
Miscellanées phytosociologiques X : syntaxons nouveaux, validés ou complétés
Title
Phytosociological miscellanées X: new, validated or completed syntaxa
Résumé
Ce dixième numéro de Miscellanées phytosociologiques apporte de nouvelles données sur des fourrés xérophiles, des landes et tourbières, des pelouses thérophytiques amphibies et xérophiles et surtout des forêts s’étendant des monts Cantabriques à la Slovénie, ainsi que qu’au Danemark.
Abstract
This tenth issue of Miscellanées phytosociologiques provides new data on xerophilous shrubs, heaths and peat bogs, amphibious and xerophilous therophytic grasslands and, above all, forests stretching from the Cantabrian Mountains to Slovenia, as well as Denmark.
Cette dixième livraison de Miscellanées phytosociologiques apporte quelques précisions syntaxonomiques, quelques corrections et validations, et surtout de nouvelles réinterprétations sur des syntaxons forestiers allant des de montagnes ibéro-cantabriques à la Slovénie et au Danemark.
1. Des boisements ibéro-cantabriques
Le tableau 1 synthétise la végétation herbacée du sous-bois de plusieurs forêts ibéro-cantabriques souvent décrites comme « Polysticho setiferi-Fraxinetum Tüxen & Oberd. 1958 » ou « Hyperico androsaemi-Alnetum glutinosae Rivas Mart. in Loidi 1983 ». Les végétations arborescentes et arbustives ont été laissées de côté car leur interprétation restait assez délicate ; pour les premières, ce sont des aulnaies (à Alnus glutinosa), des chênaies-frênaies (à Quercus robur, Fraxinus excelsior)… et pour les secondes souvent des saulaies arbustives à Salix atrocinerea.
Dans ces données du tableau 1, deux unités supérieures se séparent. L’ensemble formé par les colonnes 1 à 8 correspond à des boisements mésophiles caractérisés par le groupe sociologique à Helleborus viridis. Les colonnes 2 à 7 paraissent définir de simples variations d’une large association pouvant être nommée Helleboro viridis-Mercurialetum perennis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 1 du tableau 109 in Durán Gómez (2023, Paisaje vegetal del antiguo valle de Alfoz… : 484-485, en changeant Helleborus occidentalis en H. viridis, incl. subsp. occidentalis). La colonne 8 en est assez distincte : Pulmonario affinis-Polystichetum setiferi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 5 du tableau 39 in Loidi (1983, Estudio de la flora y vegetación de las cuencas… : 122-123) publié ici :
De Vergala à Gorla, 230 m d’altitude, 200 m², 21 taxons (hors juvéniles de taxons ligneux)
- combinaison caractéristique : Lonicera periclymenum *p. 2.2, Hypericum androsaemum1, Arum italicum 1.1, Dioscorea communis 1.1, Polystichum setiferum +.2, Pulmonaria affinis +.2, Ranunculus tuberosus +, Brachypodium sylvaticum +.2, Ruscus aculeatus +.2, Viola reichenbachiana +, Poa nemoralis *n. +.2, Dryopteris filix-mas +, Rubia peregrina *p. +, Athyrium filix-femina +.2, Struthiopteris spicant +,
- autres taxons : Hedera helix5, Rubus sp. 2.2, Ajuga reptans 1.2, Cystopteris fragilis +, Betonica officinalis 1.1, Pteridium aquilinum +.
L’ensemble formé par les colonnes 9 à 16 correspond à une végétation plus hygrophile comme le montre la présence de taxons de mégaphorbaies (Carex pendula…) et de microphorbiaies. Les colonnes 9 à 11 peuvent être décrites comme Hyperico androsaemi-Dryopteridetum affinis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 9 du tableau 93 in Durán Gómez (2023, Paisaje vegetal del antiguo valle de Alfoz… : 456-458). La colonne 13 est définie comme Crepido lampsanoidis-Polystichetum setiferi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 5 du tableau 2 in Amigo et al. (1987, V Jornadas de fitosociologia, Vegetacion de riberas de agua dulce, Universidad de la Laguna, Ser. Informes 22 : 171-172). La colonne 14 est reconnue comme Senecioni bayonnensis-Caricetum reuterianae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 17 du tableau 1 in Amigo et al. (1987, V Jornadas de fitosociologia, Vegetacion de riberas de agua dulce, Universidad de la Laguna, Ser. Informes 22 non paginé, en changeant Carex broteriana en C. reuteriana). Affine de la précédente, la colonne 15 est plutôt le Valeriano pyrenaicae-Polystichetum setiferi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 11 du tableau 3 in Amigo et al. (1987, V Jornadas de fitosociologia, Vegetacion de riberas de agua dulce, Universidad de la Laguna, Ser. Informes 22 non paginé).
Cette végétation herbacée intraforestière est foncièrement aérohygrophile comme le montre l’importance des fougères telles que Dryopteris affinis subsp. a., D. dilatata, Struthiopteris spicant, Polystichum setiferum, Athyrium filix-femina, Asplenium scolopendrium. À ce titre elle se rapproche de végétations océaniques armoricaines rattachées au Primulo vulgaris-Ruscion aculeati (de Foucault, 2020). Pour tester leur éventuel rattachement à ce dernier, on va s’appuyer sur le tableau 2 qui synthétise les deux ensembles du tableau 1 dans les colonnes H-Ps et C-Da de ce tableau 2, en y associant la colonne synthétique de l’alliance armoricaine en P-Ra (d’après de Foucault, 2020, tableau H2 : colonne PR), ainsi que, en colonne H-Ug, la donnée de Vanden Berghen (1968) décrivant la végétation herbacée du sous-bois de son tableau I (« ormaie à Ulmus glabra et Hypericum androsaemum »). L’étude de ce tableau 2 montre d’abord que les deux colonnes ibéro-cantabriques correspondent à deux alliances distinctes, celles-ci par ailleurs non réductibles au Primulo-Ruscion aculeati. On peut donc reconnaître
- pour la colonne H-Ps, l’Helleboro viridis-Polystichion setiferi nov. hoc loco, caractérisé ou différencié des alliances voisines (et surtout de la suivante) par Helleborus viridis, Rubia peregrina subsp. longifolia, Asplenium onopteris, Betonica officinalis, Polygonatum multiflorum, Symphytum tuberosum, Iris foetidissima, Mercurialis perennis, Hepatica nobilis, Sanicula europaea, Carex sylvatica; typus nominis hoc loco : le Pulmonario affinis-Polystichetum setiferi nov. ;
- pour la colonne C-Da, le Carici pendulae-Dryopteridion affinis nov. hoc loco, caractérisé ou différencié des alliances voisines (surtout de la précédente) par Carex pendula, C. remota, C. reuteriana subsp. r., C. paniculata subsp. lusitanica, Filipendula ulmaria, Lysimachia nemorum, Equisetum telmateia *t., Solanum dulcamara, Cyclosorus pozoi, Lythrum salicaria, Eupatorium cannabinum, Jacobaea aquatica, Chaerophyllum hirsutum, Senecio bayonnensis (photo 1), Osmunda regalis, Luzula sylvatica subsp. henriquesii, Oenanthe crocata, Myosotis martinii, Mentha aquatica, Valeriana pyrenaica; typus nominis hoc loco : le Crepido lampsanoidis-Polystichetum setiferi nov.
Ces deux alliances s’intègrent clairement aux Oxalidetalia acetosellae B. Foucault 2020 et aux Anemono nemorosae-Caricetea sylvaticae Gillet ex Gillet & B. Foucault in B. Foucault 2020, tout comme la colonne H-Ug, celle-ci cependant à intégrer dans une alliance encore différente de celles citées ici, qui ne peut d’ailleurs être définie dans ce document car les données sources sont synthétiques.

2. Des boisements de Croatie occidentale et du Kosovo
Dans leur beau tableau 1, Jelinčić et al. (2024) complètent l’étude phytosociologique de l’Epimedio alpini-Carpinetum betuli (Horvat 1938) Bohridi 1963 en Croatie occidentale. La végétation arborescente est surtout caractérisée par Carpinus betulus (V sur 14 relevés), Quercus robur (V), Tilia cordata (IV), Fraxinus ornus (III), Acer campestre (III), Quercus cerris (II)… : Fraxino orni-Carpinetum betuli ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (A) du relevé 6 du tableau 1 in Jelinčić et al. (2024, Šumarski list (1-2) : 11-15). La végétation arbustive du sous-bois a été récemment (de Foucault, 2026) rattachée à l’Euonymo verrucosi-Cornetum maris dans une race à Staphylea pinnata (photo 2).
La végétation herbacée du sous-bois est très originale, combinant des fougères aérohygrophiles (Polystichum setiferum, P. aculeatum, Dryopteris filix-mas, Asplenium scolopendrium, Athyrium filix-femina), des géophytes vernales (Epimedium alpinum, Galanthus nivalis, Erythronium dens-canis, Crocus vernus, Isopyrum thalictroides, Asarum europaeum, Anemone nemorosa, Scilla bifolia, Corydalis solida…) : Hacquetio epipactidis-Epimedietum alpini ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (C) du relevé 6 du tableau 1 in Jelinčić et al. (2024, Šumarski list (1-2) : 11-15).
