
Nouvelles approches phytosociologiques de la végétation africaine
Title
New phytosociological approaches on African vegetation
Résumé
Cette synthèse sur la végétation africaine abordée selon une approche moderne a permis d’aborder la végétation aquatique vivace (deux alliances nouvelles), la végétation littorale (une alliance), des mégaphorbiaies tropicales et oro-africaines (une alliance), diverses mares temporaires (une alliance) et des inselbergs (deux alliances), la végétation arborescente des forêts du mont Kenya (sept alliances, trois ordres et deux classes) ; une alliance et un ordre nouveaux se rapportent par ailleurs à la végétation arborescente des forêts tropicales de basse altitude. Enfin la considération de pelouses à Alchemilla et de fourrés afro-alpins a permis la définition de deux autres alliances.
Abstract
This overview of African vegetation using a modern approach has made it possible to deal with perennial aquatic vegetation (two new alliances), littoral vegetation (one alliance), tropical and oro-African tall herbs formations (one alliance), various temporary pools (one alliance) and inselbergs (two alliances), and tree vegetation in the forests of Mount Kenya (seven alliances, three orders and two classes); in addition, one new alliance and one new order relate to the tree vegetation of low-altitude tropical forests. Finally, the consideration of Alchemilla grasslands and Afro-Alpine thickets has led to the definition of two other alliances.
Dans la poursuite de l’exploration de la végétation afrotropicale initiée dans un article récent sur le Niger (de Foucault, 2024), on va reprendre ici d’autres publications phytosociologiques sur l’Afrique tropicale sous un angle méthodologique plus fin et plus moderne que celui suivi par leurs auteurs. La nomenclature des taxons a été mise à jour selon le référentiel Plants of the World Online (POWO). Pour information, au niveau des genres non représentés en Europe, on a indiqué dans le texte (pas dans les tableaux) la famille d’appartenance. Les taxons de présence inférieure ou égale à I n’ont pas toujours été repris dans les tableaux joints au texte.
1. La végétation aquatique
Quelques aspects de la végétation aquatique du Zaïre ont été publiés par Lisowski & Malaisse (1989). Si leur Ottelio cylindricae-Brasenietum schreberi est correctement décrit et typifié (leur tableau II ; à noter la présence de Mayaca baumii, Mayacaceae, seul représentant en Afrique d’un genre de quatre espèces plutôt néotropicales), leur tableau I est biologiquement hétérogène. On peut distinguer une composante thérophytique (présence sur 45 relevés) : Eriocaulon setaceum (Eriocaulaceae) IV, Rotala myriophylloides (Lythraceae) III, Stenops zairensis (Asteraceae) II, Hydrothauma manicatum (Poaceae) II, Trichanthecium parvifolium (Poaceae) II, Xyris capensis (Xyridaceae) II, Cyperus waillyi I, Najas baldwinii I, Lindernia conferta I, Sacciolepis myosuroides (Poaceae) I, Crepidorhopalon schweinfurthii (Linderniaceae) I, Persicaria strigosa I, Phyllanthus leonardianus (Phyllanthaceae) I, Leersia nematostachya +, Schoenoplectiella rhodesica + : Rotalo myriophylloidis-Eriocauletum setacei ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 9 du tableau I in Lisowski & Malaisse (1989, Groupements végétaux des mares et des anses calmes… hors texte), qui apparaît comme vicariant de la végétation européenne du Crassulo vaillantii-Lythrion borysthenici B. Foucault 1988. En conséquence, la composante vivace peut être renommée Bothrioclino monocephalae-Isolepidetum fluitantis ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 14 (donc le même que le type du « Rotalo myriophylloidis-Bothrioclinetum monocephalae » initial) du tableau I in Lisowski & Malaisse (1989, Groupements végétaux des mares et des anses calmes… hors texte) ; avec Isolepis fluitans, cette végétation oligotrophile paraît vicariante de celle de l’Hyperico elodis-Sparganion Braun-Blanq. & Tüxen ex Oberd. 1957.
De leur côté, du même pays, Mandango et al. (1996) évoquent sans relevé une végétation thérophytique colonisant des vases humides avec Eclipta prostrata, Rorippa madagascariensis, Ethulia conyzoides, Struchium sparganophorum (deux Asteraceae), Sphenoclea zeylanica (Sphenocleaceae), qui rappellent la végétation des Elatino triandrae-Cyperetalia fusci B. Foucault ex B. Foucault 2023 ; on notera notamment la présence d’un Rorippa vicariant de R. palustris, ce dernier bien inféodé à cet ordre tempéré.
2. Les roselières et prairies flottantes
Le tableau 1 synthétise un certain nombre de roselières afrotropicales, qui partagent notamment Phragmites australis, qui occupe une place importante dans les roselières européennes.
La colonne 2 décrit une prairie flottante à Vossia cuspidata (Poaceae ; Schnell, 1977 : figure 294 p. 210) et Echinochloa pyramidalis. L’une d’elles a été décrite par Germain (1965) du Congo et dénommée finalement Vossietum cuspidatae Robyns ex Germain 1965 (de Foucault, 2024 : 4). Celle évoquée ici, du Sénégal, en est assez distincte (celle du Congo est synthétisée en colonne 3 pour comparaison) : Echinochloo stagninae-Vossietum cuspidatae Trochain ex nov. hoc loco [syn. : groupement à Echinochloa stagnina-Vossia cuspidata Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 87)], typus nominis hoc loco : relevé 13 du tableau V in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 89). On la reconnaît aussi parmi les données de Guillaumet & Adjanohoun (1971 : 188) provenant de Côte-d’Ivoire. Des côtes togolaises, de Foucault et al. (1999, tableau 3 : relevé 11) publient un relevé qui ne semble pas se rattacher à cette association. Les données de Sillans (1958 : 157) provenant de République centrafricaine sont trop imprécises pour décider à quel Vossietum cuspidatae les rattacher. Quoi qu’il en soit, ces deux associations à Vossia cuspidata peuvent être rapprochées dans une alliance nouvelle, l’Echinochloo pyramidalis-Vossion cuspidatae all. nov. hoc loco, caractérisée par Vossia cuspidata, Echinochloa pyramidalis, Urochloa mutica, typus nominis hoc loco : l’Echinochloo stagninae-Vossietum cuspidatae Trochain ex B. Foucault.
La colonne 4 est une roselière sénégalaise à Typha domingensis décrite aussi par Trochain (1940 : 95) en tant que groupement à T. australis qu’on peut élever au rang d’association comme Cypero divitis-Typhetum domingensis Trochain ex hoc loco, typus nominis hoc loco : relevé 8 du tableau VII in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 98).
Les colonnes 5 et 6 décrivent plutôt des savanes hygrophiles à Chrysopogon nigritanus (= Vetiveria nigritana, Poaceae). Laissant de côté celle du Sénégal assez peu caractérisée (colonne 5), la colonne 6 est le Vetiverietum nigritanae Adjanohoun 1964 (Mém. ORSTOM 7 : 101), muté ici en Chrysopogonetum nigritani Adjanohoun 1964 nom. mut. hoc loco, lectotypus nominis hoc loco : relevé 3 du tableau X in Adjanohoun (1964, Mém. ORSTOM 7 hors texte). Des côtes togolaises, de Foucault et al. (1999, tableau 5) publient deux relevés à Ch. nigritanus qui ne semblent pas non plus se rattacher à cette association
Les colonnes 7 à 10 correspondent à des savanes hygrophiles oligohalophiles de lagunes littorales à Panicum repens et Paspalum vaginatum. En particulier la colonne 8, des côtes sénégalaises, est le groupement à Paspalum vaginatum Trochain 1940 (Mém. Inst. Franç. Afrique noire 2 : 122), avec Gomphrena vermicularis (= Blutaparon vermiculare, Philoxerus vermicularis ; Amaranthaceae), qui peut être validé comme Gomphreno vermicularis-Paspaletum vaginati Trochain ex hoc loco, typus nominis hoc loco : relevé 1 du tableau X in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 123). La colonne 9 en est une forme vicariante du littoral congolais, mais statistiquement faiblement définie. La colonne 10, enfin, décrit une végétation différente reconnue sur le littoral congolais : Cypero polystachyi-Paspaletum vaginati Lebrun ex hoc loco [syn. : groupements à Paspalum vaginatum-Sporobolus robustus Lebrun 1969 et à Paspalum vaginatum-Panicum repens Lebrun 1969 (Mém. Acad. Roy. Sci. outre-mer, classe des sciences naturelles et médicales, n. s., 18 (1) : 95 et 102 respectivement)], typus nominis hoc loco : relevé 6 du tableau XXIV in Lebrun (1969, Mém. Acad. Roy. Sci. d’outre-mer, classe des sciences naturelles et médicales, n. s., 18 (1) : 103). Tout cet ensemble semble bien correspondre à une alliance caractérisée par Paspalum vaginatum, Panicum repens, Sporobolus robustus, Gomphrena vermicularis : Paspalo vaginati-Panicion repentis all. nov. hoc loco, typus nominis hoc loco : le Cypero polystachyi-Paspaletum vaginati Lebrun ex B. Foucault.
3. La végétation littorale
La végétation littorale des côtes africaines a fait l’objet d’assez nombreuses publications. Le tableau 2 synthétise certaines de ces données pour l’Afrique occidentale et permet de distinguer deux associations, avec cependant un syntaxon intermédiaire.
Les colonnes 1 à 9 décrivent une association caractérisée par Cyperus crassipes, Scaevola plumieri (Goodeniaceae), Sporobolus spicatus, observée sur les côtes sénégalaises (Trochain, 1940 ; Vanden Berghen, 1979, 1982, 1990) et ivoiriennes (Guillaumet & Adjanohoun, 1971 : 211). Vanden Berghen (1990) la décrit sous le nom valide, quoique peu expressif, de Schizachyrietum pulchelli. Cette association possède plusieurs variations significatives, mais il est difficile de les relier à celles de l’auteur qui distingue les typicum et diodietosum serrulatae (Diodia serrulata est appelé Hexasepalum serrulatum dans le tableau 2, Rubiaceae) :
- les colonnes 1 à 5 sont différenciées par Sporobolus spicatus, avec des variantes à Launaea brunneri (colonne 1, Asteraceae ; Sénégal septentrional), à Fimbristylis cymosa (5, Sénégal méridional) ;
- les colonnes 6 à 9 correspondent au pergularietosum daemiae subass. nov. hoc loco, différencié par deux Apocynaceae, Pergularia daemia et Leptadenia hastata, ainsi que Hexasepalum serrulatum et Ipomoea stolonifera, typus nominis hoc loco: relevé 6 du tableau 5 in Vanden Berghen (1979, Jard. Bot. Belgique 49 : 196).
Les colonnes 12 à 14 décrivent une association un peu différente de la précédente, avec Cyperus pedunculatus (= Remirea maritima), Sporobolus virginicus, S. robustus ; elle a déjà été décrite sous le nom de Remireo maritimae-Canavalietum roseae Lebrun 1969, nom qui peut être muté en Cypero pedunculati-Canavalietum roseae nom. mut. hoc loco et lectotypifié par le relevé 2 du tableau XII in Lebrun (1969, Mém. Acad. Roy. Sci. Outre-Mer 18 (1) : 69) ; elle présente deux variations à conforter, l’une différenciée par Stenotaphrum secundatum (colonne 13), la seconde par Hexasepalum serrulatum et Vigna unguiculata (14, incluant le type, donc à valeur de typicum). Cette association semble aussi présente sur le littoral de l’île Annonbon, en Guinée-Équatoriale (Schnell, 1977 : 147).
Les colonnes 10 et 11, correspondant au littoral du Bénin (Paradis, 1976) et du Togo (de Foucault et al., 2000), se placent comme des relais entre ces deux associations, empruntant Cyperus crassipes et Scaevola plumieri à la première, Cyperus pedunculatus et Sporobolus virginicus à la seconde. Dans la publication sur le littoral togolais, on avait choisi de la rattacher à celle-ci plutôt qu’au Schizachyrietum pulchelli ; il s’avère cependant qu’il s’agit d’une race originale du Cypero pedunculati-Canavalietum. On la reconnaît aussi chez Guillaumet (1967) de Côte-d’Ivoire, toutefois guère dans son texte, mais plutôt sur la photo 26.
Ces deux associations partagent Ipomoea pes-caprae (photo 1), I. stolonifera, Schizachyrium pulchellum (Poaceae), Canavalia rosea (Fabaceae), Euphorbia glaucophylla, Alternanthera littoralis var. maritima, certains taxons s’avérant plutôt caractéristiques de rang élevé (ordre ou classe : Ipomoea pes-caprae, I. stolonifera, Canavalia rosea), d’autres plus strictement inféodés aux plages afro-atlantiques (Schizachyrium pulchellum, Euphorbia glaucophylla, Alternanthera littoralis var. maritima). Elles peuvent être rapprochées dans une alliance originale, l’Euphorbio glaucophyllae-Ipomoeion pedis-caprae all. nov. hoc loco, caractérisée par la combinaison des taxons qui viennent d’être cités, typus nominis hoc loco : le Cypero pedunculati-Canavalietum roseae Lebrun 1969 (Mém. Acad. Roy. Sci. Outre-Mer 18 (1) : 38, sub Remireo maritimae-Canavalietum roseae).
