
La classe des Charybdido pancratii-Asphodeletea ramosi en Méditerranée occidentale
Title
The Charybdido pancratii-Asphodeletea ramosi class in the western Mediterranean
Résumé
Initialement définie en Méditerranée centrale, la classe des Charybdido pancratii-Asphodeletea ramosi, caractérisée par des géophytes monocotylédones entomogames, peut être étendue à la Méditerranée occidentale par les Colchico filifolii-Moraeion sisyrinchii (péninsule Ibérique et France méditerranéenne) et Colchico graminei-Moraeion sisyrinchii (Maroc).
Abstract
Initially defined in the central Mediterranean, the Charybdido pancratii-Asphodeletea ramosi class, characterized by monocotyledonous entomogamous geophytes, can be extended to the western Mediterranean by Colchico filifolii-Moraeion sisyrinchii (Iberian Peninsula and Mediterranean France) and Colchico graminei-Moraeion sisyrinchii (Morocco).
Parmi les classes de végétation de pelouses vivaces définies assez récemment, celle des Charybdido pancratii-Asphodeletea ramosi a été créée en Italie par Biondi et al. (2016, 2017), donc plutôt en Méditerranée centrale. Elle est caractérisée surtout par des géophytes entomogames relevant de diverses familles monocotylédones (Amaryllidaceae, Araceae, Asparagaceae, Asphodelaceae, Colchicaceae, Iridaceae). Longtemps et encore récemment, de par ses contacts et ses intrications avec des pelouses thérophytiques, souvent très riches, ce type de végétation a été inclus dans les classes dédiées à de telles pelouses (surtout Stipo capensis-Trachynietea distachyae mutés en Stipellulo capensis-Brachypodietea distachyae, parfois les Tuberarietea guttatae).
Pour en avoir une vue d’ensemble, a été élaboré le tableau synthétique 1 à partir du tableau 7 de Biondi et al. (2017), avec quelques modifications (présence, nomenclature selon POWO). En particulier la colonne 9 des auteurs, correspondant au Verbasco garganici-Asphodeletum fistulosi Biondi et al. 2016, unique association connue de l’Asphodelion fistulosi Biondi et al. 2016, comporte une composante thérophytique non négligeable (Asphodelus fistulosus, Dasypyrum villosum, éventuellement Euphorbia terracina) et des liens synfloristiques faibles ou nuls avec le reste de la classe (notamment absence des géophytes caractéristiques) et n’est pas retenue ici. Il faut bien reconnaître que les associations décrites par les phytosociologues italiens sont statistiquement assez faiblement caractérisées : sur les seize syntaxons du tableau 7 des auteurs (syntaxons 1 à 17, moins le n° 9), la moyenne du nombre de relevés par syntaxon est de seulement 3,8.
Les auteurs italiens subdivisent leur classe en deux ordres :
- les Asphodeletalia ramosi Biondi in Biondi et al. 2016, avec trois alliances (hors Asphodelion fistulosi), le Charybdido pancratii-Asphodelion ramosi Biondi et al. 2016 (tableau 1 : colonnes 1 à 5), l’Asphodelo ramosi-Ferulion communis Biondi et al. 2016 (tableau 1 : colonnes 6 et 7) et l’Asphodelo ramosi-Ferulion glaucae Biondi et al. 2016 (tableau 1 : colonne 8) ;
- les Bellido sylvestris-Arisaretalia vulgaris Biondi in Biondi et al. 2017, avec quatre alliances, le Leontodonto tuberosi-Bellidion sylvestris Biondi et al. 2001 (tableau 1 : colonnes 10 à 12), le Cyclamini hederifolii-Ranunculion bullati Biondi et al. 2017 (tableau 1 : colonnes 13 et 14), le Cyclamini hederifolii-Arisarion vulgaris Biondi et al. 2017 (tableau 1 : colonnes 15 et 16) et le Ranunculion bullati Pérez Latorre & Cabezudo in Pérez Latorre et al. 2008 (tableau 1 : colonne 17).