De leur côté, dans leur tableau 1, Berisha et al. (2019) décrivent un « Ramondo-Ostryetum carpinifoliae » où d’évidence Ramonda nathaliae caractérise une végétation strictement chasmophytique ombragée par des arbres (surtout Ostrya carpinifolia, Fraxinus ornus, Acer campestre ; voir leur figure 4) et une communauté arbustive ainsi constituée (sur 10 relevés) : Cotoneaster tomentosus V, Euonymus latifolius IV, Rosa canina II, Crataegus monogyna II, Juniperus communis *c. +, soit l’Euonymo latifolii-Cotoneastretum tomentosi ass. nov hoc loco, typus nominis hoc loco : composante arbustive du relevé 9 du tableau 1 in Berisha et al. (2019, Hacquetia 18 (2) : 326-327). Au contact d’une pelouse thérophytique à Bupleurum flavicans, Clinopodium acinos, Sedum caespitosum et Arenaria serpyllifolia, la végétation chasmophytique est caractérisée par Ramonda nathaliae (V), Saxifraga sempervivum (IV), Thliphthisa purpurea (IV), Arabis procurrens (III), A. caucasica (III), Achnatherum calamagrostis (III), Cytisus hirsutus (III), Centaurea stoebe (II), Teucrium chamaedrys (II), Sedum acre (II), Hylotelephium telephium (II), Pseudoturritis turrita (II), Helleborus odorus (II), Asplenium ruta-muraria *r. (I), A. trichomanes (I), A. ceterach (I) et des taxons de présence + : Saxifrago sempervivi-Ramondetum nathaliae ass. nov hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 5 du tableau 1 in Berisha et al. (2019, Hacquetia 18 (2) : 326-327), à placer dans le Ramondion nathaliae Horvat ex Simon 1958, manifestement vicariant du Saxifrago longifoliae-Ramondetum myconi Braun-Blanq. in Meier & Braun-Blanq. 1934 pyrénéen et des Ramondeta nathaliae serpentinicoles tels que le Scorzonero austriacae-Ramondetum nathaliae Stevanović & Matevski in Stevanović et al. 2014 prov. et l’Asplenio cuneifolii-Ramondetum nathaliae Stevanović & Stevanović 1985 (dans les deux cas ‘…-Ramondaetum…’ art. 41b). Stevanović et al. (2014) placent ces associations serpentinicoles dans le Ramondion nathaliae serpentinicum Stevanović & Matevski in Stevanović et al. (2014, Botanica Serbica 38 (1) : 88) nom. inval. et illeg. (art. 3e, 3o, 5, 34a), qu’on peut valider en Asplenio cuneifolii-Ramondenion nathaliae (Stevanović & Matevski 2014) suball. nov. hoc loco, typus nominis hoc loco : l’Asplenio cuneifolii-Ramondetum nathaliae Stevanović & Stevanović 1985 (Bull. Mus. Hist. Nat. Belgrade, B, 40 non consulté), caractérisé par des taxons liés aux rochers de serpentinites tels que Hemionitis marantae, Asplenium cuneifolium et sans doute d’autres géographiquement plus localisés, voire endémiques. Elle s’oppose à l’Eu–Ramondenion nathaliae non serpentinicole. Une synthèse plus complète montrera peut-être que la première sous-alliance pourra être relevée au rang d’alliance.

3. Des forêts d’Italie
Hruska (1988) a étudié la végétation de Monti della Laga, au nord-est de Rome, en Italie centro-orientale. Son tableau 1 décrit la châtaigneraie acidiphile locale relevée entre 600 et 990 m d’altitude. La végétation arborescente correspond à la liste suivante (sur 15 relevés) : Castanea sativa V, Sorbus torminalis III, S. domestica II, Ostrya carpinifolia II, Prunus avium II, Quercus pubescens II, Q. petraea II, Acer obtusatum II, Fraxinus ornus II et de présence I Populus tremula, Fagus sylvatica, Quercus cerris, décrivant l’Ostryo carpinifoliae-Castaneetum sativae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 3 du tableau 1 in Hruska (1988, Braun-Blanquetia 2 : 120-121).
La végétation arbustive du sous-bois étant peu structurée, passons à la végétation vivace : Melampyrum italicum V, Teucrium siculum V, Hieracium lachenalii V, Pteridium aquilinum IV, Festuca heterophylla IV, Viola reichenbachiana IV, Luzula forsteri III, Veronica officinalis III, Solidago virgaurea *v. III, Clinopodium vulgare III, Campanula trachelium III, Brachypodium sylvaticum III, Anthoxanthum odoratum III, Poa nemoralis *n. III, Geranium nodosum III, Circaea lutetiana III, Hypericum montanum III, H. androsaemum II, Genista tinctoria II, Chamaecytisus hirsutus II, Danthonia decumbens II, Serratula tinctoria II, Campanula persicifolia II, Helleborus multifidus *bocconei II, Oenanthe pimpinelloides II, Brachypodium rupestre II, Pyrola minor II, Astragalus glycyphyllos II, Pentanema squarrosum II, Fragaria vesca II, Melica uniflora II, Hepatica nobilis II, Prenanthes purpurea II, Dryopteris filix-mas II, Stachys sylvatica II, Euphorbia amygdaloides II, Carex sylvatica II, Veronica urticifolia II, Primula vulgaris II, Sanicula europaea II, Salvia glutinosa II, Daphne laureola II, Lactuca muralis II, Platanthera bifolia II, Agrimonia agrimonioides II, A. eupatoria II, Anemone nemorosa II et de présence I Vaccinium myrtillus, Geranium sanguineum, Betonica officinalis, Bromus ramosus, soit le Teucrio siculi-Melampyretum italici ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 11 du tableau 1 in Hruska (1988, Braun-Blanquetia 2 : 120-121).
De leur côté, Tammaro et Poldini (1988) ont décrit une forêt développée sur le versant adriatique. Dans leur tableau 1, on reconnaît une végétation arborescente pouvant se rattacher au Fraxino orni-Carpinion orientalis B. Foucault 2026 récemment décrit (de Foucault, 2026), mais avec Quercus ilex (sur 12 relevés) : Carpinus orientalis V, Quercus ilex V, Q. pubescens V, Fraxinus ornus V, Acer campestre III, Quercus cerris II, Sorbus domestica II, Ulmus minor I, Acer opalus *obtusatum (incl. A. neapolitanum) I, soit le Querco ilicis-Carpinetum orientalis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 1 du tableau 1 in Tammaro & Poldini (1988, Braun-Blanquetia 2 : 129).
La végétation arbustive du sous-bois est notamment caractérisée par Hippocrepis emerus subsp. emeroides, taxon plutôt centre- et est-méditerranéen, mais aussi différentiel d’une alliance italienne, l’Oleo europaeae-Paliurion spinae-christi (Biondi et al. 2014) B. Foucault 2021 (de Foucault, 2021b : 17), à laquelle on peut le rattacher, sans qu’il s’identifie à un syntaxon déjà connu, étant aussi marqué par quelques taxons caractérisant les Rhamno-Prunetea spinosae (Cornus sanguinea, Crataegus monogyna, Ligustrum vulgare) : Corno sanguineae-Hippocrepidetum emeroidis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 1 in Tammaro & Poldini (1988, Braun-Blanquetia 2 : 129). On le retrouve dans le relevé 2 du tableau 3 des mêmes auteurs.
Quant à la végétation herbacée intraforestière, elle est caractérisée par Festuca drymeja : Lithospermo purpureocoerulei-Festucetum drymejae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 2 du tableau 1 in Tammaro & Poldini (1988, Braun-Blanquetia 2 : 129), qui s’intègre bien aux Rubio peregrinae-Asplenietea onopteridis B. Foucault 2020.
Cette fois sur le versant tyrrhénien (est de Salerno), Corbetta et al. (2004) décrivent plusieurs types de végétation, notamment le « Seslerio autumnalis-Aceretum obtusati », où l’on peut notamment mettre en exergue la végétation arborescente (sur 10 relevés) : Acer opalus *obtusatum V, Ostrya carpinifolia V, Quercus cerris V, Fraxinus ornus IV, Pyrus communis *c. II et de présence I Quercus pubescens I, Alnus cordata, Malus sylvestris I, une association originale relevant de l’Aceri obtusati-Quercion cerridis B. Foucault 2025 (de Foucault, 2025), l’Ostryo carpinifoliae-Aceretum obtusati ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 24 in Corbetta et al. (2004, Braun-Blanquetia 36 : 34-35).
4. Des pinèdes de Bulgarie
Tzonev et al. (2018) ont décrit des pinèdes à Pinus nigra subsp. pallasiana, dont ils ont réuni plus de quatre-vingt-dix relevés. Elles viennent compléter celles étudiées en Grèce (de Foucault, 2022b).
Le tableau 3 en synthétise la végétation arborescente en intégrant quatre syntaxons de l’étude citée (colonnes 6 à 9) dans la partie Pinion pallasianae du tableau 1 de la synthèse de 2022b (colonnes 1 à 5). Les colonnes 6 et 7 peuvent être interprétées comme deux variations (à Fagus sylvatica et Quercus frainetto en 6, à Quercus pubescens et Q. petraea en 7) d’un unique Querco dalechampii-Pinetum pallasianae ass. nov. hoc loco, développé en-dessous de 1 040 m d’altitude, typus nominis hoc loco : composante arborescente du relevé 1 du tableau 1 in Tzonev et al. (2018, Hacquetia 17 (2) : 140-147). Les colonnes 8 et 9 occupent les altitudes supérieures à 1 100 m (sauf le relevé 21, à 990 m) et décrivent plutôt l’Abieti albae–Pinetum pallasianae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 22 du tableau 2 in Tzonev et al. (2018, Hacquetia 17 (2) : 148-161).