Signalons pour en finir avec cette végétation psammophile que Cyperus pedunculatus existe aussi sur les plages de Guyane française et du Surinam dans le Canavalio maritimae-Ipomoeetum brasiliensis Lindeman 1953, muté ici en Canavalio roseae-Ipomoeetum pedis-caprae nom. mut. hoc loco (Lindeman, 1953 ; de Foucault, 2014). Sa large répartition dans le monde tropical (carte in https://powo.science.kew.org/taxon/um.lsid:ipni.ord:names:305500-1) pourrait même faire de C. pedunculatus un taxon caractéristique de classe.

4. Les mégaphorbiaies
Parmi les mégaphorbiaies tropicales, d’ailleurs fort peu connues, a été décrit l’Aframomo alboviolacei-Palisotetum hirsutae des monts Nimba (de Foucault, 2022, d’après Schnell, 1952a, b). Cette association se reconnaît encore dans d’autres tableaux de Schnell (1952b) que ceux retenus en 2022, par exemple tableaux II, VI (relevé 5), VII (relevé 5), XI, XXIV (relevé 1). Peu d’informations sont cependant données par l’auteur sur cette composante des forêts marécageuses qu’il a étudiées ; en p. 249, il indique une hauteur de deux à trois mètres, ce qui confirme bien sa valeur de mégaphorbiaie. Schnell (1976 : 154, 162) parle de fourrés secondaires à grandes Marantacées ou de brousses denses à Marantacées.
Dans les données de Guillaumet (1967), notamment celles de l’annexe 14 (p. 239-241), on peut aussi reconnaître une végétation à Halopegia azurea, Marantochloa purpurea (deux Marantaceae), Maschalocephalus dinklagei (seul représentant non américain de la famille des Rapateaceae), Staurogyne paludosa (Acanthaceae), Mapania macrantha (Cyperaceae), dont on retrouve quelques éléments dans l’annexe 3 (p. 215-217, « bas-fond ») et dans l’annexe 4 (p. 217-219, « bas de pente »), avec en plus Sarcophrynium brachystachyum, Marantochloa conferta (deux autres Marantaceae) dans celle-ci. Dans un paragraphe plus général intitulé Les « brousses à Marantacées » sur sol beige à hydromorphie de profondeur (p. 103), l’auteur ajoute encore Costus afer (Costaceae), Hypselodelphys violacea, Marantochloa cuspidata, Sarcophrynium prionogonium (trois Marantaceae), Setaria megaphylla, Smilax anceps, l’ensemble de cette flore permettant l’intégration de cette végétation au moins aux Marantochloo purpureae-Costetalia aferis B. Foucault 2022 (de Foucault, 2022), végétation pouvant être décrite sous le nom de Maschalocephalo dinklagei-Marantochloetum purpureae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace (types biologiques G, hc et Ch) du relevé de l’annexe 14 in Guillaumet (1965, Recherches sur la végétation et la flore de la région du bas Cavally… : 239-241).
Affine de telles végétations, mais de transition vers les roselières, Schnell (1952b, tableau III) livre encore une végétation ainsi caractérisée (sur huit relevés) : Impatiens irvingii (photo 2) V, Limnophyton obtusifolium (Alismataceae) V, Rhynchospora corymbosa V, Thelypteris striata V, Ludwigia hyssopifolia IV, Crinum ornatum (Amaryllidaceae) IV, Hibiscus rostellatus II, Aspilia africana (Asteraceae) II, Commelina cf. macrospatha II, Cyperus distans II, Lasimorpha senegalensis (Araceae) II, Echinochloa pyramidalis II, Costus lucanusianus (Costaceae) II, Aneilema beniniense (Commelinaceae) II, Fuirena umbellata (Cyperaceae) I, Thalia geniculata (Marantaceae ; photo 3) I et autres taxons de présence I : Thelypterido striatae-Impatientetum irvingii ass. nov. hoc loco, typus nominis hoc loco : composante ‘Herbacées et suffrutescents’ du relevé 4 du tableau III in Schnell (1952b, Mém. Inst. Franç. Afrique Noire 22 : 147-148). Sillans (1958 : 159) évoque aussi un tel groupement, mais il est difficile de préciser s’il doit y être rattaché.


5. La végétation des mares temporaires de Côte-d’Ivoire
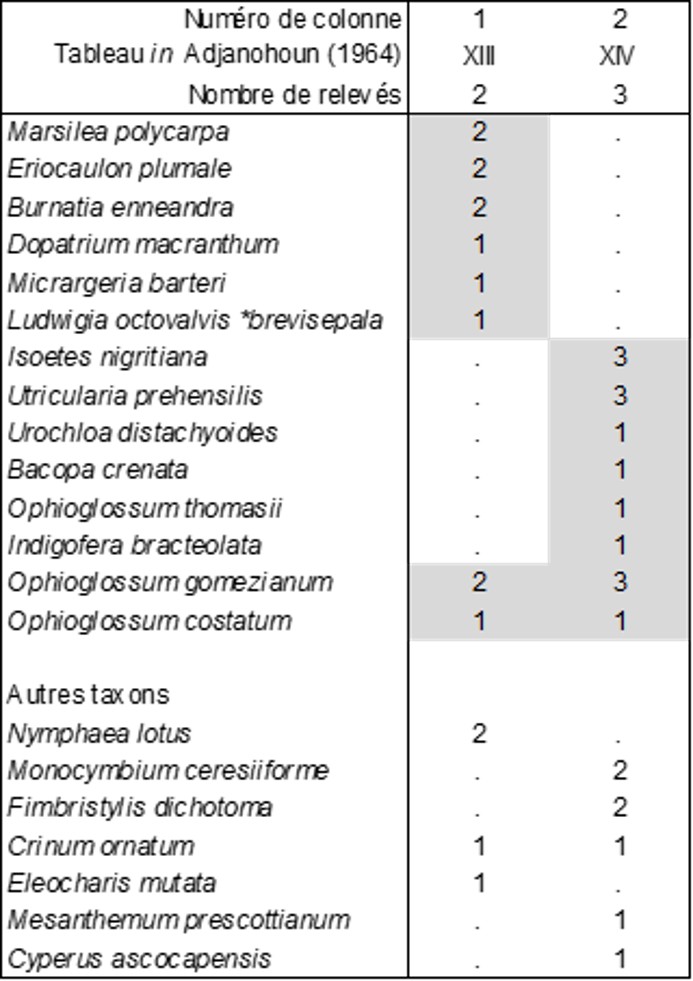
Dans son mémoire devenu un classique de la phytosociologie africaine, Adjanohoun (1964) décrit notamment la végétation de mares temporaires ivoiriennes, reprise ici dans les tableaux 3 et 4, qu’on cherchera à rapprocher des structures fonctionnelles de ce type de végétation (de Foucault, 1988). Le premier de ces deux tableaux analyse la végétation vivace de ces mares et sépare deux syntaxons :
- l’un, biologiquement hétérogène (« Dopatrio senegalensis-Marsileetum polycarpae Adjanohoun 1964 »), dont on peut extraire la composante vivace (colonne 1) à Marsilea polycarpa, Eriocaulon plumale (Eriocaulaceae), Burnatia enneandra (Alismataceae)…, Eriocaulo plumalis-Marsileetum polycarpae Adjanohoun & B. Foucault nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 2 du tableau XIII in Adjanohoun (1964, Mém. ORSTOM 7 hors texte), à rapprocher des végétations amphibies de bas niveau topographique, telles que l’Isoetetum setaceae Braun-Blanq. 1931 ex 1936 et l’Antinorio agrostideae-Isoetion velatae B. Foucault 1988 muté en Antinorio agrostideae-Isoetion longissimae nom. mut. hoc loco ;
- l’autre (colonne 2) plus homogène, riche de trois Ophioglossum, décrit sous le nom de Ophioglosso gomeziani-Isoetetum nigritianae Adjanohoun 1964, lectotypus nominis hoc loco: composante vivace du relevé 2 du tableau XIV in Adjanohoun (1964, Mém. ORSTOM 7 hors texte), à rapprocher des végétations amphibies de niveau topographique moyen, accueillant d’autres espèces du genre Ophioglossum, telles que l’Ophioglosso lusitanici-Isoetion histricis Foucault 1988.
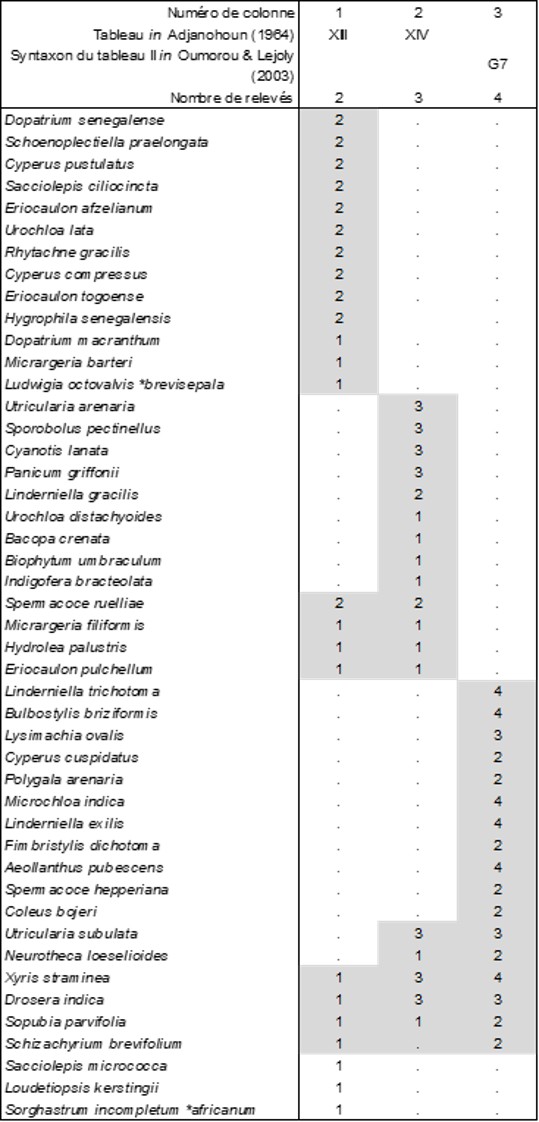
Le tableau 4 (le syntaxon G7 est abordé en partie 6) fait le même exercice pour la végétation thérophytique associée :
- l’un, biologiquement hétérogène (« Dopatrio senegalensis-Marsileetum polycarpae Adjanohoun 1964 »), dont on peut extraire la composante thérophytique (colonne 1) à Dopatrium senegalense (Plantaginaceae), Hygrophila senegalensis (Acanthaceae), riche de trois espèces d’Eriocaulon…, Eriocaulo afzeliani-Dopatrietum senegalensis Adjanohoun & B. Foucault nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 2 du tableau XIII in Adjanohoun (1964, Mém. ORSTOM 7 hors texte), à rapprocher des végétations amphibies de bas niveau topographique, telles que celles des Lythretalia borysthenici B. Foucault 2022 ;
- l’autre dont on peut extraire la composante thérophytique (colonne 2) sous le nom de Utriculario subulatae-Xyridetum stramineae Adjanohoun & B. Foucault nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 3 du tableau XIV in Adjanohoun (1964, Mém. ORSTOM 7 hors texte), à rapprocher des végétations amphibies de niveau topographique moyen telles que celles des Nanocyperetalia flavescentis Klika 1935.
De République centrafricaine, Sillans (1958) évoque aussi des végétations thérophytiques de mare temporaire (nomenclature actualisée) :
- 168, végétation à Sacciolepis indica (Poaceae), Neurotheca loeselioides, Canscora alata, C. diffusa (trois Gentianaceae), Cyperus submicrolepis;
- 169, végétation à Rotala pterocalyx (Lythraceae), Bulbostylis angolensis, Cyperus macrostachyos, C. capillifolius, Schoenoplectiella erecta ou à Scleria globonux (Cyperaceae), Dopatrium macranthum (Plantaginaceae), Rotala mexicana, Ammannia involucrata (deux Lythraceae).
Sous ces listes, on reconnaît de nouvelles réalisations de la structure systématique S(Lythraceae-Gentianaceae) (de Foucault, 1988).
6. La végétation des inselbergs
Les inselbergs sont des éléments géomorphologiques assez fréquents en région tropicale. Leur colonisation par la végétation offre de nombreux thèmes de recherches notamment abordés par Parmentier 2001, 2003a…).
