
Données sur des communautés végétales à Lathyrus nudicaulis (Willk.) Amo du Pays basque occidental (Labourd, Pyrénées-Atlantiques, France)
Title
Data on plant communities with Lathyrus nudicaulis (Willk.) Amo from the western Basque Country (Labourd, Pyrénées-Atlantiques, France)
Résumé
Rare espèce endémique ibérique, d’Espagne et du Portugal, Lathyrus nudicaulis (Willk.) Amo a d’abord été découverte en France sur la côte basque, puis plus en retrait de cette dernière (zone rétro-littorale) au cours des années récentes. Elle est proche de L. palustris L., à laquelle elle a été subordonnée au rang de variété et de sous-espèce, et de L. linifolius (Reichard) Bässler var. linifolius. En France, au Pays basque occidental, elle est présente dans des associations végétales de prés hygrophiles aussi bien littoraux qu’intérieurs, de pelouses et de landes littorales, qui sont présentées en détail dans cet article. Deux associations végétales nouvelles à L. nudicaulis sont proposées : le Lathyro nudicaulis-Schoenetum nigricantis et le Galio borealis-Pseudarrhenatheretum longifolii. L’intérêt patrimonial de l’espèce et de celui des types de communautés végétales auxquels elle est associée justifie fortement une protection réglementaire de cette dernière.
Abstract
Rare endemic Iberian species, from Spain and Portugal, Lathyrus nudicaulis (Willk.) Amo was first discovered in France on the Basque coast, then further back (retro-coastline) in recent years. It is close to L. palustris L., to which it was subordinated to the rank of variety and subspecies, and to L. linifolius (Reichard) Bässler var. linifolius. In France, in the western Basque Country, it is present in plant associations of wet meadows, both on the coast and inland, of grasslands and heathlands of the coastline, that are presented in detail in this article. Two new plant associations with L. nudicaulis are proposed: the Lathyro nudicaulis-Schoenetum nigricantis and the Galio borealis-Pseudarrhenatheretum longifolii. The patrimonial interest of the species and that of plant communities where it is strongly justify its regulatory protection.
En prolongement d’une partie des travaux que nous avons réalisés au sein du Conservatoire botanique national Sud-Atlantique (CBNSA, antenne méridionale de Saint-Jean-de-Luz basée au Jardin botanique littoral Paul-Jovet) sur la flore et la végétation du Pays basque occidental en France (province du Labourd, département des Pyrénées-Atlantiques), Lathyrus nudicaulis (Willk.) Amo et les communautés végétales auxquelles l’espèce est associée à des degrés divers ont fait l’objet de nouvelles recherches dont les résultats sont présentés dans cet article. Au préalable, une synthèse bibliographique des connaissances portant sur sa distribution, ses critères de détermination et sur son écologie, est présentée.
1. Données bibliographiques sur Lathyrus nudicaulis
1.1 Distribution générale
L’espèce est endémique de la péninsule Ibérique où elle possède deux aires nettement séparées, l’une nordique en Espagne atlantique (des Asturies au Pays basque, Rioja, provinces de Burgos et de Soria), l’autre au sud du Portugal et de l’Espagne (Dupont, 2015 ; carte 1). Sa présence en France n’a été découverte que récemment, en mai 2006, par un groupe de botanistes (Gatignol, Arlot, Chastenet, Jeanneau, Perroche, Provost et Zunino) au niveau du littoral basque sur la commune d’Hendaye (Gatignol, 2007 ; Coulot & Rabaute, 2016).
1.2 Détermination
Selon Coulot & Rabaute (2016), elle peut être confondue avec Lathyrus palustris L. et avec L. linifolius (Reichard) Bässler dont elle se distingue dans les deux cas par des tiges aptères dans leur partie supérieure, présentant tout au plus deux côtes étroites, alors qu’à ce niveau les ailes sont courtes mais bien développées chez L. palustris (0,2-1 mm) et nettes chez L. linifolius (au minimum 0,5 mm). Les autres différences morphologiques portent d’après les auteurs sur la présence de bractéoles dans les inflorescences (absentes chez L. nudicaulis, présentes chez L. palustris surtout sur les fleurs inférieures), la forme et la taille des stipules des feuilles (très petites et étroites ne dépassant pas 10 mm de long chez L. palustris, dépassant 12 mm de long et plus larges sur les feuilles supérieures de L. nudicaulis) et sur la présence d’un mucron (L. linifolius) ou d’une vrille simple ou ramifiée (L. nudicaulis) au niveau des feuilles supérieures (planches 1 et 2).


Tison et de Foucault (2014) différencient d’abord ces trois espèces sur la présence (L. nudicaulis, L. palustris) ou l’absence (L. linifolius) d’une vrille, au moins sur les feuilles supérieures. La distinction se fait ensuite dans les deux cas sur le caractère aptère de la tige, exceptionnellement à ailes ≤ 0,4 mm à leur plus grande largeur chez L. nudicaulis, à ailes ≥ 0,8 mm à leur plus grande largeur chez L. palustris, ≥ 0,5 mm de large au moins à sa partie supérieure pour L. linifolius.
La flore du Pays basque et des territoires limitrophes (Aizpuru et al., 1999) distingue quant à elle L. nudicaulis de L. linifolius respectivement sur la base de tiges non ailées, parfois anguleuses, ou de tiges ailées, ainsi que sur la présence ou non de vrilles. Deux variétés de L. linifolius y sont mentionnées : var. linifolius caractérisée par ses folioles étroitement linéaires, et var. montanus (Bernh.) Bässler, aux folioles larges, lancéolées à elliptiques-oblongues. L. palustris n’y est pas décrite, mais L. nudicaulis est mise en synonymie de L. palustris L. subsp. nudicaulis (Willk.) P.W. Ball.
Parmi les espèces du genre Lathyrus répertoriées et décrites par Gallego (1999) dans Flora Iberica, L. palustris ne figure pas non plus. Elle apparaît seulement au travers de la correspondance qui y est donnée entre L. palustris var. nudicaulis Willk. et L. palustris subsp. nudicaulis (Willk.) P.W. Ball avec L. nudicaulis (Willk.) Amo. L’auteur fournit une fiche de description très détaillée de cette dernière (dont ses variabilités morphologique, écologique et chorologique) qui a été en partie reprise par Coulot & Rabaute (2016).
Des critères supplémentaires de détermination, portant sur les parties souterraine et aérienne des plantes, sont donnés par Gallego pour distinguer L. linifolius de L. nudicaulis. Selon l’auteur, ils sont utiles à utiliser dans certaines populations du nord de l’Espagne qui ne développent pas de vrilles, où elles se ressemblent alors beaucoup et cohabitent parfois. L. linifolius présente des rhizomes avec des tubérosités tandis que L. nudicaulis possède un rhizome lignifié avec des racines épaisses plus ou moins cylindriques. En ce qui concerne la partie aérienne, la distinction se fait aussi sur le rachis (généralement ailé chez L. linifolius, cannelé chez L. nudicaulis) ainsi que sur les bractées de l’inflorescence qui sont présentes chez L. linifolius (petites, dentées et blanchâtres) et absentes chez L. nudicaulis (Gallego, 1999).
Flora Europaea (Tutin et al., 1968) subordonne également L. nudicaulis à L. palustris (L. palustris L. subsp. nudicaulis (Willk.) P.W. Ball). Les critères de distinction donnés entre les sous-espèces palustris et nudicaulis de L. palustris portent sur les ailes de la tige (étroitement ailée chez L. p. subsp. palustris, non ailée chez L. p. subsp. nudicaulis), sur la couleur de la corolle des fleurs (bleu-violacé pour L. p. subsp. palustris, violet-rouge vif pour L. p. subsp. nudicaulis) et sur celle des graines (tachetées dans L. p. subsp. palustris, noires et non tachetées dans L. p. subsp. nudicaulis). L’aire de répartition de L. p. subsp. palustris ne couvre pas la péninsule Ibérique où seule L. p. subsp. nudicaulis est présente, dans les parties nord et ouest de la péninsule, d’après Tutin et al. (1968).
La flore d’Andalousie occidentale (Valdés et al., 1987) ne mentionne aussi que L. nudicaulis qui est rattaché à L. palustris : L. palustris L. (Sp. Pl. 733, 1753) subsp. nudicaulis (Willk.) P.W. Ball (Feddes Repert. 79 : 47, 1968), mis en synonymie par les auteurs avec L. palustris var. nudicaulis Willk. (Flora (Regensb.) 34 : 630, 1851).
En revanche, la flore du Portugal (Couthino, 1913) cite à la fois L. palustris (L. palustris L.) et L. nudicaulis (L. nudicaulis (Willk.) Samp.). D’après cet ouvrage, L. palustris possède des tiges étroitement ailées, des pédoncules minces, un peu plus longs que les feuilles, 2 à 8 fleurs, des gousses de 7-8 mm de large avec les faces nervées, des graines panachées de brun et de noir, des folioles membraneuses, glabres, elliptiques à oblongues ou linéaires-lancéolées, des stipules semi-hastées, fréquemment petites. L. nudicaulis a des tiges non ailées, des pédoncules à la fin épais et beaucoup plus grands que les feuilles, des gousses de 5 mm de large avec des faces sans nervures, des graines noires, des folioles subcoriaces, plus ou moins glanduleuses à la partie inférieure, oblongues ou étroitement linéaires (forme angustifolius), des stipules semi-hastées, médiocres ou grandes.
Enfin, Willkomm (1852) différencie L. nudicaulis (L. palustris L. var. nudicaulis. Pl. exs. coll. ven. n. 100) de L. palustris sur la base d’une tige non ailée obtuse-anguleuse.
1.3 Variabilités écologique, morphologique et chorologique
En France, Coulot et Rabaute (2016) indiquent que Lathyrus nudicaulis pousse dans des landes à Erica vagans et à brachypode, sur substrat assez minéral peu humide, et que ces conditions stationnelles diffèrent nettement de celles du sud de la péninsule Ibérique où elle croît dans des milieux franchement humides ou tourbeux (cas du Parc national de Doñana en Andalousie).
Dans la péninsule Ibérique, à l’instar de Gallego (1999), les auteurs relèvent que c’est une espèce polymorphe qui possède deux morphologies liées à deux écologies différentes. Au sud-ouest de l’Espagne et dans le sud du Portugal, elle affectionne des milieux humides, tourbeux, comme L. palustris. Elle a alors « toutes les caractéristiques d’un vicariant ibérique de la gesse des marais. La plante est alors assez grande, à longues feuilles terminées par une vrille ramifiée ». Au nord, elle fréquente des habitats moins hygrophiles tels que des prés, des landes et des lisières forestières où « elle est alors moins développée, à feuilles plus courtes, étroites, terminées par une vrille simple (les inférieures par un long mucron) avec un « aspect général intermédiaire entre L. palustris et L. linifolius comme le précise Gallego » ; « C’est le cas des populations françaises de la plante, tant quant à l’écologie qu’aux aspects morphologiques » (Coulot & Rabaute, 2016). Gallego (1999) fournit des précisions sur la diversité des habitats et de la morphologie de l’espèce en péninsule Ibérique : prés, clairières de broussailles, rochers, falaises, herbages des marges de fleuves et de rivières, de mares, de ruisseaux et de marais, dans des lieux très humides ou détrempés, sur des sols sableux, argileux, hygro-tourbeux ou calcaires, entre 0 et 1 400 m d’altitude.
En ce qui concerne sa variabilité morphologique (dont la hauteur de la plante qui varie de 17 à 160 cm), corrélée au moins en partie à sa chorologie et à son écologie, Gallego (1999) note que les populations du nord de l’Espagne sont formées par des individus de taille petite ou moyenne, avec des tiges ascendantes ou plus ou moins dressées, qui ne développent pas de vrilles, ou qui en ont, mais alors fines, simples ou trifurquées, situées seulement dans les feuilles supérieures. L’autre extrême de la variation se rencontre au sud-ouest de la péninsule où les populations sont constituées par des plantes beaucoup plus robustes, grimpantes, avec des vrilles très développées et ramifiées. Ces dernières s’inscrivent uniquement dans des communautés de berges établies sur des sols sableux et hygro-tourbeux. Entre ces deux extrêmes existe tout un gradient de variations (Gallego, 1999).
1.4 Phytosociologie et écologie
En France
Lazare (2017) relève L. nudicaulis au sein de la lande maritime à Erica vagans et Smilax aspera (Smilaco asperae-Ericetum vagantis Lazare & Bioret in Lazare 2017) dont il différencie une sous-association lathyretosum nudicaulis Lazare 2017 à côté de la sous-association typicum (photo 1). Ce type de lande, qui s’étend sur les falaises maritimes situées entre Hendaye au sud, Urrugne au centre (la Corniche basque) et Saint-Jean-de-Luz au nord, est caractérisé par les deux espèces éponymes de l’association et par Rubia peregrina. Elle se distingue de la lande maritime à Glandora prostrata et Erica vagans (Glandoro prostratae-Ericetum vagantis Lazare 2017), plus acidiphile et située plus au nord, entre Guéthary et Biarritz, par l’absence d’Ulex gallii (forme humilis), de Glandora prostrata et de toute autre Ericacée qu’Erica vagans.

