?) être subordonnée à la subsp. trilobus compte-tenu de sa distribution (Argelès-sur-Mer) et de sa description alors connue reprise de la Flore de France de Rouy & Foucaud (1893).
En ce qui concerne les flores régionales, la Flore des Pyrénées-Orientales de Gautier (1898) se doit d’être citée ici car elle inclut vraisemblablement, et à l’époque du moins, les stations connues les plus pérennes de R. trilobus de France continentale. Le traitement taxonomique dissocie bien au rang spécifique R. trilobus de R. sardous en décrivant une situation encore observable actuellement, notamment lorsqu’il est fait référence aux effectifs des deux taxons (espèces données comme communes) et à leur syntopie récurrente. Cependant, elle fait mention d’une var. xatartii, très rare et représentée à Argelès, qu’elle subordonne à R. sardous, probablement par transposition simple du traitement de Rouy & Foucaud (1893) qui en faisait une sous-espèce de ce même taxon. Le traitement pertinent de R. trilobus dans ce catalogue départemental n’est vraisemblablement pas sans lien avec le fait que les deux espèces y soient syntopiques et relativement communes.
À ces traitements taxonomiques parfois très descriptifs et qui nous semblent aujourd’hui assez rébarbatifs, s’ajoutent très vraisemblablement, et sans vouloir faire offense à nos illustres et anciens botanistes, de très probables erreurs d’identification. C’est ce que nous avons pu constater en effectuant quelques vérifications sur un échantillonnage modeste (21 parts facilement accessibles via le site du GBIF avec un filtre sur la localité : France). Près d’un quart devrait être vraisemblablement plutôt attribué à R. sardous. C’est notamment le cas pour des exsiccata montpellierains (University of Oslo, 2023) qui semblent provenir du classique lieu d’herborisation local des collecteurs, la fameuse « mare de Grammont » ! Deux planches provenant de Corse (Asco) sont plus délicates à assigner de manière univoque à R. sardous, néanmoins leur habitus en semble plus proche que de R. trilobus (Bijmoer et al., 2022). Enfin, une planche provenant d’Ampus (Var) semble également se rapporter à R. sardous (MNHN & Chagnoux, 2023). En revanche, deux exemplaires de Toulon et un d’Antibes sont bien conformes à R. trilobus. De récentes données d’observation (https://nature.silene.eu/espece/117257) semblent confirmer la présence probablement ancienne et continue de l’espèce dans la région toulonnaise qui abrite pratiquement les derniers paluds littoraux (souvent cultivés depuis longtemps) vers l’est. Ce type d’habitat est conforme au préférendum écologique de l’espèce et pourrait donc signifier un indigénat potentiel, conjugué à une rareté originelle, sur le territoire provençal.
Les autres parts d’herbiers se rapportent en majorité aux stations connues jadis (fin xixe au début du xxe siècle) et proviennent pratiquement toutes d’Argelès-sur-Mer (Pyrénées-Orientales), lieu de récolte privilégié de la subsp. ou var. xatartii. Selon nous, ils correspondent sans équivoque à R. trilobus.
4. Écologie et répartition actuelles de l’espèce à différentes échelles
Il s’agit d’une espèce répertoriée essentiellement au niveau de marais doux, en général, et de mouillères (mare de champ cultivé), en particulier. Ce penchant pour les espaces cultivés inondables est notoire en Espagne où l’espèce est commune notamment dans le Sud-Ouest, bien arrosés au printemps et dont les espaces cultivés de plaine sont constellés de vastes mouillères (photo 4). C’est également l’habitat qui est évoqué pour le Maroc ou l’Algérie, pays au sein duquel l’espèce est commune. Le Maroc atlantique offre notamment de vastes espaces de marais alluviaux temporaires de plaine au sein desquels l’espèce est commune et qui constitue sans doute l’habitat originel ou primaire de l’espèce. Dans la zone tyrrhénienne sensu lato (Corse, Sardaigne, Sicile, Italie continentale, etc.), l’espèce serait également répertoriée au niveau de pelouses humides (Gamisans, 1999) et de mares temporaires méditerranéennes (Tomaselli et al., 2020).