6.1. Au Bénin
Oumorou et Lejoly (2003) ont étudié la végétation d’un inselberg béninois selon une méthodologie quelque peu obsolète ; on va reprendre ici leurs tableaux en les actualisant sur le plan nomenclatural et en les réorganisant selon des principes plus modernes.
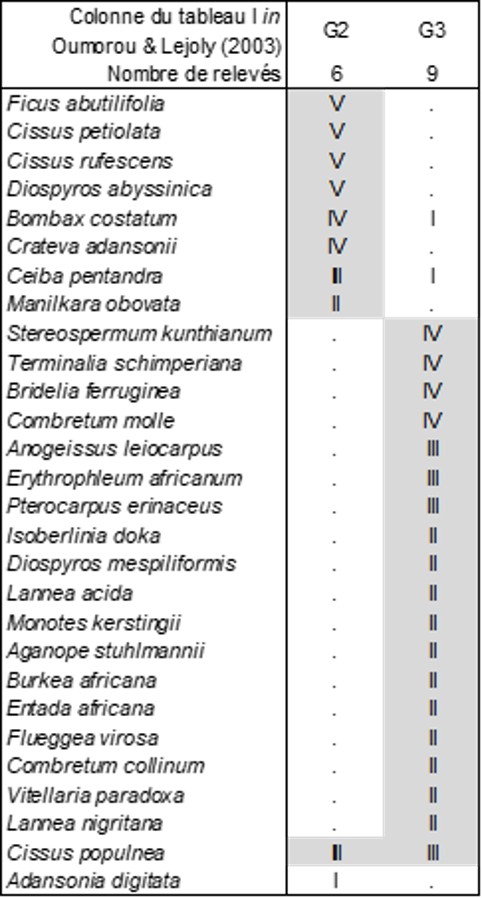
Le tableau 5 décrit ainsi la végétation arborescente que ces auteurs ont reconnue dans leur tableau I (quasi absente de leur tableau II). La colonne G2 est une forêt à Ficus abutilifolia que l’on peut décrire sous le nom de Diospyro abyssinicae-Ficetum abutilifoliae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 31 du tableau I in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 79-82). La colonne G3 en est fort différente et en outre bien plus riche en essences arborescentes : Terminalio schimperianae-Stereospermetum kunthiani ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 31 du tableau I in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 79-82, en changeant Terminalia glaucescens en T. schimperiana) ; dans cette association, on peut notamment relever la présence de Monotes kerstingii, représentant ici la famille des Dipterocarpaceae d’optimum plutôt sud-est-asiatique.
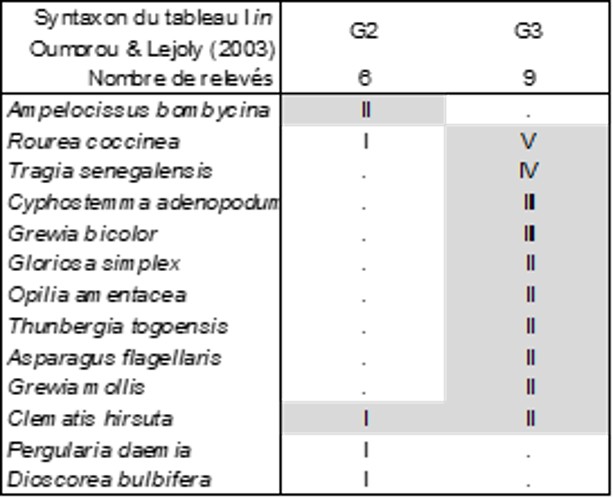
Le tableau 6 décrit la végétation arbustive, où seule la colonne G3 est floristiquement significative : Tragio senegalensis-Roureetum coccineae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 22 du tableau I in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 79-82).
Le tableau 7 est consacré à la végétation herbacée vivace de cette formation rocheuse. Les colonnes G2 et G3 partagent Commelina erecta, Adiantum philippense et Chlorophytum pusillum (Asparagaceae) ; la colonne G2 peut être reconnue comme Oplismeno hirtelli-Amorphophalletum dracontioidis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 18 du tableau I in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 79-82) ; la colonne G3 est originale avec la Poacée Andropogonée Andropogon tectorum : Siphonochilo aethiopici-Andropogonetum tectorum ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 13 du tableau I in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 79-82). Les colonnes G5 à G7 partagent surtout Afrotrilepis pilosa (Cyperaceae), taxon caractéristique des inselbergs afrotropicaux (Schnell, 1976 : 306, 309 ; Jaeger & Adam, 1981 ; Parmentier, 2001, 2003a, b). De ces trois colonnes, seul le syntaxon G7 paraît significatif, notamment avec Ophioglossum costatum qui traduit une humidification temporaire de la pelouse d’inselberg : Ophioglosso costati-Sporoboletum festivi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 7 du tableau II in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 83-84).

Le tableau 8, enfin, synthétise la végétation thérophytique étudiée sur cet inselberg. On s’intéressera particulièrement aux colonnes G5 à G7 qui partagent Spermacoce hepperiana (Rubiaceae), Aeollanthus pubescens et Coleus bojeri (deux Lamiaceae) ; la colonne G5 en est la forme la plus xérophile, Spermacoco hepperianae-Cyanotidetum lanatae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 25 du tableau II in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 83-84) ; avec la colonne G6, s’amorce un début d’humidification de cette végétation avec l’apparition de Linderniella exilis (Linderniaceae), Drosera indica, Canscora alata (Gentianaceae) ; en G7, cette dérive édaphique se précise plus fortement avec un lot conséquent de thérophytes mésohygrophiles oligotrophiles, pelouse associée à l’Ophioglosso costati-Sporoboletum festivi déjà cité : Xyrido stramineae-Lindernielletum trichotomae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 19 du tableau II in Oumorou & Lejoly (2003, Acta Bot. Gallica 150 (1) : 83-84).
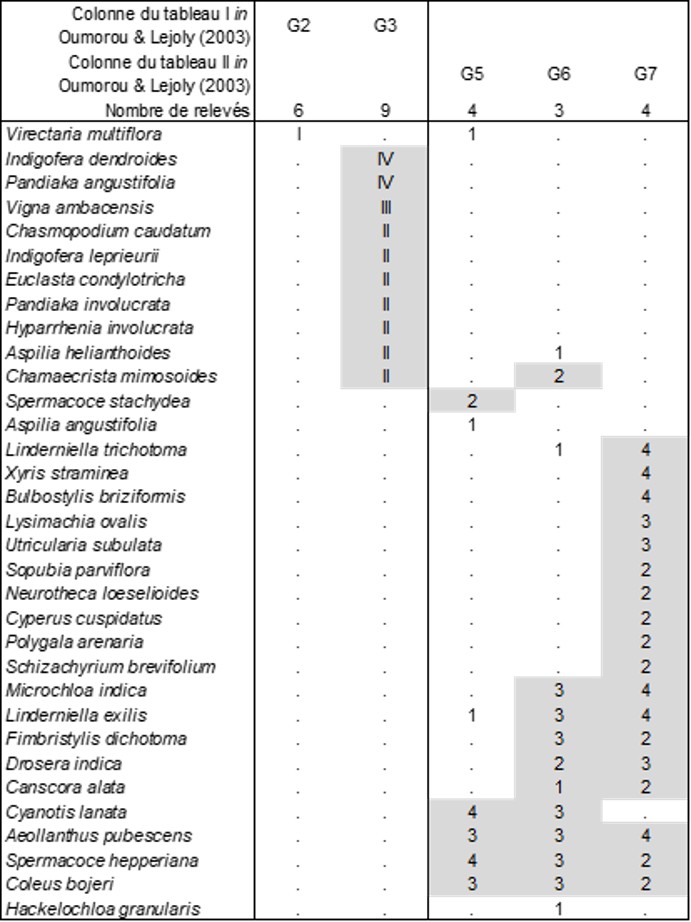
Dans la partie 5, on a reconnu les Eriocaulo afzeliani-Dopatrietum senegalensis et Utriculario subulatae-Xyridetum stramineae pour la Côte-d’Ivoire, lesquels partagent un certain nombre de taxons avec le Xyrido stramineae-Lindernielletum trichotomae béninois. Il est alors intéressant de rapprocher ces trois associations pour en préciser les affinités. Les deux premiers sont placés dans le tableau 4, on peut alors ajouter dans ce dernier la colonne G7 du tableau 8. On y voit alors se dégager une unité syntaxonomique de rang supérieur, à répartition au moins ouest-afrotropicale, de pelouses thérophytiques plus ou moins hygrophiles oligotrophies vicariantes d’une partie des Juncetea bufonii de l’hémisphère nord, caractérisée par Xyris straminea (Xyridaceae), Drosera indica, Sopubia parvifolia (Orobanchaceae), Utricularia subulata, Neurotheca loeselioides, Eriocaulon pumilum, Linderniella gracilis, L. trichotoma, Schizachyrium brevifolium (Poaceae) : Drosero indicae-Xyridion stramineae all. nov. hoc loco, typus nominis hoc loco : l’Utriculario subulatae-Xyridetum stramineae nov. Cette alliance est présente aussi au Nigeria comme le montrent les informations floristiques de Hambler (1964), ainsi qu’au Mali (Raynal & Raynal, 1961) et en Guinée (monts Nimba ; Schnell, 1952, tableau XXIX : Utriculario-Eriocauletum pulchelli nom. mut. hoc loco), et est remplacée, vers l’Afrique centrale, par l’Utricularion scandentis Mullend. 1954 (Public. INEAC 61 : 218, sub Utricularion schweinfurthii) nom. mut. hoc loco, lectotypifié par le Xyrido stramineae-Crepidorhopaletum latibracteati Mullend. 1954 (Public. INEAC 61 : 218, sub Xyrido stramineae-Craterostigmetum latibracteati) nom. mut. hoc loco, reconnaissable aussi chez Lebrun (1947 : bas de p. 526).
6.2. En Guinée-Équatoriale
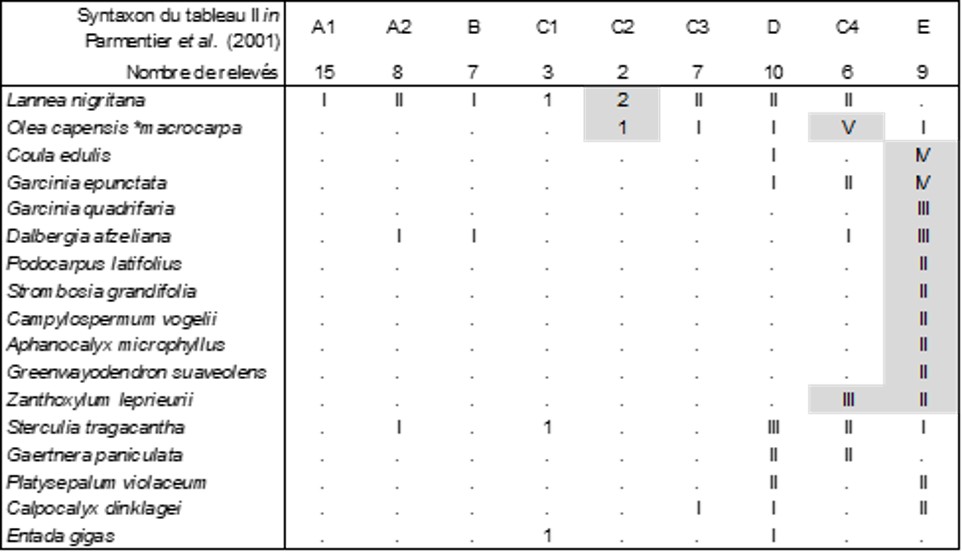
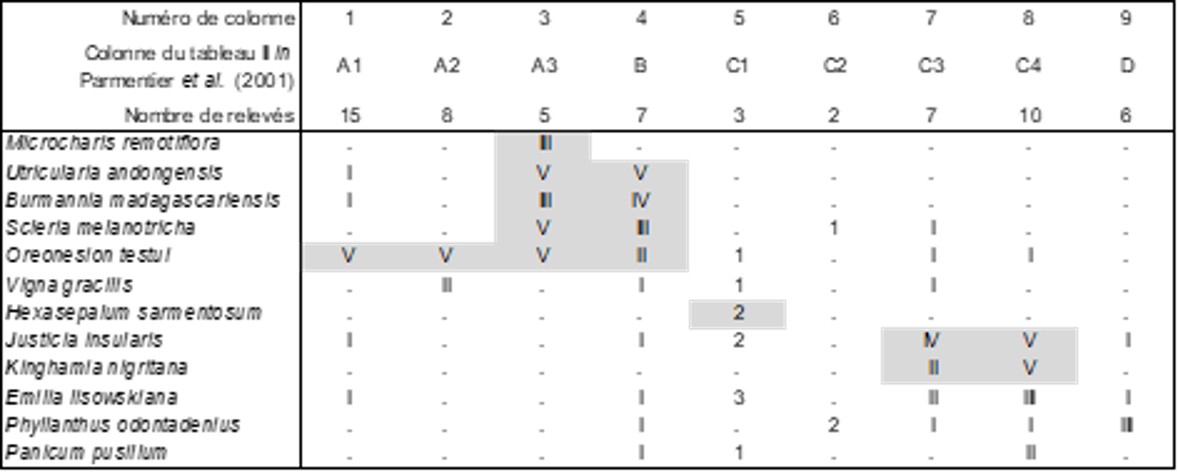
Dans une belle publication, Parmentier et al. (2001) ont décrit la diversité de la végétation des inselbergs de Piedra Nzas, en Guinée-Équatoriale. Leur approche méthodologique reste cependant très traditionnelle, mélangeant les types biologiques, y compris les épiphytes, d’où des relevés plutôt hétérogènes. On va préciser dans la suite certaines associations.