La sous-association typicum correspond d’après l’auteur à une « lande fermée assez rase (hauteur 25 à 50 cm) très anémomorphosée des pentes fortes des hauts de falaise et rebords de la corniche », établie sur sol argileux assez mince à pH basique (8,5). La sous-association lathyretosum nudicaulis, dont les relevés ont été réalisés à Hendaye (domaine d’Abbadia) et à Urrugne (parties centrale et orientale de la Corniche basque), « atteint son optimum sur les replats situés en arrière du rebord de la Corniche à une altitude d’au maximum 50 m environ et présente une végétation chaméphytique très fermée et plus élevée (50 à 90 cm de hauteur) participant à l’élaboration d’un sol plus profond, argileux, basique (pH 7,5-8), au complexe argilo-humique saturé en bases ». « Lathyrus nudicaulis (Willk.) Amo peut être assez abondant dans certains relevés et la plupart de ses individus présentent des vrilles fourchues et atteignent jusqu’à 80 cm de hauteur, caractères précisément non conformes à ceux indiqués par Tison & de Foucault (2014) et Coulot & Rabaute (2016) ». Cette sous-association présente une variante plus hygrophile à Pulicaria dysenterica, Molinia caerulea, Schoenus nigricans et Potentilla erecta (Lazare, 2017).
En Espagne
Si l’espèce est répertoriée dans des catalogues floristiques, des atlas ou dans des flores avec des informations plus ou moins précises sur son écologie et sur sa distribution, il est plus difficile de trouver des références bibliographiques sur ses appartenances phytosociologiques. Par exemple L. nudicaulis n’est pas mentionné dans l’étude des végétations du Pays basque de Braun-Blanquet (1967) ni dans le synsystème phytosociologique de l’ensemble de la péninsule Ibérique (Rivas-Martínez et al., 2001 ; Rivas-Martínez et al., 2002) ou de celui focalisé sur le Portugal (Costa et al., 2012).
Au nord de son aire ibérique (Pays basque et territoires limitrophes), Aizpuru et al. (1999) associent L. nudicaulis à des landes et à des herbages frais. Tout près de la frontière avec la France, Catalán et Aizpuru (1985) indiquent qu’il s’observe fréquemment à proximité des ruisseaux du mont Jaizkibel (entre 15 et 200 m d’altitude).
À l’est des Asturies, Nava et al. (1990) relèvent une station dans un pré de fauche hygrophile à 600 m d’altitude, et Carlón et al. (2010) au sein d’herbages entre des roches calcaires à 530 m. Dans le catalogue floristique de la région (Díaz González et al., 1994), L. nudicaulis est noté parmi les prés ou les prairies des Molinio caeruleae-Arrhenatheretea elatioris.
En Navarre où sa présence est jugée douteuse par Dupont (2015), le taxon est cité par Berastegi Gartziandia (2013) parmi les plantes liées aux communautés de prés et de prairies, mais il n’apparaît ni dans les relevés ni dans les textes phytosociologiques de l’auteur. C’est également le cas à l’est de la province de Cantabrie (bassin versant du fleuve Asón) où Herrera (1995) le donne rare sur des sols humides aux étages collinéen et montagnard, mais sans plus de précisions synsystématiques.
À l’extrémité septentrionale de la province de Burgos (Castille-et-León), elle croît dans des prairies hygrophiles, des marges de ruisseaux, des lisières de forêts caducifoliées et d’autres habitats non précisés, depuis les zones basses de la vallée de l’Ebre jusqu’à la Cantabrie (Alejandre Sáenz et al., 2006).
À la limite nord de la même province située au contact de la Cantabrie, Loidi et al. (2014) notent l’espèce dans un relevé sur quatorze qu’ils rattachent au pré pâturé du Merendero pyrenaicae-Cynosuretum cristati Oberd. & Tüxen in Tüxen & Oberd. 1958. Cette association est intégrée à l’alliance du Cynosurion cristati Tüxen 1947, à l’ordre des Arrhenatheretalia elatioris Tüxen 1931 et à la classe des Molinio caeruleae-Arrhenatheretea elatioris Tüxen 1937 dans le synsystème ibérique.
Au nord-ouest de la province de Burgos, dans les monts Obarenes, L. nudicaulis apparaît également dans un relevé phytosociologique sur cinq que García-Mijangos (1997) rattache au pré hygrophile du Scirpo holoschoeni-Molinietum caeruleae (Braun-Blanq. 1931) García-Mijangos 1997 (= Molinietum mediterraneum Braun-Blanq. 1931 d’après l’auteur), association qui relève dans le synsystème ibérique de l’alliance du Molinio-Holoschoenion vulgaris Braun-Blanq. ex Tchou 1948, de l’ordre des Holoschoenetalia vulgaris Braun-Blanq. ex Tchou 1948 et de la classe des Molinio caeruleae-Arrhenatheretea elatioris Tüxen 1937 (Rivas-Martínez et al., 2001 ; Rivas-Martínez et al., 2002). L’association correspond à des communautés végétales dominées par Molinia caerulea et par Scirpoides holoschoenus, caractérisées aussi notamment par Succisa pratensis, Dactylorhiza elata subsp. sesquipedalis, Cirsium pyrenaicum et Oenanthe lachenalii. Elles occupent des lieux plats, sur des sols marneux qui subissent un engorgement d’eau prolongé et où la nappe phréatique se maintient proche de la surface, y compris pendant la période estivale (García-Mijangos, 1997).
À l’extrême sud de l’aire de l’espèce, en Andalousie occidentale, Rivas-Martínez et al. (1980) connaissent L. palustris subsp. nudicaulis dans la Réserve biologique du Parc national de Doñana où ils situent l’espèce parmi les végétations de la classe des Rhamno-Prunetea spinosae Rivas Goday & Borja 1961 (Rhamno-Prunetea spinosae Rivas Goday & Borja ex Tüxen 1962 dans Rivas-Martínez et al., 2001). La classe rassemble des végétations arbustives et épineuses qui en général bordent ou se substituent aux bois caducifoliés développés sur des sols profonds méso-eutrophes pourvus d’une matière organique bien humifiée (mull forestier). Dans leur territoire d’étude, ces végétations se localisent exclusivement au sein de l’écosystème hygrophile riverain de la frênaie (Rivas-Martínez et al., 1980). Le contexte phytosociologique de l’espèce à Doñana est précisé par Rivas-Martínez au travers des travaux de Sáenz (1980) sur les pollens de 83 taxons du site, dont ceux de L. nudicaulis. Elle est située dans des ronciers du Lonicero hispanicae-Rubetum ulmifolii. Cette dernière association décrite dans la zone d’étude des auteurs se développe sur des sols profonds, humides et sableux, méso-oligotrophes, mais non acides en superficie en raison de l’influence de la matière organique apportée par la litière (Rivas-Martínez et al., 1980). L. nudicaulis n’apparaît pas dans le tableau des neuf relevés présentés par les auteurs.
Dans le même secteur, plus récemment (Andalousie, province de Huelva, commune d’Almonte), Garrido et al. (2002) mentionnent une station de Lathyrus nudicaulis proche d’un ruisseau au sein d’une pinède (un seul pied), avec dans les environs Juncus emmanuelis, Eleocharis multicaulis, Ranunculus ophioglossifolius, Baldellia ranunculoides, Scirpus cernuus (Isolepis cernua) et Isoetes histrix. Valdés et al. (2007, 2010) citent également l’espèce dans la région du Parc de Doñana, rare, parmi des formations riveraines sur des sols hygro-tourbeux, sans précisions phytosociologiques.
Au Portugal
Au Portugal, Clemente et al. (2021) résument l’écologie de l’espèce à des prés hygrophiles et à des jonchaies situés en marge de cours d’eau et de zones marécageuses, moins fréquemment en bordure de landes hygrophiles, sur des sols sableux. Carapeto et al. (2014) la signalent au sud-est de Lisbonne au sein de l’habitat d’intérêt communautaire 6410 Prairies à Molinia sur sols calcaires, tourbeux ou argilo-limoneux (Molinion caeruleae), dans des jonchaies acidiphiles dominées par Juncus acutiflorus (J. a. subsp. rugosus, J. rugosus) ou par J. effusus, établies sur des sols sableux très humides de zones basses engorgées une grande partie de l’année. Les auteurs citent parmi les principales espèces liées à ces végétations Bolboschoenus maritimus, Hypericum undulatum, Lythrum junceum, Holcus lanatus et Hyacinthoides vicentina subsp. transtagana.
Bilan
Endémique ibérique, L. nudicaulis est par nature une espèce patrimoniale particulièrement rare à l’échelle mondiale, mais aussi au sein de la péninsule où sa distribution est restreinte au nord, au centre-sud et au sud-ouest de l’Espagne, ainsi qu’au Portugal. Elle est aussi menacée dans une partie de son aire puisqu’elle est classée en danger (EN) dans la Liste rouge d’Andalousie (Cabezudo et al., 2005) et vulnérable (VU) dans celle du Portugal (Carapeto et al., 2020). De même à l’échelle de l’ensemble des Pyrénées tel qu’il est admis par Gómez et al. (2020), les auteurs retiennent L. nudicaulis parmi une liste de 363 plantes rares susceptibles d’intégrer une liste rouge d’espèces dont la conservation est prioritaire. Elle y figure parmi un groupe de 104 taxons présents dans cinq à dix mailles UTM de 10 km de côté avec des populations très réduites.
À la variabilité des conditions écologiques dans lesquelles elle se trouve, souvent quand même dans des zones plus ou moins humides et au nord de son aire dans des prairies au sens large (classe des Molinio caeruleae-Arrhenatheretea elatioris du synsystème ibérique), variabilité elle-même en partie liée à sa chorologie qui « pose un problème » à Dupont (2015), se superpose une variabilité morphologique (Gallego, 1999 ; Coulot & Rabaute, 2016). Ces variabilités pourraient peut-être aussi s’expliquer par une confusion partielle de L. nudicaulis avec L. palustris dans la partie sud de son aire, basée sur le caractère non ailé ou étroitement ailé de la tige (existence de populations de L. palustris à tige aptère et différentes de L. nudicaulis ?), auquel cas L. nudicaulis serait encore plus rare qu’il ne l’est déjà.
Comme le soulignent Coulot & Rabaute (2016), « une étude plus poussée de différentes populations serait vivement souhaitable pour retracer la phylogénie de ce taxon ». Des investigations phytosociologiques également.
2. Méthode d’étude de l’espèce au Pays basque occidental
2.1 Évolution récente des connaissances sur sa distribution et sur sa sociologie
Cette étude s’appuie sur une nouvelle exploitation partielle des observations que nous avons recueillies en 2017 et en 2018, très ponctuellement en 2019, au sein du Conservatoire botanique national Sud-Atlantique (CBNSA) avec le soutien de ses partenaires financiers (planche 3). Les flores de référence utilisées sont celles de Aizpuru et al. (1999) et de Tison & de Foucault (2014).

En 2017, dans le cadre de l’Observatoire de la biodiversité végétale du littoral des Pyrénées-Atlantiques, le littoral basque a fait l’objet d’une étude de sa flore et de sa végétation portant sur près de 40 km de côte situés entre Hendaye au sud (fleuve Bidassoa qui marque la frontière avec l’Espagne) et Anglet au nord (fleuve Adour). Elle a donné lieu à la rédaction d’un rapport général (Dufay et al., 2018), de fiches sur les espèces patrimoniales (Meslage, 2018) et sur les sites à enjeux de la zone d’étude (Meslage & Hardy, 2018). C’est à cette occasion que près de deux cents relevés phytosociologiques (186) ont été réalisés selon la méthode sigmatiste (Guinochet, 1973 ; Géhu & Rivas-Martínez, 1981 ; de Foucault, 1986a) sur le littoral sableux, rocheux et vaseux du Pays basque, relevés situés en général à moins de 500 m de l’océan, à l’exception de la partie amont de quelques fleuves (Bidassoa, Untxin et Uhabia).
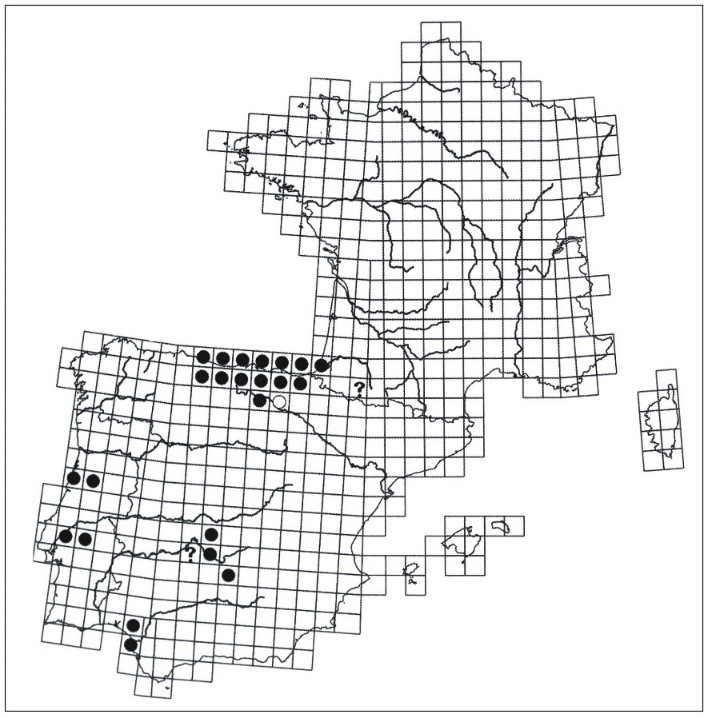
Déjà connu à Hendaye et à Urrugne où les connaissances sur sa distribution ont été affinées, Lathyrus nudicaulis s’est avéré être présent à Bidart, sa limite nord s’établissant au niveau de la localité d’Ilbarritz en limite de Biarritz. Sa distribution présentait en revanche un hiatus important sur le littoral des communes de Ciboure, de Saint-Jean-de-Luz, de Guéthary et du sud de Bidart. L’espèce a été notée au sein de pelouses, de prés et de landes du littoral rocheux.
En 2018, au cours du programme d’inventaires systématiques de la flore du département des Pyrénées-Atlantiques dans un secteur s’étendant en moyenne à une quinzaine de kilomètres en retrait est du littoral (rétro-littoral), nous avons découvert des stations de Lathyrus nudicaulis situées plus à l’intérieur des terres où l’espèce n’avait jamais été signalée. C’est ainsi que nous l’avons découverte sur les communes d’Ahetze, Arbonne, Halsou, Larressore, Saint-Jean-de-Luz, Saint-Pée-sur-Nivelle et Ustaritz. Une trentaine de relevés phytosociologiques (34) ont alors été effectués dans des pelouses et dans des prés qui comportaient ou non l’espèce. En complément, neuf ont été faits en 2019 dans les mêmes types de végétations de ce secteur. Cette découverte récente de la présence de l’espèce en dehors du littoral (en France) s’explique probablement par une pression d’observation plus forte, mais aussi par une confusion partielle avec L. linifolius var. linifolius qui lui ressemble beaucoup. Pour identifier L. nudicaulis avec certitude, nous nous sommes basés sur la présence d’une vrille au moins au niveau d’une feuille avec une tige non ailée.
2.2 Exploitation de relevés phytosociologiques et recherches bibliographiques
Une centaine de relevés phytosociologiques qui possédaient ou non L. nudicaulis (106) a été extraite des travaux réalisés entre 2017 et 2019. Ceux de pelouses et de prés (81 au total) et ceux de landes (25) ont été analysés séparément.
Des recherches bibliographiques ont été effectuées pour identifier des associations végétales auxquelles ces relevés étaient susceptibles d’être rattachés, sur les synsystèmes, ainsi que sur les appartenances phytosociologiques et écologiques des espèces. Les références suivantes ont en particulier été consultées : Julve (1993), Bardat et al. (2004), Bensettiti et al. (2005), de Foucault (1984, 1986b, 1986c, 2008, 2012, 2016), Lazare & Bioret (2006), Royer et al. (2006), de Foucault & Catteau (2012), Lazare (2017), Lafon et al. (2020). Au tableau des 81 relevés de pelouses et de prés ont été ajoutés 63 relevés tirés de la bibliographie (de Foucault, 1984, 1986b ; Lazare & Bioret, 2006) : 22 du tableau princeps du Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008, 6 du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 nom. ined., 8 sur 22 du groupement à Lychnis flos-cuculi–Carum verticillatum B. Foucault 1984 nom. ined., 1 sur 29 de l’Anagallido tenellae-Pinguiculetum lusitanicae (Rivas Goday 1964) B. Foucault 2008, 10 sur 33 du tableau princeps du Carici piluliferae-Pseudarrhenatheretum longifolii (P. Allorge 1941) B. Foucault 1986, 5 du groupement à Brachypodium pinnatum–Pseudarrhenatherum longifolium B. Foucault 1986, 11 du tableau princeps de l’Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006.
Ces 144 relevés ont fait l’objet d’un traitement statistique en présence-absence des espèces via le programme Twinspan du logiciel Juice (Tichý, 2002). Certains d’entre eux ont ensuite été redéplacés manuellement dans les différents groupements végétaux issus de l’analyse en s’appuyant sur leur ensemble spécifique et sur le contexte dans lequel ils avaient été réalisés, c’est-à-dire sans tenir compte exclusivement de la présence ou de l’absence de Lathyrus nudicaulis.
La nomenclature taxonomique utilisée suit la version consultée en 2020 du référentiel national français Taxref (MNHN & OFB, Inventaire national du patrimoine naturel).
3. Résultats
3.1. Le pré hygrophile à Lathyrus nudicaulis et Schoenus nigricans (Lathyro nudicaulis-Schoenetum nigricantis)
Synfloristique et physionomie
Le tableau 1 rassemble 32 relevés, dont trois d’origine bibliographique, réalisés dans ce pré de fauche hygrophile (mésohygrophile à mésophile) particulièrement original et varié floristiquement, qui est caractérisé (planche 4) par Schoenus nigricans (V), Lathyrus nudicaulis (V), Narcissus gigas (III), Sanguisorba officinalis (III), Silaum silaus var. angustifolium (III), Linum catharticum (III), Carex hostiana (II) et Allium ericetorum (II).