La végétation du sous-bois arbustif n’est vraiment significative que dans le « Seslerio latifoliae-Pinetum pallasianae haberleetosum » et peut être décrit selon cette liste (en retenant les 19 relevés suivants : 17, 19 à 21, 24, 26 à 28, 30 à 38, 41, 42) : Viburnum lantana IV, Euonymus verrucosus III, Cornus mas III, Corylus avellana III, Cotoneaster integerrimus III, Lonicera xylosteum III, Rosa pendulina III, Juniperus communis *c. II, Clematis vitalba II, Rubus idaeus II, Rhamnus cathartica II et de présence I Lonicera caerulea *c., Crataegus monogyna, Syringa vulgaris, Daphne mezereum, Cotinus coggygria, Juniperus deltoides. On peut l’interpréter comme un remarquable représentant d’altitude de l’Euonymo verrucosi-Cornion maris décrit de Roumanie (de Foucault, 2026), le Cotoneastro integerrimi-Cornetum maris ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 33 du tableau 2 in Tzonev et al. (2018, Hacquetia 17 (2) : 148-161).
Le sous-bois herbacé vivace est synthétisé dans le tableau 4, qui révèle une coupure majeure entre les colonnes 2 et 3. La colonne 1 peut être décrite comme Lathyro laxiflori-Melicetum uniflorae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 14 du tableau 1 in Tzonev et al. (2018, Hacquetia 17 (2) : 140-147). Les colonnes 3 et 4 sont fortement différenciées par le groupe sociologique à Calamagrostis arundinacea :
- en 3, le Galio schultesii-Seslerietum latifoliae ass. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 68 du tableau 2 in Tzonev et al. (2018, Hacquetia 17 (2) : 148-161) ;
- en 4, l’Agrimonio agrimonioidis-Calamagrostietum arundinaceae ass. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 17 du tableau 2 in Tzonev et al. (2018, Hacquetia 17 (2) : 148-161) ; à noter la présence dans cette association d’Haberlea rhodopensis, une Gesneriaceae relictuelle des périodes chaudes du Tertiaire à l’image de Ramonda myconi dans les Pyrénées et R. nathaliae au Kosovo.
5. Des charmaies de Slovénie occidentale
Čušin (2002) a décrit une charmaie à Carex alba, avec deux variations, entre 280 et 400 m d’altitude. Le tableau 5 en synthétise la végétation arborescente, avec
- en colonne 1, une association originale combinant Picea abies, Ostrya carpinifolia et Fraxinus ornus, le Piceo abietis-Ostryetum carpinifoliae ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (E3) du relevé 4 du tableau 1 in Čušin (2002, Hacquetia 1 (1) : 102-107) ;
- en colonne 2, une aulnaie-ormaie proche du Fraxino excelsioris-Alnetum incanae Julve ex B. Foucault & Julve in B. Foucault 2020.
Le tableau 6 rapporte la végétation arbustive du sous-bois, avec en colonne 1 un fourré mésophile pouvant se rattacher au Berberidion vulgaris Tüxen 1962 et en colonne 2 un fourré mésohygrophile pouvant se rattacher au Salici cinereae-Rhamnion catharticae (Géhu et al. 1983) B. Foucault & J.-M. Royer 2016 (de Foucault & Royer, 2016). Le premier peut être décrit comme Daphno mezerei-Cornetum maris ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (E2) du relevé 1 du tableau 1 in Čušin (2002, Hacquetia 1 (1) : 102-107).
Plus originale est la végétation herbacée vivace intraforestière (les deux coefficients séparés par un trait d’union se rapportent à chacune des deux variations, sur 15 et 9 relevés ; 0 indique absence du taxon ; en bold les coefficients des taxons différentiels de variations) :
- combinaison caractéristique : Carex alba V-V, Leucojum vernum V-V, Omphalodes verna V-V, Crocus neapolitanus V-V, Galanthus nivalis V-V, Primula vulgaris V-V, Anemone trifolia V-V, Cyclamen purpurascens V-V, Hepatica nobilis V-V, Vinca minor V-V, Aegopodium podagraria V-V, Mercurialis perennis V-V, Knautia drymeja V-IV, Polygonatum multiflorum V-IV, Carex digitata IV-V, Salvia glutinosa V-IV, Neottia ovata III-V, Aconitum lycoctonum s. l. IV-IV, Pulmonaria officinalis IV-IV, Anemone ranunculoides IV-IV, Brachypodium sylvaticum V-III, Oxalis acetosella III-V, Paris quadrifolia III-V, Fragaria vesca V-III, Allium ursinum III-V, Veratrum nigrum III-IV, Viola reichenbachiana IV-III, Helleborus odorus III-IV, Ornithogalum pyrenaicum III-IV, Galium laevigatum III-III, Euphorbia amygdaloides III-III, Aposeris foetida III-III, Lathyrus vernus *v. III-III, Lamium galeobdolon *flavidum II-IV, Anemone nemorosa III-III, Rubus caesius II-III, Euphorbia carniolica II-III, Melica nutans III-II, Symphytum tuberosum II-III, Arum maculatum II-III, Isopyrum thalictroides II-II, Epimedium alpinum II-II, Campanula trachelium II-II, Dryopteris filix-mas II-II, Thalictrum aquilegiifolium II-II, Senecio ovatus II-II, Lathraea squamaria I-II, Colchicum autumnale I-II, Epipactis helleborine s. l. II-I, Cardamine trrifolia II-I, pentaphyllos II-I, C. bulbifera I-I, C. enneaphyllos 0-I, Angelica sylvestris *s. I-I, Actaea spicata I-I, Lilium martagon I-I, Stellaria nemorum *montana 0-I, Aruncus dioicus I-0, Clematis recta I-0, Campanula rapunculoides I-0,
- taxons différentiels de variation
- 1) Cruciata glabra V-I, Scilla bifolia IV-II, Allium carinatum III-I, Heracleum sphondylium *s. III-0, Aquilegia vulgaris s. l. III-0, Neottia nidus-avis II-0, Vincetoxicum hirundinaria II-0, Viola riviniana II-0, Peucedanum verticillare II-0,
- 2) Carex sylvatica I-V, Lamium orvala II-V, Cerastium sylvaticum I-IV, Asperula taurina II-IV, Filipendula ulmaria 0-III, Geum urbanum I-III, Petasites hybridus 0-II, Euphorbia dulcis s. l. 0-II, Ranunculus lanuginosus +-II, Adoxa moschatellina 0-II, Ficaria verna +-II, Gagea lutea 0-II, Glechoma hederacea 0-II.
Cette association appartient au groupe des sous-bois géophytiques vernaux représenté en France par le Scillion bifoliae Gillet ex Gillet & B. Foucault in B. Foucault 2020 et peut être défini comme Omphalodo vernae-Caricetum albae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace (E1) du relevé 6 du tableau 1 in Čušin (2002, Hacquetia 1 (1) : 102-107). La variation 1 en est le typicum, différencié par Cruciata glabra, Scilla bifolia…, la 2 est le caricetosum sylvaticae subass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace (E1) du relevé 18 du tableau 1 in Čušin (2002, Hacquetia 1 (1) : 102-107).
6. Des boisements du Danemark
Au moyen de deux tableaux, le phytosociologue néerlandais Sissingh (1970) rapporte des données sur des hêtraies du Danemark. Dans ces boisements, la végétation arbustive est assez pauvre. Sur hydromull (tableau 1 de cette publication de 1970), on relève Viburnum opulus, Rubus idaeus, Ribes uva-crispa, Corylus avellana, Crataegus laevigata, Sambucus nigra, une végétation qu’il serait possible de placer dans le Rubo idaei-Viburnion opuli B. Foucault & J.-M. Royer 2016 à caractère boréo-montagnard (de Foucault & Royer, 2016). Sur moder-mor (le tableau 2), ce sont plutôt Frangula alnus subsp. a., Rubus idaeus, avec de jeunes Sorbus aucuparia, qui orientent cette végétation vers le Sorbo aucupariae-Franguletum alni Julve & Gillet ex B. Foucault, Julve & Gillet in B. Foucault 2018 connu de Pologne et du nord de la France ; l’autorité de cette association écrite initialement comme ‘Julve & Gillet ex B. Foucault 1994’ et reprise par de Foucault & Royer (2014 : 86 et 96) est incorrecte.