Des neuf colonnes de la végétation arborescente observée (tableau 9), seule la colonne E est véritablement significative ; elle décrit le Coulo edulis-Garcinietum epunctatae Parmentier, Lejoly & Nguema ex ass. nov. hoc loco [syn. : forêt saxicole à Garcinia epunctata et Coula edulis Parmentier et al. 2001 (Acta Bot. Gallica 148 (4) : 353) nom. inval.], une forêt saxicole de 10 à 16 m de hauteur, typus nominis hoc loco : composante arborescente du relevé r de la colonne E du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364).
Des neuf colonnes de la végétation arbustive observée (tableau 10), seules les colonnes A2 à E sont véritablement significatives ; les colonnes C3 à E, voire C1 et C2 partagent un nombre assez conséquent de taxons, Memecylon collinum (Melastomataceae), Asparagus drepanophyllus (Asparagaceae), Clappertonia polyandra (Malvaceae), Psychotria peduncularis (Rubiaceae), Scleria boivinii (Cyperaceae)… Au sein de ces six colonnes, seules les D et C4 autorisent la définition d’une association, le Memecylo collini-Asparagetum drepanophylli ass. nov. hoc loco, une lisière arbustive d’inselberg de 3 à 6 m de hauteur, typus nominis hoc loco : composante arbustive du relevé r de la colonne D du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364). L’ensemble des colonnes A2 à E possède vraisemblablement la valeur d’une alliance caractérisée par Memecylon collinum, Asparagus drepanophyllus, Clappertonia polyandra, Psychotria peduncularis, Scleria boivinii, Strychnos campicola (Loganiaceae), Adenia cissampeloides (Passifloraceae), Loeseneriella iotricha (Celastraceae), Secamone brevipes (Apocynaceae), Laccosperma secundiflorum (Arecaceae), Asparago drepanophylli-Memecylion collini all. nov. hoc loco, typus nominis hoc loco : Memecylo collini-Asparagetum drepanophylli nov. ; des espèces plus largement répandues telles que Jasminum dichotomum, Hymenodictyon floribundum (Rubiaceae), Smilax anceps permettraient un lien avec des alliances encore méconnues pour définir des unités de rang plus élevé.
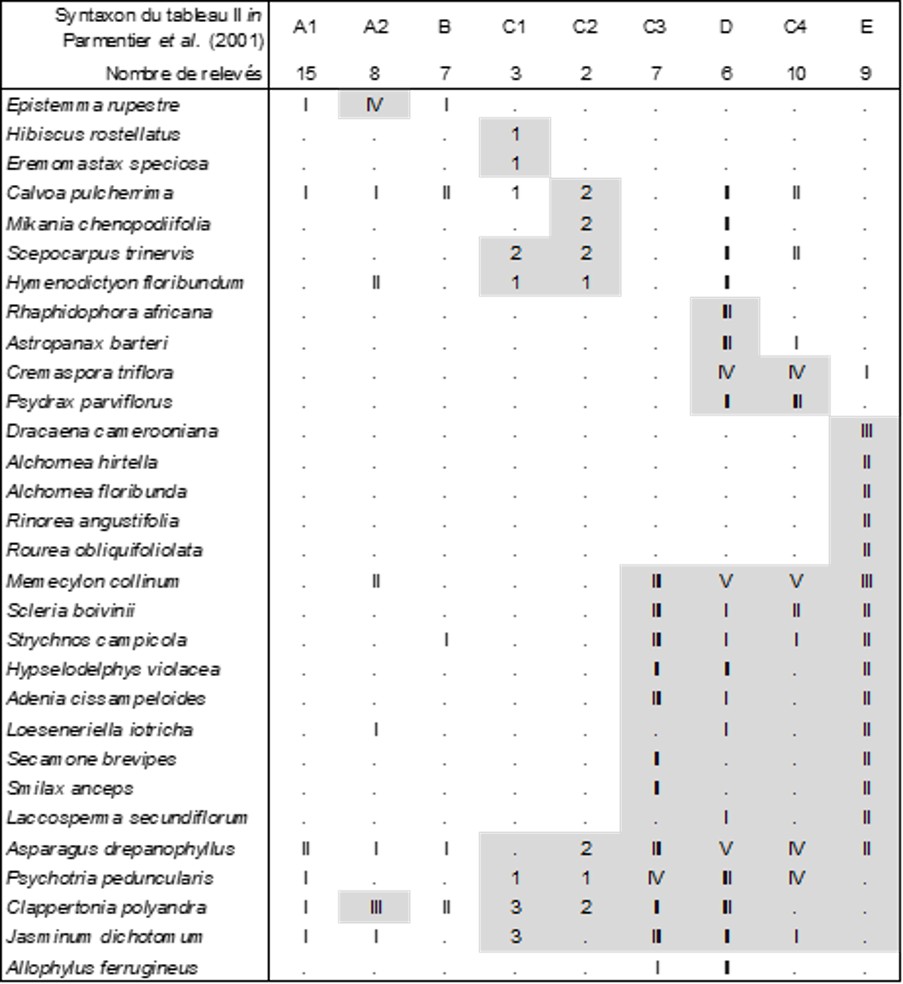
Il est regrettable que des épiphytes aient été relevés avec le reste de la végétation phorophytique (un « phorophyte » est un hôte, arbre ou arbuste, pour la végétation épiphytique), des Orchidaceae, des ptéridophytes ; c’est surtout le cas de la colonne D des auteurs (sur six relevés) : Microgramma lycopodioides (Polypodiaceae) V, Begonia eminii IV, Microsorum punctatum (Polypodiaceae) IV, Calyptrochilum emarginatum (Orchidaceae ; voir photo in Schnell, 1976 : 158) III, Rhaphidophora africana (Arecaceae) III, Solenangis scandens (Orchidaceae) II, Peperomia molleri I, Bulbophyllum cf. comatum (Orchidaceae), soit le Calyptrochilo emarginati-Begonietum eminii ass. nov. hoc loco, typus nominis hoc loco : composante épiphytique (‘e’) du relevé r de la colonne D du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364).
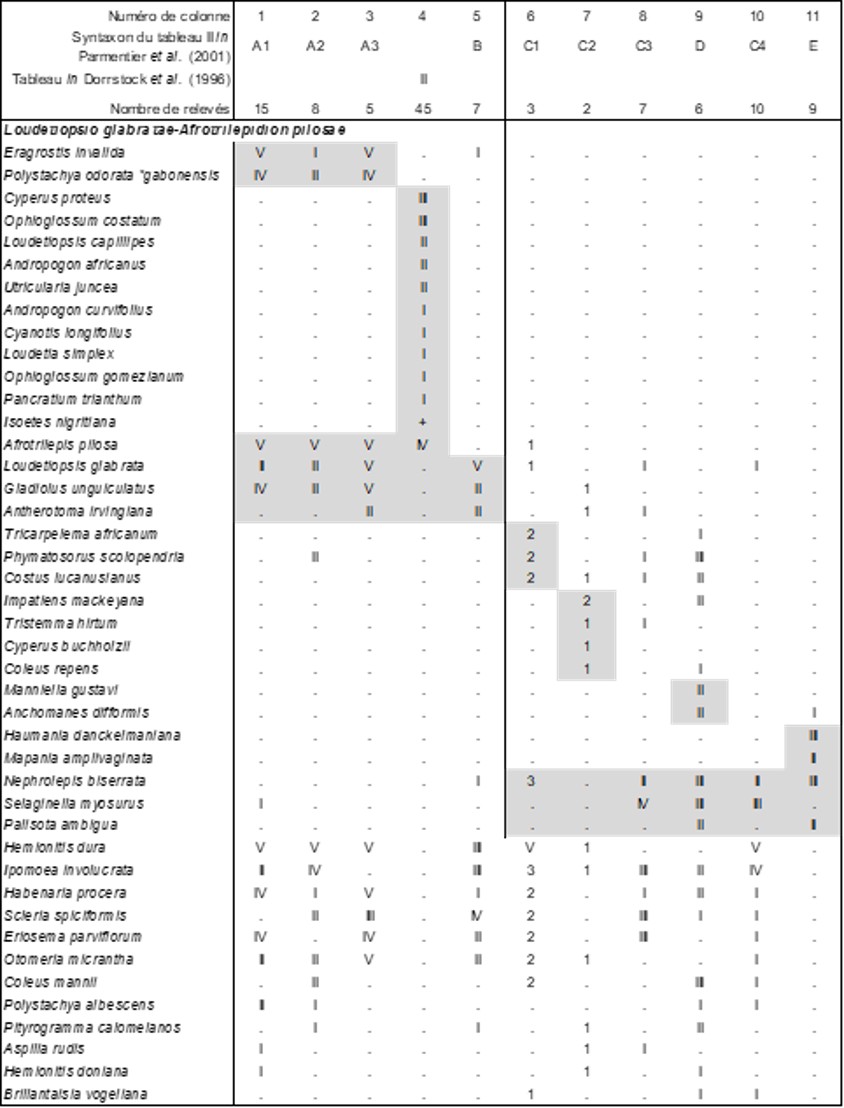
La végétation herbacée vivace est synthétisée dans le tableau 11, au niveau duquel se séparent distinctement deux ensembles : colonnes 1 à 5 d’un côté, 6 à 11 d’un autre. Dans le premier, différencié essentiellement par Loudetiopsis glabrata (Poaceae), Gladiolus unguiculatus, Antherotoma irvingiana (Melastomataceae), on peut mettre au moins en exergue l’ensemble 1 + 3 comme Habenario procerae-Afrotrilepidetum pilosae Parmentier, Lejoly & Nguema ex ass. nov. hoc loco [syn. : prairies sèches et mésoxériques à Oreonesion testui et Afrotrilepis pilosa Parmentier et al. 2001 (Acta Bot. Gallica 148 (4) : 347), prairies mésoxériques à Habenaria procera var. gabonensis et Afrotrilepis pilosa Parmentier et al. 2001 (Acta Bot. Gallica 148 (4) : 350) nom. inval.], pelouse vivace d’inselberg africain à Afrotrilepis pilosa, cypéracée xérophile typique de cet habitat, typus nominis hoc loco : composante herbacée vivace du relevé r de la colonne A3 du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364). Cet ensemble formé par les colonnes 1 à 5 a la valeur d’une alliance caractérisée par Afrotrilepis pilosa, Loudetiopsis glabrata (Poaceae), Gladiolus unguiculatus, Antherotoma irvingiana (Melastomataceae), Eragrostis invalida, Polystachya odorata subsp. gabonensis (Orchidaceae), le Loudetiopsio glabratae-Afrotrilepidion pilosae all. nov. hoc loco, typus nominis hoc loco : Habenario procerae-Afrotrilepidetum pilosae Parmentier, Lejoly & Nguema ex B. Foucault. Des monts Nimba (Guinée), Schnell (1952, tableau XXXI) définit un Melastomastro porteresi-Afrotrilepidetum pilosae nom. mut. hoc loco qui ne relève peut-être pas de cette dernière alliance.
Dans le second ensemble du tableau 11, différencié plutôt par Selaginella myosurus, Nephrolepis biserrata (Polypodiaceae), Palisota ambigua (Commelinaceae), se dégage surtout D, mais la présence la plus élevée étant III, il est difficile d’y définir une association.
Enfin dans la végétation thérophytique de ces inselbergs (tableau 12), paucispécifique, se dégage assez bien
- l’ensemble des colonnes 3 et 4 comme Burmannio madagascariensis-Utricularietum andongensis nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé r de la colonne A3 du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364) ; cette association est proche de la végétation thérophytique des mares temporaires citées précédemment, bien que manquent ici des Lythraceae, des Xyridaceae, des Drosera ;
- l’ensemble des colonnes 7 et 8 comme Kinghamio nigritanae-Justicietum insularis nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé r de la colonne C3 du tableau II in Parmentier et al. (2001, Acta Bot. Gallica 148 (4) : 358-364).
