
Contribution à une synthèse des Lygeo sparti-Macrochloetea tenacissimae
Title
Contribution to an overview of the Lygeo sparti-Macrochloetea tenacissimae class
Résumé
Sur la base d’un tableau synthétique et de tableaux par alliance, les auteurs proposent une ordination des steppes à Lygeum spartum et Macrochloa tenacissima (= Stipa tenacissima), avec trois ordres (dont un nouveau) et seize alliances (dont quatre nouvelles).
Abstract
On the basis of a synthetic table and tables by alliance, the authors propose an overview of the steppes with Lygeum spartum and Macrochloa tenacissima (= Stipa tenacissima), with three orders (of which one is new) and sixteen alliances (of which four are new).
1. Introduction
Les steppes vivaces nord-sahariennes et ibériques à Lygeum spartum et Macrochloa tenacissima (= Stipa tenacissima) ont été rassemblées dans les Lygeo-Stipetea tenacissimae par Rivas-Martínez (1978). Dans le prodrome de Bardat et al. (2004), cette classe a été étendue à la France où elle serait représentée par les Lygeo sparti-Stipetalia tenacissimae Braun-Blanq. & O. Bolòs 1958 (avec une alliance), les Hyparrhenietalia hirtae Rivas Mart. 1978 (une alliance) et les Brachypodietalia phoenicoidis Braun-Blanq. ex Molin. 1934 (une alliance). Plus récemment (de Foucault & Jasprica, 2019), ont été mis en évidence des phénomènes d’eutrophisation/perturbation dans les pelouses et garrigues méditerranéennes qui provoquent par suite un grand développement de taxons rudéraux thermophiles, dont Dittrichia viscosa ; la conséquence en a été la reconnaissance d’une classe méditerranéenne méconnue, les Dittrichietea viscosae Trinajstić, B. Foucault & Jasprica 2019, laquelle inclut au moins une partie des Brachypodietalia phoenicoidis, d’ailleurs déjà exclus des Lygeo- Stipetea par Rivas-Martínez (1978). Pour vérifier la présence et le contenu du reste des Lygeo-Stipetea dans notre pays, nous en proposons ici une synthèse selon des principes phytosociologiques modernes. Elle s’articule avec celle des Rosmarinetea officinalis (de Foucault, 2020) et vient compléter une seconde synthèse antérieure sur d’autres steppes nord-africaines (de Foucault & Lazare, 2020).
2. Méthodes et conventions
Depuis les travaux historiques fondamentaux de Guinochet (1951), Lemée (1953) et Quézel (1954, 1965) sur les steppes sahariennes, la méthodologie de description de la végétation s’est bien affinée. Déjà en 1973, puis en 1978, Guinochet reconnaît la nécessité de bien distinguer phytocénoses, individus d’association (IA) et stations. L’un de nous a discuté le choix de séparer des IA vivaces et des IA thérophytiques dans les phytocénoses biologiquement complexes (de Foucault 1993, 1999, 2013). Outre les stratégies adaptatives différentes entre les deux formes biologiques, ces deux types d’IA ont généralement des phénologies asynchones dans une même phytocénose complexe. Ces idées sont émises et/ou suivies indépendamment par Nègre (1978 : 30), Kaabèche (1990, 1995, 1996), Djebaili (1990), Aidoud-Lounis et al. (1999), de Foucault & Lazare (2020). C’est aussi le choix que nous suivrons ici, ce qui nous amènera à redéfinir des unités originales. C’est d’ailleurs par la volonté de clarifier la synsystématique des pelouses méditerranéennes en distinguant des communautés thérophytiques autonomes rassemblées au rang de classe et des communautés vivaces que Rivas-Martínez (1978) en vient à proposer la classe des Lygeo-Stipetea. Le choix de relever ou non les thérophytes, naturellement omniprésents dans les ouvertures des pelouses vivaces méditerranéennes, n’est donc pas en soit un débat fondamental si l’on s’accorde sur le fait que c’est bien la composante vivace de la phytocénose qui doit servir de support à la description et la classification. L’inverse bien entendu s’applique aux communautés annuelles.