Physionomiquement, les espèces à la fois fréquentes et souvent abondantes sont Schoenus nigricans (V), Molinia caerulea (V), Erica vagans (V), Festuca sp. (IV) ou encore Brachypodium rupestre (III). Les Carex globalement y sont souvent bien représentés (C. hostiana, C. panicea, C. pulicaris, C. flacca ; planche 5). Les fleurs de Narcissus gigas ponctuent de jaune le pré au mois de mars et celles d’Allium ericetorum de blanc à partir du mois de septembre.

Comparaison synfloristique et synsystématique
L’association végétale à laquelle ce groupement pourrait très bien être rattaché est celle du Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008 dans sa variante à Schoenus nigricans B. Foucault 1984. On retrouve en effet un fond majoritaire d’espèces partagées avec cette moliniaie à Cirsium filipendulum établie sur des sols minéraux argileux du Pays basque (Pyrénées-Atlantiques) et de Chalosse (sud des Landes) que de Foucault (1984) décrit sur la base de 22 relevés et caractérise par Molinia caerulea, Pseudarrhenatherum longifolium, Erica vagans, Cirsium filipendulum, Tractema umbellata, Betonica officinalis, Ranunculus serpens, Laserpitium prutenicum subsp. dufourianum et Serratula tinctoria subsp. seoanei.
L’auteur distingue une sous-association typicum, notamment différenciée par des espèces d’ourlets préforestiers (Lathyrus linifolius var. linifolius, Euphorbia angulata, Pulmonaria longifolia, Viola riviniana), et une sous-association caretosum verticillati interprétée comme un peu plus acidiphile sur sol un peu plus organique, différenciée par Trocdaris verticillatum (= Carum verticillatum), Dactylorhiza maculata et Juncus acutiflorus. Au sein de la première sous-association, il met en évidence « une variante à Schoenus nigricans, Rubia peregrina, Linum catharticum observée sur les falaises maritimes situées entre Saint-Jean-de-Luz et Hendaye », dans laquelle devait probablement « se localiser Galium boreale sur la côte basque (Lloyd, 1898) » (de Foucault, 1984). Compte-tenu de l’endroit où cette variante a été observée, il est très probable que Lathyrus nudicaulis côtoyait ou remplaçait L. linifolius var. linifolius qui est noté dans les trois relevés de la variante (L. montanus angustifolius). En outre Carex hostiana et Silaum silaus (vraisemblablement sa variété angustifolium) apparaissent dans un relevé, de même que Narcissus sp. dans un autre et qui correspond sans doute à N. gigas.
Nous proposons d’intégrer cette variante (de Foucault, 1984 : relevés 1, 2 et 3 du tableau 138 de l’auteur) à une variation d’une association végétale nouvelle, sous le nom de Lathyro nudicaulis-Schoenetum nigricantis ass. nov. hoc loco (typus nominis hoc loco : relevé 9 = 201901 du tableau 1 hoc loco repris ci-après ; planche 6), interprétée comme plus basiphile que le Cirsio-Molinietum, et qui relève comme lui de la sous-alliance du Serratulo seoanei-Molinienion caeruleae B. Foucault 2008, de l’alliance du Juncion acutiflori Braun-Blanq. in Braun-Blanq. & Tüxen 1952 ou de celle du Molinion caeruleae W. Koch 1926, de l’ordre des Molinietalia caeruleae W. Koch 1926 et de la classe des Molinio caeruleae-Juncetea acutiflori Braun-Blanq. 1950.

Typus nominis hoc loco du Lathyro nudicaulis-Schoenetum nigricantis
Commune d’Ustaritz (64), Bexinenea, 12 juin 2019, 95%, 30 m2, 31 taxons, relevé réalisé par F. Hardy & N. Meslage (tableau 1 hoc loco, relevé 9 = 201901) :
- combinaison caractéristique du Lathyro nudicaulis-Schoenetum nigricantis: Schoenus nigricans 4, Lathyrus nudicaulis 1, Narcissus gigas +, Sanguisorba officinalis 1, Silaum silaus angustifolium +, Carex hostiana +, Allium ericetorum +,
- différentielles du typicum et variantes : Juncus acutiflorus +, Succisa pratensis 1, Pulicaria dysenterica +, Mentha aquatica r, Agrostis canina 1, Trocdaris verticillatum 1, Salix atrocinerea +, Juncus conglomeratus +,
- unités supérieures : Molinia caerulea 3, Cirsium filipendulum r, Serratula tinctoria seoanei +, Carex pulicaris +, Carex panicea 1, Scorzonera humilis +,
- autres : Potentilla erecta +, Danthonia decumbens +, Festuca 2, Erica vagans 1, E. cilaris r, Betonica officinalis r, Pteridium aquilinum r, Lotus corniculatus r, Quercus robur r, Quercus rubra r.
En ce qui concerne l’alliance, si le Serratulo-Molinienion est généralement rattaché au Juncion acutiflori (Bardat et al., 2004 ; Bensetitti et al., 2005 ; Lafon et al., 2020), mais parfois aussi bien au Molinion qu’au Juncion (Bensetitti et al., 2005 ; de Foucault, 2008), nous aurions tendance à intégrer le Lathyro-Schoenetum au Molinion ou à une unité intermédiaire entre Juncion et Molinion dans laquelle se placerait le Serratulo-Molinienion et l’association.
Enfin, outre les espèces de la combinaison caractéristique qui différencient le Lathyro-Schoenetum du Cirsio-Molinietum, mis à part Narcissus gigas et Allium ericetorum qui sont sans doute partagés par les deux associations, même si ces espèces n’apparaissent pas dans le tableau princeps du Cirsio-Molinietum (dates des relevés peu favorables à leur identification), il faut noter l’absence de Calluna vulgaris dans la première alors qu’elle est relativement fréquente dans la deuxième (40 à 60 % des relevés) à l’instar d’autres espèces acidiphiles mais moins notées (Carex pilulifera, Viola lactea, Teucrium scorodonia, Luzula multiflora).
On se reportera au tableau 2 pour comparer en détail les colonnes synthétiques du Lathyro-Schoenetum (colonne LS et ses trois variations LS1, LS2 et LS3) et du Cirsio-Molinietum tel qu’il a été posé initialement avec sa variante à Schoenus nigricans (colonne CMG1) et sans cette variante (colonne CMG2) intégrée au Lathyro-Schoenetum. Trois relevés atypiques dans la mesure où ils ne comportaient pas Schoenus nigricans (relevés 7, 8 et 9 du tableau 2), réalisés dans des végétations de contact, ont été ajoutés et rattachés tout de même à l’association. Le relevé 7 correspond à un microsuintement de moins d’1 m² à Pinguicula grandiflora.
Variations, synécologie et synchorologie
Trois variations principales ayant valeur de sous-association sont proposées au sein du Lathyro nudicaulis-Schoenetum nigricantis (colonne synthétique LS des tableaux 1 et 2) : typicum (relevés 1 à 17 du tableau 1 ; colonne synthétique LS1 des tableaux 1 et 2), trocdarietosum verticillati subass. nov. hoc loco (relevés 18 à 21 du tableau 1 ; typus nominis hoc loco : relevé 18 = 201810 du tableau 1 hoc loco repris ci-après ; colonne synthétique LS2 des tableaux 1 et 2) et potentilletosum montanae subass. nov. hoc loco (relevés 22 à 32 du tableau 1 ; typus nominis hoc loco : relevé 22 = 396364 du tableau 1 hoc loco repris ci-dessous ; colonne synthétique LS3 des tableaux 1 et 2).
Typus nominis hoc loco du trocdarietosum verticillati
commune de Saint-Pée-sur-Nivelle (64), déchèterie (est), 25 mai 2018, 100%, 175 m2, 33 taxons, relevé réalisé par J.-M. Dupont, F. Hardy, P. Lamouroux et F. Rossmann (tableau 1 hoc loco, relevé 18 = 201810) :
- combinaison caractéristique du Lathyro nudicaulis-Schoenetum nigricantis : Schoenus nigricans r, Lathyrus nudicaulis 2, Narcissus gigas +, Silaum silaus angustifolium r, Linum catharticum r, Carex hostiana r, Allium ericetorum +,
- différentielles du trocdarietosum verticillati (partagées avec le typicum) : Trocdaris verticillatum 2, Salix atrocinerea r, Frangula alnus r,
- unités supérieures : Molinia caerulea 5, Cirsium filipendulum r, Serratula tinctoria seoanei +, Carex pulicaris 2, Carex panicea r,
- autres : Potentilla erecta 2, Danthonia decumbens 1, Festuca +, Pseudarrhenatherum longifolium 1, Polygala vulgaris r, Erica vagans 5, E. ciliaris +, Ulex europaeus r, Betonica officinalis +, Pteridium aquilinum 2, Lotus corniculatus 2, Carex flacca +, Ranunculus serpens +, Crataegus monogyna r, Cruciata glabra r, Plantago lanceolata r, Carex caryophyllea r, Rosa sp. r.
Typus nominis hoc loco du potentilletosum montanae
commune d’Urrugne (64), littoral (la Corniche basque), Kauteren Borda (partie sud), 29 juillet 2017, 100%, 50 m2, 25 taxons, relevé réalisé par F. Hardy (tableau 1 hoc loco, relevé 22 = 396364 du tableau 1) :
- combinaison caractéristique du Lathyro nudicaulis-Schoenetum nigricantis : Schoenus nigricans 1, Lathyrus nudicaulis +, Sanguisorba officinalis +, Silaum silaus angustifolium r, Linum catharticum r, Allium ericetorum r,
- différentielles du potentilletosum montanae et variantes : Tractema umbellata r, Potentilla montana +, Pimpinella saxifraga +, Galium boreale 1, Agrostis capillaris 2, Agrostis curtisii 1,
- unités supérieures : Molinia caerulea 2, Cirsium filipendulum r, Serratula tinctoria seoanei +,
- autres : Potentilla erecta 1, Danthonia decumbens 1, Festuca 3, Pseudarrhenatherum longifolium +, Erica vagans 3, Ulex gallii +, Betonica officinalis r, Lotus corniculatus +, Brachypodium rupestre 2, Euphorbia dulcis / E. angulata r.
Trocdaris verticillatum, Salix atrocinerea, Frangula alnus et beaucoup plus rarement Juncus conglomeratus sont des espèces différentielles communes au typicum et au trocdarietosum verticillati. La sous-association typicum, interprétée comme correspondant au pôle le plus hygrophile de l’association (mésohygrophile), est en outre différenciée (planche 7) par Juncus acutiflorus, Lysimachia tenella, Succisa pratensis, Laserpitium prutenicum subsp. dufourianum, Pulicaria dysenterica, Mentha aquatica, Dactylorhiza maculata et des espèces moins fréquentes (Pinguicula lusitanica, Gentiana pneumonanthe, Galium uliginosum, Valeriana dioica, Isolepis setacea, Osmunda regalis, Agrostis canina, Cirsium dissectum, Samolus valerandi, Carex echinata, Pinguicula grandiflora, Juncus bulbosus).

La sous-association potentilletosum montanae, plus mésophile et peut-être plus thermophile, a été observée uniquement sur la côte à Urrugne (la Corniche basque), contrairement aux deux autres sous-associations qui n’ont été notées qu’à l’intérieur (communes d’Ahetze, Arbonne, Halsou, Saint-Pée-sur-Nivelle, Ustaritz). Elle est différenciée (planche 8) par Potentilla montana, Tractema umbellata, Pimpinella saxifraga, Galium boreale, Agrostis capillaris, Asphodelus albus, Glandora prostrata, Rubia peregrina, Agrostis curtisii, Euphorbia flavicoma, Smilax aspera, Poterium sanguisorba, Carex punctata et Leontodon hispidus.

Dans tous les cas le Lathyro-Schoenetum se trouve dans des prés de fauche plus ou moins hygrophiles, généralement comme le Cirsio-Molinietum dans le système mésophile situé en-dehors des fonds de vallée (différent du système hygrophile) tel que le décrit de Foucault (1986b), assez souvent sur des pentes plus ou moins prononcées et sur des sols argileux compacts pour ceux dont nous avons examiné la partie superficielle. Nous n’avons pas étudié leur pH, mais nous pensons qu’il est globalement neutre à basique, parfois neutre à plus acide, si l’on se réfère à l’écologie donnée dans la littérature d’une partie des espèces du cortège floristique de l’association et de ses variations, en particulier Schoenus nigricans, Sanguisorba officinalis, Silaum silaus, Linum catharticum, Carex hostiana, Pulicaria dysenterica, Galium boreale auxquels nous ajoutons Lathyrus nudicaulis. En fait, tels qu’ils sont définis par de Foucault (1984) dans son étude des prairies hygrophiles des plaines atlantiques françaises, le Lathyro-Schoenetum s’insère visiblement dans des systèmes intermédiaires entre système acide et système alcalin, ici mésophiles, planitiaires et sous climat thermo-ombro-atlantique de l’ouest du Pays basque (province du Labourd).
Contacts
En dehors du littoral (photo 2), le Lathyro nudicaulis-Schoenetum nigricantis est fréquemment associé à la pelouse plus mésophile et acidiphile du Carici piluliferae-Pseudarrhenatheretum longifolii (P. Allorge 1941) de Foucault 1986 (voir tableau 4), ce qui est surprenant dans la mesure où l’on aurait pu s’attendre à trouver un groupement mésophile de système intermédiaire, par exemple analogue au groupement à Brachypodium pinnatum–Pseudarrhenatherum longifolium B. Foucault 1986.

Quand le niveau d’humidité le permettait, nous avons observé très ponctuellement un contact avec l’association hygrophile du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 nom. ined. (relevés 1, 2 et 3, colonnes synthétiques CJTO et CJB, tableau 2) et avec un groupement qui s’en rapproche mais avec des particularités floristiques (groupement à Carex echinata et Schoenus nigricans : relevés 4, 5 et 6 du tableau 2, colonne synthétique CJTI).
De même rarement, aussi bien à l’intérieur que sur le littoral, nous avons noté l’association à proximité de végétations que nous avons rattachées au pré hygrophile du Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008 (relevés 10 à 15, colonnes synthétiques CMT, CMG1 et CMG2, tableau 2) malgré la présence diffuse d’espèces du Lathyro-Schoenetum.
Sur la bande littorale étroite à Hendaye et à Urrugne, deux autres types de pelouses remplacent le Carici piluliferae-Pseudarrhenatheretum longifolii bien que nous ayions identifié l’association ponctuellement, qui est par ailleurs bien présente plus en retrait de la côte. La première est une pelouse neutro-acidiclinophile à Brachypodium rupestre qui relève de l’Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006 (tableau 3) et l’autre d’un groupement mésophile très particulier qui présente un caractère intermédiaire entre cette association, le Lathyro-Schoenetum (et le Cirsio-Molinietum) et le Carici-Pseudarrhenatheretum. C’est ce dernier groupement qui comporte notamment Lathyrus nudicaulis, Pseudarrhenatherum longifolium, Galium boreale, Brachypodium rupestre, Potentilla montana, Euphorbia flavicoma, Tractema umbellata, Glandora prostrata, Helictochloa marginata, Silaum silaus var. angustifolium, Agrostis curtisii… (tableau 5 : Galio borealis-Pseudarrhenatheretum longifolii), actuellement connu seulement de trois sites de la Corniche basque à Urrugne, que nous avons observé au moins une fois non loin du Lathyro-Schoenetum potentilletosum montanae qu’il relaie en conditions plus mésophiles. La distinction entre ce groupement et la sous-association potentilletosum montanae du Lathyro-Schoenetum n’est pas toujours aisée à faire si l’on ne se base que sur la présence-absence des espèces.
Syndynamisme
Sur le plan syndynamique, la présence et l’abondance dans le Lathyro-Schoenetum d’espèces des Calluno vulgaris-Ulicetea minoris Braun-Blanq. & Tüxen ex Klika in Klika & Hadač 1944, excepté Calluna vulgaris (Erica vagans surtout, E. ciliaris, Ulex gallii, U. europaeus, parfois sur le littoral Glandora prostrata et exceptionnellement Erica cinerea), laissent présager une évolution possible de l’association vers des landes à Erica vagans.
À l’intérieur, ces landes (photo 3) pourraient correspondre à celles que Rivas-Martínez et al. (1984) rapprochaient du Cirsio filipenduli-Ericetum ciliaris Braun-Blanq., P. Silva & Rozeira 1964 ericetosum vagantis Rivas Mart. 1979 dans leur étude de la vallée de la Bidassoa (Espagne) et qu’ils mettaient en synonymie de l’Ulici-Ericetum ciliaris Braun-Blanq. 1967. Loidi et al. (1996) retiennent ce dernier nom (dans lequel l’espèce d’Ulex n’est pas précisée) en désignant un relevé type de l’association et de la sous-association ericetosum vagantis Rivas Mart. 1979 réalisé par Braun-Blanquet à San Sebastian (1967) où figurent Ulex europaeus et U. minor, ce dernier noté selon eux de manière erronée par Braun-Blanquet et qu’ils remplacent par U. gallii. L’aire du Cirsio filipenduli-Ericetum ciliaris, qui avait été étendue initialement au Pays basque, est limitée par les auteurs à la Galice et au Portugal. Les deux associations sont mentionnées par Rivas-Martínez et al. (2001) dans le synsystème ibérique, avec une mutation du nom d’Ulici-Ericetum ciliaris en Ulici gallii-Ericetum ciliaris Braun-Blanq. 1967. Le Lathyro-Schoenetum (sous-associations typicum et trocdarietosum verticillati) pourrait évoluer vers l’Ulici-Ericetum ciliaris ericetosum vagantis, peut-être au travers d’une variation plus basiphile à Schoenus nigricans dans laquelle Lathyrus nudicaulis se maintiendrait parfois.