6.3. En Côte-d’Ivoire
De leur côté, Dorrstock et al. (1996) ont décrit une végétation comparable de Côte-d’Ivoire avec un tableau de 45 relevés. Là encore, celui-ci est biologiquement hétérogène.
Ainsi, la composante vivace est à nouveau caractérisée par Afrotrilepis pilosa, mais le reste de la flore est très différent de ce qui a été observé en Guinée-Équatoriale, elle est reprise dans la colonne 4 du tableau 11 et se rattache à la même alliance ; un tri plus fin des relevés permettrait d’ailleurs d’augmenter la présence des taxons. Quoi qu’il en soit, on peut définir ici le Cypero protei-Afrotrilepidetum pilosae ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 4 du tableau II in Dorrstock et al. (1996, Candollea 51 (2) hors texte, en changeant Ascolepis protea en Cyperus proteus). La présence de deux Ophioglossum et d’Isoetes nigritana montre un début d’humidification du substrat ; ces taxons rappellent l’Ophioglosso gomeziani-Isoetetum nigritanae Adjanohoun 1964 des mares temporaires du même pays (cf. supra).
La composante thérophytique est ici particulièrement diversifiée, reprise dans la colonne 6 du tableau 8. Elle se rattache clairement au Drosero indicae-Xyridion stramineae défini aussi en Côte-d’Ivoire, dans une association originale, le Bulbostylido congolensis-Utricularietum pubescentis ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 1 du tableau II in Dorrstock et al. (1996, Candollea 51 (2) hors texte). On peut émettre l’hypothèse que cette pelouse oligotrophile hygrophile dérive d’une pelouse initiale plus xérophile à Trichanthecium tenellum (Poaceae), Cyanotis lanata (Commelinaceae), Sporobolus pectinellus, Panicum griffonii… sous l’effet d’une humidification édaphique.
L’humidification des pelouses à Afrotrilepis pilosa a aussi été mise en évidence en Sierra Leone par Jaeger & Adam (1981) qui évoquent des sols tourbeux avec
- des taxons vivaces comme Mesanthemum prescottianum (Eriocaulaceae), Genlisea africana (Lentibulariaceae), Xyris festucifolia (Xyridaceae), Exochaenium gracile (Gentianaceae), Bulbostylis bulbostyloides (Cyperaceae), Anadelphia leptocoma (Poaceae), Lycopodiella cernua, Osmunda regalis, Drosera pilosa;
- des taxons annuels comme Mesanthemum auratum, Utricularia subulata, pubescens, Eriocaulon pulchellum, Bulbostylis briziformis, Panicum pusillum, Swertia mannii, Schizachyrium brevifolium, Lobelia rubescens, Cyperus atrorubidus, Nerophila gentianoides (Melastomataceae) ;
on reconnaît dans les deux listes la structure systématique S(Xyridaceae-Eriocaulaceae) (de Foucault, 1988), qui peut donc se réaliser concrètement tant en végétation vivace que thérophytique, et dans la seconde un cortège rappelant le Drosero indicae-Xyridion stramineae.
Des végétations comparables sont aussi évoquées du mont Niangbo par Porembski et al. (1996 : 257).
7. Les forêts du mont Kenya
Le mont Kenya, dont le nom signifie « montagne de l’Autruche » chez les Wakamba, l’une des populations vivant à son pied, est le point culminant du Kenya et le deuxième plus haut sommet d’Afrique, après le Kilimandjaro (sur le territoire de la Tanzanie), culminant à 5 199 m à la pointe Batian. Il se situe au centre du pays, juste au sud de l’équateur, à approximativement cent cinquante kilomètres au nord-nord-est de la capitale Nairobi. Ce mont est un volcan rouge (volcan effusif émettant des laves généralement basaltiques, suffisamment fluides pour ne pas s’accumuler au point de sortie, et formant des fontaines et des coulées de lave plus ou moins longues progressant parfois à plusieurs kilomètres par heure) né il y a environ trois millions d’années de l’ouverture du rift est-africain. Il a été recouvert pendant des millénaires par une importante calotte glaciaire qui a fortement érodé ses pentes et lui a donné son relief particulier, avec de nombreuses vallées qui descendent du sommet. Il reste aujourd’hui une douzaine de petits glaciers en phase de retrait rapide, malgré des températures souvent négatives, avec un climat très variable au cours des millénaires et des siècles, des saisons et des jours. La montagne demeure une source d’eau essentielle pour une grande partie du pays. Elle possède huit étages de végétation entre le bas et le sommet, avec notamment une vaste couronne de forêts. De nombreuses espèces sont endémiques ou très caractéristiques du mont Kenya, comme des Lobelia (L. gregoriana, L. telekii), des Dendrosenecio (D. keniensis, D. battiscombei ; Asteraceae) ou, pour la faune, le Daman du Cap (Procavia capensis). C’est pourquoi une zone de 715 km2 autour du sommet est protégée par le parc national du mont Kenya, classé par l’Unesco au patrimoine mondial de l’humanité (toute cette partie est reprise et légèrement modifiée de Wikipédia [25.12.2024, 28.12.2024]).
Bussmann et Beck (1995) ont publié une importante monographie sur la végétation forestière de ce mont (plus de six cents relevés mis en tableaux, en dessous de l’étage des Lobelia et Dendrosenecio), dans laquelle malheureusement tous les types de formation sont mélangés (végétations arborescente, arbustive, herbacée vivace, épiphytique…). En outre, les syntaxons de rang supérieur à l’association (alliances…) sont typifiés par des relevés au lieu de syntaxons de rang juste inférieur (associations pour les alliances…). Suite à ce constat, on va tenter de reprendre ici sur une base plus actuelle et plus rigoureuse une partie de cette végétation forestière.
7.1. La végétation arborescente
Le tableau 13 synthétise une partie des nombreux syntaxons qui se dégagent de l’analyse. On y distingue trois groupes de colonnes.
Les colonnes 1 à 9 tout d’abord sont caractérisées par le groupe sociologique à Astropanax volkensii (Araliaceae). On peut notamment mettre en exergue
- les colonnes 1 et 2 comme Apodyto dimidiatae-Kuloetum usambarensis nov. hoc loco, typus nominis hoc loco: composante arborescente (taxons notés T dans les tableaux de 1995) du relevé 10 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538, en changeant Ocotea usambarensis en Kuloa u.) ;
- la colonne 7 comme Cussonio holstii-Kuloetum usambarensis nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 246 du tableau 5 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 539-543, en changeant Ocotea usambarensis en Kuloa u.) ;
- la colonne 8 comme Faureo salignae–Kuloetum usambarensis nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 220 du tableau 5 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 539-543, en changeant Ocotea usambarensis en Kuloa u.).
L’ensemble de ces neuf colonnes, caractérisé par Astropanax volkensii, Cornus volkensii, Lepidotrichilia volkensii (Meliaceae), Psychotria orophila (Rubiaceae), Cussonia holstii (Araliaceae), Euphorbia abyssinica, Polyscias fulva (Araliaceae), Faurea saligna (Proteaceae), Ocotea kenyensis (Lauraceae), Dracaena afromontana (Asparagaceae), possède la valeur d’une alliance originale, le Corno volkensii-Astropanacion volkensii all. nov. hoc loco, typus nominis hoc loco : le Cussonio holstii-Kuloetum usambarensis nov.
Dans l’ensemble formé par les colonnes 10 à 22, bien caractérisé par le groupe sociologique à Anthocleista grandiflora, on peut au moins distinguer
- la colonne 10 comme Galiniero saxifragae-Mussaendetum odoratae ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 15 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538, en remplaçant Galiniera coffeoides par saxifraga et Moussaenda odorata par Mussaenda microdonta subsp. o.) ;
- les colonnes 12 et 13 comme Myriantho holstii-Tabernaemontanetum stapfianae nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 51 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538) ;
- la colonne 15 comme Cordio africanae-Harunganetum madagascariensis nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 68 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538).
Le Myriantho holstii-Tabernaemontanetum stapfianae nov. peut être retenu comme typus nominis hoc loco de cet ensemble de treize colonnes à valeur d’alliance, le Zanthoxylo gilletii-Anthocleistion grandiflorae all. nov. hoc loco, caractérisé par Anthocleista grandiflora (Gentianaceae), Zanthoxylum gilletii (Rutaceae), Trichilia dregeana (Meliaceae), Garcinia volkensii (Clusiaceae), Casearia battiscombei (Salicaceae), Croton macrostachyus (Euphorbiaceae), Albizia gummifera (Fabaceae), Myrianthus holstii (Urticaceae), Xymalos monospora (Monimiaceae), Ochna insculpta (Ochnaceae), Vitex fischeri var. keniensis (Lamiaceae), Peddiea fischeri (Thymelaeaceae), Psychotria fractinervata (Rubiaceae).
L’ensemble formé par les colonnes 23 à 28 est globalement peu caractérisé ; on peut cependant y définir
- la colonne 25 comme Ekebergio capensis-Syzygietum capensis ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 25 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538) ;
- la colonne 27 comme Oleo macrocarpae-Macarangetum kilimandscharicae ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 167 du tableau 5 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 539-543, en changeant Olea capensis hochstetteri en O. c. subsp. macrocarpa).
Relativement aux deux premières alliances, il convient de définir celle-ci comme Oleo macrocarpae-Kuloion usambarensis all. nov. hoc loco, surtout caractérisé par Olea capensis subsp. macrocarpa et Psychotria fractinervata (Rubiaceae), typus nominis hoc loco : l’Oleo macrocarpae-Macarangetum kilimandscharicae nov.
Tout l’ensemble constitué par les colonnes 1 à 28 peut alors être interprété comme un ordre caractérisé par Kuloa usambarensis (= Ocotea u., Lauraceae), Tabernaemontana stapfiana (Apocynaceae), Strombosia scheffleri (Olacaceae), Macaranga kilimandscharica (Euphorbiaceae), Lasianthus kilimandscharicus (Rubiaceae), Podocarpus latifolius (Podocarpaceae), Apodytes dimidiata (Icacinaceae), Aningeria adolfi-friederici (Sapotaceae), Cassipourea malosana (Rhizophoraceae), Neoboutonia macrocalyx (Euphorbiaceae), Ekebergia capensis (Meliaceae), Bersama abyssinica (Melianthaceae), Ilex mitis (Aquifoliaceae), Newtonia buchananii (Fabaceae), Syzygium guineense (Myrtaceae), les Strombosio scheffleri-Kuloetalia usambarensis ord. nov. hoc loco, typus nominis hoc loco : le Zanthoxylo gilletii-Anthocleistion grandiflorae nov.
Le tableau 14 synthétise l’autre partie des syntaxons arborescents du mont Kenya. Son interprétation est plus délicate que celle du tableau 13.
Les colonnes 29 à 38 paraissent correspondre à un groupe assez structuré, où l’on peut au moins distinguer
- les colonnes 32 et 33 (plus peut-être 34 en tant que variation) comme Psydraco schimperiani-Lepidotrichilietum volkensii ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 270 du tableau 7 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 545-549) ;
- les colonnes 34 à 37 (et peut-être 38) comme Ilici mitis-Podocarpetum latifolii ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 296 du tableau 7 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 545-549).
Cet ensemble caractérisé par Nuxia congesta (Stilbaceae), Vepris nobilis (Rutaceae), Juniperus procera, Olea europaea subsp. cuspidata, Psydrax schimperianus (Rubiaceae), Halleria lucida (Stilbaceae), Calodendrum capense (Rutaceae) peut être considéré comme une alliance, le Nuxio capensis-Oleion macrocarpae all. nov. hoc loco, typus nominis hoc loco : le Psydraco schimperiani-Lepidotrichilietum volkensii nov., alliance de transition vers les Strombosio-Kuloetalia usambarensis.
Les colonnes 39 à 49 forment un second ensemble :
- la colonne 39 décrit l’Agaristo salicifoliae-Podocarpetum latifolii nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 377 du tableau 8 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 550-553) ;
- les colonnes 40 à 42 peuvent se rattacher à un unique Myrsino melanophlei-Podocarpetum latifolii nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 388 du tableau 8 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 550-553, en remplaçant Rapanea melanophleos par Myrsine m.) ;
- les colonnes 43 à 45 sont faiblement caractérisées et restent en attente d’une interprétation définitive ;
- la colonne 46 est un Ekebergio capensis-Polysciadetum kikuyuensis ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (T) du relevé 471 du tableau 8 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 550-553) ;
- les colonnes 47 à 49 sont assez bien différenciées, mais le nombre de relevés de chacune d’elles n’est que de cinq.