Il faudra sans doute aussi intégrer à ces unités les Ophryo praecocis-Dactylidetum hispanicae, Anacamptido longicornus-Dactylidetum hispanicae et Anacamptido collinae-Dactylidetum hispanicae décrits de Sardaigne nord-occidentale par Farris et al. (2007, respectivement sub Ophrydo praecocis-Dactyletum hispanicae, Orchido longicorni-Dactyletum hispanicae et Orchido collinae-Dactyletum hispanicae nom mut. et corr.). La présence significative dans ces pelouses sardes de taxons comme Scabiosa atropurpurea, Foeniculum vulgare, Oloptum miliaceum, Dittrichia viscosa, Cichorium intybus indique qu’elles sont soumises à des pressions biotiques.
En dehors de l’Italie et du Ranunculion bullati décrit du sud de l’Espagne (Pérez Latorre et al., 2008), cette classe n’est guère connue, bien que des associations pouvant s’y rattacher sont décrites depuis longtemps. Ainsi, les colonnes 18 à 29 du tableau 1 synthétisent les données suivantes (en respectant éventuellement l’écriture des noms des syntaxons donnée par les auteurs, qui pourra être corrigée par la suite) :
- 18 – de Bolòs et al. (1970, tableau 14, ‘Allietum chamaemoly’)
- 19 – Molinier (1959, tableau VI, ‘Allietum chamaemolyi’)
- 20 – Molinier (1953a, 2 relevés p. 68 ; 1953b, 1 relevé p. 29)
- 21 – Gesti Perich (2006, tableau 39, ‘Allietum chamaemolly’)
- 22 – de Bolòs & Molinier (1958, tableau 12, ‘Allietum chamaemoly’)
- 23 – de Bolòs & Molinier (1958, tableau 11, ‘Irideto-Stipetum retortae’)
- 24 – de Bolòs (1996, p. 116, ‘Irideto-Stipetum retortae’)
- 25 – Fernandez Casas (1970, p. 282, ‘Scillo-Narcissetum serotinae’)
- 26 – Brullo (1985, tableau 9, ‘Allietum lojaconoi’)
- 27 – Paradis & Pozzo di Borgo (1999, tableau 6)
- 28 – Braun-Blanquet & Maire (1924, 1 relevé p. 39)
- 29 – Braun-Blanquet & Maire (1924, p. 53-54 : relevés 2 et 3).
Les colonnes 19 et 20, respectivement du Cap Corse et du Var, paraissent constituer deux races d’une même association, le Romuleo columnae-Allietum chamaemolyos Molin. 1953 (on a repris le nom initial donné par l’auteur, 1953a : 67, contemporain de l’Allietum chamaemolyos de Molinier, 1953b : 29), souvent abrégé en Allietum chamaemolyos par Molinier lui-même (sub Allietum chamaemoly). La colonne 18, de Minorque, est assez voisine de celles-ci, mais suffisamment différenciée pour être distinguée comme Colchico filifolii-Allietum chamaemolyos (O. Bolòs, Molin. & P. Monts. 1970) ass. nov. hoc loco [syn. : Allietum chamaemolyos O. Bolòs, Molin. & P. Monts. 1970 (Acta Geobot. Barcinon. 5 : 71) nom. illeg., non Molin. 1953], typus nominis hoc loco : composante vivace du relevé 3 du tableau 14 in de Bolòs et al. (1970, Acta Geobot. Barcinon. 5 : 72-73). Les colonnes 21 (de Catalogne) et 22 (de Majorque) ne peuvent se rattacher à l’association de Molinier et peuvent être distinguées respectivement comme
- Lobulario maritimae-Allietum chamaemolyos (Gesti Perich 2006) nov. hoc loco [syn. : Allietum chamaemolyos Gesti Perich 2006 (Arxius CXXXVIII : 583) nom. illeg., non Molin. 1953], typus nominis hoc loco: composante vivace du relevé 4 du tableau 39 in Gesti Perich (2006, Arxius CXXXVIII : 584-585) ;
- Romuleo assumptionis-Allietum chamaemolyos (O. Bolòs & Molin. 1958) nov. hoc loco [syn. : Allietum chamaemolyos O. Bolòs & Molin. 1958 (Collect. Bot. (Barcelona) V (III) : 787) nom. illeg., non Molin. 1953], typus nominis hoc loco: composante vivace du relevé 2 du tableau 12 in de Bolòs & Molinier (1958, Collect. Bot. (Barcelona) V (III) : 788-790).