Sur le littoral, les landes à Erica vagans du plateau des falaises relèvent du Smilaco asperae-Ericetum vagantis Lazare & Bioret in Lazare 2017 et du Glandoro prostratae-Ericetum vagantis Lazare 2017, deux associations qui à notre avis ne s’excluent pas géographiquement, contrairement à ce qu’indique Lazare (2017). Pour la seconde, nous n’avons pas préciséé s’il s’agissait de la forme humilis d’Ulex gallii dans nos relevés, mais la présence de cette espèce (et d’autres) nous a conduits à rattacher plusieurs d’entre eux à l’association bien qu’ils aient été réalisés dans des secteurs où le Smilaco-Ericetum était aussi identifié, en particulier sur la Corniche basque à Urrugne (tableaux 6 et 7). On peut imaginer une dynamique possible du Lathyro-Schoenetum potentilletosum montanae vers les variantes hygrophiles des deux associations.
Par ailleurs, il nous semble que la lande littorale du Leucanthemo crassifolii-Ericetum vagantis (P. Allorge & Jovet 1941) Géhu & Géhu-Franck 1981, reconsidérée par Lazare (2017), existe bien et qu’elle se situe dans la partie supérieure des pentes très prononcées, et par suite difficilement accessibles, de falaises littorales situées à Hendaye, Urrugne et Saint-Jean-de-Luz, évoquant une lande à Leucanthemum crassifolium et Erica vagans « suspendue » au-dessus de l’océan. Nous n’avons pu faire que deux relevés dans ce type de végétation (non présentés dans le cadre de ce travail) mais nous avons bien observé le télescopage étonnant des deux espèces éponymes au sein de landes rases « suspendues » dans lesquelles les têtes fleuries de Leucanthemum crassifolium ressortaient d’un tapis dense d’Erica vagans.
Intérêt patrimonial
La présence constante de Lathyrus nudicaulis dans le Lathyro nudicaulis-Schoenetum nigricantis confère à ce pré de fauche mésohygrophile à mésophile, varié floristiquement (nombre spécifique moyen : 29), qui est par ailleurs rare et de distribution restreinte en France à l’extrémité occidentale du Pays basque (littorale et rétro-littorale, Labourd), un très fort intérêt patrimonial. Parmi les autres espèces remarquables de la combinaison caractéristique de l’association, la variété angustifolium de Silaum silaus est également indiquée comme rare à l’échelon national français par Tison & de Foucault (2014), répertoriée dans le Jura, le Massif central et les Pyrénées. Carex hostiana est classé quant à lui vulnérable (VU) dans la Liste rouge d’ex-Aquitaine (CBNSA, CBNMC & CBNPMP, Observatoire de la biodiversité végétale de Nouvelle-Aquitaine, 2021) et figure parmi la Liste des espèces déterminantes ZNIEFF de la Nouvelle-Aquitaine (Abadie et al., 2019), comme Schoenus nigricans en-dehors du triangle landais, Allium ericetorum, Narcissus gigas et Sanguisorba officinalis.
C’est également le cas, au sein des différentes variations de l’association ou parmi les espèces des unités supérieures (planche 9), de Laserpitium prutenicum subsp. dufourianum, Pinguicula lusitanica, Gentiana pneumonanthe, Valeriana dioica à l’échelon des Pyrénées-Atlantiques, Carex echinata hors massif pyrénéen et Massif central, Carex pulicaris hors massif pyrénéen, Tractema umbellata, Smilax aspera, Carex punctata en dehors du triangle landais, Galium boreale et Glandora prostrata (Abadie et al., 2019). Les deux dernières espèces sont protégées réglementairement, respectivement au niveau régional (JORF, 2002) et au niveau national (JORF, 1982), et sont toutes deux considérées comme quasi menacées (NT) dans la Liste rouge d’ex-Aquitaine.

Bien que ne figurant pas dans une liste d’espèces à statut, Il faut aussi souligner la présence « abyssale » de Pinguicula grandiflora à une cinquantaine de mètres d’altitude dans un relevé attribué au Lathyro-Schoenetum réalisé à Ahetze (relevé 6 du tableau 1) et dans un autre au sein d’une communauté végétale qui s’en rapproche également rattachée à l’association (relevé 7 du tableau 2), effectué sur la commune d’Ustaritz à un niveau altitudinal du même ordre.
Enfin, la position synsystématique du Lathyro-Schoenetum (sous-alliance du Serratulo seoanei-Molinienion caeruleae, alliances du Juncion acutiflori et/ou du Molinion caeruleae, ordre des Molinietalia caeruleae et classe des Molinio caeruleae-Juncetea acutiflori) l’intègre à l’habitat d’intérêt communautaire des Prairies à Molinia sur sols calcaires, tourbeux ou argilo-limoneux (Molinion caeruleae) (code Eur 28 : 6410 ; Romão, 1997 ; Bensettiti et al., 2005 ; Anonyme, 2007).
3.2 Les autres prés hygrophiles à Lathyrus nudicaulis
Le pré à Cirsium filipendulum et Molinia caerulea (Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008)
Le tableau 2 présente six relevés (relevés 10 à 15 synthétisés dans la colonne CMT, réalisés sur le littoral et à l’intérieur à Ahetze, Larressore, Saint-Pée-sur-Nivelle, Urrugne, Ustaritz) que nous rattachons à cette moliniaie du système mésophile (de Foucault, 1986b) présentée plus haut et qui s’insère dans l’habitat d’intérêt communautaire des Prairies à Molinia sur sols calcaires, tourbeux ou argilo-limoneux (Molinion caeruleae) (code Eur 28 : 6410), plus précisément au sein de l’habitat élémentaire des Prés humides acidiphiles thermo-atlantiques sur sol à assèchement estival (code Eur 28 : 6410-10) (photo 4). Les colonnes synthétiques CMG1 et CMG2 de ce tableau correspondent respectivement à la synthèse des 22 relevés de la description initiale de l’association (de Foucault, 1984) et à celle de cette dernière moins les trois relevés de la variante à Schoenus nigricans identifiée par de Foucault (1984) qui ont été intégrés au Lathyro nudicaulis-Schoenetum nigricantis, soit un total de 19 relevés.

Trocdaris verticillatum et Juncus acutiflorus permettent d’identifier la sous-association caretosum verticillati B. Foucault 2008 dans les relevés 10 à 14, le relevé 15 se rattachant plus à la sous-association typicum malgré la présence de Dactylorhiza maculata. Nous interprétons l’ensemble de ces relevés comme des variations à espèces du Lathyro-Schoenetum dans le Cirsio-Molinietum (selon les cas Lathyrus nudicaulis ou/et Carex hostiana, Narcissus gigas, Schoenus nigricans, Silaum silaus var. angustifolium, Linum catharticum, Galium boreale, Pimpinella saxifraga), mais sans avoir d’explication sur les processus écologiques qui déterminent la présence diffuse de ces espèces et l’éventuel passage graduel d’une association à l’autre : acidification progressive du Lathyro-Schoenetum ou alcalinisation du Cirsio-Molinietum et, si c’est le cas, selon quelles modalités détaillées ? Lathyrus nudicaulis a été noté dans trois relevés sur six rapportés à cette association, dont un dans lequel figurait simultanément Lathyrus linifolius, tous au sein du caretosum verticillati.
Le pré à Trocdaris verticillatum et Juncus acutiflorus (Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 nom. ined.)
Description, synécologie et synsystématique
Ce pré hygrophile de marais tourbeux, dont de Foucault (1984) rapporte six relevés qu’il a réalisés dans des fonds de vallées du Pays basque occidental, est caractérisé par Juncus acutiflorus, Lysimachia tenella, Carex pulicaris, Trocdaris verticillatum, Scutellaria minor, Carex echinata, Wahlenbergia hederacea, Carex hostiana, et globalement par la rareté des espèces prairiales et par l’abondance des espèces de bas-marais. L’auteur note qu’il se différencie d’autres végétations rattachées au Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 (« eu-atlantique ») par l’absence de Cirsium dissectum, Hydrocotyle vulgaris, Carex nigra, et l’interprète comme une race basque originale, qui, par la présence de Carex hostiana et de C. pulicaris, se rapproche des variations basiclinophiles de l’association, ce qui « prouve que les substrats des marais basques étudiés ici ne sont que modérément acides » (de Foucault, 1984).
Dans le synsystème, l’association est intégrée à la sous-alliance du Caro verticillati-Juncenion acutiflori B. Foucault & Géhu 1980, l’alliance du Juncion acutiflori Braun-Blanq. in Braun-Blanq. & Tüxen 1952, l’ordre des Molinietalia caeruleae W. Koch 1926 et à la classe des Molinio caeruleae-Juncetea acutiflori Braun-Blanq. 1950. Elle fait partie de l’habitat élémentaire d’intérêt communautaire des Prés humides et bas-marais acidiphiles atlantiques (code Eur 28 : 6410-6).
Synchorologie, comparaison synsystématique, synécologie et contacts
Nous présentons trois relevés que nous rattachons à cette association (relevés 1, 2 et 3 du tableau 2 effectués à Saint-Pée-sur-Nivelle et Ustaritz, colonne synthétique CJTO, comparés à la colonne synthétique CJB des six relevés de la race basque B. Foucault 1984 du Caro-Juncetum), dont un à Lathyrus nudicaulis et Narcissus gigas. Ce dernier (relevé 3) correspond à une petite jonchaie mouilleuse à Juncus acutiflorus et Trocdaris verticillatum (avec également Cirsium dissectum et Agrostis canina) qui curieusement était située au contact supérieur du Lathyro-Schoenetum typicum (et non inférieur comme on aurait pu l’imaginer), sous une moliniaie fermée et paucispécifique non échantillonnée (probablement du Cirsio-Molinietum), et sur une pente en-dehors d’un fond de vallée. L’ensemble était environné d’une lande haute à Ulex gallii et Erica vagans, et de fourrés à Salix atrocinerea.
Les deux autres relevés rattachés au Caro-Juncetum basque (sans Lathyrus nudicaulis), localisés au contact l’un de l’autre (le n° 2 du tableau 2 au niveau inférieur du n° 1 ; photo 5), concernent des jonchaies hygrophiles à Juncus acutiflorus, Trocdaris verticillatum et Eleocharis multicaulis effectivement situées dans le fond d’un petit vallon. En outre, on y relève notamment selon les cas Carex hostiana (présent et abondant dans le niveau supérieur), Sphagnum sp., Pinguicula lusitanica, Ranunculus flammula, Carex demissa, Hypericum elodes, Lotus pedunculatus, Alnus glutinosa, Isolepis setacea. Parmi les espèces de la combinaison caractéristique du Caro-Juncetum race basque, il y manque Carex echinata et Wahlenbergia hederacea. Sur le plan écologique, le sol de ces jonchaies était argileux, très humide, et non tourbeux.

À leur niveau supérieur, en bas de pente et développée sur un sol portant sec, se trouvait une pelouse mésophile à Brachypodium rupestre, Festuca sp. et Pteridium aquilinum que nous avons rattachée au Cirsio filipenduli-Molinietum caeruleae caretosum verticillati (relevé 14 du tableau 2) dans une variation à espèces du Lathyro-Schoenetum (Carex hostiana +, Schoenus nigricans r, Linum catharticum r) malgré la faible abondance de Molinia caerulea (+). Elle-même était située au contact inférieur d’une pelouse à Agrostis curtisii, Pteridium aquilinum et Pseudarrhenatherum longifolium (plus, hors relevé, Helictochloa marginata abondant dans la partie supérieure de la parcelle) du Carici piluliferae-Pseudarrhenatheretum longifolii (P. Allorge 1941) B. Foucault 1986 brachypodietosum pinnati B. Foucault 1986, dans sa variante plus hygrophile à Molinia caerulea, Cirsium filipendulum, Dactylorhiza maculata (relevé 6 du tableau 4), qui occupait la majeure partie de la parcelle.
En conclusion, Lathyrus nudicaulis s’observe donc parfois (un relevé sur trois) dans la race basque du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962, race qui pourrait éventuellement être différenciée pour tout ou partie des relevés en sous-association plus basiphile avec notamment Carex hostiana, espèce partagée avec la sous-association juncetosum subnodulosi B. Foucault 2008 du Caro-Juncetum acutiflori.
Le groupement à Carex echinata et Schoenus nigricans
Description et synchorologie
Ce pré hygrophile de fauche est assez proche de la jonchaie à Juncus acutiflorus du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 nom. ined. avec notamment Carex echinata, Eleocharis multicaulis, Lotus pedunculatus et Carex hostiana, mais il en diffère globalement par la présence d’espèces caractéristiques du Lathyro-Schoenetum (Schoenus nigricans, Lathyrus nudicaulis et Narcissus gigas en plus de Carex hostiana) ou partagées par ce dernier avec le Cirsio-Molinietum (Betonica officinalis, Serratula tinctoria subsp. seoanei, Laserpitium prutenicum subsp. dufourianum, Erica vagans et Cirsium filipendulum). Nous disposons de trois relevés de ce groupement effectués dans deux sites différents (relevés 4 et 5 à Saint-Jean-de-Luz et relevé 6 à Ahetze, colonne synthétique CJTI, tableau 2).
Intérêt patrimonial, synécologie, contacts et synsystématique
Pinguicula lusitanica est présent avec Sphagnum sp. dans deux relevés (n° 5 et n° 6), dont l’un avec l’espèce protégée nationalement Drosera rotundifolia (JORF, 1982). Le troisième (n° 4) comporte Epipactis palustris, espèce caractéristique de bas-marais neutrobasiphiles comme Schoenus nigricans et Carex hostiana (de Foucault, 1984 ; Julve, 1993), et qui est protégée régionalement en Nouvelle-Aquitaine (JORF, 2002). On y trouve aussi Valeriana dioica. Ce groupement présente donc un fort intérêt patrimonial et, comme ceux présentés précédemment, mêle étonnamment des espèces basiphiles et d’autres acidiphiles.
Lathyrus nudicaulis y a été noté dans les trois relevés, avec un coefficient d’abondance-dominance particulièrement élevé de 3 sur une surface de 50 m² dans l’un d’entre eux (relevé 4 du tableau 2 ; effectifs de la population de Lathyrus compris entre 100 et 500 tiges), issu d’un pré ras à Succisa pratensis abondant (coefficient 5), ainsi que Juncus acutiflorus (3) et Schoenus nigricans (3), avec Epipactis palustris (+) déjà évoqué (photo 6). Il était situé en-dehors d’un fond de vallée au contact d’un pré ras ouvert à Eleocharis multicaulis (3), Carex echinata (3), Schoenus nigricans (2) et Succisa pratensis (2) dans lequel se trouvait notamment Sphagnum sp., Pinguicula lusitanica et Drosera rotundifolia (relevé 5 du tableau 2), au sein d’une gouille circulaire d’une bordure de petite zone en eau évoquant une résurgence. Le reste de la parcelle, exploitée par fauche, était occupé au moins en partie par une pelouse de la variante à Molinia caerulea du Carici piluliferae-Pseudarrhenatheretum longifolii B. Foucault 1986 typicum (Molinia caerulea 3, Cirsium filipendulum 1, Laserpitium prutenicum subsp. dufourianum +, Trocdaris verticillatum +, Dactylorhiza maculata +, Serratula tinctoria subsp. seoanei +, Scorzonera humilis +, Erica ciliaris +) qui fait la transition avec le Cirsio filipenduli-Molinietum caeruleae B. Foucault 2008 dont elle est proche et pas toujours facile à distinguer, où se trouvait également l’espèce protégée Glandora prostrata (relevé 10 du tableau 4).