L’ordination hiérarchique des classes (tableau 1) est déduite de tableaux synthétiques (tableaux 2 à 12), dont les sources sont données dans le texte ci-après. Les noms des associations biologiquement complexes sont placés entre guillemets ; conformément à nos choix méthodologiques, nous en extrairons et validerons parfois des associations biologiquement plus homogènes.
La nomenclature taxonomique suit Flora Gallica (Tison & de Foucault, 2014) pour les taxons présents en France (sauf Stipella parviflora nommé ici Stipellula parviflora) et Euro+Med PlantBase pour les autres. Les noms des sous-espèces ou variétés autonymes (c’est-à-dire portant la même épithète que l’espèce à laquelle elles se rattachent) seront réduits à leur initiale (exemple : Daucus carota subsp. c.) ; le signe * remplace ‘subsp.’ et ‘var.’ dans les tableaux phytosociologiques.
3. Déclinaison de la classe
Le tableau synthétique 1 met en évidence trois ordres et seize unités du rang alliance floristiquement bien caractérisées, quoique pas toujours nommées ; malheureusement, nous n’avons pas toujours pu préciser leurs caractères écologiques et chorologiques.
LYGEO SPARTI-MACROCHLOETEA TENACISSIMAE Rivas Mart. 1978 (Colloq. Phytosoc. VI : 58, sub Lygeo-Stipetea) nom. mut. hoc loco
[syn. : Thero-Brachypodietea ramosi Braun-Blanq. ex A. Bolòs & O. Bolòs 1950 nom. amb. propos. in Rivas Mart., Fern. Gonz. & Loidi 1999 (Itinera Geobot. 13 : 389)]
Typus classis : Lygeo sparti-Macrochloetalia tenacissimae Braun-Blanq. & O. Bolòs 1954 (Collect. Bot. (Barcelona) 4 (2) : 236) nom. mut. hoc loco.
Pelouses steppiques surtout xérophiles méditerranéennes à hautes graminées, notamment Lygeum spartum (photo 1), Stipa div. sp. et Poaceae Andropogoneae, développées sur des sols assez peu rocailleux. À défaut de taxons significativement communs aux trois ordres reconnus ici, la classe est caractérisée par des taxons traversant au moins deux d’entre eux : Macrochloa tenacissima (photo 2), Stipellula parviflora, Lygeum spartum, Plantago albicans, Seriphidium herba-alba, Hyparrhenia hirta (photo 4), Heteropogon contortus ; d’autres taxons plus ou moins bien représentés se retrouvent aussi dans des classes de pelouses vivaces méditerranéennes : Brachypodium retusum, Dactylis glomerata subsp. hispanica, Sedum sediforme subsp. s. (photo 3), Poa bulbosa subsp. b. Pour cette raison, ces quatre taxons ne peuvent être considérés comme vraiment caractéristiques des Lygeo-Macrochloetea et on propose ici de ne conserver dans cette classe que les pelouses élevées significativement pourvues d’Andropogoneae et quelques autres Poaceae, laissant aux Rosmarinetea officinalis les pelouses-garrigues plus basses dépourvues de ces Poaceae (de Foucault, 2020). Le positionnement en particulier de l’alliance du Phlomido lychnitidis-Brachypodion retusi G. Mateo 1983, traditionellement placée dans les Lygeo-Macrochloetea et transférée dans l’ordre des Rosmarinetalia officinalis Braun-Blanq. ex Molin. 1934 par l’un de nous (de Foucault, 2020), méritera certainement des développements complémentaires en lien avec la proposition récente d’isoler une classe d’ourlets nord-ouest méditerranéens, les Charybdido pancratii-Asphodeletea ramosi Biondi 2016 (Biondi et al., 2016, 2017).