En France, La Flore des champs cultivés (Jauzein, 1995) mentionne bien cette espèce comme adventice rare au rang subspécifique R. sardous subsp. trilobus. Flora Gallica indique comme habitats les friches méditerranéennes hygrophiles. Nos observations peuvent se ranger effectivement dans ce type d’habitat, mais elles permettent, selon nous, d’apporter quelques précisions sur la partie languedocienne de sa distribution. Nous avons rencontré des stations de cette espèce au sein de vignes (deux stations), prairies pâturées avec drains (deux stations), culture de blé dur (trois stations), jachères (deux stations). Le point commun entre ces stations est le caractère inondable et pionnier des habitats, ainsi qu’un caractère non oliogotrophile souvent accusé, justifiant bien le qualificatif de « friche ». Cependant, la plupart des stations ne possèdent parfois pas pour autant un caractère eutrophile très marqué, avec des plantes compagnes telles que Lythrum tribracteatum, ou L. thymifolia (Els Estanyots, Villeneuve-de-la-Raho ; La Garenque, Sérignan). La texture argileuse semble assez constante également, au niveau de substrats tant acides (alluvions villafranchiennes) que basiques (anciens étangs).
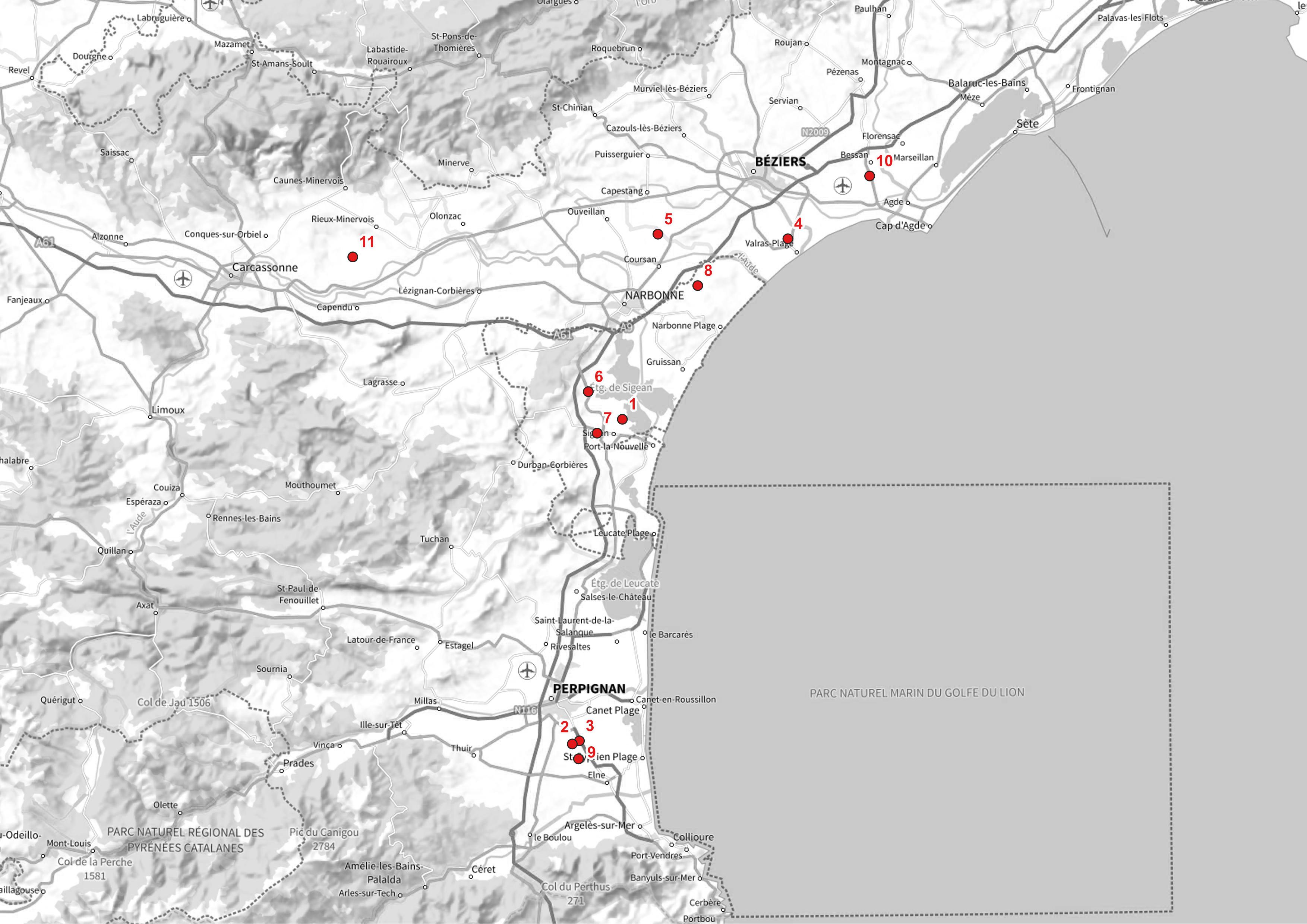
Au-delà du biotope, au niveau du paysage, la localisation régionale apparaît de fait fortement dépendante des anciens étangs temporaires nichés dans les dépressions éoliennes et qui sont assez nombreux en Languedoc. Sur les onze stations exposées dans le présent article (carte 1), huit sont en effet liées à d’anciens étangs. Les autres sont tout de même au sein de mouillères sur des terrasses alluviales anciennes (Bessan, Sérignan), trahies souvent dans la toponymie par des termes suggestifs (Estang, Estagnols, Estanyots, etc.). L’hypothèse d’un indigénat de la plante en Languedoc au sein de ces milieux, tout à fait conformes à ceux supposés primaires et observés, notamment en Espagne et au Maroc, se fait alors jour. La majorité de ces anciens étangs ou paluds naturels a été largement drainée, asséchée et mise en culture dès le Moyen