Tout cet ensemble 39 à 49 est caractérisé ou différencié par Nuxia congesta, Polyscias kikuyuensis (Araliaceae), Dombeya torrida subsp. t. (Malvaceae), Faurea saligna (Proteaceae), Myrsine melanophleos (Primulaceae), ce qui justifie la définition du Nuxio congestae-Juniperion procerae all. nov. hoc loco, typus nominis hoc loco : le Myrsino melanophlei-Podocarpetum latifolii nov.
Le Nuxio-Oleion macrocarpae et le Nuxio-Juniperion procerae peuvent être rapprochés dans l’ordre des Nuxio congestae-Juniperetalia procerae ord. nov. hoc loco, caractérisé par Nuxia congesta, Juniperus procera, Afrocarpus falcatus (Podocarpaceae), Olea europaea subsp. cuspidata, typus nominis hoc loco : le Nuxio congestae-Juniperion procerae nov.
Dans l’ensemble 50 à 54 du tableau 14, seule la colonne 50 est statistiquement et floristiquement assez définie pour justifier un Olinio rochetianae-Hagenietum abyssinicae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T) du relevé 594 du tableau 10 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 557-558). Cet ensemble possède cependant la valeur d’une alliance, l’Oleo cuspidatae-Hagenion abyssinicae all. nov. hoc loco, de transition vers le Nuxio congestae-Juniperion procerae et caractérisée par Lasiosiphon glaucus (Thymelaeaceae), Prunus africana, Olinia rochetiana (Penaeaceae), Ekebergia capensis, Podocarpus latifolius, typus nominis hoc loco : le Myrsino melanophlei-Podocarpetum latifolii nov.
Enfin, l’ensemble des colonnes 55 et 56 décrit l’unique Hyperico revoluti-Hagenietum abyssinicae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T) du relevé 588 du tableau 10 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 557-558), par ailleurs typus nominis hoc loco de l’Hyperico revoluti-Hagenion abyssinicae all. nov. hoc loco, caractérisé par Hypericum revolutum (photo 4), Myrsine melanophleos et Hagenia abyssinica (Rosaceae), en outre unique alliance et typus nominis hoc loco des Hyperico revoluti-Hagenietalia abyssinicae ord. nov. hoc loco, dont les taxons caractéristiques coïncident avec ceux caractérisant l’alliance type.
Il reste encore à traiter les bambouseraies d’altitude à Oldeania alpina, que l’on peut rattacher aux boisements par leurs chaumes ligneux et leur taille. Sur les 52 relevés qui les définissent, 19 combinent ce taxon à Podocarpus latifolius, Nuxia congesta et parfois Olea capensis subsp. macrocarpa, les autres étant réduits au seul O. alpina. On peut alors décrire le Podocarpo latifolii-Oldeanietum alpinae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (T) du relevé 516 du tableau 9 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 554-556, en changeant Sinarundinaria alpina en Oldeania a.).

Le tableau synthétique global 15 reprend toutes les unités définies ici en condensant chaque alliance en une seule colonne, montrant que les alliances CAv, ZAg et OKu ne partagent guère que Podocarpus latifolius avec les autres ; elles peuvent donc être réunies dans un ordre et une classe uniques, les Strombosio scheffleri-Kuloetalia usambarensis définis plus haut et les Strombosio scheffleri-Kuloetea usambarensis classis nov. hoc loco, caractérisés par les mêmes taxons que l’ordre. Il n’est peut-être pas nécessaire de scinder l’ensemble des quatre autres alliances (NOm à HHa) en deux classes : on peut retenir ici les Junipero procerae-Podocarpetea latifolii classis nov. hoc loco, caractérisés par Podocarpus latifolius, Juniperus procera, Olea europaea subsp. cuspidata, Myrsine melanophleos, typus nominis hoc loco : les Nuxio congestae-Juniperetalia procerae nov., classe qui existe aussi sur les Cherangani Hills, au nord-ouest du Kenya (Mabberley, 1975). Le Podocarpo latifolii-Oldeanietum alpinae pourrait alors être inclus dans le Nuxio capensis-Oleion macrocarpae. Il sera peut-être possible d’inclure dans cette dernière classe les forêts à J. procera, Hagenia abyssinica et Afrocarpus gracilior d’Éthiopie (Guillaumet, 2009).
7.2. Quelques groupements subordonnés aux forêts kenyanes
Il serait d’un grand intérêt de poursuivre l’exercice précédent avec la végétation arbustive intraforestière, mais il reste plutôt complexe à finaliser. Contentons-nous de signaler un fourré lié aux boisements du Nuxio congestae-Juniperion procerae, décrit par les relevés 370 à 437, 441 à 445 et 491 à 495 du tableau 8 de Bussmann & Beck (1995, soit 78 relevés) : Berberis holstii V, Dovyalis abyssinica (Salicaceae) IV, Rubus volkensii III, Mimulopsis alpina (Acanthaceae) II, Podocarpus latifolius j V, Juniperus procera j IV, Myrsine melanophleos j III, Cornus volkensii j II, Cussonia spicata (Araliaceae) j II, soit le Dovyali abyssinicae-Berberidetum holstii ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 406 du tableau 8 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 550-553). M. alpina et C. spicata différencient une sous-association mimulopsietosum alpinae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 373 du tableau 8 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 550-553).
Sous diverses canopées relevant des Strombosio scheffleri-Kuloetalia usambarensis, on peut remarquer une mégaphorbiaie d’altitude à (d’après le tableau 4 de Bussmann & Beck, 1995 : relevés 1 à 29, soit 29 relevés) Thalictrum rhyncocarpum V, Dryopteris kilemensis V, Oplismenus hirtellus (Poaceae) V, Impatiens meruensis V, Panicum calvum V, Arisaema mildbraedii (Araceae) IV, Pteris pteridioides IV, P. catoptera III, Cynanchum abyssinicum III, Carex chlorosaccus III, Setaria megaphylla I, décrivant le Panico calvi-Thalictretum rhynchocarpi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 11 du tableau 4 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 534-538).
Il en existe une autre décrite par les relevés 215 à 254 (soit 40 relevés) de leur tableau 5 : Carex chlorosaccus V, Dryopteris kilemensis V, Thalictrum rhynchocarpum IV, Impatiens mildbraedii IV, Cynanchum abyssinicum IV, C. altiscandens III…, que l’on peut décrire comme Cynancho altiscandentis-Thalictretum rhynchocarpi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 220 du tableau 5 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 539-543).
Une troisième mégaphorbiaie à Th. rhynchocarpum est décrite par les relevés 531 à 534 : Th. rhynchocarpum 4, Pteris catoptera 4, Carex chlorosaccus 4, C. vallis-rosetto 4, C. johnstonii 4, Senecio synringifolius 4, Carici johnstonii-Thalictretum rhynchocarpi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 531 du tableau 9 in Bussmann & Beck (1995, Phytocoenologia 25 (4) : 554-556). Ces trois associations affines partagent donc Thalictrum rhynchocarpum, Carex chlorosaccus, Dryopteris kilemensis, Pteris catoptera, Cynanchum abyssinicum, qui caractérisent une alliance de mégaphorbiaie oro-africaine, le Carici chlorosacci-Thalictrion rhynchopetali all. nov. hoc loco, typus nominis hoc loco : le Cynancho altiscandentis-Thalictretum rhynchocarpi nov.
Au voisinage du Panico calvi-Thalictretum, on remarque aussi une microphorbiaie à Lobelia baumannii, Hydrocotyle mannii, Centella asiatica (Apiaceae), trois taxons à feuilles suborbiculaires souvent caractéristiques de ce type de formation végétale (de Foucault, 2018) ; il en est de même au voisinage du Carici johnstonii-Thalictretum avec Lobelia duriprati et peut-être Clinopodium uhligii var. obtusifolium.
8. D’autres végétations ligneuses
Dans ce paragraphe et le tableau 16, on évoque d’abord des végétations sénégalaises issues des données de Trochain (1940). On a en outre repris ceux décrits du Niger (colonnes 1 à 11 ; de Foucault, 2024) pour comparaison. La colonne 12 s’individualise en tant que Guiero senegalensis-Phyllanthetum pentandri ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 8 du tableau XXII in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 191-193) ; la comparaison avec les fourrés nigériens montre qu’il est possible de le rattacher aussi au Combreto micranthi-Guierion senegalensis B. Foucault 2024. Les colonnes 13 et 14 paraissent se rapporter à un unique Bauhinio rufescentis-Ziziphetum mauritianae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 1 du tableau XVIII in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 167-168), lequel se rattache plutôt mal à l’alliance nigérienne.
Dans ce paragraphe et le tableau 17, on évoque maintenant des végétations arborescentes sénégalaises toujours issues des données de Trochain (1940). On a en outre repris ceux décrits du Niger (colonnes 1 à 14 ; de Foucault, 2024) pour comparaison. Les colonnes 15 et 16 paraissent se rapporter à un unique Balanito aegyptiacae-Vachellietum raddianae ass. nov. hoc loco [syn. : groupement à Acacia raddiana Trochain 1940 (Mém. Inst. Franç. Afrique noire 2 : 167-169)], typus nominis hoc loco : composante arborescente du relevé 13 du tableau XVIII in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 167-169). La colonne 19 est un Vachellio seyal-Senegalietum ataxacanthae ass. nov. hoc loco [syn. : faciès à Acacia ataxacantha Trochain 1940 (Mém. Inst. Franç. Afrique noire 2 : 252)], typus nominis hoc loco : composante arborescente du relevé 4’ du tableau XXXI in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 256-257). Enfin la colonne 20 peut être interprétée comme Faidherbio albidae-Adansonietum digitatae ass. nov. hoc loco [syn. : péniclimax à Faidherbia albida Trochain 1940 (Mém. Inst. Franç. Afrique noire 2 : 257)], typus nominis hoc loco : composante arborescente du relevé 7 du tableau XXXII in Trochain (1940, Mém. Inst. Franç. Afrique noire 2 : 262-263). Dans leur ensemble, les colonnes 17 à 20 se rattachent à une unité supérieure à valeur d’alliance distincte du Bauhinio rufescentis-Vachellion raddianae défini au Niger, caractérisée par Adansonia digitata (Malvaceae), Sclerocarya birrea (Anacardiaceae), Vachellia sieberiana, Borassus aethiopum (Arecaceae), Afromorus mesozygia (Moraceae), Celtis toka (Cannabaceae), Pterocarpus erinaceus (Fabaceae), Khaya senegalensis (Meliaceae), Senegalia ataxacantha (Tamarindus indica n’est que naturalisé ici) : Sclerocaryo birreae-Adansonion digitatae all. nov. hoc loco, typus nominis hoc loco : le Faidherbio albidae-Adansonietum digitatae nov. Le Bauhinio rufescentis-Vachellion raddianae et le Sclerocaryo birreae-Adansonion digitatae peuvent cependant être rapprochés dans l’ordre des Vachellio seyal-Balanitetalia aegyptiacae ord. nov. hoc loco, caractérisé par Balanites aegyptiaca (Zygophyllaceae), Vachellia seyal, Senegalia macrostachya, S. senegal, Terminalia avicennioides et Anogeissus leiocarpus (Combretaceae), Diospyros mespiliformis (Ebenaceae), Hyphaene thebaica (Arecaceae), typus nominis hoc loco : le Sclerocaryo birreae-Adansonion digitatae nov.
De leur côté, Mandango et al. (1996) évoquent des forêts juvéniles à Anthocleista schweinfurthii (Gentianaceae), Harungana madagascariensis (Hypericaceae), Macaranga spinosa, M. monandra, Mallotus oppositifolius (deux genres d’Euphorbiaceae), Musanga cecropioides (Urticaceae), Trema orientale (Cannabaceae), vicariant du Musangetum cecropioidis Yongo & B. Foucault 2007 de République centrafricaine, mais floristiquement plus riche (Yongo & de Foucault, 2007).
9. La végétation afro-alpine
Tout comme Schnell (1976, 1977) traite de la végétation des hautes montagnes d’Afrique dans ses ouvrages sur l’Afrique tropicale, pour achever cette étude, on va quitter les étages tropicaux pour s’élever dans les étages afro-alpins, surtout est-africains, sous climat plus tempéré, célèbres notamment pour les Dendrosenecio et Lobelia scapeux (Hauman, 1934 ; Hedberg, 1951, 1964 ; Schnell, 1977 ; photo 5). À côté de ces fourrés, il existe cependant aussi des pelouses remarquables par la représentation d’espèces originales du genre Alchemilla.