Ordre 1. LYGEO SPARTI-MACROCHLOETALIA TENACISSIMAE Braun-Blanq. & O. Bolòs 1954 (Collect. Bot. (Barcelona) 4 (2) : 236) nom. mut. hoc loco (tableau 1 : colonnes ALs à Sp)
[syn. : Thero-Brachypodietalia Braun-Blanq. ex Bharucha 1933 p. p.]
Typus ordinis : Agropyro cristati-Lygeion sparti Braun-Blanq. & O. Bolòs 1954 (Collect. Bot. (Barcelona) 4 (2) : 237, sub Eremopyro cristati-Lygeion sparti) nom. mut. Rivas Mart., Fern. Gonz. & Loidi 1999.
Pelouses steppiques plutôt ouest-méditerranéennes, surtout étudiées dans la péninsule Ibérique, associant Macrochloa tenacissima, Stipellula parviflora ou Lygeum spartum à diverses Lamiaceae partagées avec les Rosmarinetea officinalis (Teucrium pseudochamaepitys, T. capitatum subsp. c., Thymus vulgaris subsp. v.) et quelques espèces des Festuco-Brometea erecti (Koeleria vallesiana s. l., Eryngium campestre).
Alliance ALs. Agropyro pectinati-Lygeion sparti Braun-Blanq. & O. Bolòs 1954 (Collect. Bot. (Barcelona) 4 (2) : 237) nom. mut. Rivas Mart., Fern. Gonz. & Loidi 1999 (Itinera Geobot. 13 : 389) (tableau 1 : colonne ALs ; tableau 2)
Typus allianciae : Lygeo sparti-Stipelluletum parviflorae Braun-Blanq. & O. Bolòs 1954 (Collect. Bot. (Barcelona) 4 (2) : 237, sub Lygeo sparti-Stipetum parviflorae) nom. mut.
Steppes ibériques associant Lygeum spartum, Seriphidium herba-alba et Plantago albicans à Stipellula parviflora, Stipa lagascae ou Agropyrum cristatum subsp. pectinatum ; cartographie in Brullo et al. (2002, figure 4) et Marcenò et al. (2019, figure 2).
ALs1. Lygeo sparti-Stipelluletum parviflorae Braun-Blanq. & O. Bolòs 1954 nom. mut. hoc loco (Sivim, consulté le 01/11/2019).
ALs2. Atractylido humilis-Lygeetum sparti Braun-Blanq. 1936 (Braun-Blanquet, 1936 : 160).
ALs3. Agropyro pectinati-Lygeetum sparti Braun-Blanq. & O. Bolòs 1958 nom. mut. Rivas Mart., T.E. Díaz, Fern. Gonz., Izco, Loidi, Lousã & Penas 2002 (Braun-Blanquet & de Bolòs, 1957, tableau 28, sub Eremopyro cristati-Lygeetum).
ALs4. Dactylido hispanicae-Lygeetum sparti Rivas Mart. ex Alcaraz 1984 (Sivim, consulté le 27/08/2020).
Alliance Mt. Macrochloion tenacissimae Rivas Mart. 1978 (Colloq. Phytosoc. VI : 58, sub Stipion tenacissimae) nom. mut. hoc loco (tableau 1 : colonne Mt ; tableau 3 : colonnes Mt1 à Mt3)
Lectotypus allianciae : Fumano thymifoliae-Macrochloetum tenacissimae Braun-Blanq. & O. Bolòs 1958 (Anales Est. Exp. Aula Dei 5 (1-4) : 161).
Steppes à large répartition dans le centre, le sud-est et l’est de la péninsule Ibérique, à Macrochloa tenacissima, Rosmarinus officinalis subsp. o., Fumana laevipes… ; photo 5.