Le relevé 6 est un relevé linéaire de quelques m² (10) localisés sur une partie de la bordure supérieure d’un fossé à Schoenus nigricans et Molinia caerulea dont la végétation a été rattachée au Lathyro nudicaulis-Schoenetum nigricantis typicum (relevé 2 du tableau 1) qui était lui-même situé au niveau inférieur d’un pré sur pente relevant du même syntaxon (relevé 13 du tableau 1). Comme dans le site précédent, la variante à Molinia caerulea du Carici piluliferae-Pseudarrhenatheretum longifolii typicum a été identifiée dans les environs (relevé 14 du tableau 4).
La position synsystématique du groupement à Carex echinata et Schoenus nigricans paraît identique à celle du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984.
3.3. Les pelouses à Lathyrus nudicaulis
La pelouse littorale à Euphorbia flavicoma et Silaum silaus (Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006)
Description, synécologie, syndynamisme et synsystématique
Lazare et Bioret (2006) caractérisent cette pelouse à Brachypodium rupestre, décrite à partir de onze relevés provenant du littoral d’Hendaye (domaine d’Abbadia) et d’Urrugne (Corniche basque), par Euphorbia flavicoma DC. subsp. occidentalis Laínz (d’après les auteurs mis en synonymie par Flora iberica avec E. polygalifolia Boiss. & Reut. subsp. vasconensis Vivant ex Kerguélen & Lambinon), Cirsium filipendulum et Silaum silaus (L.) Schinz & Thell. var. pyrenaicus Burnat. Ce dernier taxon semble correspondre à S. silaus var. angustifolium (Wallr.) Thell. dans Flora Gallica (Tison & de Foucault, 2014) qui précise qu’il s’agit d’une variété rare en France (Jura, Massif central, Pyrénées, mais particulièrement commune au Pays basque) et qui « pourrait mériter un rang plus élevé (à étudier) ».
Au sujet d’Euphorbia polygalifolia subsp. vasconensis (« vasconcensis Vivant »), Flora Gallica indique qu’il a été « décrit de Saint-Jean-de-Luz d’après une population glabre » et qu’il « a été souvent amalgamé à tort avec E. flavicoma subsp. occidentalis Laínz du NO de l’Espagne », ce dernier « en fait bien distinct par sa pilosité de type mariolensis/costeana et par son écologie strictement calcicole ». L’ouvrage inclut le taxon dans E. flavicoma subsp. verrucosa (Fiori) Pignatti (planche 10).

La pelouse de l’Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006 (photo 7) se développe sur substrat carbonaté dont le sol est légèrement lessivé et décalcifié superficiellement (pH 6,5 à 7), et « présente un caractère nettement thermophile, occupant les pentes bien exposées (SE à SW en majorité) situées à proximité du littoral de la Corniche basque ». Elle évolue rapidement, via des stades enrichis en espèces landicoles, vers des landes à Erica vagans (Leucanthemo crassifolii-Ericetum vagantis pour les stations les plus proches des falaises littorales, Ulici gallii-Ericetum ciliaris pour celles qui sont plus en retrait) si « elle n’est pas entretenue par une gestion agropastorale très extensive (fauche et/ou pacage légers) » (Lazare & Bioret, 2006). Comme cela a déjà été évoqué et discuté plus haut dans cet article (voir le Lathyro-Schoenetum littoral : potentilletosum montanae), il faut rappeler ici que le Leucanthemo crassifolii-Ericetum vagantis a été redéfinie par Lazare en 2017 qui a décrit un Smilaco asperae-Ericetum vagantis (typicum et lathyretosum nudicaulis, dont une variante hygrophile dans cette dernière sous-association) et un Glandoro prostratae-Ericetum vagantis (typicum et ericetosum cinereae, l’auteur distinguant également une variante hygrophile dans le deuxième cas).

Enfin, Lazare et Bioret (2006) intègrent l’Euphorbio occidentalis-Silaetum pyrenaici à l’alliance du Potentillo montanae-Brachypodion rupestris Braun-Blanq. 1967 corr. Rivas Mart. et al. 2002, l’ordre des Brometalia erecti W. Koch 1926 et la classe des Festuco valesiacae-Brometea erecti Braun-Blanq. & Tüxen ex Braun-Blanq. 1949. Ils la situent par ailleurs dans l’habitat d’intérêt communautaire Pelouses calcicoles mésophiles des Pyrénées et du piémont nord-pyrénéen (code Eur 28 : 6210-6).
Synchorologie, comparaison synsystématique et variations
Le tableau 3 rassemble dix relevés de pelouses à Brachypodium rupestre effectués à Hendaye (domaine d’Abbadia) et à Urrugne (Corniche basque) que nous rapprochons de cette association (relevés 1 à 10, colonne synthétique ESG), dont certains sont très particuliers et pourraient être classés ailleurs mais placés ici par défaut. Ces relevés et leur synthèse peuvent être comparés à la colonne synthétique ES1 qui est celle du tableau princeps de l’Euphorbio occidentalis-Silaetum pyrenaici (tableau 2 de Lazare & Bioret, 2006). La colonne ES2 correspond à la même source dont nous avons extrait un relevé (relevé 11 du tableau 2 de Lazare & Bioret = relevé 11 du tableau 3 hoc loco) qui a été intégré à un autre groupement original décrit plus loin (Galio borealis-Pseudarrhenatheretum longifolii, tableau 5).
Nous avons distingué différentes variations qui s’entrecroisent :
- à Galatella linosyris, Rubia peregrina et Smilax aspera de transition dynamique avec la lande du Smilaco-Ericetum vagantis (relevé 1),
- à Tephroseris helenitis macrochaeta (relevé 2),
- à Leucanthemum crassifolium et Leontodon hispidus (relevés 2, 3 et 4),
- à Rhinanthus angustifolius et Galium verum qui présente un caractère prairial assez prononcé avec notamment Lathyrus pratensis, Trifolium pratense, Vicia cracca, Dactylis glomerata, Linum usitatissimum angustifolium, Arrhenatherum elatius, Trifolium patens… et paraissant susceptible d’évoluer par eutrophisation vers la prairie du Lino biennis-Cynosuretum cristati P. Allorge ex Tüxen & Oberd. 1958 dont la sous-association oenanthetosum pimpinelloidis B. Foucault 1986 a été reconnue à Hendaye (relevés 5 et 6),
- à Succisa pratensis et plusieurs espèces hygrophiles telles que Lotus pedunculatus, Mentha aquatica, Lythrum salicaria et Juncus conglomeratus (relevé 7),
- à Galium boreale (relevés 7, 8 et 9),
- à Sanguisorba officinalis (relevés 9 et 10).
Intérêt patrimonial
Lathyrus nudicaulis est présent dans huit relevés sur dix et semble pouvoir être intégré à la combinaison caractéristique de l’Euphorbio occidentalis-Silaetum pyrenaici. Lathyrus linifolius, que nous n’avons pas noté, figure dans cinq relevés sur onze de Lazare & Bioret (2006), mais il s’agissait probablement de L. nudicaulis dans au moins une partie des stations.
D’autres espèces plus ou moins fréquentes dans les relevés présentés renforcent le fort intérêt patrimonial de ces communautés végétales (planche 11). Ainsi Tephroseris helenitis subsp. macrochaeta, qui n’est connu en France qu’au Pays basque et qui est globalement rare à l’échelle mondiale puisqu’il s’agit d’un taxon vasco-cantabrique (Tison et de Foucault, 2014), est protégé nationalement (JORF, 1982), tout comme Leucanthemum crassifolium également vasco-cantabrique et qui normalement se rencontre sur le pentes des falaises basques, ainsi que Glandora prostrata. Galium boreale, déjà mentionné dans le Lathyro-Schoenetum potentilletosum montanae, est quant à lui protégé régionalement (JORF, 2002). Par ailleurs, Galatella linosyris, sans doute représenté ici par sa très rare variété armoricana (Rouy) Bock & Tison, pour l’instant uniquement connue en France sur le littoral du Morbihan (Bretagne), est inscrit sur la Liste des espèces déterminantes ZNIEFF de la Nouvelle-Aquitaine (Abadie et al., 2019). C’est également le cas de Lathyrus nudicaulis, Schoenus nigricans (hors triangle Landais), Sanguisorba officinalis, Trifolium patens, Prunella hastifolia (hors Massif pyrénéen), Tractema umbellata, Smilax aspera et des espèces protégées citées précédemment.

La pelouse à Carex pilulifera et Pseudarrhenatherum longifolium (Carici piluliferae-Pseudarrhenatheretum longifolii B. Foucault 1986)
Description, synécologie et synsystématique
C’est une pelouse oligotrophile, acidiphile et mésophile décrite au Pays basque par de Foucault (1986b) qui la définit par l’assemblage de trois ensembles sociologiques :
- des hémicryptophytes et des géophytes oligotrophiles thermo-atlantiques ou à répartition plus large (Pseudarrhenatherum longifolium, Agrostis curtisii, Helictochloa marginata, Tractema umbellata, Carex pilulifera, Carex caryophyllea, Polygala serpyllifolia, Potentilla erecta, Pilosella officinarum, Veronica officinalis, Avenella flexuosa, Ranunculus serpens…) qui constituent d’après l’auteur le noyau caractéristique du groupement,
- des hémicryptophytes de forêts ou d’ourlets acidiphiles à neutrophiles atlantiques (Pteridium aquilinum, Hypericum pulchrum, Solidago virgaurea, Euphorbia angulata, Asphodelus albus),
- et des chaméphytes de landes ibéro-atlantiques (Erica vagans, cinerea, Calluna vulgaris, Ulex europaeus, plus rarement Glandora prostrata surtout au voisinage de l’océan).
Pseudarrhenatherum longifolium, Carex pilulifera, Agrostis curtisii, Ranunculus serpens, Tractema umbellata, Polygala serpyllifolia et Helictochloa marginata forment la combinaison caractéristique du Carici piluliferae-Pseudarrhenatheretum longifolii B. Foucault 1986. L’auteur distingue une sous-association brachypodietosum pinnati B. Foucault 1986 et une sous-association typicum, respectivement interprétées comme correspondant aux volets le plus et le moins neutrophiles de l’association, puis trois variantes homologues au sein de chacune d’entre elles. Il précise notamment que les troisièmes variantes de l’une ou de l’autre sous-association, visiblement plus hygrophiles et qui ont en commun Molinia caerulea, Cirsium filipendulum, Dactylorhiza maculata, Betonica officinalis, Succisa pratensis, Serratula tinctoria subsp. seoanei puis selon les cas Trocdaris verticillatum, Laserpitium prutenicum subsp. dufourianum, Erica tetralix, Gentiana pneumonanthe ou Scorzonera humilis « rappellent fortement le Cirsio filipenduli-Molinietum ». Il estime que « les trois variantes de chaque sous-association constituent autant d’étapes d’une série édapho-dynamique menant du Carici-Pseudarrhenatheretum à une forme du Cirsio-Molinietum, avec disparition progressive de quelques espèces typiques de la pelouse et apparition d’espèces du Molinion (Serratulo seoanei-Molinienion), les premières indicatrices de cette dynamique étant surtout Betonica officinalis et Cirsium filipendulum » (de Foucault, 1986b).
L’association (photo 8) relève de l’alliance de l’Agrostion curtisii B. Foucault 1986, de l’ordre des Nardetalia strictae Oberd. ex Preising 1950 et de la classe des Nardetea strictae Rivas Goday in Rivas Goday & Rivas Mart. 1963. Elle fait partie de l’habitat élémentaire d’intérêt communautaire prioritaire Pelouses acidiphiles thermo-atlantiques (code Eur 28 : 6230*-5).

Comparaison synfloristique, synchorologie, synécologie et contacts
Le tableau 4 présente quinze relevés phytosociologiques que nous rapportons à cette association. Les relevés 1 à 6 (colonne synthétique CPTB) se rapprochent du brachypodietosum pinnati par Brachypodium rupestre, et les relevés 7 à 15 (colonne synthétique CPTT) du typicum. L’ensemble est synthétisé dans la colonne CPTG qui peut être comparée aux colonnes synthétiques initiales de description de l’association (CPBP pour les dix-sept relevés du brachypodietosum pinnati, CPT pour les seize du typicum, CP pour l’ensemble de l’association ; de Foucault, 1986b, tableau 1 de l’auteur). Nous avons ajouté la colonne synthétique en nombre d’occurrences des cinq relevés princeps du groupement à Brachypodium pinnatum et Pseudarrhenatherum longifolium B. Foucault 1986 (colonne GBBP ; de Foucault, 1986b, tableau 3 de l’auteur) qui est plus nettement neutrophile que le Carici-Pseudarrhenatheretum (de Foucault, 2012).
Globalement, on note par rapport à la description initiale de l’association et de ses variations que ne figurent pas dans nos relevés Pilosella officinarum, Avenella flexuosa, Hypericum pulchrum, Blechnum spicant, Eupatorium cannabinum et Erica cinerea, ainsi que Galium verum et Teucrium scorodonia pour la sous-association brachypodietosum pinnati, Erica tetralix pour la variante hygrophile du typicum. À l’inverse, Carex flacca, Ulex gallii et Erica ciliaris sont présents et assez fréquents. Carex pilulifera est très peu noté.
Les relevés effectués (à l’intérieur et sur le littoral à Ahetze, Bidart, Hendaye, Saint-Jean-de-Luz, Saint-Pée-sur-Nivelle, Urrugne, Ustaritz) ont tous un caractère hygrophile comme l’indique la présence constante de Cirsium filipendulum, avec neuf à Molinia caerulea qui est parfois abondant (relevés 10, 11 et 12 du tableau 4), ce qui peut conduire, avec la présence conjuguée d’autres espèces (dans ce cas Laserpitium prutenicum subsp. dufourianum, Serratula tinctoria subsp. seoanei, Dactylorhiza maculata, Scorzonera humilis, Trocdaris verticillatum, Carex panicea), à des hésitations de rattachement des relevés concernés au Cirsio-Molinietum (photo 9).