Le tableau 18 synthétise quelques-unes d’entre elles que l’on peut décrire en tant qu’association :
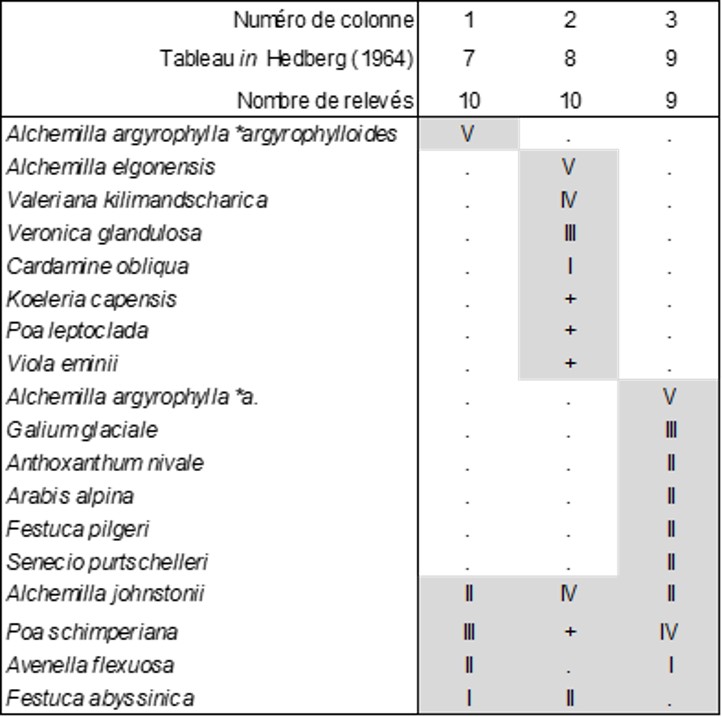
- la colonne 1 de ce tableau est le Poo schimperianae-Alchemilletum argyrophylloidis Hedberg & B. Foucault nov. hoc loco, du Rwenzori (Ouganda et République démocratique du Congo), vers 3 900 m d’altitude, typus nominis hoc loco: le relevé 9 du tableau 7 in Hedberg (1964, Acta Phytogeogr. Suec.49 : 111) ; du même massif, Hauman (1933) évoque aussi des pelouses à A. johnstonii, A. stuhlmannii, A. argyrophylla subsp. argyrophylloides, A. subnivale, Festuca abyssinica, Afrosciadium kerstenii (Apiaceae) ;
- la colonne 2 est le Valeriano kilimandscharicae-Alchemilletum elgonensis Hedberg & B. Foucault nov. hoc loco, du mont Elgon (Ouganda et Kenya), vers 3 700 m d’altitude, typus nominis hoc loco: le relevé 4 du tableau 8 in Hedberg (1964, Acta Phytogeogr. Suec.49 : 112) ; photo 6 ;
- la colonne 3 (avec Hypericum revolutum, photo 4) est le Poo schimperianae-Alchemilletum argyrophyllae Hedberg & B. Foucault nov. hoc loco, du mont Kenya (Kenya), vers 4 200 m d’altitude, typus nominis hoc loco: le relevé 8 du tableau 9 in Hedberg (1964, Acta Phytogeogr. Suec.49 : 113).

Ces trois associations partagent suffisamment de taxons, notamment Alchemilla johnstonii (photo 7), Poa schimperiana, Avenella flexuosa, Festuca abyssinica, pour que cet ensemble acquière la valeur d’une alliance de pelouses vivaces afro-alpines : Poo schimperianae-Alchemillion johnstonii Hedberg & B. Foucault all. nov. hoc loco, typus nominis hoc loco : le Poo schimperianae-Alchemilletum argyrophyllae Hedberg & B. Foucault nov.

Dans des boisements afro-alpins à Hagenia abyssinica (Rosaceae) du mont Kenya décrits plus haut, Bussmann et Beck (1995) citent aussi de telles prairies, mais avec une composition floristique assez différente : Alchemilla argyrophylla, A. johnstonii, A. fischeri, Cyperus kerstenii, Kniphofia thomsonii (Asphodelaceae), Poa schimperiana, Luzula johnstonii, Geranium kilimandscharicum, Polygonum afromontanum, Silene afromontana, Agrostis kilimandscharica…, sans doute à rattacher à une alliance différente de la précédente.
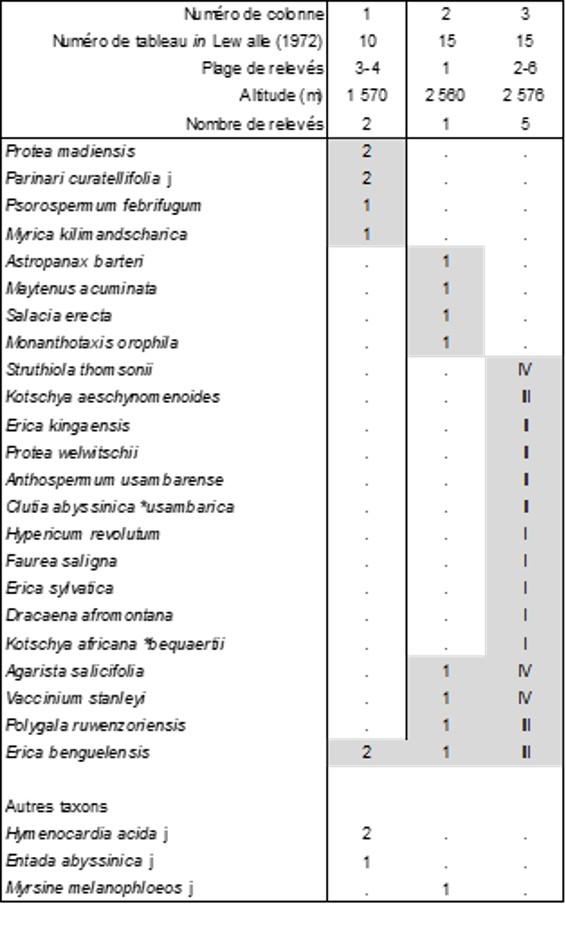
Le botaniste et phytogéographe belge José Lewalle (voir annexe) a laissé une belle étude de montagnes du Burundi occidental (1972). Parmi la végétation décrite, on a d’abord extrait une pelouse à succulentes (d’après les relevés 12 et 13 du tableau 15 de l’auteur, Ch + H, vers 2 500 m) : Aloe lateritia 2, Crassula alba 2, Helichrysum tillandsiifolium 2, Oleandra distenta (Polypodiaceae) 2, Alvesia clerodendroides (Lamiaceae) 2, Hemionitis inaequalis (Pteridaceae) 2, H. pectiniformis 2, Anemia lepigera (Schizaeaceae) 2, Otiophora lebruniana *l. (Rubiaceae) 2, O. lebruniana *devrediana 1, Lobelia holstii 1, Asplenium friesiorum 1, Microchloa kunthii (Poaceae) 1, soit le Crassulo albae-Aloetum lateritiae Lewalle & B. Foucault ass. nov. hoc loco, typus nominis hoc loco : composante hémicryptophyto-chaméphytique du relevé 13 du tableau 15 in Lewalle (1972, Bull. Jard. Bot. Natl. Belg. 42 (1-2) : 142-145). Cette combinaison d’Aloe et de Crassula montre des affinités avec la végétation homologue d’Afrique du Sud.
Le tableau 19 hoc loco reprend maintenant quelques végétations nanophanérophytiques. La colonne 1 décrit un fourré de moyenne altitude à Protea madiensis : Erico benguelensis-Proteetum madiensis Lewalle & B. Foucault ass. nov. hoc loco, typus nominis hoc loco : composante nanophanérophytique du relevé 4 du tableau 10 in Lewalle (1972, Bull. Jard. Bot. Natl. Belg. 42 (1-2) : 102-103, en changeant Philippia benguelensis en Erica b.). Les colonnes 2 et 3 décrivent plutôt des fourrés d’altitude supérieure, dont seule la 3 est significative : Vaccinio stanleyi-Agaristetum salicifoliae Lewalle & B. Foucault ass. nov. hoc loco, typus nominis hoc loco : composante nanophanérophytique du relevé 2 du tableau 15 in Lewalle (1972, Bull. Jard. Bot. Natl. Belg. 42 (1-2) : 142-145, en changeant Agauria salicifolia en Agarista s.). Partageant au moins Erica benguelensis, Agarista salicifolia, Vaccinium stanleyi, Polygala ruwenzoriensis, ces colonnes 2 et 3 peuvent être retenues pour définir une alliance originale de fourrés afro-alpins à Protea et Erica, le Vaccinio stanleyi-Ericion benguelensis Lewalle & B. Foucault all. nov. hoc loco, typus nominis hoc loco : le Vaccinio stanleyi-Agaristetum salicifoliae Lewalle & B. Foucault nov. Comme la pelouse à Aloe lateritia du paragraphe précédent, ces fourrés montrent de réelles affinités avec ceux bien plus développés à des latitudes et altitudes plus basses, dans les fynbos de la région du Cap (Afrique du Sud) où plusieurs genres de Proteaceae, de nombreux Erica, des Struthiola (Thymelaeaceae), Myrica, Anthospermum (Rubiaceae), Clutia (Peraceae) s’associent pour structurer des fourrés austraux. De Tanzanie, Kerfoot (1963-64 : 197) évoque aussi des fourrés à Protea angolensis var. divaricata, P. heckmanniana, P. welwitschii, P. rubrobracteata, Erica sylvatica, de même que Schnell (1976 : 391) pour des fourrés à Protea gaguedi et Agarista salicifolia ; le même auteur (p. 393) en cite du Zimbabwe. De même, dans la chaîne des Bambuttos (Cameroun), Portères (1946 : 355-356) cite un fourré à Erica mannii et Protea madiensis, taxons associés à Adenocarpus mannii et Pittosporum viridiflorum. Au Kilimandjaro (Tanzanie), Protea afra subsp. kilimandscharica est associé à Erica mannii, E. arborea, E. trimera subsp. meruensis, Adenocarpus mannii, Anthospermum usambarense (Hedberg, 1951 : 186). Du Rwenzori, Hauman (1933) évoque aussi un fourré à Hypericum bequaertii et Erica trimera.
En lien spatial avec le Vaccinio stanleyi-Agaristetum salicifoliae, il existe aussi une lande (tableau 15 de Lewalle 1972 : relevés 2 à 6) : Alvesia clerodendroides (Lamiaceae) V, Helichrysum cymosum V, Loudetia simplex (Poaceae) IV, Eragrostis hispida III, Helichrysum tillandsiifolium III, Exotheca abyssinica (Poaceae) III, Pteridium aquilinum III, Crotalaria caudata (Fabaceae) III, Hemionitis inaequalis (Pteridaceae) III, Microchloa kunthii (Poaceae) II, Monocymbium ceresiiforme (Poaceae) II, Lycopodium clavatum II, Viola eminii II, Anemia lepigera II, Bulbostylis atrosanguinea (Cyperaceae) II, Otiophora lebruniana *devrediana I, Lobelia holstii I, soit l’Helichryso cymosi-Alvesietum clerodendroidis Lewalle & B. Foucault ass. nov. hoc loco, typus nominis hoc loco : composante hémicryptophyto-chaméphytique du relevé 3 du tableau 15 in Lewalle (1972, Bull. Jard. Bot. Natl. Belg. 42 (1-2) : 142-145, en changeant Plectranthastrum clerodendroides en Alvesia c.). Notons la présence de Lycopodium clavatum, hôte de plusieurs types de landes à travers le monde.
10. Conclusion
Cette synthèse sur la végétation africaine abordée selon une approche moderne a donc permis de présenter la végétation aquatique vivace (herbiers, prairies flottantes, roselières, avec deux alliances nouvelles), la végétation littorale (une alliance), des mégaphorbiaies tropicales et oro-africaines (une alliance), diverses mares temporaires (une alliance) et des inselbergs (deux alliances). La végétation arborescente des forêts du mont Kenya a fait l’objet d’une partie importante avec la reconnaissance de sept alliances, trois ordres et deux classes ; une alliance et un ordre nouveaux se rapportent par ailleurs à la végétation arborescente des forêts de basse altitude. Enfin la considération de pelouses à Alchemilla et de fourrés afro-alpins a permis la définition de deux autres alliances.
Annexe 1 : Édouard Joshua Adjanohoun (1928-2016)
(d’après Wikipedia [31/12/2024] ; photo 8)

Édouard Joshua Adjanohoun est un universitaire béninois né le 5 novembre 1928 à Ouidah et mort le 16 janvier 2016. Il a été le premier botaniste noir et premier recteur de l’université du Dahomey aujourd’hui Université d’Abomey-Calavi (UAC), boulevard Édouard-Adjanohoun, à l’entrée du campus d’Abomey-Calavi.
Après avoir obtenu, en juin 1956, un DESS en sciences naturelles à l’université d’Orsay, il soutient en 1963 dans la même université une thèse d’État consacrée à la végétation en Côte-d’Ivoire, publiée en 1964. En 1970, il crée le jardin botanique et zoologique de l’université d’Abomey-Calavi. Il a été professeur titulaire de la chaire de botanique et biologie végétale de l’université d’Abidjan en 1966, professeur de botanique et de biologie végétale à l’université de Bordeaux-III de 1976 à 1990. Il a également été vice-président du Conseil scientifique africain, président du Comité interafricain de l’Organisation de l’unité africaine pour la médecine traditionnelle et les plantes médicinales africaines en 1988.