* Groupe d’associations peu différencié, incluant le type
Mt1. Arrhenathero albi-Macrochloetum tenacissimae Rivas Mart. ex Izco 1969 (Sivim, consulté le 01/11/2019, sub Arrhenathero-Stipetum tenacissimae).
Mt2. Helianthemo squamati-Macrochloetum tenacissimae Pérez-Raya ex A. Garcia in Cano et al. 1995 (Sivim, consulté le 27/08/2020, sub Helianthemo squamati-Stipetum tenacissimae).
Mt3. Fumano thymifoliae-Macrochloetum tenacissimae Braun-Blanq. & O. Bolòs 1958 (Braun-Blanquet & de Bolòs, 1957, tableau 34, sub Fumano thymifoliae-Stipetum tenacissimae).

Alliance LMt. Lapiedro martinezii-Macrochloion tenacissimae all. nov. hoc loco (tableau 1 : colonne LMt ; tableau 3 : colonnes LMt1 à LMt5)
Le second groupe d’associations traditionellement placées dans l’alliance précédente est fortement différencié par Brachypodium retusum… (tableau 3), ce qui suggère une alliance originale caractérisée ou différenciée du Macrochloion tenacissimae par Brachypodium retusum, Fumana laevipes, F. ericoides, Helictochloa bromoides subsp. b., Stipa offneri, Phagnalon saxatile, Hyparrhenia sinaica, Lapiedra martinezii, Phlomis purpurea, Lavandula multifida…
Typus nominis hoc loco : Lapiedro martinezii-Macrochloetum tenacissimae Rivas Mart. & Alcaraz in Alcaraz 1984 (Flora y vegetación del NE de Murcia, non consulté).
LMt1. Heteropogono contorti-Macrochloetum tenacissimae M.B. Crespo in De la Torre, M.B. Crespo & Solanas 1997 (Sivim, consulté le 01/11/2019, sub Heteropogono contorti-Stipetum tenacissimae).
LMt2. Lapiedro martinezii-Macrochloetum tenacissimae Rivas Mart. & Alcaraz in Alcaraz 1984 (Sivim, consulté le 02/11/2019, sub Lapiedro martinezii-Stipetum tenacissimae).
LMt3. Sedo dianii-Macrochloetum tenacissimae De la Torre, M.B. Crespo & Solanas 1997 (Sivim, consulté le 02/11/2019), sub Sedo dianii-Stipetum tenacissimae ; Sedum sediforme subsp. ou var. dianium n’est plus reconnu par les référentiels actuels.
LMt4. Thymo gracilis-Macrochloetum tenacissimae Pérez-Raya & Molero 1988 (Sivim, consulté le 02/11/2019, sub Thymo gracile-Stipetum tenacissimae).
LMt5. Artemisio barrelieri-Anabasietum articulatae López Guadalupe & Esteve 1978 (López Guadalupe & Esteve, 1978, tableau 1).
Alliance Sp. Stipellulion parviflorae De la Torre, Alcaraz & Vicedo 1996 (Acta Bot. Malac. 21 : 298, sub Stipion parviflorae) nom. mut. hoc loco (tableau 1 : colonne Sp ; tableau 4)
Typus allianciae : Plantagini albicantis-Stipelluletum parviflorae De la Torre, Alcaraz & Vicedo 1996 (Acta Bot. Malac. 21 : 298) désigné par les auteurs (1996, à la même page).
Steppes surtout dispersées à l’est et au nord-est de la péninsule Ibérique à Stipellula parviflora (= Stipa parviflora), Sideritis hirsuta, Teucrium gnaphalodes.
Sp1. Plantagini albicantis-Stipelluletum parviflorae De la Torre, Alcaraz & Vicedo 1996 (Sivim, consulté le 2/11/2019).
Sp2. Salvio verbenacae-Plantaginetum albicantis O. Bolòs & Molin. ex O. Bolòs & Vigo in Folch 1981 (Sivim, consulté le 2/11/2019).