Cette pelouse se rencontre dans des parcelles exploitées en fauche, souvent sur de vastes surfaces à l’intérieur, et elle est fortement dominée par Pteridium aquilinum à partir de l’été. Nous l’avons aussi observée ponctuellement sur le littoral à Saint-Jean-de-Luz en lisière étroite de sentier située au contact de la lande du Glandoro prostratae-Ericetum vagantis ericetosum cinereae (variante non hygrophile). Les autres contacts notés sont avec le Lathyro nudicaulis-Schoenetum nigricantis typicum présent au niveau inférieur (au moins à quatre reprises sur les communes d’Ahetze, Halsou et Ustaritz) et avec le Cirsio filipenduli-Molinietum caeruleae caretosum verticillati dans sa variation à espèces du Lathyro-Schoenetum (un cas à Ustaritz avec Carex hostiana +, Schoenus nigricans r et Linum catharticum r dans le Cirsio-Molinietum).
Intérêt patrimonial
Lathyrus nudicaulis ne semble présent qu’exceptionnellement dans le Carici piluliferae-Pseudarrhenatheretum longifolii car l’espèce n’apparaît que dans un relevé sur quinze, lequel a été effectué sur le littoral à Hendaye (relevé 3 du tableau 4). Cela s’explique probablement par le caractère trop acidiphile et mésophile de l’association.
Cependant, l’association comporte parfois d’autres espèces patrimoniales (planche 12) telles que Serapias cordigera, exceptionnellement le rarissime Romulea bulbocodium qui n’est connu dans les Pyrénées-Atlantiques que dans une seule commune du littoral (relevés 7 et 8 du tableau 4). Ces espèces sont toutes deux protégés régionalement en Nouvelle-Aquitaine (JORF, 2002). Glandora prostrata, protégé en France (JORF, 1982), apparaît également dans quatre relevés réalisés à Ahetze, Saint-Jean-de-Luz et Urrugne (relevés 1, 9, 10 et 11 du tableau 4). On peut encore citer Tractema umbellata, Gentiana pneumonanthe, Simethis mattiazzii, Laserpitium prutenicum subsp. dufourianum et Viola lactea qui sont inscrits sur la liste des espèces déterminantes ZNIEFF de la flore vasculaire de Nouvelle-Aquitaine (Abadie et al., 2019).

Bien qu’elle ne figure pas dans nos relevés, Erythronium dens-canis est une autre espèce déterminante ZNIEFF (hors Massif pyrénéen) qui peut être rencontrée dans ce type de pelouse en tout début d’année (février-mars). En fin de saison à l’automne (octobre-novembre), après que la pelouse ait été fauchée au cours de la période estivale, c’est Crocus nudiflorus qui y fleurit fréquemment et parfois abondamment. L’espèce ne possède pas de statut particulier dans les Pyrénées-Atlantiques mais c’est une belle sub-pyrénéo-cantabrique eu-atlantique montagnarde (Dupont, 2015).
La pelouse littorale à Galium boreale et Pseudarrhenatherum longifolium (Galio borealis-Pseudarrhenatheretum longifolii)
Synfloristique, physionomie et synchorologie
Ce groupement original est une pelouse littorale landicole à Brachypodium rupestre observée uniquement dans trois sites proches situés sur le plateau des falaises de la Corniche basque à Urrugne (planche 13 ; photo 10), qui est intermédiaire entre l’Euphorbio occidentalis-Silaetum pyrenaici, avec lequel il partage notamment les deux espèces éponymes (Euphorbia flavicoma constant, Silaum silaus var. angustifolium), le pré hygrophile du Lathyro nudicaulis-Schoenetum nigricantis potentilletosum montanae et la pelouse acidiphile du Carici piluliferae-Pseudarrhenatheretum longifolii.


Les espèces de landes y sont fréquentes et abondantes, excepté Calluna vulgaris qui est absent : Erica vagans surtout (V), Glandora prostrata (IV), Erica ciliaris (III), Ulex gallii (III), Ulex europaeus (III), rarement Erica cinerea (I). Il est caractérisé par Euphorbia flavicoma (V), Tractema umbellata (V), qui peut être très abondant au cours de sa floraison printanière en avril, Pseudarrhenatherum longifolium (V), Lathyrus nudicaulis (V), Serratula tinctoria subsp. seoanei (IV), Galium boreale (IV), Asphodelus albus (IV), Glandora prostrata (IV), Erica ciliaris (III), Carex pulicaris (III), Helictochloa marginata (III), Silaum silaus var. angustifolium (III), Agrostis capillaris (III), Hypericum pulchrum (II), Agrostis curtisii (II) et Polygala serpyllifolia (II).
À la fin de l’été puis à l’automne, entre les mois d’août et d’octobre, le groupement est parfois coloré par les belles floraisons successives de Gentiana pneumonanthe, d’Allium ericetorum, puis de Crocus nudiflorus. À l’opposé, au tout début du printemps (mars-avril), ce sont les fleurs de Narcissus gigas qui peuvent y apparaître.
Comparaison synfloristique, synécologie, variations et synsystématique
Le groupement (onze relevés, tableau 5) diffère de l’Euphorbio occidentalis-Silaetum pyrenaici (colonnes synthétiques ES1, ES2 et ESG, tableau 5), auquel nous avons été tentés de le rattacher, à la fois :
- par la présence ou par la fréquence plus marquée d’espèces acidiphiles de la pelouse du Carici piluliferae-Pseudarrhenatheretum longifolii, de l’Agrostion curtisii et des Nardetea strictae (en particulier Pseudarrhenatherum longifolium, Tractema umbellata, Helictochloa marginata, Agrostis curtisii, Agrostis capillaris, Polygala serpyllifolia), d’ourlets acidiphiles des Melampyro pratensis-Holcetea mollis (Asphodelus albus, Hypericum pulchrum, Cruciata glabra), des landes fraîches ibéro-atlantiques à Erica vagans (Glandora prostrata, Erica ciliaris),
- et par celle d’espèces plus hygrophiles des prés hygrophiles du Serratulo seoanei-Molinienion caeruleae et des Molinio caeruleae-Juncetea acutiflori (Serratula tinctoria seoanei et Carex pulicaris, ainsi que Gentiana pneumonanthe non noté dans les relevés présentés).
On y relève aussi l’absence de Pulicaria dysenterica. Il est aussi proche floristiquement du Lathyro nudicaulis-Schoenetum nigricantis potentilletosum montanae (colonne synthétique LS3 du tableau 5), plus hygrophile, avec lequel la distinction n’est pas toujours aisée à faire en ne se basant que sur la présence-absence des espèces (nécessité d’intégrer parfois dans l’analyse l’abondance de certaines espèces et le contexte, comme pour la variante hygrophile à Molinia caerulea du Carici piluliferae-Pseudarrhenatheretum longifolii par rapport au Cirsio filipenduli-Molinietum caeruleae).
Compte-tenu de son originalité, nous proposons de classer ce groupement de pelouse thermo-ombro-atlantique, dont la distribution est réduite au plateau argileux des falaises littorales du Pays basque (Urrugne, la Corniche basque) et s’insérant dans un système intermédiaire entre système basique et système acide (de Foucault, 1984), dans une association végétale nouvelle sous le nom de Galio borealis-Pseudarrhenatheretum longifolii ass. nov. hoc loco (typus nominis hoc loco : relevé 5 = 396186 du tableau 5 hoc loco, publié plus bas). Nous y incluons un relevé de Lazare & Bioret (relevé 11, tableau 2 des auteurs, 2006) attribué initialement par ces derniers à l’Euphorbio occidentalis-Silaetum pyrenaici. La colonne ES1 correspond à la colonne synthétique princeps basée sur les onze relevés de cette dernière association, ES2 également moins un relevé (relevé 11, tableau 2, in Lazare & Bioret, 2006), et ESG aux dix relevés que nous avons rattachés à l’Euphorbio-Silaetum (voir tableau 3).
Typus nominis hoc loco du Galio borealis-Pseudarrhenatheretum longifolii
commune d’Urrugne (64), littoral (la Corniche basque), Kauteren Borda (partie sud), 21 juin 2017, 95%, 100 m2, 27 taxons, relevé réalisé par J. Dufay (tableau 5 hoc loco, relevé 5 = 396186)
- combinaison caractéristique : Pseudarrhenatherum longifolium 3, Galium boreale 2, Helictochloa marginata 2, Agrostis curtisii 2, Erica ciliaris 2, Tractema umbellata 1, Euphorbia flavicoma 1, Serratula tinctoria seoanei +, Glandora prostrata +, Carex pulicaris +, Silaum silaus var. angustifolium +, Polygala serpyllifolia r,
- différentiels de variations : Carex panicea 1, Schoenus nigricans +, Scorzonera humilis +, Dactylorhiza maculata +,
- unités supérieures : Brachypodium rupestre 2, Carex flacca 1, Linum catharticum +, Pimpinella saxifraga r,
- autres taxons : Cirsium filipendulum 1, Danthonia decumbens 2, Potentilla erecta 3, Pteridium aquilinum +, Erica vagans 2, Ulex gallii 2, Centaurea r.
L’association se développe dans des parcelles fauchées pendant l’été (avec exportation) ou gyrobroyées pour un site. Elle présente quelques fines variations : à Leucanthemum crassifolium, Smilax aspera et Schoenus nigricans avec aussi selon les cas Neottia ovata, Lysimachia tenella, Serapias lingua, Anacamptis pyramidalis et Blackstonia perfoliata (relevés 1 et 2 du tableau 5), à Leucanthemum crassifolium, Lathyrus linifolius, Linum usitatissimum subsp. angustifolium, Rhinanthus angustifolius, Pulmonaria longifolia et Solidago virgaurea (relevé 3), à Plantago maritima, Smilax aspera, Rhinanthus angustifolius et Allium ericetorum (relevé 4), à Carex panicea, Scorzonera humilis, Dactylorhiza maculata et Schoenus nigricans (relevé 5), à Sanguisorba officinalis (relevé 6), à Narcissus gigas, Viola lactea ou/et Erica cinerea (relevés 7, 8 et 9 du tableau 5).
Le Galio borealis-Pseudarrhenatheretum longifolii semble relever des mêmes unités synsystématiques que l’Euphorbio occidentalis-Silaetum pyrenaici, c’est-à-dire l’alliance du Potentillo montanae-Brachypodion rupestris Braun-Blanq. 1967 corr. Rivas Mart. et al. 2002, l’ordre des Brometalia erecti W. Koch 1926 et la classe des Festuco valesiacae-Brometea erecti Braun-Blanq. & Tüxen ex Braun-Blanq. 1949. Il pourrait s’agir d’une sous-unité plus hygrophile (mésohygrophile) de l’alliance.
Contacts, synécologie et syndynamisme
Ce type de pelouse a notamment été observé au contact supérieur de l’Euphorbio occidentalis-Silaetum pyrenaici dont il semble dériver en partie par acidification et humidification conjuguées. Si l’acidification se poursuit, il paraît pouvoir évoluer vers le Carici piluliferae-Pseudarrhenatheretum longifolii B. Foucault 2006. Dans des conditions édaphiques plus humides, il est remplacé par le Lathyro nudicaulis-Schoenetum nigricantis potentilletosum montanae et par le Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008, d’abord dans sa variation à présence disséminée d’espèces du Lathyro-Schoenetum puis, si l’acidité est plus marquée, sans ces espèces.
Sur le plan syndynamique, le groupement est susceptible d’évoluer en l’absence de fauche vers la variante hygrophile à Erica ciliaris ou Molinia caerulea de la lande littorale du Glandoro prostratae-Ericetum vagantis Lazare 2017, identifiée sur la Corniche basque bien que nous n’ayions pas pu préciser s’il s’agissait de la « forme humilis » d’Ulex gallii.
Intérêt patrimonial
La pelouse littorale du Galio borealis-Pseudarrhenatheretum longifolii possède un très fort intérêt patrimonial de par son originalité phytocoenotique, sa distribution actuellement connue, qui se réduit à trois sites de la Corniche basque (Urrugne), et par la présence plus ou moins forte de plusieurs espèces patrimoniales déjà citées dans d’autres groupements : Lathyrus nudicaulis (neuf relevés sur onze), avec les espèces protégées réglementairement Galium boreale (sept relevés), Glandora prostrata (sept relevés) et Leucanthemum crassifolium (trois relevés). Sa richesse spécifique, comprise entre 27 et 41 espèces, est élevée (nombre spécifique moyen : 32).
3.4 Les landes littorales à Erica vagans et Lathyrus nudicaulis
Nous n’avons noté Lathyrus nudicaulis dans des landes qu’en contexte littoral. L’espèce y a été observée à la fois :
- dans le Smilaco asperae-Ericetum vagantis Lazare & Bioret in Lazare 2017 lathyretosum nudicaulis Lazare 2017 (relevés 6 à 9 du tableau 6 ; colonne synthétique SET des neuf relevés rapportés à l’association comparée aux colonnes synthétiques SE et GE, qui correspondent respectivement aux colonnes synthétiques princeps du Smilaco-Ericetum vagantis et du Glandoro-Ericetum vagantis),
- dans le Glandoro prostratae-Ericetum vagantis Lazare 2017 (tableau 7 ; seize relevés ; colonne synthétique GET1 des sept relevés rattachés à la sous-association typicum ; colonne synthétique GET2 des neuf relevés rattachés à la sous-association ericetosum cinereae Lazare 2017 ; colonne synthétique globale GET des seize relevés comparée aux colonnes synthétiques princeps GE et SE, respectivement du Glandoro-Ericetum vagantis et du Smilaco-Ericetum vagantis).
Dans ce dernier type de lande (Glandoro-Ericetum vagantis), L. nudicaulis est présent dans six relevés sur seize au total, dont un sur sept de la sous-association typicum (relevé 7, tableau 7) et cinq sur neuf de la variante hygrophile à Molinia caerulea, Erica ciliaris ou Cirsium filipendulum de la sous-association ericetosum cinereae (relevés 9 à 13 du tableau 7). La limite nord absolue de l’espèce actuellement connue sur le littoral basque se trouve au sein de cette variation dans une lande rase située sur une pente prononcée exposée au vent et aux embruns du golf d’Ilbarritz à Bidart (photo 11), où Schoenus nigricans est également présent (relevé 10 du tableau 7).

Comme nous l’avons déjà évoqué, nous ne pensons pas que les associations du Smilaco asperae-Ericetum vagantis et du Glandoro prostratae-Ericetum vagantis (planche 14) s’excluent géographiquement, contrairement à ce qu’indique Lazare (2017), et nous les avons notées toutes les deux à Urrugne au sud et à Bidart au nord. À l’instar des pelouses et des prés hygrophiles, il est possible d’imaginer un phénomène d’acidification progressive qui conduit de la première à la seconde.