La colonne 23 (Majorque) est définie sur seulement trois relevés, mais est bien distincte de la colonne 24 pourtant rattachée à celle-ci. Le ‘Scillo-Narcissetum serotinae’ peut être corrigé et muté ici en Prosperini autumnalis-Narcissetum serotini Fern. Casas 1970 nom. corr. et mut. (colonne 25). La colonne 26 (de Malte et Gozo) est correctement décrite sous le nom de Allietum lojaconoi Brullo 1985. Le syntaxon du plateau de Bonifacio (colonne 27) mérite d’être élevé au rang d’association : Moraeo sisyrinchii-Asphodeletum ramosi B. Foucault, Paradis & Pozzo di Borgo ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 4 du tableau 6 in Paradis & Pozzo di Borgo (1999, Bull. Soc. Bot. Centre-Ouest, n. s., 30 : 20) ; Gamisans (2001 : 126, 128) publie de son côté une liste d’une végétation à Moraea sisyrinchium, Kundmannia sicula, Brachypodium retusum, Gladiolus byzantinus, Ambrosina bassii, Romulea requienii, Drimia maritima présente aussi sur le plateau calcaire de Bonifacio, mais paraissant différente de la précédente ; une monographie phytosociologique de ce plateau calcaire corse serait la bienvenue…
L’ensemble formé par les colonnes 18 à 27 est bien différencié et paraît avoir la valeur d’une alliance originale caractérisée ou différenciée des alliances voisines essentiellement par Moraea sisyrinchium, Romulea columnae subsp. c., Colchicum filifolium, Brachypodium retusum, Allium chamaemoly, à chorologie plutôt ouest-méditerranéenne (péninsule Ibérique à France méditerranéenne), le Colchico filifolii-Moraeion sisyrinchii all. nov. hoc loco, typus nominis hoc loco : l’Allietum lojaconoi Brullo 1985 (Doc. Phytosoc., n. s., IX : 11). Deux groupes d’associations peuvent toutefois être distingués : l’un à Allium chamaemoly (colonnes 18 à 22), l’autre à Moraea sisyrinchium et Drimia maritima (colonnes 23 à 27). Quelques données apportées par Braun-Blanquet et Maire (1924 : 39 et 53-54 : relevés 2 et 3, respectivement repris en colonnes 28 et 29 du tableau 1), au nord de Rabat et à l’est de Fès, montrent la présence de cette alliance et de ce dernier groupe au Maroc méditerranéen. Les listes de Sauvage (1933-1934) du Zerhoun confirment cette affiliation avec des taxons comme Asphodelus ramosus, Arisarum simorrhinum subsp. subexertum, Drimia maritima, Hyacinthoides lingulata, Ranunculus bullatus, Allium chamaemoly, Prospero autumnale, Narcissus papyraceus, N. cantabricus, Erophaca baetica, Scilla peruviana. On peut encore intégrer cette alliance aux Bellido sylvestris-Arisaretalia vulgaris.
Sous l’effet d’une humidification édaphique, certaines de ces associations accueillent Isoetes duriei et Ophioglossum lusitanicum. C’est le cas notamment du Colchico filifolii-Allietum chamaemolyos (de Bolòs et al., 1970 : 76-77 ; Argagnon, 2023 : fiche 69-04, alors au sein de l’Allietum chamaemolyos).
Ajoutons que, en sous-bois d’un fourré à Pistacia lentiscus de Minorque, de Bolòs et al. (1970, tableau 3 : relevés 2 à 8) évoquent un syntaxon à Araceae (Arum pictum, Helicodiceros muscivorus, Arisarum vulgare) pouvant se rattacher encore à cette alliance, mais il est dépourvu de Moraea sisyrinchium et d’Allium chamaemoly, peut-être par suite de sa position intraforestière.