Âge. Néanmoins, la plupart de ces dépressions présente des mises en eau partielles, mais régulières, qui conviennent encore à cette espèce, en quelque sorte préadaptée à la mise en culture de jadis. L’hypothèse alternative, dans la distribution fragmentée actuelle de l’espèce, d’une colonisation récente de ces anciens étangs après leur mise en culture nous semble moins vraisemblable, mais demeure possible compte tenu du pouvoir colonisateur supposé important de l’espèce au sein des espaces cultivés humides : production de semences importante, épizoochorie potentielle (Lone et al., 2014). Le statut d’archéophyte ibérique, adventice de cultures, à l’image d’autres espèces comme Nigella hispanica var. hispanica, demeure donc une hypothèse possible.
La répartition actuellement connue de l’espèce couvre les secteurs de plaine du bassin méditerranéen occidental avec un barycentre situé sur la zone atlanto-méditerranéenne (Maroc atlantique, Andalousie, Portugal) et des poches importantes à l’est au niveau des secteurs bien arrosées (bioclimat subhumide à humide) : îles, montagnes maghrébines. Des mentions de l’espèce dans l’est du bassin (peu nombreuses semble-t-il) mériteraient des vérifications car des confusions avec l’espèce vicariante Ranuculus marginatus nous semblent probables, même si nous ne connaissons pas cette espèce in natura. Dans ce cadre, le Languedoc constitue un prolongement classique et naturel des stations catalanes qui sont également localisées aux vallées des grands fleuves et aux marais littoraux.
En France, l’espèce est peu répandue, sauf en Corse où elle est donnée commune par Gamisans et traitée au rang subspécifique R. sardous subsp. trilobus (Gamisans, 1999). Sur le continent, deux régions de présence se détachent, le littoral centre-atlantique et la partie occidentale de l’arc méditerranéen.
Dans le domaine atlantique, les zones occupées sont assez ténues et bien localisées aux secteurs de grands marais du Médoc et des deux îles charentaises Oléron et Ré. L’espèce est même protégée au niveau régional dans l’ancienne région Poitou-Charentes.