Sp3. Dactylido hispanicae-Stipetum junceae Penas, M.E. García, De Paz, L. Herrero, R. Alonso & F. Salegui in Rivas Mart., T.E. Díaz, Fern. Gonz., Izco, Loidi, Lousã & Penas 2002 (Rivas-Martínez et al., 2002, tableau 34).
Sp4. Teucrio gnaphalodis-Astragaletum macrorrhizi A. Velasco 1983 (Sivim, consulté le 3/11/2019).
Sp5. Delphinio gracilis-Lygeetum sparti Conesa 1990 (Conesa, 1990, tableau I).
Sp6. Aristido coerulescentis-Hyparrhenietum sinaicae Rivas Mart. & Alcaraz in Alcaraz 1984 ; a – race ibérique (Sivim, consulté le 3/11/2019 ; Brullo et al., 1997, tableau 3 : relevés 12 à 25) ; b – race marocaine (Brullo et al., 1997, tableau 3 : relevés 1 à 11), peut-être plutôt à rapprocher du troisième ordre de cette classe (cf. infra).
Ordre 2. HYPARRHENIO HIRTAE-BRACHYPODIETALIA RAMOSI Horvatić 1957 ex 1962 (Acta Bot. Croatica XX-XXI : 250, sub Cymbopogo-Brachypodietalia ramosi)
[syn. : Cymbopogono-Brachypodietalia ramosi Horvatić 1957 (Acta Bot. Croatica XVI : 35) nom. inval. ; Cymbopogono-Brachypodietalia ramosi Horvatić 1958 (Acta Bot. Croatica XVII : 15) nom. inval. ; Hyparrhenietalia hirtae Rivas Mart. 1978 (Colloq. Phytosoc. VI : 58) nom. illeg.]
Typus ordinis : Hyparrhenio hirtae-Brachypodion ramosi Horvatić 1957 ex 1962 (Acta Bot. Croatica XX-XXI : 250, sub Cymbopogo-Brachypodion ramosi).
Steppes plutôt nord-méditerranéennes à Hyparrhenia hirta, Phagnalon saxatile, Aristida adscensionis subsp. coerulescens, Piptatherum coerulescens, Micromeria graeca subsp. g., Carlina hispanica subsp. globosa, Andropogon distachyos, Galium lucidum s. l., Thapsia garganica, Lotus cytisoides.
Selon la synthèse déjà citée des Dittrichietea viscosae, l’Hyparrhenion hirtae Braun-Blanq., P. Silva & Rozeira 1956 et le Saturejo graecae-Hyparrhenion hirtae O. Bolòs 1962, validés respectivement par le Carici depressae-Hyparrhenietum hirtae Braun-Blanq., P. Silva & Rozeira 1956 et l’Andropogonetum hirto-pubescentis Braun-Blanq., A. Bolòs & O. Bolòs in A. Bolòs 1950, n’appartiennent pas aux Lygeo-Macrochloetea tenacissimae. Quant au Dauco criniti-Hyparrhenion hirtae O. Bolòs 1962 (El paisaje vegetal barcelonés : 73), il semble bien que ce soit un synonyme illégitime, car postérieur, de l’Hyparrhenion hirtae Braun-Blanq., P. Silva & Rozeira 1956, et invalide (nom. nud.). Cette comparaison justifie que la première alliance ci-après doit correspondre à une entité nouvelle.
Les deux premières alliances, ouest- et sud-ouest-méditerranéennes, assurent le passage des Lygeo-Macrochloetalia tenacissimae aux Hyparrhenio hirtae-Brachypodietalia ramosi ; elles manquent de taxons communs aux sept autres de l’ordre : Asphodelus ramosus s. l., Charybdis maritima…
Alliance Hsh. Hyparrhenion sinaico-hirtae all. nov. hoc loco (tableau 1 : colonne Hsh ; tableau 5)
Typus nominis hoc loco : Heteropogono contorti-Hyparrhenietum sinaicae M.C. Crespo in Rivas Mart., T.E. Díaz, Fern. Gonz., Izco, Loidi, Lousã & Penas 2002 (Itinera Geobot. 15 : 117).