Plus au sud, par exemple toujours au Maroc (entre Casablanca et Marrakech, donc bien plus au sud que le Colchico filifolii-Moraeion sisyrinchii), on retrouve encore des pelouses xérophiles à géophytes souvent mélangées aux pelouses thérophytiques en mosaïque (Nègre, 1959, 1978 ; de Foucault & Lazare, 2020). Ce sont, avec la nomenclature de 1978 qui serait à améliorer
- « Rumiceto-Plantaginetum»
- « Plantagineto-Vagarietum»
- « Tuniceto-Spergularietum»
- « Rumiceto-Plantaginetum à Androcymbium gramineum» (Nègre, 1959 : 170)
- « Stipeto-Resedetum»
- « Androcymbio-Iridetum»
- « Notocero-Plantaginetum»
- « Androcymbio-Notoceretum»
- « Notocero-Astragaletum»
- « Aizoeto-Spergularietum»
- « Haloxyleto-Plantaginetum»
- « Artemisieto-Plantaginetum»
- « Lycieto-Plantaginetum»
- « Spergularieto-Hannonietum» (Nègre, 1959, tableau III)
- « Leysereto-Mesembryanthemetum»
- « Leysereto-Mesembryanthemetum»
- « Iflogeto-Aizoetum»
- « Haloxyleto-Matthioletum»
- « Notocero-Leyseretum»
- « Haloxyleto-Spergularietum»
- « Haloxyleto-Erucetum».
Le tableau 2 synthétise notamment ces syntaxons issus des tableaux 1 et 2 de Nègre (colonnes 1 à 20 ; le nombre de relevés synthétisés n’est pas indiqué par l’auteur, sauf indication contraire de données en 4 et 14 provenant de Nègre, 1959) où on remarque une présence significative de Moraea sisyrinchium, Biarum dispar (Araceae), Colchicum gramineum (= Androcymbium g., Colchicaceae), associés à Hannonia hesperidum (Amaryllidaceae). Certains de ces syntaxons sont recevables selon le paradigme phytosociologique actuel, notamment l’« Androcymbio-Iridetum » et le « Spergularieto-Hannonietum »pour les pelouses vivaces, qui sont donc par ailleurs valides et dont les noms peuvent être mutés respectivement en Colchico graminei-Moraeetum sisyrinchii Nègre 1959 et Spergulario fimbriatae-Hannonietum hesperidi Nègre 1959, lesquels ne peuvent toutefois être lectotypifiés car les données publiées sont synthétiques.
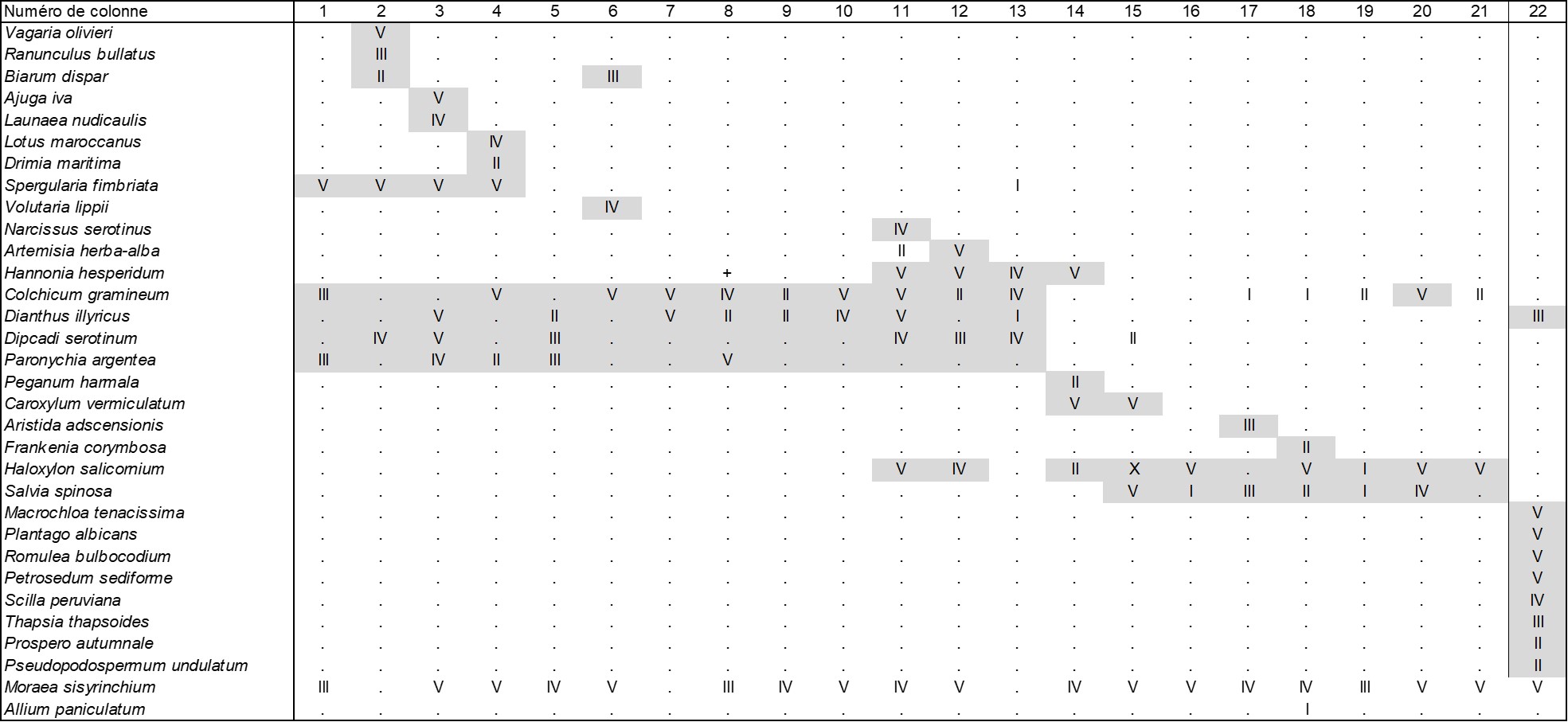
Notre tableau synthétique 2 montre que les colonnes 1 à 13 constituent une alliance originale caractérisée par Moraea sisyrinchium, Biarum dispar, Dipcadi serotinum, Colchicum gramineum, Hannonia hesperidum, Dianthus illyricus, le Colchico graminei-Moraeion sisyrinchii all. nov. hoc loco, typus nominis hoc loco : le Colchico graminei-Moraeetum sisyrinchii Nègre 1959 (Recherches phytogéographiques… : 179, sub Androcymbio-Iridetum). Cette alliance s’intègre toutefois assez mal à la classe considérée ici, seul Moraea sisyrinchium le reliant au Colchico filifolii-Moraeion sisyrinchii, à moins de la considérer comme une fin de classe vers le sud.