Dans la zone méditerranéenne, l’espèce est plus répandue car son aire s’étend actuellement des Pyrénées-Orientales au Var, avec cependant un hiatus important centré sur l’est de l’Hérault et la Camargue. Nous pouvons néanmoins la considérer comme une espèce rare et localisée au sein des trois départements qui regroupent actuellement les plus grandes populations : Hérault (ouest), Aude et Pyrénées-Orientales. Nous avons personnellement répertorié dix stations depuis 2007, localisées au sein de huit communes. Ces stations sont les suivantes (les numéros sont reportés sur la carte 1) ordonnées par date d’inventaire :
- nord du Grand Salin, 25/04/2007, Sigean (Aude) : vigne (aujourd’hui en friche) ;
- lieu-dit Els Estanyots, 06/04/2013, Villeneuve-de-la-Raho (Pyrénées-Orientales) : culture inondable (mouillère) abritant des végétations de friches hygrophiles et de mares temporaires méditerranéennes mésotrophiles ; espèces représentées dans le même biotope : Ranunculus sardous, Lythrum tribracteatum, thymifolium, Pulicaria vulgaris, etc. (photo 5) ;
- lieu-dit Les Salleses, 06/04/2013, Villeneuve-de-la-Raho (Pyrénées-Orientales) : friche post-culturale inondable (mouillère) abritant des végétations de friches hygrophiles ;
- lieu-dit La Garenque, 05/05/2013, Sérignan (Hérault) : mare temporaire méditerranéenne (sur friche ou jachère très ancienne) sur alluvions villafranchiennes, abritant une belle population de Lythrum thymifolium, en compagnie de quelques pieds de Polygonum romanum et Ranunculus muricatus;
- partie sud de l’étang de Capestang au nord du canal de l’Aiguille de Londres, 13/05/2013, Coursan (Aude) : cultures et jachères inondables (mouillères) abritant des végétations de friches hygrophiles et de mares temporaires méditerranéennes (Lythrum tribracteatum, Crypsis schoenoides ; association du Lythro tribracteati-Crypsietum aculeatae Foucault 2021 ; de Foucault, 2021) ;
- lieu-dit Les Venderelles, 15/05/2013, Peyriac-de-Mer (Aude) : ancien étang dans une dépression éolienne, culture de blé inondable ;
- étang de Sainte-Croix, 09/05/2018, Sigean (Aude) : ancien étang dans une dépression éolienne ; plusieurs secteurs occupés dans l’ancien étang, dont une vigne ;
- étang de Taraillan, 01/06/2020, Fleury/Vinassan (Aude) : ancien étang dans une dépression éolienne ; plusieurs secteurs occupés dans l’ancien étang dont vignes, fossés inondables, cultures de blé ;
- lieu-dit Val Marie, 14/04/2021, Villeneuve-de-la-Raho (Pyrénées-Orientales) : ancien palud aujourd’hui occupé par des pâtures à chevaux ; secteur qui appartient au vaste système de marais du Prat de la Vila;
- lieu-dit Les Estagnols, 23/06/2022 (identifié sec et en fruit), Béssan (Hérault) : mare temporaire méditerranéenne (sur friche ou jachère très ancienne) sur alluvions anciennes, abritant une belle population de Pulicaria vulgaris. Les autres espèces sont : Polygonum romanum, Dittrichia viscosa, Symphyotrichum squamatum, Polypogon maritimus, Rumex criqpus, etc.

À ces stations qui nous sont directement connues de visu, s’ajoute celle récemment découverte près de l’étang de Marseillette par notre confrère Christophe Savon (Les Arjalats, 5 mai 2022, station 11 sur la carte 1). Cette petite station (moins de dix pieds) se situe au niveau d’une friche inondable à scirpe maritime. Elle étire loin vers l’ouest la zone de présence potentielle de l’espèce, mais elle demeure dans les limites connues du pays des grands étangs/paluds languedociens.