Steppes encore ouest-méditerranéennes à Hyparrhenia sinaica, Stipellula parviflora.
Hsh1. Andryalo laxiflorae-Hyparrhenietum hirtae Peinado, Martínez-Parras & Alcaraz ex Díez Garretas & Asensi 2002 (Rivas-Martínez et al., 2002 : 34).
Hsh2. Dauco criniti-Hyparrhenietum sinaicae Rivas Mart. in Rivas Mart., Fern. Gonz. & Sánchez-Mata 1986 corr. Díez-Garretas & Asensi 1999 (Sivim, consulté le 3/11/2019).
Hsh3. Heteropogono contorti-Hyparrhenietum sinaicae M.C. Crespo in Rivas Mart., T.E. Díaz, Fern. Gonz., Izco, Loidi, Lousã & Penas 2002 (Rivas-Martínez et al., 2002, tableau 49).
Hsh4. Dactylido hylodis-Hyparrhenietum sinaicae Capelo, J.C. Costa, Lousã, Fontinha, Jardim, Sequeira & Rivas Mart. 2000 (Capelo et al., 2000, tableau 14).
Hsh5. Piptathero coerulescentis-Brachypodietum retusi Noble & Baret (Noble & Baret, in prep.).
Hsh6. Phagnalo saxatilis-Andropogonetum distachyi Molin. 1954 (Molinier, 1954, tableau p. 303-304) ; photo 6.
Hsh7. Paragymnopterido marantae-Kengietum serotinae R.J. Loisel 1970 nom. mut. hoc loco (Loisel, 1976, tableau XXII : relevés 328 à 339, sub Cheilantho marantae-Diplachnetum serotinae) ; cf. discussion ci-dessous.
Les pelouses mésohygrophiles à xérophiles franco-méditerranéennes connues à Andropogoneae se classent diversement selon leurs traits écologiques. Avec Anacamptis morio subsp. champagneuxii, Serapias lingua, S. cordigera, plus rarement S. neglecta et S. longipetala, l’Oenantho lachenalii-Chysopogonetum grylli Aubert & R.J. Loisel 1971 peut se ranger dans le Serapiadion cordigero-neglectae B. Foucault 2012 (Aubert & Loisel, 1971 ; Loisel, 1976). Le groupement à Chrysopogon gryllus décrit par Loisel (1976, tableau 1) ne semble pas devoir se ranger dans les Lygeo-Macrochloetea. Enfin le Paragymnopterido marantae-Kengietum serotinae R.J. Loisel 1970 est plus complexe : la sous-association hyparrhenietosum hirtae, qui correspond à l’optimum de Kengia serotina et qui peut être considérée comme le typicum, est justement la plus riche en Andropogonae (genres Hyparrhenia, Heteropogon, Bothriochloa, Andropogon ; Loisel, 1970, 1976). Il semble alors possible d’élever les deux variations au rang d’association, l’hyparrhenietosum hirtae gardant le nom de Paragymnopterido marantae-Kengietum serotinae R.J. Loisel 1970 [lectotypus nominis hoc loco : relevé 2 du tableau 2 in Loisel (1970, Ann. Fac. Sci. Marseille XLIII (8) : 202] et relevant de l’Hyparrhenion sinaico-hirtae, l’euphorbietosum spinosae pouvant plutôt se rattacher aux Rosmarinetea officinalis et au Phlomido lychnitidis-Brachypodion retusi G. Mateo 1983, dont Ruta angustifolia et Iris lutescens subsp. l. sont de bons taxons caractéristiques, en tant qu’association originale : Euphorbio spinosae-Sedetum ochroleuci (Loisel 1970) ass. nov. et stat. nov. hoc loco [basionyme : Paragymnopterido marantae-Diplachnetum serotinae euphorbietosum spinosae R.J. Loisel 1970 (Ann. Fac. Sci. Marseille XLIII (8) : 203)], typus nominis hoc loco : relevé 16 du tableau 2 in Loisel (1970, Ann. Fac. Sci. Marseille XLIII (8) : 202).