La colonne 22 de ce même tableau 2 rapporte une donnée d’Algérie (Wojterski & Beloued, 1992, tableau 2), synthèse d’un ensemble de six listes floristiques non quantifiées ordonnées le long d’un transect topographique, montrant des liens avec une steppe à Macrochloa tenacissima et d’autres réduits à Moraea sisyrinchium et Dianthus illyricus avec l’alliance marocaine. La réserve étudiée se situant vers la latitude
35° 12’ N, donc assez loin des plaines telliennes, cela n’exclut pas que le Colchico graminei-Moraeion sisyrinchii puisse être présent bien plus au nord.
En conclusion on peut considérer que les Charybdido-Asphodeletea initialement décrits de Méditerranée centrale peuvent s’étendre à la Méditerranée occidentale (péninsule Ibérique, France méridionale, une partie du Maroc). Cette végétation dominée par les géophytes monocotylédones zoogames apparaît comme une timide répétition dans l’hémisphère nord de la végétation pouvant être observée en Afrique du Sud sous un climat un peu comparable et caractérisée par des familles et genres tels que (entre parenthèses l’indication du nombre approximatif d’espèces pour juger de la richesse de cette flore, d’après Leistner, 2000 ; contour des genres revu selon Mabberley, 2008)
- Amaryllidaceae : Amaryllis (2), Ammocharis (3), Boophone (2), Brunsvigia (209), Crinum (20), Crossyne (2), Cybistetes (1), Cyrtanthus (50), Gethyllis (32), Haemanthus (22), Hessea (10), Nerine (23), Scadoxus (3), Strumaria (incl. Carpolyza; 24), Tulbaghia (20 ; photo 1) ;
- Asparagaceae (incl. Eriospermaceae) : Bowiea (2), Caesia (2), Chlorophytum (36), Daubenya (incl. Amphisiphon, Androsiphon, Neobakeria; 4), Dipcadi (14), Drimia (incl. Litanthus, Rhadamanthus, Schizobasis, Thuranthos, Urginea ; 44), Eriospermum (100), Eucomis (10), Lachenalia (incl. Polyxena ; 115), Ledebouria (incl. Drimiopsis ; 43), Massonia (incl. Whiteheadia ; 10), Ornithogalum (incl. Albuca, Galtonia, Neopatersonia, Tenicroa ; 142), Scilla (6), Veltheimia (2) ;
- Asphodelaceae : Astroloba (7), Bulbine (46 ; photo 2), Bulbinella (17), Chortolirion (1), Gasteria (8), Kniphofia (47 ; photo 3), Trachyandra (49) ;
- Colchicaceae : Baeometra (1), Camptorhiza (1), Colchicum (incl. Androcymbium, 28), Hexacyrtis (1), Ornithoglossum (8), Wurmbea (18) ;
- Haemodoraceae : Dilatris (4), Wachendorfia (4) ;
- Hypoxidaceae : Empodium (9), Hypoxis (45), Pauridia (2) ;