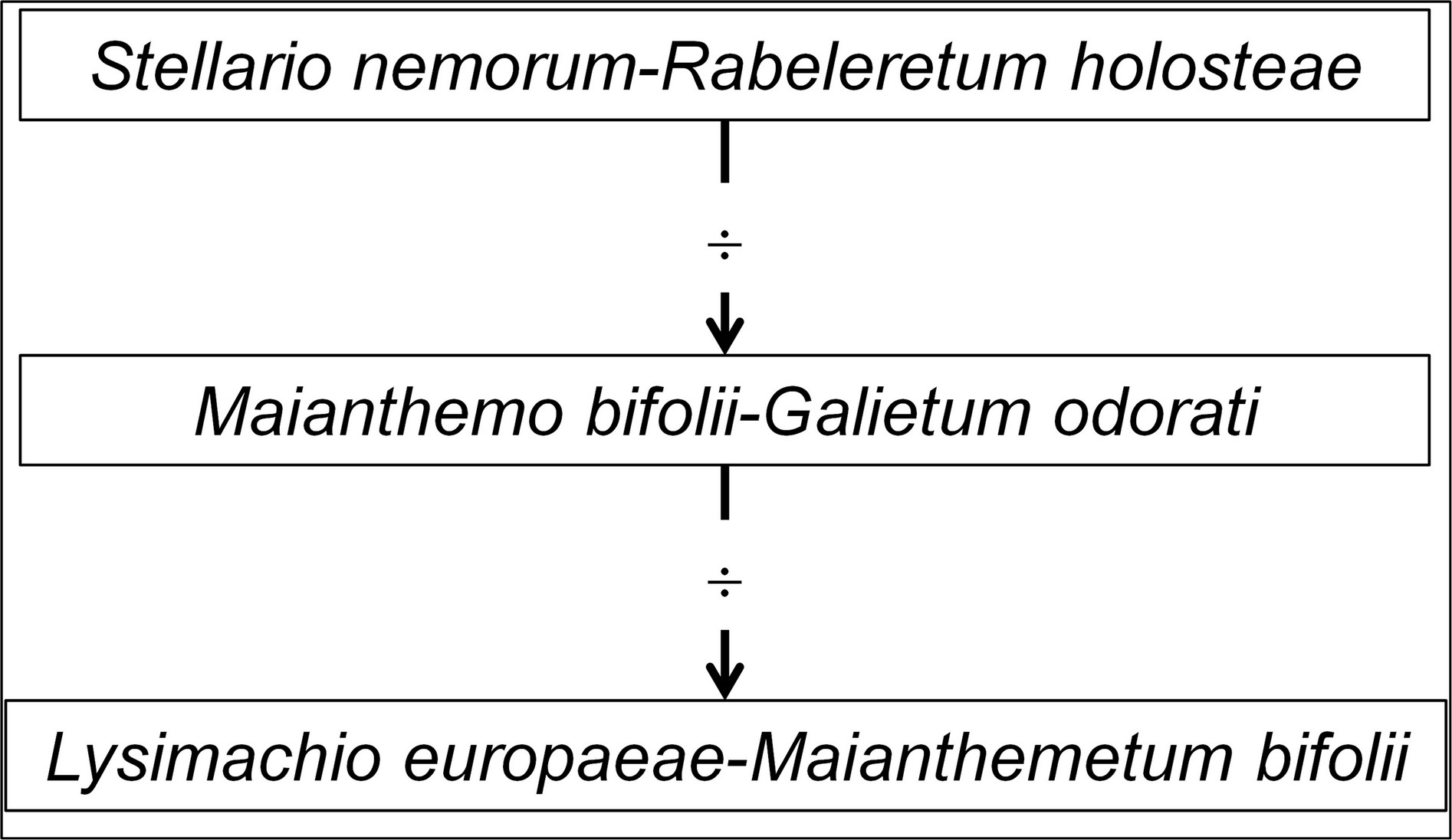
Le tableau 7 hoc loco met en évidence trois syntaxons herbacés intraforestiers. La colonne 1 décrit une association sur hydromull actif caractérisée notamment par Stellaria nemorum, que l’on va pouvoir comparer à des associations vicariantes de plus basses latitudes (de Foucault, 2026) : c’est au Lolio gigantei-Stellarion nemorum B. Foucault 2026 qu’elle se rattache le mieux, mais sans s’identifier à l’une des associations connues de cette alliance. On peut donc définir ici le Stellario nemorum-Rabeleretum holosteae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 17 du tableau 1 in Sissingh (1970, Vegetatio 21 (4-6) hors texte en remplaçant Stellaria holostea par Rabelera h.).
La colonne 2 accueille des taxons plus acidiphiles et a perdu les plus exigeants des taxons du Stellario-Rabeleretum au plan édaphique : Maianthemo bifolii-Galietum odorati ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 34 du tableau 1 in Sissingh (1970, Vegetatio 21 (4-6) hors texte).
L’importance des taxons franchement acidiphiles indicateurs de moder/mor incite à rattacher la colonne 3 au Melampyrion pratensis H. Passarge 1967 ex 1969 et à une association nouvelle, le Lysimachio europaeae-Maianthemetum bifolii ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 3 du tableau 2 in Sissingh (1970, Vegetatio 21 (4-6) : 250-251).
Globalement, le tableau 7 décrit une série temporelle d’oligotrophisation (figure 1 ; ¸ symbolise l’oligotrophisation édaphique) menant du Stellario nemorum-Rabeleretum holosteae au Lysimachio europaeae-Maianthemetum bifolii, le Maianthemo bifolii-Galietum odorati s’avérant intermédiaire ; il montre la disparition progressive des taxons de l’hydromull et leur remplacement par ceux du moder/mor, alors que le groupe sociologique à Oxalis acetosella supporte l’oligotrophisation. Le troisième stade pourrait précéder une lande à Vaccinium myrtillus, mais celle-ci n’est pas évoquée par l’auteur néerlandais. On a là une nouvelle réalisation de la série structurale décrite antérieurement (de Foucault, 2020 : 67). Cette interprétation en terme temporel d’une série avant tout spatiale est autorisée par le fait qu’elle se déroule au sein d’un système caractérisé par une géologie et un climat à peu près homogènes : des matériaux d’origine glaciaire (moraines argileuses et dépôts fluvio-glaciaires) recouvrant un sous-sol d’âge crétacé et tertiaire (Nordmann, 1928), climat boréo-atlantique.
Quelques bribes de végétation thérophytique intraforestière montrent une combinaison de Moehringia trinervia, Galeopsis tetrahit, Lapsana communis, plus rarement Impatiens noli-tangere.
7. Des fourrés xérophiles
En 2021, puis en 2023a, j’ai décrit puis apporté des compléments à un beau fourré de moyenne montagne méditerranéenne, l’Aceri monspessulani-Rhamnetum alpinae à partir de relevés réalisés dans les Corbières audoises puis les Cévennes méridionales (de Foucault, 2021a, 2022a, 2023a). En randonnant au-dessus de Montdardier (Gard), dans la forêt domaniale de la Vis (environ N 43° 55’, E 3° 37’), vers 700 m d’altitude, en versant nord, je l’ai revu avec Rhamnus alpina (photo 3), Rh. cathartica, Juniperus communis subsp. c., Prunus mahaleb, Hippocrepis emerus subsp. e., Lonicera xylosteum, Buxus sempervirens, Clematis vitalba, Crataegus monogyna, Amelanchier ovalis subsp. o., Cytisophyllum sessilifolium, Ilex aquifolium, Viburnum lantana, Ribes alpinum et de jeunes Acer monspessulanum (photo 4), Laburnum anagyroides, Fagus sylvatica, Sorbus aria, Fraxinus excelsior, sans avoir la possibilité d’en réaliser un relevé plus précis. Il est à confirmer aussi sur le causse du Larzac, aux environs du Caylar, notamment sur les flancs du Roc Castel (N 43° 51’ 55’’, E 3° 19’ 15’’, vers 740 m).
Sur la montagne de l’Hortus, en face du pic Saint-Loup (Hérault), vers 300 m d’altitude, s’étendent des fourrés à Juniperus phoenicea dont le tableau 8 rapporte deux relevés. Ils rappellent assez le Bupleuro fruticosi-Juniperetum phoeniceae B. Foucault 2022 de l’Aude rangé dans l’Amelanchiero ovalis-Juniperion phoeniceae B. Foucault 2021 (de Foucault, 2021b, 2022), mais ici Erica arborea est remplacé par Arbutus unedo. Pour interpréter ce groupement, le mieux est d’actualiser la synthèse de l’alliance (tableau 9) à partir du tableau 11 de la publication de 2021b en ajoutant le Bupleuro fruticosi-Juniperetum phoeniceae B. Foucault 2022 (colonne BJp d’après de Foucault, 2021b : tableau 12, et 2023a : tableau 2) et le présent groupement (AJp, tableau 8 hoc loco). On voit qu’on ne peut réduire ce dernier au Bupleuro fruticosi-Juniperetum phoeniceae audois et qu’on peut le définir comme Arbuto unedonis-Juniperetum phoeniceae ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 8 hoc loco publié ci-dessous
Valflaunès (34), montagne de l’Hortus, Gourdou, N 43° 49’ 36,6’’, E 3° 51’ 38,7’’, 318 m, le 25/09/2025, 75 %, 100 m², 19 taxons ; photo 5
- Juniperus phoenicea *ph. 3, oxycedrus *o. 2, Amelanchier ovalis *o. 2, Buxus sempervirens 2, Phillyrea media 2, Arbutus unedo 1, Pistacia terebinthus 1, Smilax aspera 1, Quercus coccifera 1, Genista scorpius +, Lonicera implexa +, Rhamnus saxatilis *s. +, Phillyrea latifolia +, Rhamnus alaternus *a. +, Viburnum tinus +, Clematis flammula +,
- autres taxons : Pinus halepensis j 2, Salvia rosmarinus ! 2, Quercus ilex j 2, Prunus mahaleb +.