Alliance AHh. Aristido coerulescentis-Hyparrhenion hirtae Brullo, Scelsi & Spampinato 1997 (Fitosociologia 32 : 190) (tableau 1 : colonne AHh ; tableau 6)
Typus allianciae : Cenchro ciliaris-Hyparrhenietum sinaicae Wildpret & O. Rodríguez in Rivas Mart., Wildpret, Del Arco, O. Rodríguez, Pérez de Paz, García Gallo, Acebes, T.E. Díaz & Fern. Gonz. 1993 (Itinera Geobot. 7 : 248) désigné par les auteurs de l’alliance (1997 : 190).
Steppes canariennes à Hyparrhenia hirta, Cenchrus ciliaris, Forsskaolea angustifolia, Launaea arborescens, Lavandula canariensis, Lotus sessilifolius, Tetrapogon villosus, empruntant parfois quelques taxons aux Kleinio neriifoliae-Euphorbietea canariensis.
AHh1. Cenchro ciliaris-Hyparrhenietum sinaicae Wildpret & O. Rodríguez in Rivas Mart., Wildpret, Del Arco, O. Rodríguez, Pérez de Paz, García Gallo, Acebes, T.E. Díaz & Fern. Gonz. 1993 corr. Díez-Garretas & Asensi 1999 ; a – variation type (Sivim, consulté le 3/11/2019 ; Brullo et al., 1997, tableau 1 : relevés 1 à 23) ; b – variation à Tricholaena teneriffae (Brullo et al., 1997, tableau 2 : relevés 1 à 10, sub Tricholaeno teneriffae-Hyparrhenietum hirtae).
AHh2. Piptathero coerulescentis-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Brullo et al., 1997, tableau 6 : relevés 1 à 8).
AHh3. Dichanthio foveolati-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Brullo et al., 1997, tableau 8, sub Eremopogono foveolati-Hyparrhenietum hirtae).
Alliance PHh. Phagnalo saxatilis-Hyparrhenion hirtae all. nov. hoc loco (tableau 1 : colonne PHh ; tableau 7)
Typus nominis hoc loco : Bothriochloo panormitanae-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Fitosociologia 32 : 198).
Steppes centre-méditerranéennes voisines de celles de l’alliance suivante, mais moins riches floristiquement, à Ampelodesmos mauritanicus, Hyoseris radiata, Anthyllis vulneraria subsp. maura.
* Groupe d’associations à Lobularia maritima subsp. m. et Andropogon distachyos
PHh1. Bothriochloo panormitanae-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Brullo et al., 1997, tableau 5).
PHh2. Heteropogono contorti-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Brullo et al., 1997, tableau 4).
PHh3. Groupement de Sicile (Brullo et al., 1997, tableau 1 : relevés 24 à 29).
PHh4. Carlino globosae-Hyparrhenietum hirtae ass. nov. hoc loco (Brullo et al., 1997, tableau 2 : relevés 11 à 31). Les relevés 1 à 10 du tableau du Tricholaeno teneriffae-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 représentent un syntaxon validé par un relevé provenant de Ténériffe, il s’avère en fait coïncider avec la sous-association tricholaenetosum teneriffae du Cenchro ciliaris-Hyparrhenietum sinaicae Wildpret & O. Rodríguez in Rivas Mart., Wildpret, Del Arco, O. Rodríguez, Pérez de Paz, García Gallo, Acebes, T.E. Díaz & Fern. Gonz. 1993 corr. Díez-Garretas & Asensi 1999 lui-même canarien (voir supra syntaxon AHh1). Les relevés 11 à 31 décrivent alors une association originale d’Italie méridionale définie ici et validée par le relevé 11 (typus nominis hoc loco) du tableau 2 in Brullo et al. (1997, Fitosociologia 32 : 194).