Au niveau dynamique, sur la Corniche basque (Urrugne), on peut émettre l’hypothèse que le Smilaco-Ericetum vagantis est plus fréquemment associé à la pelouse de l’Euphorbio occidentalis-Silaetum pyrenaici et que le Glandoro-Ericetum vagantis l’est à celle du Galio borealis-Pseudarrhenatheretum longifolii et aux prés hygrophiles (Lathyro nudicaulis-Schoenetum nigricantis potentilletosum montanae, Cirsio filipenduli-Molinietum caeruleae).
Enfin, rappelons que le Leucanthemo crassifolii-Ericetum vagantis (P. Allorge & Jovet 1941) Géhu & Géhu-Franck 1981 (photo 12) nous paraît bien exister. Il se trouve là où il a été défini initialement, c’est-à-dire à la limite supérieure des pentes souvent très accentuées des falaises littorales d’Hendaye, d’Urrugne et de Saint-Jean-de-Luz. Il y est comme suspendu au-dessus de l’océan, « accrochée aux pentes des falaises jusqu’au rebord du plateau sommital » (Géhu & Géhu-Franck, 1981), en situation de télescopage entre les landes du plateau décrites par Lazare (2017) et les pelouses des pentes. Ces dernières relèvent du Leucanthemo crassifolii-Helichrysetum stoechadis (P. Allorge & Jovet 1941) Géhu & Géhu-Franck 1981 et du Leucanthemo crassifolii-Festucetum pruinosae Géhu & Géhu-Franck 1981 corr. Prieto & Loidi 1984, avec plus bas le Crithmo maritimi-Plantaginetum maritimae (Pavillard 1928) Guinea 1949.

3.5 Bilan
Les groupements végétaux à Lathyrus nudicaulis
Le tableau 8 présente les colonnes synthétiques de l’ensemble des groupements végétaux dans lesquels Lathyrus nudicaulis a été observé au Pays basque occidental (province du Labourd) :
- pré hygrophile du Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 ined. (colonne 1, CJB, des six relevés princeps de la race définie par de Foucault, 1984 ; colonne 2, CJTO, de nos trois relevés, dont deux à Lathyrus nudicaulis) ;
- pré hygrophile du groupement à Carex echinata et Schoenus nigricans (colonne 3, CJTI, trois relevés, tous à Lathyrus nudicaulis) ;
- pré hygrophile (mésohygrophile à mésophile) basiclinophile du Lathyro nudicaulis-Schoenetum nigricantis (colonne 4, LS, 32 relevés ; présence constante de Lathyrus nudicaulis dans les 29 relevés récents, et présence probable dans les trois relevés d’origine bibliographique effectués par de Foucault, 1984), sous-associations typicum (colonne 5, LS1, dix-sept relevés), trocdarietosum verticillati (colonne 6, LS2, quatre relevés) et potentilletosum montanae (colonne 7, LS3, onze relevés) ;
- prés atypiques, sans Schoenus nigricans, du Lathyro-Schoenetum (colonne 8, LSA, trois relevés, tous à Lathyrus nudicaulis) ;
- pelouse littorale neutro-acidiclinophile et thermophile de l’Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006 (colonne 9, ES1, onze relevés initiaux de Lazare & Bioret, 2006 ; colonne 10, ES2, relevés initiaux de Lazare & Bioret moins un relevé rattaché au Galio borealis-Pseudarrhenatheretum longifolii ; colonne 11, ESG, dix relevés récents rattachés à l’association ou proches de cette dernière, dont huit à Lathyrus nudicaulis);
- pelouse littorale neutro-acidiclinophile et hygroclinophile du Galio borealis-Pseudarrhenatheretum longifolii (colonne 12, GP, onze relevés, dont un d’origine bibliographique extrait de l’Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006 ; présence de Lathyrus nudicaulis dans neuf relevés) ;
- pré hygrophile (mésohygrophile à mésophile) acidiclinophile ou acidiphile du Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008 (colonne 13, CMG1, vingt-deux relevés princeps, de Foucault, 1984 ; colonne 14, CMG2, dix-neuf relevés, vingt-deux relevés princeps moins trois relevés de la variante à Schoenus nigricans intégrés au Lathyro nudicaulis-Schoenetum nigricantis ; colonne 15, CMT2, quatorze relevés, sous-association typicum, dix-sept relevés princeps moins trois relevés de la variante à Schoenus nigricans intégrés au Lathyro-Schoenetum ; colonne 16, CMCA1, cinq relevés princeps de la sous-association caretosum verticillati Foucault 2008, de Foucault, 1984 ; colonne 17, CMT, six relevés récents rattachés à l’association dans des variations à espèces du Lathyro-Schoenetum, dont cinq au caretosum verticillati et un au typicum, avec présence de Lathyrus nudicaulis dans trois relevés de la sous-association caretosum) ;
- pelouse mésophile acidiclinophile ou acidiphile du Carici piluliferae-Pseudarrhenatheretum longifolii Foucault 1986 (colonne 18, CP, trente-trois relevés princeps de l’association, de Foucault, 1986b, tableau 1 de l’auteur ; colonne 19, CPBP, dix-sept relevés princeps de la sous-association brachypodietosum pinnati B. Foucault 1986 ; colonne 20, CPT, seize relevés princeps de la sous-association typicum; colonne 21, CPTB, six relevés récents rattachés au brachypodietosum pinnati, dont un à Lathyrus nudicaulis ; colonne 22, CPTT, neuf relevés récents du typicum ; colonne 23, CPTG, synthèse des quinze relevés récents) ;
- lande littorale neutro-basiclinophile à acidiclinophile du Glandoro prostratae-Ericetum vagantis Lazare 2017 (colonne 25, GE, douze relevés princeps de l’association, Lazare, 2017, tableau V de l’auteur ; colonne 26, GET, seize relevés rattachés à l’association dont six à Lathyrus nudicaulis) ;
- lande littorale basiphile du Smilaco asperae-Ericetum vagantis Lazare & Bioret in Lazare 2017 (colonne 27, SE, dix-huit relevés princeps de l’association dont dix à Lathyrus nudicaulis, Lazare, 2017, tableau I de l’auteur ; colonne 28, SET, neuf relevés rattachés à l’association dont quatre à Lathyrus nudicaulis).
La colonne synthétique du groupement à Brachypodium pinnatum et Pseudarrhenatherum longifolium B. Foucault 1986 (colonne 24, GBBP : cinq relevés princeps, de Foucault, 1986b, tableau 3 de l’auteur), plus neutrophile que le Carici-Pseudarrhenatheretum (de Foucault, 2012), non observé mais que l’on aurait imaginé trouver à l’intérieur au contact supérieur du Lathyro-Schoenetum à la place du Carici-Pseudarrhenatheretum, a également été insérée au tableau 8 afin d’avoir une vision plus globale des végétations.
Dans la même optique, le lecteur est invité à consulter plusieurs travaux phytosociologiques réalisés sur les prairies mésophiles de fauche à Linum usitatissimum subsp. angustifolium et Oenanthe pimpinelloides avec parfois Trifolium patens (en particulier sur le Lino biennis-Cynosuretum cristati P. Allorge ex Tüxen & Oberd. 1958 et sur le Lino angustifolii-Brometum hordeacei B. Foucault 2016 ; de Foucault, 1986c, 2016), assez fréquentes dans la zone d’étude (photo 13), et qui semblent constituer la suite de séries trophiques d’eutrophisation de certaines associations présentées (Carici-Pseudarrhenatheretum, Euphorbio-Silaetum), peut-être dans certains cas par l’intermédiaire de pelouses à Rhinanthus angustifolius et espèces prairiales qui seraient à étudier en détail. Nous n’y avons pas noté Lathyrus nudicaulis.

Discussion phytoécologique
Ce qui revient constamment dans les essais et dans les difficultés d’interprétation phytoécologique des groupements végétaux présentés, c’est l’originalité d’appartenance supposée d’une grande partie d’entre eux à un système intermédiaire entre système acide et système basique tel que défini par de Foucault (1984), à laquelle se superposent des questions d’humidité, puis de situation littorale ou intérieure. Au niveau trophique, ils semblent pouvoir être qualifiés d’oligotrophiles, établis pour ceux dont nous avons examiné la partie superficielle des sols sur des terrains argileux compacts.
La première grande séparation nette entre groupements clairement hygrophiles (Caro verticillati-Juncetum acutiflori, groupement à Carex echinata et Schoenus nigricans) et groupements plus mésophiles par rapport à ces derniers (tous les autres), qui d’ailleurs avait été faite par le traitement statistique initial de nos relevés, ne pose pas de problèmes d’interprétation et de qualification en ce qui concerne le facteur humidité. Le caractère interprété comme acidiclinophile ou acidiphile du Carici piluliferae-Pseudarrhenatheretum longifolii et du Cirsio filipenduli-Molinietum caeruleae semble également crédible.
Par contre, la conjugaison d’acidité et de basicité, d’après l’écologie théorique des espèces constitutives des cortèges floristiques, est difficile à cerner, à qualifier et à expliquer dans le Caro verticillati-Juncetum acutiflori race basque (à Carex hostiana surtout), le groupement à Carex echinata et Schoenus nigricans, le Lathyro nudicaulis-Schoenetum nigricantis et le Galio borealis-Pseudarrhenatheretum longifolii. Pour la pelouse littorale de l’Euphorbio occidentalis-Silaetum pyrenaici (Corniche basque à Urrugne et son prolongement à Hendaye), Lazare et Bioret (2006) l’expliquent par une « acidification superficielle du sol due au lessivage des cations par les précipitations » de cette association « développée sur substrat carbonaté (bancs calcaires du flysch sénonien à maestrichtien) ». On peut imaginer que le même phénomène se produise avec le Galio borealis-Pseudarrhenatheretum longifolii qui se trouve dans le même secteur et en partie au contact de la précédente association sur la Corniche, conduisant ensuite au Carici piluliferae-Pseudarrhenatheretum longifolii si l’acidification se poursuit. Mais est-ce qu’il intervient aussi dans des conditions plus humides et dans ce sens avec une évolution graduelle du Lathyro nudicaulis-Schoenetum nigricantis (potentilletosum montanae sur le littoral) vers le Cirsio filipenduli-Molinietum caeruleae, ou dans le sens inverse d’une alcalinisation progressive du Cirsio-Molinietum par accumulation des cations issus des lessivages précédents ?
Le même type de question se pose à l’intérieur, où nous ne comprenons pas les facteurs écologiques qui expliquent en détail la présence ou l’absence de tel ou tel groupement et les processus d’évolution de l’un vers l’autre. En plus du Carici-Pseudarrhenatheretum fréquemment observé, nous aurions dû trouver des groupements de pelouses mésophiles de système intermédiaire homologues de celles du littoral (avec peut-être Lathyrus nudicaulis), relayés en conditions plus humides par le Lathyro-Schoenetum ou par le Cirsio-Molinietum selon le niveau d’acidité, puis par le Caro-Juncetum race basque ou par le groupement à Carex echinata et Schoenus nigricans.
Résumé et discussion synsystématiques
La position synsystématique actuelle des groupements végétaux à Lathyrus nudicaulis du Pays basque occidental est donnée ci-après :
Molinio caeruleae-Juncetea acutiflori Braun-Blanq. 1950
Molinietalia caeruleae W. Koch 1926
Juncion acutiflori Braun-Blanq. in Braun-Blanq. & Tüxen 1952
Caro verticillati-Juncenion acutiflori B. Foucault & Géhu 1980
Caro verticillati-Juncetum acutiflori (G. Lemée 1937) Korneck 1962 race basque B. Foucault 1984 nom. ined.
Groupement à Carex echinata et Schoenus nigricans
Juncion acutiflori ou Molinion caeruleae W. Koch 1926
Serratulo seoanei-Molinienion caeruleae B. Foucault 2008
Lathyro nudicaulis-Schoenetum nigricantis nov.
typicum
trocdarietosum verticillati nov.
potentilletosum montanae nov.
Cirsio filipenduli-Molinietum caeruleae (P. Allorge 1941) B. Foucault 2008
caretosum verticillati B. Foucault 2008
Nardetea strictae Rivas Goday in Rivas Goday & Rivas Mart. 1963
Nardetalia strictae Oberd. ex Preising 1950
Agrostion curtisii B. Foucault 1986
Carici piluliferae-Pseudarrhenatheretum longifolii (P. Allorge 1941) B. Foucault 1986
brachypodietosum pinnati B. Foucault 1986 variante hygrophile
Festuco valesiacae-Brometea erecti Braun-Blanq. & Tüxen ex Klika & Hadač 1944
Brometalia erecti W. Koch 1926
Potentillo montanae-Brachypodion rupestris Braun-Blanq. 1967 corr. Rivas Mart. et al. 2002
Euphorbio occidentalis-Silaetum pyrenaici Lazare & Bioret 2006
Galio borealis-Pseudarrhenatheretum longifolii nov.
Calluno vulgaris-Ulicetea minoris Braun-Blanq. & Tüxen ex Klika in Klika & Hadač 1944
Ulicetalia minoris Quantin 1935
Dactylido oceanicae-Ulicion minoris Géhu 1975
Smilaco asperae-Ericetum vagantis Lazare & Bioret in Lazare 2017
lathyretosum nudicaulis Lazare 2017
Glandoro prostratae-Ericetum vagantis Lazare 2017
ericetosum cinereae Lazare 2017
variante non hygrophile
variante hygrophile Lazare 2017
Au sein des Molinio-Juncetea, il manque peut-être une alliance qui rassemblerait des groupements végétaux de systèmes intermédiaires entre systèmes acides et systèmes alcalins, intermédiaire entre le Juncion acutiflori et le Molinion, dans laquelle se placerait au moins une partie des associations du Serratulo-Molinienion (en particulier le Lathyro-Schoenetum) et des variations basiphiles d’autres associations du Juncion, mais ce qui impliquerait pour ces dernières d’élever leur rang synsystématique. Par ailleurs pour le Galio borealis-Pseudarrhenatheretum longifolii, que nous avons classé dans le Potentillo montanae-Brachypodion rupestris et les Festuco-Brometea, il pourrait s’agir d’une unité plus hygrophile (mésohygrophile) de cette alliance ou d’une autre alliance homologue au Pays basque du Tetragonolobo maritimi-Bromenion erecti J.-M. Royer et al. 2006.
4. Conclusion
L’intérêt patrimonial de Lathyrus nudicaulis (photo 14), et des communautés végétales dans lesquelles l’espèce se rencontre au Pays basque occidental (Labourd), est particulièrement fort. L’espèce est rare à l’échelle mondiale, puis au sein de son aire ibérique où des études complémentaires seraient par ailleurs à entreprendre pour préciser sa distinction avec Lathyrus palustris surtout, et avec Lathyrus linifolius var. linifolius, en particulier sur les plans génétique et phytosociologique. Malgré la découverte récente de plusieurs stations situées plus en retrait du littoral en France, elle n’en demeure pas moins une espèce rare inféodée à des groupements végétaux très originaux, dont certains sont peut-être synendémiques du Pays basque nord (France) et sud (Espagne).

L’ajout de Lathyrus nudicaulis à une liste d’espèces protégées réglementairement « pour prévenir la disparition d’espèces végétales menacées et permettre la conservation des biotopes correspondants » (JORF, 1982), serait une première étape minimale pour assurer sa conservation et celle des végétations auxquelles elle est associée.