Annexe du tableau 8 – rel. 1 : Valflaunès (34), montagne de l’Hortus, Gourdou, 25/09/2025 ; rel. 2 : Rouet (34), Gourdou, vers le menhir de Juoilles, 25/09/2025.



8. La végétation des monts de l’Espinouse et du Caroux
Dans l’Hérault, les monts de l’Espinouse (parfois écrit Espinouze) ont été partiellement décrits au plan phytosociologique, mais attendent toujours une monographie plus large. Ils furent l’objet de la thèse de Baudière (1970) et l’examen de celle-ci montre des affinités entre la végétation de ce massif granitique et celle du versant sud de la Montagne Noire (de Foucault, 2023b). Ainsi en page 196, le relevé 558 semble se rattacher au Lavandulo stoechadis-Ericetum cinereae Géhu ex B. Foucault 2021 ; aux pages 263-264, avec Ilex aquifolium et les deux taxons éponymes, on peut reconnaître le Sorbo ariae-Cytisetum scoparii B. Foucault 2019 ; en pages 298, 306 et 314, l’auteur évoque le sous-bois herbacé à géophytes vernales de hêtraies (notamment au pont de la Mouline), Scilla bifolia, Tractema lilio-hyacinthus, Erythronium dens-canis, Corydalis solida, Narcissus pseudonarcissus, Conopodium majus subsp. m., à rattacher au Tractemo lilio-hyacinthi-Erythronietum dentis-canis B. Foucault 2020 ; malheureusement, aucune précision n’est apportée par la publication ultérieure sur les forêts acidiphiles de l’Espinouse (Baudière, 1975a). En page 166, dans la strate arbustive du relevé 32, on reconnaît le Cytiso scoparii-Ericetum arboreae B. Foucault 2015. En pages 164, puis 191 et 199, ainsi que dans Baudière (1972b : 22), dans la végétation herbacée intraforestière, avec Carex distachya, Rubia peregrina et Asplenium onopteris, on reconnaît la classe des Rubio peregrinae-Asplenietea onopteridis B. Foucault 2020.
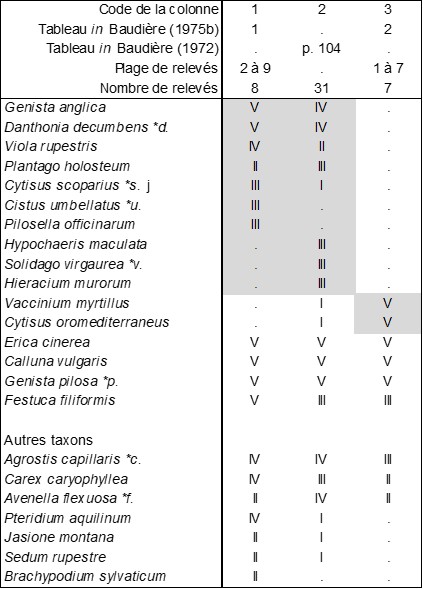
Le même auteur (1972a, 1975b) apporte des données plus précises sur les landes à Ericaceae du même massif, qui ne paraissent pas avoir été prises en compte dans la synthèse récente de Glemarec et al. (2025). Le tableau 10 synthétise cet apport. La synthèse de classe de 2025 montre que la première de ces landes (colonnes 1 et 2) peut se ranger dans l’Erico cinereae-Callunion vulgaris Glemarec et al. 2025, mais ne coïncide avec aucune des associations listées : Genistetum anglico-pilosae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 1 in Baudière (1975b, Colloq. Phytosoc. II : 103). La troisième (colonne 3) se rattache plutôt au Cytiso oromediterranei-Vaccinietum myrtilli B. Foucault 2017 décrit des Cévennes, qui se place dans le Cytiso oromediterranei-Ericion cinereae B. Foucault 2021.
Baudière (1972c) livre aussi une lande du versant nord du Caroux assez différente qui paraît oubliée des phytosociologues, le Vaccinio-Saxifragetum pedatifidae, modernisé ici en Saxifrago prostii-Vaccinietum myrtilli Baudière 1972 nom. mut. et inv. hoc loco, lectotypus nominis hoc loco : relevé 8 du tableau 1 in Baudière (1972c, Bull. Soc. Bot. France 119, 92e sess. extr. dans le Languedoc : 112).
9. La végétation des tourbières bombées
En 2019, dans une contribution à une monographie phytosociologique du pays de Sault (de Foucault, 2019a, tableau 35), j’ai notamment décrit le Calluno vulgaris-Sphagnetum capillifolii. Il s’avère que ce nom est illégitime devant le Calluno vulgaris-Sphagnetum capillifolii Fern. Prieto et al. 1987 (Lazaroa 7 : 464) nord-ouest-ibérique, distinct notamment par Erica tetralix, Carex nigra (subsp. carpetana non retenu par les référentiels actuels), Trichophorum cespitosum subsp. germanicum, l’absence de Molinia caerulea ; on renomme le nom de 2019 en Calluno vulgaris-Sphagnetum magellanici (B. Foucault 2019) nom. nov. hoc loco, typus nominis hoc loco : le même que celui de 2019, soit le relevé 1 du tableau 35 in de Foucault (2019a, Evaxiana 6 : 110).
10. La végétation de pelouses amphibies
Le « Ranunculo paludosi-Lythretum portulae » Molero & Pujadas in Molero 1984 (Molero, 1984, tableau II) est une association biologiquement complexe où l’on peut surtout isoler une végétation thérophytique (sur 8 relevés) : Lythrum thymifolia V, Juncus bufonius V, L. portula IV, Lotus angustissimus IV, Lysimachia talaverae IV, Trifolium dubium V, T. lappaceum IV (photo 6), T. strictum III, Isolepis setacea III, Polypogon monspeliensis III, Linum trigynum III, Leontodon rothii III, Sisymbrella aspera II, Centaurium pulchellum II, Blackstonia perfoliata II, Euphorbia exigua II, Sagina apetala II, Spergularia segetalis (photo 7) II, Juncus pygmaeus II, J. capitatus I, qui semble devoir se rattacher au Centaurio pulchelli-Blackstonion perfoliatae (Müller‑Stoll & Pietsch 1965) B. Foucault ex B. Foucault 2023, dans une association originale, le Trifolio lappacei-Lythretum thymifoliae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 4 du tableau II in Molero (1984, Buttl. Inst. Cat. Hist. Nat. 51, Bot. : 144).
Par ailleurs, Bergmeier et Raus (1999) apportent des données sur quelques végétations amphibies de Grèce reprises ici dans les tableaux 11 (végétation thérophytique) et 12 (végétation vivace). La colonne 1 du tableau 11 est le Myosuro minini-Ranunculetum lateriflori Bergmeier & Raus 1999, une association srpentinicole orophile (relevé type à 1 240 m d’altitude) définie sur la base de seulement deux relevés. La colonne 5 est le Cypero micheliani-Fimbristylidetum bisumbellatae Horvatić 1954 qui paraît devoir être rangé dans le Centaurio pulchelli-Blackstonion perfoliatae (Müller-Stoll & Pietsch 1965). B. Foucault 1988 ex 2023 relié à une prairie vivace à Samolus valerandi et Polypogon viridis (tableau 12 : colonne 4 ; Paspalo distichi-Polypogonion viridis Braun-Blanq. in Braun-Blanq. 1952). La colonne 4 relève du Sporobolion schoenoidis Braun-Blanq. ex Rivas Goday 1956 et du Fimbristylido bisumbellatae-Cyperetum fusci Oberd. 1952 corr. Raus 1997, relié au Paspalo distichi-Polypogonetum viridis Braun-Blanq. 1936 (tableau 12 : colonne 3). Possédant une écologie plus subhalophile, la colonne 6 relève plutôt des Sporoboletea aculeati Vicherek 1973 et du Cypero-Spergularion marinae Slavnić 1948 ; d’ailleurs le contact vivace est un pré à Aeluropus littoralis (tableau 12 : colonne 5). La colonne 8 est le Cypero pygmaei-Glinetum lotoidis Mucina in Bergmeier & Raus 1999. Reliée à une pelouse vivace à Isoetes histrix (Ophioglosso lusitanici-Isoetion histricis B. Foucault 1988 fragmentaire ; O. lusitanicum est bien cité dans les colonnes 16 et 18 du tableau 5 des auteurs), la colonne 9 offre une association originale du Cicendio filiformis-Solenopsion laurentiae Brullo & Minissale 1998, difficile à valider. Avec Crassula vaillantii, la colonne 11 évoque le Crassulo vaillantii-Lythrion borysthenici B. Foucault 1988. La colonne 12 enfin semble représenter une association des Saginetea maritimae ou peut-être plutôt des Tuberarietea guttatae ou des Stipellulo capensis-Brachypodietea distachyae.


11. La végétation de pelouses xéro-acidiphiles thérophytiques
Dans une synthèse récente (de Foucault, 2024a), on a passé en revue la plupart des alliances des Tuberarietea guttatae, mais on doit encore y revenir pour la compléter.
Il n’y a rien à ajouter à l’Hispidello hispanicae-Festucion delicatulae B. Foucault 1999. Le Crassulo tillaeae-Sedion caespitosi B. Foucault 1999 n’est pas tout à fait correct car il correspond à un nouveau statut attribué au Sedenion caespitosi Rivas Mart. 1978 (Colloq. Phytosoc. VI : 59) et doit donc être dénommé Sedion cespitosi (Rivas Mart. 1978) B. Foucault all. nov. et stat. nov. hoc loco, typus nominis hoc loco : le Sedo caespitosi-Crassuletum tillaeae Rivas Goday 1957 (Anales Inst. Bot. Cavanilles 15 : 582).