Au niveau mondial, l’adventice Ranunculus trilobus a d’ores et déjà bien engagé sa colonisation d’autres territoires éloignés de son aire d’origine puisqu’elle est bien présente dans le sud-est des États-Unis (littoral du Texas, Alabama, Louisiane, Floride et Caroline-du-Sud) et le sud de l’Australie (région de Perth à l’ouest et entre Adélaïde et Melbourne, au sud-est). En Eurasie, elle est signalée jusque dans l’Himalaya (Lone et al., 2014), ainsi que ponctuellement en Chine. Elle demeure donc dans la zone tempérée chaude, mais non strictement sous climat méditerranéen, notamment aux États-Unis et en Chine, dans des secteurs soumis plutôt au climat cantonnais (ou chinois) typique des masses continentales orientales situées aux latitudes concernées. Aussi, bien que probablement originaire du bassin ouest-méditerranéen, l’espèce semble montrer également un tropisme vers les climats non strictement méditerranéens à l’échelle mondiale. Son origine atlanto-méditerranéenne (ou méditerranéo-atlantique) nécessitant des précipitations importantes en saison froide l’exclurait potentiellement de climats méditerranéens à saison sèche trop marquée et à hivers froids. À noter qu’elle semble bien représentée également dans la région macaronésienne, sauf sur les deux îles les plus arides, Fuerteventura et Lanzarote.
5. Conclusion
Ranunculus trilobus demeure une espèce rare et localisée dans la zone méditerranéenne de France continentale. Elle est répertoriée à ce jour sur quinze communes à l’échelle régionale (interface du SINP Occitanie, consultée le 16/01/2023) : neuf dans les Pyrénées-Orientales (communes de la plaine du Roussillon), quatre dans l’Aude et deux dans l’Hérault. Bien que considérée antérieurement comme occasionnelle, ou potentiellement naturalisée, nous pensons que l’hypothèse d’un indigénat y est très vraisemblable. Cette hypothèse apparaît également crédible pour l’aire atlantique de cette répartition française. En effet, son aire de répartition actuelle méditerranéo-atlantique, même fragmentée, demeure concordante avec de nombreuses espèces thermophiles, notamment parmi les taxons méditerranéens possédant des besoins hydriques plus importants en période de végétation printanière, dont les espèces de mares temporaires. En guise d’illustration, citons, par exemple, plusieurs espèces de Lythrum, Damasonium alisma, Elatine macropoda, Exaculum pusillum, Isoetes histrix, Isoetes velata, etc. Par ailleurs, le taxon est connu depuis plus de deux siècles à la fois dans la région de Bordeaux et celle du Roussillon. Enfin, les stations catalanes françaises sont finalement peu éloignées de celles de Catalogne espagnole, également localisées (espèce rare d’après de Bolòs & Vigo, 1995) mais qui possèdent une écologie conforme : grandes vallées alluviales, marais littoraux, mares temporaires.
Plusieurs facteurs semblent pouvoir expliquer la paradoxale omission de cette espèce indigène en Languedoc-Roussillon durant plusieurs décennies, alors que la région héberge potentiellement ses plus grands effectifs français :
- la désuétude des ouvrages de référence conjuguée à leur traitement souvent inadéquat du taxon, comme exposé au-dessus ;
- la rareté et la localisation de l’espèce en France en lien avec ses exigences écologiques notoires (espèce thermohygrophile amphibie mésotrophile) ;
- l’irrégularité du développement de la plante en zone méditerranéenne, à l’instar d’autres plantes inféodées aux mares temporaires ;
- les confusions entretenues par les anciens auteurs qui attribuaient à cette espèce des formes un peu singulières, notamment en ce qui concerne le découpage des feuilles caulinaires de sardous;
- la description du taxon xatartii par Lapeyrouse ; notre sentiment est qu’en lisant le descriptif un botaniste de terrain peut se dire que Lapeyrouse s’est trompé d’affinité et qu’il s’agit plutôt d’un trilobus, mais à grandes fleurs, ce qui fait sortir l’espèce du cadre de sa variabilité et incite à garder, au bénéfice du doute, un taxon chimère (ou hybridogène, au mieux), c’est-à-dire un R. trilobus à fleurs de R. sardous;
- l’aspect effectif de la plante qui, lorsqu’elle n’est pas bien développée, ressemble à un sardous d’aspect malingre, assez facile à négliger. A contrario, lorsqu’elle bénéficie de conditions favorables (inondations de ses stations qui se prolongent au début du printemps), les tiges étalées dépassent facilement 40 à 50 cm et le caractère « pétale court » ne l’assimile alors pas à une R. sardous à pétales un peu avortés.