Bibliographie
Abadie J.-C., Nawrot O., Vial T., Caze G. & Hamdi E., 2019. Liste des espèces déterminantes ZNIEFF de la flore vasculaire de Nouvelle-Aquitaine. Conservatoire botanique national (CBN) Sud-Atlantique, CBN Massif central, CBN Pyrénées et de Midi-Pyrénées, Audenge, Chavaniac-Lafayette, Bagnères-de-Bigorre, 108 p + annexes.
Aizpuru I., Aseginolaza C., Uribe-Echebarría P.M., Urrutia P. & Zorrakin I. (eds.), 1999. Claves ilustradas de la Flora del País Vasco y territorios limítrofes. Gobierno Vasco, Instituto Alavés de la Naturaleza, Sociedad de Ciencias Aranzadi, Vitoria-Gasteiz, 831 p.
Alejandre Sáenz J.A., García-López J.M. & Mateo Sanz G. (eds.), 2006. Atlas de la flora vascular silvestre de Burgos. Junta de Castilla y León, Caja Rural de Burgos, Burgos, 924 p.
Anonyme, 2007. Interpretation Manual of European Union Habitats. EUR 27. European Commission (DG Environment, Nature and biodiversity), 142 p.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Muséum national d’Histoire naturelle (Patrimoines naturels 61), Paris, 171 p.
Bensettiti F., Boullet V., Chavaudret-Laborie C. & Deniaud J. (eds.), 2005. « Cahiers d’habitats » Natura 2000. Connaissance et gestion des habitats et des espèces d’intérêt communautaire. 4 – Habitats agropastoraux. MEDD, MAAPAR, MNHN (Ed. La Documentation française), Paris, 2 volumes : 445 p. et 487 p.
Berastegi Gartziandia A., 2013. Prados y pastizales en Navarra : descripción, tipificación y ecología. Guineana 19 : 1-505.
Braun-Blanquet J., 1967. Vegetationsskizzen aus dem Baskenland mit Ausblicken auf das weitere Ibero-Atlantikum, II Teil. Vegetatio 14 (1-4) : 1-126.
Cabezudo B. (coord.), Talavera S. (coord.), Blanca G., Salazar C., Cueto M., Valdés B., Hernández Bermejo J.E., Herrera C.M., Rodríguez Hiraldo C. & Navas D. (eds.), 2005. Lista roja de la flora vascular de Andalucía. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla, 126 p.
Carapeto A., Pinto da Silva P., Porto M. & Cardoso P.E., 2014. Estudo de caracterização da flora e vegetação da Reserva local do sapal do rio Coina e Mata Nacional da Machada – Barreiro. Sociedade Portuguesa de Botânica, 137 p.
Carapeto A., Francisco A., Pereira P. & Porto M. (eds.), 2020. Lista Vermhela da Flora Vascular de Portugal Continental. Sociedade Portuguesa de Botânica, Associação Portuguesa de Ciência da Vegetação – PHYTOS, Instituto da Conservação de Natureza e das Florestas (coord.), Coleção « Botânica em Português » 7, Lisboa, 374 p.
Carlón L., González del Valle J.M., Laínz M., Moreno Moral G., Rodríguez Berdasco J.M. & Sánchez Pedraja Ó., 2010. Contribuciones al conocimiento de la flora cántabrica, VIII. Documentos del Jardín Botánico Atlántico (Gijón) 7 : 1-95.
Catalán P. & Aizpuru I., 1985. Aportación al catálogo florístico de la cuenca del Bidasoa (Guipúzcoa y Navarra). Munibe Ciencias Naturales 37 : 17-86.
Clemente A., Lemos P., Carapeto A., Alves P., Duarte S., Aguiar C., Clamote F. & Porto M. et al., 2021. Lathyrus nudicaulis (Willk.) Amo – mapa de distribuição. Sociedade Portuguesa de Botânica, http://www.flora-on.pt
Costa J.C., Neto C., Aguiar C., Capelo J., Espírito Santo M.D., Honrado J., Pinto-Gomes C., Monteiro-Henriques T., Sequeira M. & Lousã M., 2012. Vascular plant communities in Portugal (Continental, Azores and Madeira). Global Geobotany 2 : 1-180.
Coulot P. & Rabaute Ph., 2016. Monographie des Leguminosae de France, 4 – Tribus des Fabeae, des Cicereae et des Genisteae. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 46 : 1-902.
Coutinho A.X.P., 1913. A Flora de Portugal (Plantas vasculares) disposta em chaves dichotomicas. Eds. Aillaud, Alves & Cia, Francisco Alves & Cia, Paris, Rio de Janeiro, 766 p.
Díaz González T.E., Fernández Prieto J.A. & Nava Fernández H.S., 1994. Catálogo de la flora vascular de Asturias. Itinera Geobotanica 8 : 529-600.
Dufay J., Hardy F. & Meslage N., 2018. Observatoire de la biodiversité végétale du littoral des Pyrénées-Atlantiques. Bilan des travaux menés en 2017. Rapport général. Conservatoire botanique national Sud-Atlantique, Audenge (Saint-Jean-de-Luz), 81 p.
Dupont P., 2015. Les plantes vasculaires atlantiques, les pyrénéo-cantabriques et les éléments floristiques voisins dans la péninsule ibérique et en France. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 45 : 1-494.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’État, Universités de Rouen et de Lille II, SIP Bailleul, 674 p. (2 tomes) + 248 tableaux.
Foucault B. (de), 1986a. Petit manuel d’initiation à la phytosociologie sigmatiste. Société linnéenne du nord de la France (Amiens), Laboratoire de botanique de la Faculté de Pharmacie (Lille II), 47 p.
Foucault B. (de), 1986b. Données systémiques sur la végétation prairiale mésophile du Pays basque et des Landes de Gascogne. Documents phytosociologiques, NS, X : 203-219.
Foucault B. (de), 1986c. Contribution à une étude phytosociologique des systèmes prairiaux hygrophile et mésophile de l’Armagnac méridional (Hautes-Pyrénées et Gers, France). Documents phytosociologiques, NS, X : 221-254.
Foucault B. (de), 2008. Validation nomenclaturale de syntaxons inédits ou invalides. Journal de botanique de la Société botanique de France 43 : 43-61.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Nardetea strictae Rivas Goday in Rivas Goday & Rivas-Mart. 1963. Journal de botanique de la Société botanique de France 59 : 241-344.
Foucault B. (de), 2016. Contribution au prodrome des végétations de France : les Arrhenatheretea elatioris Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952. Documents phytosociologiques, série 3, 3 : 5-217.
Foucault B. (de) & Catteau E., 2012. Contribution au prodrome des végétations de France : les Agrostietea stoloniferae Oberd. 1983. Journal de botanique de la Société botanique de France 59 : 5-131.
Gallego M.J., 1999. Lathyrus L. In S. Talavera, C. Aedo, S. Castroviejo, C. Romero Zarco, L. Sáez, F.J. Salgueiro & M. Velayos (eds.), Flora iberica, Plantas vasculares de la Península Ibérica e Islas Baleares. Leguminosae (partim), Real Jardín Botánico, CSIC, Madrid, VII (I) : 423-482.
García-Mijangos I., 1997. Flora y vegetación de los Montes Obarenes (Burgos). Guineana 3 : 1-457.
Garrido B., Aparicio A., Pérez Porras C., Aparicio J., García Martín F., Fernández Carrillo L. & Carrasco M.A., 2002. Contribuciones a la flora vascular de Andalucía (España), Flora de interés en bosques-isla de Andalucía occidental. Acta Botanica Malacitana 27 : 295-308.
Gatignol P., 2007. Découverte d’une nouvelle espèce pour la France : Lathyrus nudicaulis (Willk.) Amo sur la Corniche basque (Pyrénées-Atlantiques). Le Monde des plantes 492 : 30-31.
Géhu J.-M. & Géhu-Franck J., 1981. Aperçu phytosociologique sur les falaises d’Hendaye et de Saint-Jean-de-Luz (Pays basque). Documents phytosociologiques, NS, V : 363-394.
Géhu J.-M. & Rivas-Martínez S., 1981. Notions fondamentales de phytosociologie. Berichte der Internationalen Symposien Vereinigung für Vegetationskunde, Syntaxonomie, (Rinteln 31.3-3.4.1980). Ed. J. Cramer, Vaduz : 5-33.
Gómez D., Lorda M., Font X., García M.B. & Aizpuru I., 2020. Rareza en la flora pirenaica. Materiales preliminares para una lista roja. Munibe Monographs. Nature Series 4 : 93-100.
Guinochet M., 1973. Phytosociologie. Collection d’écologie 1, Masson & Cie, Paris, 227 p.
Herrera M., 1995. Estudio de la vegetación y flora vascular de la cuenca del río Asón (Cantabria). Guineana 1 : 9-435.
JORF, 1982. Arrêté du 20 janvier 1982 fixant la liste des espèces végétales protégées sur l’ensemble du territoire national pour prévenir la disparition d’espèces végétales menacées et de permettre la conservation des biotopes correspondants. Journal officiel de la République française (JORF) n°111 (N.C.), du 13 mai 1982.
JORF, 2002. Arrêté du 8 mars 2002 relatif à la liste des espèces végétales protégées en région Aquitaine complétant la liste nationale. Journal officiel de la République française (JORF) n°104 du 4 mai 2002.
Julve Ph., 1993. Synopsis phytosociologique de la France (communautés de plantes vasculaires). Lejeunia, NS, 140 : 1-160. [index des syntaxons et des taxons : Roux G., 1996, inédit]
Lafon P. (coord.), Mady M., Corriol G. & Belaud A., 2020. Catalogue des végétations de Nouvelle-Aquitaine. Classification, chorologie et correspondances avec les habitats européens. Conservatoire botanique national (CBN) Sud-Atlantique, CBN Massif central, CBN Pyrénées et de Midi-Pyrénées, Audenge, Chavaniac-Lafayette, Bagnères-de-Bigorre, 272 p.
Lazare J.-J., 2017. Contribution à l’étude phytosociologique et symphytosociologique du littoral aquitain (France). Botanique 2 : 41-71.
Lazare J.-J. & Bioret F., 2006. Associations végétales nouvelles du littoral du Pays basque. Journal de botanique de la Société botanique de France 34 : 71-80.
Loidi J., Berastegui A. & García-Mijangos I., 1996. Notas nomenclaturales sobre la vegetación del norte de la Península Ibérica, V. Lazaroa 17 : 141-146.
Loidi J., Fernández Prieto J.A., Herrera M. & Bueno Á., 2014. La vegetación de la comarca burgalesa de Espinosa de los Monteros. Guineana 20 : 1-139.
Meslage N., 2018. Observatoire de la biodiversité végétale du littoral des Pyrénées-Atlantiques. Bilan des travaux menés en 2017. Fiches espèces patrimoniales. Conservatoire botanique national Sud-Atlantique, Audenge (Saint-Jean-de-Luz), 71 p.
Meslage N. & Hardy F., 2018. Observatoire de la biodiversité végétale du littoral des Pyrénées-Atlantiques. Bilan des travaux menés en 2017. Fiches sites à enjeux. Conservatoire botanique national Sud-Atlantique, Audenge (Saint-Jean-de-Luz), 247 p.
Nava H.S., Fernández Casado M.A., Zabaleta I. & Martino J., 1990. Nota floristica asturiana. Boletín de Ciencias de la Naturaleza. Real Instituto de Estudios Asturianos 40 : 127-131.
Rivas-Martínez S., Costa M., Castroviejo S. & Valdés E., 1980. La vegetación de Doñana (Huelva, España). Lazaroa 2 : 5-189.
Rivas-Martínez S., Loidi J., Cantó P., Sancho L. G. & Sánchez-Mata D., 1984. Datos sobre la vegetación del valle del río Bidasoa (España). Lazaroa 6 : 127-150.
Rivas-Martínez S., Fernández-González F., Loidi J., Lousã M. & Penas A., 2001. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica 14 : 5-341.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal. Addenda to the syntaxonomical checklist of 2001. Itinera Geobotanica 15 (I) : 5-432, 15 (II) : 433-922.
Romão C., 1997. Manuel d’interprétation des habitats de l’Union européenne, version EUR 15. Conseil de l’Europe, Bruxelles, 109 p.
Royer J.-M., Felzines J.-C., Misset C. & Thévenin S., 2006. Synopsis commenté des groupements végétaux de la Bourgogne et de la Champagne-Ardenne. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 25 : 369 p.
Sáenz C., 1980. Polen de la flora de Doñana (Huelva, España). Lazaroa 2 : 191-270.
Tichý L., 2002. Juice, software for vegetation classification. Journal of Vegetation Science 13 : 451-453.
Tison J.-M. & de Foucault B. (coords), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.
Tutin T.G., Heywood V.H., Burges N.A., Moore D.M., Valentine D.H., Walters S.M. & Webb D.A. (eds.), 1968. Flora Europaea. 2 – Rosaceae to Umbelliferae. Cambridge University Press, Cambridge, XXVII-469 p.
Valdés B., Talavera S. & Fernández-Galiano E. (eds.), 1987. Flora Vascular de Andalucía Occidental 2. Ed. Ketres, Barcelona, 640 p.
Valdés B., Girón V., Sánchez Gullón E. & Carmona I., 2007. Catálogo florístico del espacio natural de Doñana (SO de España). Plantas vasculares. Lagascalia 27 : 73-362.
Valdés B., Girón V., Sánchez Gullón E. & Carmona I., 2010. Guía de las especies de interés de la flora de Doñana y su Comarca. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla, 182 p.
Willkomm M., 1852. Sertum Florae Hispanicae. Leipzig, 173 p.
Sites internet
Associação Portuguesa de Ciência de Vegetação (PHYTOS). https://www.phytosassociation.com/
Conservatoire botanique national (CBN) Sud-Atlantique, CBN Massif central, CBN Pyrénées et de Midi-Pyrénées. Observatoire de la biodiversité végétale de Nouvelle-Aquitaine. Inventaire du Patrimoine Naturel (IPN). Flore – Fonge – Habitats. https://obv-na.fr/
Fédération des conservatoires botaniques nationaux (FCBN). SI Flore. http://siflore.fcbn.fr/
GBIF. Global Biodiversity Information Facility (Système mondial d’information sur la biodiversité). Accès libre et ouvert aux données sur la biodiversité. https://www.gbif.org/fr/
Gobierno de España (Ministerio de Agricultura, Alimentación y Medio Ambiente), Fundación Biodiversidad & Consejo Superior de Investigaciones Científicas (CSIC, Real Jardín Botánico). Anthos : Sistema de información sobre las plantas de España. http://www.anthos.es/
Muséum national d’histoire naturelle (MNHN), Office français de la biodiversité (OFB). Inventaire national du patrimoine naturel (INPN). https://inpn.mnhn.fr/
Observatorio pirenaico del cambio clímatico. Atlas de la Flora de los Pirineos. Proyecto POCTEFA. http://www.florapyrenaea.com/florapyrenaea/
Real Jardín Botánico (Consejo Superior de Investigaciones Científicas). Flora iberica : Plantas vasculares de la Península Ibérica e Islas Baleares. http://www.anthos.es/
Sociedade Portuguesa de Botânica. Flora-On : Flora de Portugal Interactiva. https://flora-on.pt/
Tela Botanica. Le réseau des botanistes francophones. https://www.tela-botanica.org/
Remerciements
Nous tenons à remercier toutes les personnes qui ont contribué de manière directe ou indirecte à la réalisation de ce travail : Bruno de Foucault, Pascal Lacroix et Patrick Gatignol (relecture de l’article), Pascal Clerc, Josselin Dufay, Jean-Marie Dupont, Marion Garnier, Ganix Grabières, Rémi Guisier, Peio Lambert, Paule Lamouroux et France Rossmann (contributions aux relevés phytosociologiques), Marc Castera, Grégory Caze, François Esnault, Yorick Ferrez, Henri Michaud, Jean-Pierre Roux, Fabienne Lissardy, Armelle Maurice, Jon Sudupe, Nicolas Van Meer Hargain et Anne-Elizabeth Wolf (contributions diverses).
Nos remerciements s’adressent également aux partenaires financiers du Conservatoire botanique national Sud-Atlantique (CBNSA) pour leur soutien aux différentes études réalisées entre 2017 et 2019 qui ont notamment permis d’améliorer les connaissances sur Lathyrus nudicaulis : Département des Pyrénées-Atlantiques, Ville de Saint-Jean-de-Luz (Jardin botanique littoral Paul-Jovet), DREAL Nouvelle-Aquitaine, Région Nouvelle-Aquitaine, Union européenne, Communauté d’agglomération Pays basque.
Avec nos pensées pour Pierre Dupont (1925-2017) et pour Jean Guittet (1939-2013).