Il n’y a pas non plus à revenir sur le Festucion ligusticae Aubert & R.J. Loisel, bien qu’on aimerait comprendre sa différenciation synécologique relativement au Tuberarion guttatae Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940. Depuis 1999, il faut ajouter à la liste des associations de ce dernier l’Alysso alyssoidis-Tuberarietum guttatae B. Foucault 2023 (de Foucault, 2023a), le Tolpido umbellatae-Micropyretum tenelli B. Foucault 2023 (de Foucault, 2023b) et, du Maroc, les Lysimachio parviflorae-Tuberarietum guttatae Titolet & B. Foucault in B. Foucault 2022 et Anthoxantho ovati-Trifolietum tomentosi Titolet & B. Foucault in B. Foucault 2022 (de Foucault, 2022c), les Crepido bellidifoliae-Plantaginetum weldenii Paradis et al. 2014 et Stipetum capensis Paradis et al. 2014 (Paradis et al., 2014). Dans cette même alliance, Roselló (1994, tableau 49) décrit le Cerastio taurici-Myosotidetum ramosissimae ; avec Geranium purpureum, Cardamine hirsuta, Valeriana calcitrapae… et, selon le commentaire associé évoquant son écologie de biotopes sombres et frais, cette association paraît cependant plus à sa place dans les Cardaminetea hirsutae.
On ne revient pas non plus sur l’Hedypnoido arenariae-Anthoxanthion ovati B. Foucault 2024, marocain, et le Tuberario guttatae-Airion praecocis B. Foucault 1999, de transition entre Tuberarion guttatae et Thero-Airion, en y ajoutant les Sclerantho annui-Tuberarietum guttatae (P. Allorge 1922) B. Foucault 2021 (de Foucault, 2021d), Miboro minimae-Tuberarietum guttatae B. Foucault 2021 (de Foucault, 2021e), Airo praecocis-Tuberarietum guttatae B. Foucault 2023 (de Foucault, 2023a), Ornithopodetum compresso-pinnati Menand, F. Kessler & Corriol in Corriol et al. (2022), Ornithopodo compressi-Tuberarietum guttatae Laigneau, Menand, F. Kessler & Corriol in Corriol et al. (2022) (Corriol et al., 2022) et Ornithopodetum perpusillo-compressi Lafon et al. 2025 (Lafon et al., 2025).
Le Vulpio-Lotion angustissimi centre-méditerranéen avait été ignoré dans la synthèse de 1999. Il faut dire que les données sur cette alliance surtout adriatique sont difficiles d’accès. D’abord son autorité hésite entre Vulpio-Lotion angustissimi Horvatić 1960 et Horvatić 1963 selon les publications. Cette alliance apparaît dans Horvatić (1962, Ljetopis JAZU 66 : 307), mais comme nomen nudum. C’est en 1963 qu’apparaît à nouveau ce nom avec un seul syntaxon validement publié, le Chrysopogono grylli-Airetum capillaris Horvatić 1963 (Hrvatska akademija znanosti i umjetnosti 4 : 96), les Ornithopodo compressi-Vulpietum Horvatić 1963, Gastridio ventricosi-Brachypodietum ramosi Horvatić 1963 et Psiluro-Trifolietum cherleri Horvatić 1963 étant des nom. nud. (les deux premiers) ou prov. (le troisième). On peut considérer que l’Ornithopodo compressi-Vulpietum Horvatić 1963 a finalement été validé dans Horvatić (1971, Simpozij o zaštiti prirode u našem kršu : 126) et doit donc être référencé comme Ornithopodo compressi-Vulpietum Horvatić 1963 ex 1971(la présence de plusieurs taxons du genre Vulpia, maintenant inclus dans Festuca, ne permet pas de préciser le nom complet). Le tableau 13 synthétise douze syntaxons se répartissant en deux groupes. Les colonnes 1 à 10, de Croatie, correspondent bien à cette alliance :
- en 1 et 2, l’Ornithopodo compressi–Festucetum Horvatić 1963 ex 1971 mut. hoc loco, 1 selon Horvatić (1971), 2 selon Hećimović (1984) ; le relevé in Horvatić (1971, Simpozij o zaštiti prirode u našem kršu: 126) en est tout naturellement le lectotypus nominis hoc loco ;
- en 3 et 4, la composante thérophytique du « Gastridio ventricosi-Brachypodietum ramosi Horvatić 1962 », originale mais qu’on ne peut valider faute d’en posséder des relevés précis ;
- en 5, un syntaxon qu’Hećimović (1984) rattache aussi au « Gastridio ventricosi-Brachypodietum ramosi Horvatić 1962 », mais bien distinct du précédent et qu’on propose d’isoler comme Galio divaricati-Phleetum echinati ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 9 du tableau 4 in Hećimović (1984, Acta Bot. Croatica 43) ;
- en 6 et 7, deux composantes thérophytiques du « Trifolio-Brachypodietum rupestris Hodak-Horvatić 1975 », deux colonnes pourtant difficiles à placer dans la même association ; quoi qu’il en soit, on peut au moins interpréter la colonne 7 comme Loto ornithopodioidis-Trifolietum echinati nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 8 du tableau 1 in Hodak-Horvatić (1975, Problems of Balkan flora and vegetation : 361-363) ;
- en 8 à 10, trois composantes thérophytiques du « Chrysopogono-Airetum capillaris Horvatić 1963 » ; la colonne 8 pourrait être redéfinie comme Loto angustissimi–Airetum elegantis mais les données sources sont synthétiques, empêchant toute validation ;
- la colonne 11 est le Psiluro-Trifolietum cherleri défini provisoirement par Horvatić (1963) sur la base d’un unique relevé.
Les colonnes 12 et 13, de Macédoine, paraissent se rattacher à une autre unité, notamment caractérisée par Petrorhagia dubia ; ce sont
- en 12, le Junco bufonii-Vulpietum ligusticae Stešević et al. 2025,
- en 13, le Tuberario guttatae-Avellinietum festucoidis Stešević et al. 2025,
ces deux associations rangées dans le Vulpio-Lotion par leurs auteurs. Le tableau 13 montre bien que les colonnes 1 à 10 se rangent correctement dans le Vulpio-Lotion angustissimi Horvatić 1963 muté en Festuco-Lotion angustissimi nom. mut. hoc loco, bien différencié des unités ouest-méditerranéennes comme le Festucion ligusticae et le Tuberarion guttatae, qu’on propose de néotypifier ici par l’Ornithopodo compressi–Festucetum Horvatić 1963 ex 1971 (Simpozij o zaštiti prirode u našem kršu : 126, sub Ornithopodo-Vulpietum). On ne peut y rattacher les deux associations de Macédoine. L’examen du tableau 13 montre encore que cette végétation centre-méditerranéenne accueille plus souvent qu’en Méditerranée occidentale des taxons habituellement considérés comme caractérisant les Stipellulo capensis-Brachypodietea distachyi, tels que Brachypodium distachyon, Linum strictum, Petrorhagia prolifera, Trifolium scabrum, Medicago minima, Catapodium rigidum.
Il est possible par ailleurs que de telles unités atteignent des régions plus orientales. Ainsi, en Crète (de Foucault, 1993 : 93) il existe des pelouses acidiphiles à Rumex bucephalophorus, Silene gallica, Tuberaria guttata, Tolpis umbellata, Aira elegans, Teesdalea coronopifolia, Anthoxanthum gracile…
Sans rien ajouter au Thero-Airion, on peut enfin passer au Sedion annui B. Foucault 2019, l’alliance la plus récemment définie et où l’on ne connaît guère que le Sclerantho annui-Sedetum annui B. Foucault, Corriol & F. Kessler in B. Foucault 2019 et l’Airo caryophylleae-Veronicetum brevistylae B. Foucault 2021 (de Foucault, 2019b, 2021f). On a aussi proposé d’y rattacher le Violo parvulae-Malcolmietum bicoloris de haute montagne grecque (de Foucault, 2022b), ne serait-ce qu’en considérant Viola parvula comme une bonne caractéristique de l’alliance ; bien que Sedum annuum n’y apparaisse pas, ce taxon est présent dans ces montagnes.
12. Une végétation de dune bulgare
De la côte bulgare de la mer Noire, Tzonev et al. (2005, tableau 2) décrivent une végétation dunaire biologiquement complexe, le « Xanthio italici-Leymetum sabulosi » que l’on peut séparer en
- une végétation vivace à Leymus racemosus et Eryngium maritimum à rattacher au Leymion racemosi Morariu 1957 (de Foucault, 2021c) ;
- une végétation thérophytique à Xanthium orientale, Salsola tragus (= ruthenica), Euphorbia peplis, Cakile maritima subsp. euxina se rattachant bien au Cakilo euxinae-Salsoletum ruthenicae Vicherek 1971 décrit aussi dans le tableau 1 des auteurs bulgares.
On voit ainsi comment la prise en compte des types biologiques peut s’avérer un « garde-fou » contre la description d’unités syntaxonomiques artificielles.