Recteur honoraire de l’université nationale du Bénin, il est titulaire de la chaire de botanique en 1982. Il crée et dirige le Centre pilote régional de la biodiversité africaine des Jardins botanique et zoologique du campus universitaire d’Abomey-Calavi au Bénin. En septembre 2012, les jardins, l’allée centrale de l’université et le bâtiment du rectorat prennent son nom. Il est professeur émérite de l’université Bordeaux-Montaigne de 1990 à 2003.
Chercheur chevronné, le professeur E.J. Adjanohoun a formé plus de cent-dix docteurs et est auteur de plus de deux cents travaux scientifiques dans les domaines de la botanique, de l’ethnobotanique, de l’écologie végétale, de la phytosociologie, de la gestion et de la conservation des ressources phytogénétiques. C’est encore à lui que l’Université d’Abomey-Calavi doit l’Herbier national du Bénin et le célèbre jardin botanique actuellement baptisé Jardin botanique et zoologique Édouard-Adjanohoun. Membre fondateur de plusieurs groupes de recherche et sociétés savantes dans le monde, il a dirigé des missions ethnobotaniques dans douze pays francophones d’Afrique, de l’océan Indien, des Caraïbes puis dans cinq pays de l’Afrique centrale et occidentale.
Annexe 2 : Karl Olov Hedberg (1923-2007)
(d’après Wikipedia [31/12/2024])
Né le 19 octobre 1923 et décédé le 24 septembre 2007, K. O. Hedberg fut professeur de botanique systématique à l’université d’Uppsala, en Suède, de 1970 à 1989 et éditeur de la Flora of Ethiopia. Il a été un pionnier dans la connaissance scientifique de la végétation afro-alpine. Notre connaissance actuelle de cet écosystème doit beaucoup aux recherches qu’il a menées avec sa femme Inga sur le Rwenzori et d’autres hautes montagnes d’Afrique de l’Est. Ses idées novatrices sont basées sur leur travail systématique sur le terrain à la fin des années 1940. L’ouvrage intitulé Features of Afroalpine Plant Ecology (Caractéristiques de l’écologie végétale afro-alpine) reste jusqu’à aujourd’hui un point de repère dans la recherche écologique sur les montagnes équatoriales et est toujours disponible dans une réédition en fac-similé de 1995.
En 1981, il a été élu membre de l’Académie royale suédoise des sciences. Il a été reconnu comme docteur jubilaire lors de la Promotion doctorale linnéenne à l’université d’Uppsala en 2007. Il mettait à jour le manuscrit sur les Ombellifères (préparé par Vernon Heywood, Stephen Jury et d’autres) lorsqu’il est décédé en 2007.
Il était membre de la British Mycological Society, a contribué aux herbiers du Natural History Museum (BM), du Jardin botanique national de Belgique (BR), des musées nationaux du Kenya (East African Herbarium), de l’Herbier national (Éthiopie), des Royal Botanic Gardens, Kew (K), du Muséum national d’histoire naturelle (P) et du Swedish Museum of Natural History Department of Phanerogamic Botany (S).
Molau (Nordic J. Bot. 8 : 194, 1988) lui a dédié le genre hémiparasite afrotropical de trois espèces Hedbergia (Orobanchaceae) proche de Bartsia.
Bibliographie
Adjanohoun E., 1964. Végétation des savanes et des rochers découverts en Côte-d’Ivoire centrale. Mémoires de l’ORSTOM 7 : 1-178.
Bamps P. & Léonard J., 2005. José Lewalle (1931-2004). Systematics and geography of plants 75 : 3-6.
Blundell M., 1987. Collins Guide to the Wild Flowers of East Africa. Collins, 464 p.
Bussmann R.W. & Beck E., 1995. The forests of Mt. Kenya (Kenya), a phytosociological synopsis. Phytocoenologia 25 (4) : 467-560.
Dorrstock S., Porembski S. & Barthlott W., 1996. Ephemeral flush vegetation on inselbergs in the Ivory Coast (West Africa). Candollea 51 (2) : 407-419.
Foucault B. (de), 1988. Les végétations herbacées basses amphibies : systémique, structuralisme et synsystématique. Dissertationes Botanicae 121 : 1-150.
Foucault B. (de), 2014. 5 – Aperçu phytosociologique. In Comptes-rendus de la 148e session extraordinaire de la Société botanique de France en Guyane française (19-29 juillet 2012). Journal de botanique de la Société botanique de France 65 : 41-54.
Foucault B. (de), 2018. Contribution au prodrome des végétations de France : les Montio-Cardaminetea amarae Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, 3e série, 7 : 1-86.
Foucault B. (de), 2022. Miscellanées phytosociologiques III : syntaxons nouveaux, validés, corrigés ou complétés. Carnets botaniques 112 : 1-23, https://doi.org/10.34971/6AX1-0K40.
Foucault B. (de), 2024. La végétation hydrophile à hygrophile de l’Ader Doutchi (République du Niger). Carnets botaniques 213 : 1-16, https://doi.org/10.34971/ZRPA-8689.
Foucault B. (de), Akpagana K., Batawila K. & Bouchet Ph., 1999. Contribution à une étude phytosociologique des végétations inondables du Sud-Togo. Belgian Journal of Botany 132(2) : 141-152.
Foucault B. (de), Akpagana K., Batawila K. & Bouchet Ph., 2000. Contribution à une étude phytosociologique du littoral sableux du Togo. Acta Botanica Gallica147 (4) : 333-344.
Germain R., 1952. Les associations végétales de la plaine de la Ruzizi (Congo belge) en relation avec le milieu. Publications de l’Institut national pour l’étude agronomique du Congo belge, série scientifique, 52 : 1-321.
Guillaumet J.-L., 1967. Recherches sur la végétation et la flore de la région du bas Cavally (Côte-d’Ivoire). Thèse de doctorat ès sciences, Orsay, 247 p., publiée par l’Orstom, Paris.
Guillaumet J.-L., 2009. La végétation des montagnes du Rift. In B. Hirsch & B. Roussel, Le Rift est-africain, une singularité plurielle, IRD et Publications scientifiques du MNHN, chapitre 12 : 195-204.
Guillaumet J.-L. & Adjanohoun E., 1971. La végétation de la Côte-d’Ivoire. In J.-M. Avenard, M. Eldin, G. Girard, J. Sircoulon, P. Touchebeuf de Lussigny, J.-L. Guillaumet, E. Adjanohoun & A. Perraud, Le milieu naturel de la Côte-d’Ivoire, Mémoires de l’ORSTOM 50 : 157-263.
Hambler D.J., 1964. The vegetation of granitic outcrops in Western Nigeria. Journal of Ecology 52 (3) : 573-594.
Hauman L., 1933. Esquisse de la végétation des hautes altitudes sur le Ruwenzori. Bulletins de l’Académie royale de Belgique, 5e série, XIX : 602-616.
Hauman L., 1934. Les « Lobelia » géants des montagnes du Congo belge. Mémoires de l’Institut royal colonial belge, section des sciences naturelles et médicales, II (1) : 1-52.
Hedberg O., 1951. Vegetation belts of the east-African mountains. Svensk Bot Tid 45 : 140-203.
Hedberg O., 1964. Features of afroalpine plant ecology. Acta Phytogeographica Suecica 49 : 1-144.
Jaeger P. & Adam J.-G., 1981. La prairie montagnarde des monts Loma (Sierra Leone). Boletim da Sociedade Broteriana LIII, 2e série : 1341-1396.
Kerfoot O., 1963-64. A preliminary account of the vegetation of the Mbeya Range, Tankanyika. Kirkia 4 : 191-202.
Lebrun J., 1947. La végétation de la plaine alluviale au sud du lac Édouard. Institut des parcs nationaux du Congo belge, Bruxelles, 800 p. et LII planches.
Lebrun J., 1969. La végétation psammophile du littoral congolais. Mémoires de l’Académie royale des sciences d’outre-mer, classe des sciences naturelles et médicales, n. s., 18 (1) : 1-166.
Lewalle J., 1972. Les étages de végétation du Burundi occidental. Bulletin du Jardin botanique national de Belgique 42 (1-2) : 1-247.
Lindeman J.C., 1953. The vegetation of coastal region of Surinam. Mededelingen van het Botanisch Museum en Herbarium van de Rijksuniversiteit te Utrecht 113 (1) : 1-135.
Lisowski S. & Malaisse F., 1989. Groupements végétaux des mares et des anses calmes des rivières du plateau des Kundelungu. In J.-J. Symoens (éd.), Exploration hydrobiologique du bassin du lac Bangweolo et du Luapula, Bruxelles : 5-41.
Mabberley D.J., 1975. Notes on the vegetation of the Cherangani Hills, N.W. Kenya. Journal of the East Africa natural history Society and national Museum 150 : 1-11.
Mandango M.A., Ngaki N. & Kayumba L., 1996. Groupes écologiques de la végétation de l’île Tundulu à Kisangani (Zaïre). Documents phytosociologiques, n. s., XVI : 193-203.
Mullenders W., 1954. La végétation de Kaniama (Entre-Lubishi-Lubilash, Congo belge). Publications de l’Institut national pour l’étude agronomique du Congo belge, série scientifique, 61 : 1-499 et XVIII planches.
Oumorou M. & Lejoly J., 2003. Écologie, flore et végétation de l’inselberg Sobakpérou (Nord-Bénin). Acta Botanica Gallica 150 (1) : 65-84.
Paradis G., 1976. Contribution à l’étude de la flore et de la végétation littorales du Dahomey. Bulletin du Muséum d’histoire naturelle, 3e série, 383, 26 : 33-60.
Parmentier I., 2001. Premières études sur la diversité végétale des inselbergs de Guinée-Équatoriale continentale. Systematics and geography of plants 71 : 911-922.
Parmentier I., 2003a. Étude de Ia végétation des inselbergs de la forêt dense d’Afrique centrale atlantique. Thèse de doctorat, Université libre de Bruxelles, 85 p. (résumé in Acta Botanica Gallica, 2005, 152 (2) : 403-408).
Parmentier I., 2003b. Study of the vegetation composition in three inselbergs from continental Equatorial Guinea (Western Central Africa): effects of site, soil factors and position relative to forest fringe. Belgian Journal of Botany, 136 (1) : 63-72.
Parmentier I., Lejoly J. & Nguema N., 2001. La végétation des inselbergs de Piedra Nzas (Guinée-Équatoriale continentale). Acta Botanica Gallica 148 (4) : 341-365.
Porembski S., Mund J.-P., Szarzynski J. & Barthlott W., 1996. Ecological conditions and floristic diversity of an inselberg in the savanna zoner of Ivory Coast: Mt Niangbo. In J.-L. Guillaumet, M. Belin & H. Puig, Phytogéographie tropicale : réalités et perspectives, ORSTOM éditions : 251-261.
Portères R., 1946. Climat et végétation sur la chaîne des Bambuttos (Cameroun). Bulletin de la Société botanique de France 93 (9) : 352-360
Raynal A. & Raynal J., 1961. Observations botaniques dans la région de Bamako. Bulletin de l’Institut français d’Afrique noire, série A, 23 : 994-1021.
Schnell R., 1952a. Contribution à une étude phytosociologique et phytogéographique de l’Afrique occidentale : les groupements et les unités géobotaniques de la région guinéenne. Mémoires de l’Institut français d’Afrique noire 18 : 43-234.
Schnell R., 1952b. Végétation et flore de la région montagneuse du Nimba. Mémoires de l’Institut français d’Afrique noire 22 : 1-604.
Schnell R., 1976. Flore et végétation de l’Afrique tropicale, 1. Gauthier-Villars, Paris, 468 p.
Schnell R., 1977. Flore et végétation de l’Afrique tropicale, 2. Gauthier-Villars, Paris, 375 p.
Sillans R., 1958. Les savanes de l’Afrique centrale française ; essai sur la physionomie, la structure et le dynamisme des formations végétales ligneuses de l’Oubangui-Chari. Thèse de doctorat ès sciences, université de Montpellier, 331 p.
Trochain J., 1940. Contribution à l’étude de la végétation du Sénégal. Mémoires de l’Institut français d’Afrique noire 2 : 1-433.
Vanden Berghen C., 1979. La végétation des sables maritimes de la basse Casamance méridionale (Sénégal). Bulletin du Jardin botanique national de Belgique 49 (3-4) : 185-238.
Vanden Berghen C., 1990. La végétation des sables maritimes de la Casamance (Sénégal méridional). Lejeunia, n. s., 133 : 1-84.
Yongo O.-D. & de Foucault B., 2007. Contribution à une caractérisation de la végétation de la forêt de Ngotto (République Centrafricaine). Acta Botanica Gallica 154 (2) : 203-220.