- Iridaceae : Aristea (45), Babiana (68), Bobartia (14), Chasmanthe (3), Crocosmia (9), Dierama (44), Dietes (5), Duthiastrum (1), Ferraria (11), Freesia (14), Geissorhiza (84), Gladiolus (165), Hesperantha (75), Ixia (50), Klattia (3), Lapeirousia (35), Melasphaerula (1), Micranthus (3), Moraea (incl. Homeria; 180), Nivenia (10), Pillansia (1), Romulea (77), Sparaxis (15), Syringodea (8), Thereianthus (7), Tritonia (28), Tritoniopsis (22), Watsonia (52), Witsenia (1), Xenoscapa (2) ;
- Lanariaceae : Lanaria (1) ;
- Tecophilaeaceae : Cyanella (7), Walleria (2),
avec donc de nouvelles familles (Haemodoraceae, Hypoxidaceae, Lanariaceae, Tecophilaeaceae) relativement à la végétation euroméditerranéenne ; notons encore la présence de genres en commun avec celle-ci : Colchicum, Dipcadi, Drimia, Gladiolus, Moraea, Ornithogalum, Romulea, Scilla.
Dans ces pelouses géophytiques sud-africaines, il existe aussi des pelouses thérophytiques associées, d’optimum vernal après les pluies d’hiver, caractérisées par des Crassula, des Grielum (Neuradaceae), des Brassicaceae (Heliophila, incl. Thlaspeocarpa), des Poaceae (Schmidtia, Tribolium), des Campanulaceae (Monopsis, Wahlenbergia), des Aizoaceae (Aethephyllum, Aizoon, Apatesia, Carpanthea, Cleretum, Dorotheanthus, Hymenogyne, Tetragonia), des Molluginaceae (Adenogramma, Glinus, Limeum, Mollugo, Pharnaceum) et surtout des Asteraceae (Amellus, Arctotheca, Arctotis, Cotula, Dimorphotheca, Felicia, Gazania, Gorteria, Gymnodiscus, Lasiospermum, Leysera, Oncosiphon, Osteospermum, Pentzia, Relhania, Rhynchopsidium, Senecio s. l., Ursinia), des Scrophulariaceae (Alonsoa, Cromidon, Diascia, Dischisma, Hebenstretia, Hemimeris, Jamesbrittenia, Lyperia, Manulea, Nemesia, Polycarena, Sutera, Zaluzianskya, incl. Reyemia).



Bibliographie
Argagnon O., 2023. Contribution au prodrome des végétations de France : les Stipo capensis – Trachynietea distachyae Brullo in Brullo, Scelsi & Spampinato 2001. Documents phytosociologiques, série 4, 3 : 3-32.
Biondi E., Pesaresi S., Galdenzi D., Gasparri R., Biscotti N., del Viscio G. & Casavecchia S., 2016. Post-abandonment dynamic on Mediterranean and sub-Mediterranean perennial grasslands: the edge of vegetation of the new class Charybdido pancratii-Asphodeletea ramosi. Plant Sociology 53 (2) : 3-18.
Biondi E., Pesaresi S., Gasparri R., Biscotti N., del Viscio G., Bonsanto D. & Casavecchia S., 2017. New contributions to the class Charybdido pancratii-Asphodeletea ramosi Biondi 2016. Plant Sociology 54 (1) : 137-144.
Bolòs O. (de), 1996. La vegetació de les Illes Balears, comunitats de plantes. Institut d’Estudis Catalans, Barcelone, 267 p.