Sur le statut taxonomique de l’espèce, sa distinction au rang spécifique par rapport à R. sardous semble appuyée compte tenu :
- des niveaux de ploïdie différents qui, s’ils sont exacts (des comptages divergent selon les dates et échantillons avec de potentiels problèmes d’identification) et assez constants sur l’ensemble de l’aire, rendraient délicate l’hybridation potentielle simple de type F1 entre les deux taxons ;
- des études phylogénétiques disponibles, qui placent les deux espèces dans des clades proches mais bien différenciés avec deux évènements parallèles (convergence évolutive) de passage, à partir de deux ancêtres vivaces distincts, vers le type biologique annuel ;
- de la rareté des individus dont on peut supposer une origine hybride entre les deux espèces. Nous n’en avons personnellement encore jamais soupçonné. Les seules mentions de ce type que nous avons répertoriées au sein de toute l’aire de trilobus émanent en majorité des écrits relatifs aux stations des Pyrénées-Orientales ; nous ne connaissons pas la station actuelle mentionnée comme abritant des individus atypiques de R. sardous et dont il est supposé que certains individus puissent être d’origine hybride avec R. trilobus (Schwab, 2020). L’existence d’hybrides reste possible mais demeure très hypothétique et sera difficile à asseoir sans recherches systématiques de terrain à large échelle ou sans analyses génétiques.
En l’état actuel de nos connaissances et des faits exposés, R. trilobus reste une espèce bien individualisée, assez exigeante et potentiellement menacée. Elle accompagne souvent des espèces protégées patrimoniales liées aux mares temporaires méditerranéennes. La responsabilité de la région Occitanie dans sa préservation pourrait donc apparaître importante. Une évaluation de son statut de menace aux échelles nationales et régionales reste à effectuer.
Bibliographie
Bijmoer R., Scherrenberg M. & Creuwels J., 2022. Naturalis Biodiversity Center (NL) – Botany. Naturalis Biodiversity Center. Occurrence dataset https://doi.org/10.15468/ib5ypt accessed via GBIF.org on 2023-01-11. https://www.gbif.org/occurrence/2516001203
Bolòs O. (de) & Vigo J., 1995. Flora dels Països catalans. Editorial Barcino, Barcelona, 1 227 p.
Coste H.,1937. Flore descriptive et illustrée de la France, de la Corse et des contrées limitrophes, I. Librairie des sciences et des arts, Paris.
Desfontaines R. L., 1800. Flora Atlantica, sive Historia Plantarum, quae in Atlante, Agro tunetano et algeriensi crescunt, 1. Blanchon.
Diosdado J.C. & Díaz J.E.P., 1993. Citotaxonomía de las especies anuales de Ranunculus, con aquenios ornamentados y hojas divididas, en la Península Ibérica. Lagascalia 17 (1), 71-87.
Foucault B. (de), 2021. Contribution à la connaissance de la végétation des étangs asséchés et des canaux du Narbonnais. Carnets botaniques 38, 1-15 + 3 tableaux ; https://doi.org/10.34971/387K-6576.
Gamisans J., 1999. La végétation de la Corse. Edisud, Aix-en-Provence.
Gautier G. 1898. Catalogue raisonné de la flore des Pyrénées-Orientales. Société agricole, scientifique et littéraire des Pyrénées-Orientales, Perpignan.
Grenier M. & Godron M. 1847. Flore de France, ou description des plantes qui croissent naturellement en France et en Corse, I. Librairie de l’Académie royale de médecine, Paris.
Hörandl E., Paun O., Johansson J.T., Lehnebach C., Armstrong T., Chen L. & Lockhart P., 2005. Phylogenetic relationships and evolutionary traits in Ranunculus s.l. (Ranunculaceae) inferred from ITS sequence analysis. Molecular Phylogenetics and Evolution 36 (2) : 305-327.