Il en est d’ailleurs un peu de même du « Xanthio italici-Ammophiletum arundinaceae Pignatti 1952 » du littoral italien (voir par exemple Brullo et al., 2001, tableau II).
13. Des pelouses des Charybdido pancratii-Asphodeletea ramosi
Dans une publication récente, on a attiré l’attention des phytosociologues sur la classe surtout géophytique des Charybdido pancratii-Asphodeletea ramosi (de Foucault, 2024b). Une lecture ultérieure a révélé un nouveau syntaxon de Corse, malheureusement connu d’un unique relevé (Paradis & Piazza, 1990 : 98), qui accueille Allium chamaemoly (photo 8), Acis rosea, Spiranthes spiralis, Serapias lingua, Romulea rollii (photo 9), Muscari comosum, Aetheorhiza bulbosa, Bellis sylvestris, Urospermum dalechampii, bien distinct du Romuleo columnae-Allietum chamaemolyos Molin. 1953 du Var et du Cap Corse et dont l’étude est à poursuivre. Ce relevé a été réalisé à Minaccia, sur la côte occidentale de l’île, mais en consultant les listes floristiques de Médail et al. (2017) puis celles de 141 îles et îlots de Corse (Paradis et al., 2021), deux de ceux-ci (site 10 : San Cyprianu, au sud-est ; site 42 : Cavallu, à l’extrême sud) hébergent probablement un groupement fort proche, avec A. chamaemoly, Acis rosea, Arisarum vulgare, Muscari comosum, Romulea requienii, Asphodelus ramosus, Charybdis undulata, et Moraea sisyrinchium à San Cyprianu.
De leur côté, Pozzo di Borgo et al. (2002, tableau 12) publient huit relevés d’une pelouse à Moraea sisyrinchium, Romulea cf. requienii, R. ramiflora, Asphodelus ramosus relevant de la même classe, mais non définie comme association, l’incertitude taxonomique régnant sur le premier Romulea cité empêchant d’aller plus loin.
Le tableau 8 publié par Paradis et al. (2006) décrit sans interprétation finale une pelouse à Thapsia meoides, Asphodelus ramosus, Leontodon tuberosus, Romulea columnae, Serapias lingua, plus rarement Arisarum vulgare et Ambrosina bassii.
Enfin, la composante cryptophyto-chaméphytique du Rostrario pubescentis-Tuberarietum villosissimae décrit de Sicile par Minissale et Sciandrello (2015, tableau 3) est caractérisée par cette liste (sur 20 relevés) : Alkanna tinctoria V, Paronychia argentea V, Moraea sisyrinchium IV, Echinophora tenuifolia IV, Romulea columnae *c. III, Ornithogalum gussonei III, Stachys arenaria III, Teucrium luteum III, Muscari comosum II, Cachrys sicula II, Stipa letourneuxii *l. II et des taxons de présence I et + dont Gagea granatelli (I), décrivant le nouveau Echinophoro tenuifoliae-Moraeetum sisyrinchii ass. nov. hoc loco, typus nominis hoc loco : composante cryptophyto-chaméphytique du relevé 10 du tableau 3 in Minissale & Sciandrello (2015, Acta Bot. Gallica: Botany Lett. 162 (1) : 70-72).


Bibliographie
Amigo J., Guitian J. & Fernández Prieto F., 1987. Datos sobre los bosques ribereños de aliso (Alnus glutinosa) cántabro-atlánticos ibéricos. V Jornadas de fitosociologia, Vegetacion de riberas de agua dulce, Universidad de la Laguna, Ser. Informes 22 : 159-176.
Baudière A., 1970. Recherches phytogéographiques sur la bordure méridionale du Massif central français (les monts de l’Espinouze) ; le climat et les formations forestières. Thèse de doctorat d’État, université de Montpellier, 317 p.
Baudière A., 1972a. Contribution à l’étude de la végétation des landes des monts de l’Espinouze (Cévennes occidentales). Bulletin de la Société botanique de France 119, 92e session extraordinaire dans le Languedoc : 65-109.
Baudière A., 1972b. La forêt de chênes verts dans les gorges d’Héric et sa signification biogéographique sur les limites de son aire. Bulletin de la Société botanique de France 119, 92e session extraordinaire dans le Languedoc : 19-64.
Baudière A., 1972c. Les groupements à Saxifraga pedemontana All. subsp. prostii (Sternb.) D.A. Webb du massif du Caroux. Bulletin de la Société botanique de France 119, 92e session extraordinaire dans le Languedoc : 111-114.
Baudière A., 1975a. Les hêtraies acidiphiles des monts de l’Espinouze (Cévennes occidentales) ; composition floristique et relations avec les chênaies caducifoliées. Colloques phytosociologiques III, La végétation des forêts caducifoliées acidiphiles : 1-15.
Baudière A., 1975b. Les callunaies des monts de l’Espinouze (Cévennes occidentales) et leurs relations avec les groupements sylvatiques. Colloques phytosociologiques II, La végétation des landes d’Europe occidentale : 1-124.
Bergmeier E. & Raus Th., 1999. Verbreitung und Einnischung von Arten der Isoëto-Nanojuncetea in Griechenland. Mitteilungen des badischen Landesvereins für Naturkunde und Naturschutz 17 (2) : 463-479.
Berisha N., Millaku F., Gashi B. & Matevski V., 2019. Ramondo-Ostryetum carpinifoliae, a new association from the hop-hornbeam forests of the Sharri Mountains, Kosovo. Hacquetia 18 (2) : 323-336.
Birač V., 1973 Vegetacija srđa i okolice Dubrovačke rijeke. Acta Botanica Croatica 32 : 135-170 (en croate).
Brullo S., Guisso del Galdo G.P., Siracusa G. & Spampinato G., 2001. Considerazioni fitogeografiche sulla vegetazione psammofila del litorali italiani. Biogeographia XXII : 93-137.
Corbetta F., Pirone G., Frattaroli A.R. & Ciaschetti G., 2004. Lineamenti vegetazionali del Parco nazional del Cilento e Vallo di Diano. Braun-Blanquetia 36 : 1-61.
Corriol G., Laigneau F., Menand M. & Kessler F., 2022. Étude phytosociologique de quelques végétations vasculaires thérophytiques, silicicoles, xérophiles du sud-ouest de la France. Carnets botaniques 96 : 1-7, https://doi.org/10.34971/tdgp-8335.
Čušin B., 2002. Pionirski gozdovi belega gabra (Carici albae-Carpinetum betuli ass. nova) na holocenskih terasah Nadiže. Hacquetia 1 (1) : 91-107.
Durán Gómez J.A., 2023. Paisaje vegetal del antiguo valle de Alfoz de Lloredo y su entorno (Cantabria), de Cóbreces a La Rabia, Cabezón de la Sal, Santillana del Mar y Suances. Jolube Consultor Botánico y Editor, Jaca (Huesca), 626 p.
Fernández Prieto J.A.,Fernández Ordóñez M.C. & Collado Prieto M.A., 1987. Datos sobre la vegetación de las « turberas de esfagnos » galaico-asturianas y orocantábricas. Lazaroa 7 : 443-471.
Foucault B. (de), 1993. II – Données phytosociologiques. In B. de Foucault & J.-P. Cornu, Itinéraire botanique d’un groupe de Linnéens en Crète, 29 avril-13 mai 1991 (2e partie). Bulletin de la Société linnéenne Nord-Picardie 11 : 85-105.
Foucault B. (de), 1999. Nouvelle contribution à une synsystématique des pelouses sèches à thérophytes. Documents phytosociologiques, n. s., XIX : 47-105.
Foucault B. (de), 2017. Données phytosociologiques nouvelles sur la végétation des Cévennes occidentales (département de la Lozère) et du Haut-Languedoc (département de l’Hérault). Bulletin de la Société botanique du Centre-Ouest, n. s., 48 : 334-359.
Foucault B. (de), 2019a. Contribution à la connaissance phytosociologique de la végétation du pays de Sault (Aude, France). Evaxiana 6 : 29-129.
Foucault B. (de), 2019b. Données phytosociologiques collectées lors des sorties du groupe botanique de la SESA durant l’année 2018. Bull. Soc. Études Sci. Aude CXVIII : 176-186
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021a. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2021b. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de), 2021c. Contribution à une synthèse des classes des Honckenyo peploidis-Leymetea arenarii et des Euphorbio paraliae-Ammophiletea arundinaceae. Carnets botaniques 64 : 1-22 + 16 tableaux, https://doi.org/10.34971/4ff3-vd67.
Foucault B. (de), 2021d. Miscellanées phytosociologiques : quelques syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 44 : 1-23 + 28 tableaux, https://doi.org/10.34971/AKGG-T733.
Foucault B. (de), 2021e. Complément à l’étude phytosociologique du littoral sableux du Languedoc-Roussillon : les dunes de Sainte-Marie-la-Mer à Argelès-sur-Mer ; comparaison avec un système arrière-dunaire atlantique. Carnets botaniques 32 : 1-13 + 6 tableaux, https://doi.org/10.34971/pw55-r859.
Foucault B. (de), 2021f. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2022a. Validation de syntaxons invalidement publiés en 2021. Carnets botaniques 79 : 1-12, https://doi.org/10.34971/3XK3-N640.