Bolòs O. (de) & Molinier R., 1958. Observations phytosociologiques dans l’île de Majorque. Collectanea Botanica (Barcelona) V (III) : 699-865.
Bolòs O. (de), Molinier R. & Montserrat P., 1970. Observations phytosociologiques dans l’île de Minorque. Acta Geobotanica Barcinonensia 5 : 1-150.
Braun-Blanquet J. & Maire R., 1924. Études sur la végétation et la flore marocaines, 1re partie – Études sur la végétation du Maroc. Mémoires de la Société des sciences naturelles du Maroc 8 (1) : 5-152.
Brullo S., 1985. Sur la syntaxonomie des pelouses thérophytiques des territoires de l’Europe occidentale. Documents phytosociologiques, n. s., IX : 1-24.
Farris E., Secchi Z. & Filigheddu R., 2007. Caratterizzazione fitosociologica dell’habitat prioritario 6220-“Percorsi substeppici di graminacee e piante annue dei Thero-Brachypodietea”: caso di studio della Sardegna settentrionale. Fitosociologia 44 (2), suppl. 1 : 271-278.
Fernandez Casas J., 1970. Notas fitosociologicas breves, I. Ars Pharmaceutica XI : 273-298.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2023. Nouvelles données sur la végétation du versant audois de la Montagne Noire. Carnets botaniques 160 : 1-30, https://doi.org/10.34971/HTCT-WD37.
Foucault B. (de) & Lazare J.-J., 2020. Contribution à une synthèse phytosociologique des steppes d’Afrique du Nord (hors Lygeo sparti-Stipetea tenacissimae). Botanique 6 : 53-116.
Gamisans J., 2001. La végétation de la Corse. In D. Jeanmonod & H.-M. Burdet (eds), Compléments au prodrome de la flore corse, annexe 2 : 1-391.
Gesti Perich J., 2006. El poblament vegetal dels Aiguamolls de l’Empordà. Arxius de les seccions de ciències CXXXVIII : 1-856.
Leistner O.A., 2000. Seed plants of southern Africa: families and genera. Strelitzia 10 : 1-775.
Mabberley D.J., 2008. Mabberley’s plant-book, a portable dictionary of plants, their classification and uses. Cambridge University Press, 1 021 p.
Molinier Re., 1953a. Observations sur la végétation de la presqu’île de Giens (Var). Bulletin du Muséum d’histoire naturelle de Marseille XIII : 57-69.
Molinier Re., 1953b. La végétation des îles des Embiers (Var). Bulletin de la Société linnéenne de Provence 19 : 26-32.
Molinier Ro., 1959. Étude des groupements végétaux terrestres du Cap Corse. Bulletin du Muséum d’histoire naturelle de Marseille XIX : 5-75.
Nègre R., 1959. Recherches phytogéographiques sur l’étage de végétation méditerranéen aride (sous-étage chaud) au Maroc occidental. Thèse de doctorat d’État, Montpellier, publication de la thèse soutenue en 1956, 385 p.
Nègre R., 1978. Données phytosociologiques sur les associations thérophytiques du Maroc aride. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 23-32.
Paradis G. & Pozzo di Borgo M.-L., 1999. Chorologie et description des stations de l’espèce protégée Gynandriris sisyrinchium (L.) Parl. (Iridaceae) en Corse. Bulletin de la Société botanique du Centre-Ouest, n. s., 30 : 3-20.
Pérez Latorre A.V., Caballero G., Casimiro-Soriguer Solanas F., Gavira O. & Cabezudo B., 2008. Vegetación del sector Malacitano-Axarquiense (Comarca de La Axarquía, Montes de Málaga y Corredor de Colmenar), Málaga (España). Acta Botanica Malacitana 33 : 215-270.
POWO (Plant of the world online), https://powo.science.kew.org.
Sauvage C., 1933-1934. Aperçu sur la végétation du Zerhoun. Bulletin de la Société botanique de France 80 : 742-755, 81 : 75-86.
Wojterski T.W. & Beloued A., 1992. La répartition et la structure de quelques groupements végétaux caractéristiques pour la partie occidentale de la réserve Mergueb près de Bou Saada en Algérie. Documents phytosociologiques, n. s., XIV : 431-446.
Remerciements
Un grand merci à Olivier Argagnon et Philippe Julve pour l’aide à l’acquisition de certaines publications.