Jauzein Ph., 1995. Flore des champs cultivés. INRA éditions, Paris, 898 p.
Jeanmonod D. & Gamisans J., 2007. Flora Corsica. Edisud, Aix-en-Provence, 921 p.
Maire R., 1952-1987. Flore de l’Afrique du Nord (Maroc, Algérie, Tunisie, Tripolitaine, Cyrénaïque et Sahara). Édit Lechevalier, Paris, 16 vol.
Lapeyrouse P. P. (de), 1813. Histoire abrégée des plantes des Pyrénées, et itinéraire des botanistes dans ces montagnes, et supplément, 1. Ed. Bellegarrigue.
Lapeyrouse P. P. (de), 1818. Supplément à l’Histoire abrégée des plantes des Pyrénées. Ed. Bellegarrigue.
Lone F.A., Dar G.H. & Wafai B.A., 2014. Ranunculus trilobus Desfontaines (Ranunculaceae), a new record for Northwest and Trans-Himalayas. Pleione 8 (1) : 171-177.
López González G., Cook C.D.K. & Grau J., 1986. Ranunculus L. In S. Castroviejo, C. Aedo, M. Laínz, F. Muñoz Garmendia, G. Nieto Feliner, J. Paiva & C. Benedí (eds.), Flora Iberica, Real Jardín Botánico, CSIC, Madrid, 1 : 279-371.
Rouy G. & Foucaud J., 1893. Flore de France, ou description des plantes qui croissent spontanément en France, en Corse et en Alsace-Lorraine, 1. Société des sciences naturelles de la Charente-Inférieure.
Schwab P., 2020. Sur la variabilité de Ranunculus sardous Crantz et Ranunculus trilobus Desf. dans les Pyrénées-Orientales. Bulletin de la Société mycologique et botanique de Catalogne nord 34 : 59-66.
Tison J.-M. & de Foucault B., 2014. Flora Gallica : Flore de France. Biotope, Mèze, xx +1 196 p.
Tison J.-M., Jauzein Ph. & Michaud H., 2014. Flore de la France méditerranéenne continentale. Naturalia publications, Turriers, 2 078 p.
Tomaselli V., Beccarisi L., Brullo S., Cambria S., Forte L., Minissale P. & Veronico G., 2020. Phytosociological research on temporary ponds in Apulia (southern Italy). Mediterranean Botany 41 (1) : 15-41.
Tutin T.G., Heywood V.H., Burges N.A., Valentine D.H., Walters S.M. & Webb D.A., 1964. Flora Europaea, 1 – Lycopodiaceae to Platanaceae. Cambridge University Press.
UICN France, FCBN, AFB & MNHN, 2018. La Liste rouge des espèces menacées en France, § Flore vasculaire de France métropolitaine. UICN, Paris, 12 p.
University of Oslo, 2023. Vascular Plant Herbarium, Oslo (O) UiO. Version 1.2060. Occurrence dataset https://doi.org/10.15468/wtlymk accessed via GBIF.org on 2023-01-11. https://www.gbif.org/occurrence/1095010938.
Sites internet
Global Biodiversity Information Facility (GBIF.org), 2023. GBIF Occurrence download [13/01/2023] https://doi.org/10.15468/dl.r2jjer.
MNHN, Chagnoux S., 2023. Herbarium specimens of Société des sciences naturelles et mathématiques de Cherbourg (CHE), version 70.291. Société des sciences naturelles et mathématiques de Cherbourg, occurrence dataset https://doi.org/10.15468/lmznjw accessed via GBIF.org on 2023-01-11. https://www.gbif.org/occurrence/1019755183.
Ranunculus marginatus d’Urv. in GBIF Secretariat (2022). GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 2023-01-13.
Ranunculus trilobus Desf. in GBIF Secretariat (2022). GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 2023-01-13.
Remerciements
Nos remerciements vont en direction de Madame Mélanie Olivera pour sa relecture attentive, ainsi qu’à Bruno de Foucault pour ses commentaires et améliorations.