Foucault B. (de), 2022b. Nouvelles considérations phytosociologiques sur la végétation de la Grèce. Carnets botaniques 109 : 1-27 + 7 tableaux, https://doi.org/10.34971/Q6KD-W822.
Foucault B. (de), 2022c. Compléments aux synthèses des Juncetea bufonii et Crypsietea aculeatae ; essai sur les Oryzetea sativae. Carnets botaniques 107 : 1-27 + 5 tableaux, https://doi.org/10.34971/vgyx-k502.
Foucault B. (de), 2023a. Miscellanées phytosociologiques IV : syntaxons nouveaux, validés ou complétés. Carnets botaniques 127 : 1-18 + 14 tableaux, https://doi.org/10.34971/1d8e-ky97.
Foucault B. (de), 2023b. Nouvelles données sur la végétation du versant audois de la Montagne Noire. Carnets botaniques 160 : 1-30 + 21 tableaux, https://doi.org/10.34971/HTCT-WD37.
Foucault B. (de), 2024a. Nouvelles interprétations phytosociologiques des subéraies marocaines. Carnets botaniques 228 : 1-8, https://doi.org/10.34971/9BFQ-G158.
Foucault B. (de), 2024b. La classe des Charybdido pancratii-Asphodeletea ramosi en Méditerranée occidentale. Carnets botaniques 226 : 1-8 + 2 tableaux, https://doi.org/10.34971/WW5N-7K72.
Foucault B. (de), 2025. Miscellanées phytosociologiques VIII : syntaxons nouveaux, validés ou complétés. Carnets botaniques 264 : 1-13 + 16 tableaux, https://doi.org/10.34971/AVAA-5634.
Foucault B. (de), 2026. Nouvelles interprétations phytosociologiques de la végétation forestière sud-est-européenne. Carnets botaniques 290 : 1-15 + 11 tableaux, https://doi.org/10.34971/DHCY-4609.
Foucault B. (de) & Royer J.-M., 2014. Contribution au prodrome des végétations de France : les Franguletea alni Doing ex V. Westh. in V. Westh. & den Held 1969. Journal de botanique de la Société botanique de France 66 : 83-106.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Glemarec E., Delassus L. & Boullet V., 2025. Contribution au prodrome des végétations de France : les Calluno vulgaris-Ulicetea minoris Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, série 4, 5 : 1-163.
Jelinčić A., Vukelić J. & Dora Papković D., 2024. Phytosociological research into the Illyrian oak-hornbeam forest on the limestone cliffs of Kupa river canyon (Kamanje, west-central Croatia). Šumarski list (1-2) : 7-18.
Hećimović M., 1984. Grassland vegetation of the island of Sipan. Acta Botanica Croatica 43 : 161-166.
Hodak-Horvatić N., 1975. Trifolio-Brachypodietum rupestris, eine neue Trockenrasen-Gesellschaft der kroatischen Küstenlandes. In Problems of Balkan flora and vegetation, Sofia : 360-364.
Horvat I., 1936. Istraživanje vegetacije planina Vardarske banovine II. Ljetopisa JAZU 48 : 211-227 (en croate).
Horvat I., Glavač V. & Ellenberg H., 1974. Vegetation Südosteuropas. Geobotanica Selecta IV : 1-767.
Horvatić S., 1962. Prilozi poznavanju vegetacije Južnohrvatskog primorja. Ljetopis JAZU 66 : 302-308 (en croate).
Horvatić S., 1963. Vegetacijska karta otoka Paga s općim pregledom vegetacijskih jedinica Hrvatskog primorja. Acta biologica / Hrvatska akademija znanosti i umjetnosti (Academia scientiarum et artium croatica) 4 : 5-181 (en croate).
Horvatić S., 1971. Osnovne vegetacijske jedinice primorskog krša i pitanje njihove pojačane zaštite. In Simpozij o zaštiti prirode u našem kršu : 109-135 (en croate).
Hruska K., 1988. I castagneti dei Monti della Laga (Italia Centrale). Braun-Blanquetia 2 : 117-125.
Lafon P., Mady M., Dufay J. & Guisier R., 2025. Les ornithopodaies et les pelouses silicicoles thérophytiques acidiphiles à trèfles nains du sud-ouest de la France (Nouvelle Aquitaine). Carnets botaniques 250 : 1-14, https://doi.org/10.34971/3RBY-KP27.
Loidi J., 1983. Estudio de la flora y vegetación de las cuencas de los rios Deva y Urola en la provincia de Guipúzcoa. Thèse, Universidad Complutense de Madrid, 298 p.
Médail F., Petit Y., Delage A., Paradis G. & Hugot L., 2017. La flore vasculaire de l’île de Cavallo (archipel des Lavezzi, Corse) : diversité, intérêt biogéographique et conservation. Ecologia Mediterranea 43 (2) : 103-158.
Minissale P. & Sciandrello S., 2015. The sabulicolous therophytic associations in Sicily: new insights through the statistical approach, stressing the continuum versus discrete model of plant communities. Acta Botanica Gallica: Botany Letters 162 (1) : 55-78.
Molero J., 1984. Contribució al coneixement fitocenològic dels Catalànids centrals (Serra de Pradesi Montsant): comunitats noves o poc conegudes. Butlletí de la Institució Catalana d’Història Natural 51, Bot. : 139-160.
Nordmann V. (rédacteur), 1928. Aperçu de la géologie du Danemark. Danmarks geologiske Undersøgelse 4 : 9-232.
Paradis G. & Piazza C., 1990. Étude de la végétation d’un site littoral sableux en voie de dégradation rapide à proximité d’Ajaccio (Corse) : le fond de l’anse de Minaccia. Bulletin de la Société botanique du Centre-Ouest, n. s., 21 : 75-112.
Paradis G., Piazza C. & Pozzo di Borgo M.-L., 2006. Description phytosociologique et cartographique de la végétation des pourtours rocheux du golfe de la Rondinara (Corse). Bulletin de la Société botanique du Centre-Ouest, n. s., 37 : 65-115.
Paradis G., Panaiotis Ch. & Piazza C., 2014. Contribution à la connaissance de la végétation thérophytique du littoral rocheux de la Corse (Saginetea maritimae, Tuberarietea guttatae, Sisymbrietea officinalis). Documents phytosociologiques, série 3, 1 : 352-391.
Paradis G., Piazza C. & Mori Ch., 2021. Les îles et îlots satellites de la Corse : état des connaissances en 2021 et enjeux de conservation. Evaxiana 8 : 69-192.
Pozzo di Borgo M.-L., Ravetto S., Lorenzoni C. & Paradis G., 2002. Description phytosociologique et cartographique de la végétation du site d’Arasu (sud-est de la Corse) proposé pour le réseau Natura 2000. Travaux scientifiques du Parc naturel régional de Corse et des réserves naturelles 60 : 3-51.
Rivas Goday S., 1957. Nuevos órdenes y alianzas de Helianthemetea annua Br.-Bl. Anales del Instituto Botanico A.J. Cavanilles 15 : 539-651
Rivas-Martínez S., 1978. Sur la syntaxonomie des pelouses thérophytiques de l’Europe occidentale. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 55-69.
Roselló Gimeno R., 1994. Catalogo floristico y vegetación de la comarca natural del Alto Mijares (Castellon). Diputacio de Castello, 650 p.
Sissingh G., 1970. Dänische Büchenwalder. Vegetatio 21 (4-6) : 245-254.
Stešević D., Stanišić-Vujačić M., Milanović Đ & Šilc U., 2025. Retrodunal dry grassland vegetation in the hinterland of Velika Plaža (Montenegro). Acta Botanica Croatica 84 (2) : 219-236.
Stevanović V. & Stevanović B., 1985. Asplenio cuneifolii Ramondaetum nathaliae – new chasmophytic community on serpentine rocks in Macedonia. Bulletin of the Natural History Museum in Belgrade, B 40 : 75-87.
Stevanović V., Jakovljević K. & Matevski V., 2014. Chasmophytic communities of endemic and relict species Ramonda nathaliae Pančić & Petrović on ophiolithic substrate in Republic of Macedonia. Botanica Serbica 38 (1) : 81-90.
Tammaro F. & Poldini L., 1988. La vegetazione della lecceta litoreana di Torino di Sangro (Chieti), nel medio versante adriatico italiano. Braun-Blanquetia 2 : 127-132.
Tzonev R., Dimitrov M. & Roussakova V., 2005. Dune vegetation of the Bulgarian Black Sea coast. Hacquetia 4 (1) : 7-32.
Tzonev R., Dimitrov M., Gussev C., Vulchev V. & Nikolov I., 2018. Classification of the relict forest communities of Palla’s Black Pine (Pinus nigra subsp. pallasiana) in Bulgaria. Hacquetia 17 (2) : 125-161.
Vanden Berghen C., 1968. Les forêts de la haute Soule (Basses-Pyrénées). Bulletin de la Société royale de botanique de Belgique 102 (1) : 107-132.
Remerciements
Sans l’aide efficace et réactive de Nenad Jasprica (phytosociologue, Dubrovnik), qui m’a envoyé plusieurs scans de publications, notamment de S. Horvatić (photo ci-contre), je n’aurai pas pu éclaircir le problème du Festuco-Lotion angustissimi croate ; par ailleurs Franck Le Driant m’a autorisé à puiser dans sa base photographique FloreAlpes (https://www.florealpes.com) ; qu’ils en soient tous les deux vivement remerciés.