PHh5. Euphorbio terracinae-Hyparrhenietum hirtae Brullo & Siracusa 1996 (Brullo & Siracusa, 1996, tableau 4).
PHh6. Ferulo communis-Hyparrhenietum hirtae Brullo & Siracusa 1996 (Brullo & Siracusa, 1996, tableau 5).
PHh7. Ferulagini nodosae-Hyparrhenietum hirtae Minissale, Sciandrello & Spampinato 2007 (Brullo et al., 2010, tableau 4, ‘Ferulago…’ art. 41b).
PHh8. Cachryo pungentis-Hyparrhenietum hirtae Brullo, Minissale & Sciandrello in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 5).
PHh9. Hyparrhenio hirtae-Festucetum humifusae Brullo & Guarino in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 6).
PHh10. Stipo gussonei-Hyparrhenietum hirtae Brullo & Scuderi in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 7).
PHh11. Thapsietum pelagicae C. Brullo & Brullo in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 2 : relevés 5 à 7).
* Groupe d’associations à Brachypodium retusum et Kundmannia sicula
PHh12. Cenchro setacei–Hyparrhenietum hirtae Gianguzzi, Ilardi & Raimondo 1996 (Brullo et al., 2010, tableau 1 : colonne 27, sub Penniseto setacei–Hyparrhenietum hirtae) ; relié aussi au groupe précédent.
PHh13. Pulicario odorae-Brachypodietum retusi Ferro & Ladero 1999 (Brullo et al., 2010, tableau 14 : colonne 1).
PHh14. Coronillo glaucae-Brachypodietum retusi C. Brullo, Brullo, Giusso & Tomaselli 2006 (Brullo et al., 2010, tableau 14 : colonnes 2 et 3).
Alliance HAm. Helictochloo cincinnatae-Ampelodesmion mauritanici Minissale ex all. nov. hoc loco (tableau 1 : colonne HAm ; tableau 8)
[syn. : Avenulo cincinnatae-Ampelodesmion mauritanici Minissale 1995 (Colloq. Phytosoc. XXI : 630) nom. inval. (art. 2d, 3o, 5)]
Typus nominis hoc loco : Helictotricho convoluti-Ampelodesmetum mauritanici Minissale 1995 (Colloq. Phytosoc. XXI : 632).
Steppes siciliennes développées sur sol profond et caractérisées par la dominance de Ampelodesmos mauritanicus (photo 8), ainsi que par la présence d’Anthyllis vulneraria subsp. maura, Asphodeline lutea, Helictochloa cincinnata (= Avenula cincinnata), Carlina gummifera, Kundmannia sicula, Micromeria graeca subsp. g., Scorzonera villosa subsp. columnae ; photo 7.
HAm1. Helichryso hyblaei-Ampelodesmetum mauritanici Minissale 1995 (Brullo et al., 2010, tableau 8 : colonne 1).
HAm2. Helictotricho convoluti-Ampelodesmetum mauritanici Minissale 1995 (Brullo et al., 2010, tableau 8 : colonnes 2 à 5).
HAm3. Seseli tortuosi-Ampelodesmetum mauritanici Minissale 1995 (‘Seselio…’, art. 41b) (Brullo et al., 2010, tableau 8 : colonnes 6 et 7).
HAm4. Galio aetnici-Ampelodesmetum mauritanici Minissale 1995 (Brullo et al., 2010, tableau 8 : colonne 8).
HAm5. Astragalo huetii-Ampelodesmetum mauritanici Minissale 1995 (Brullo et al., 2010, tableau 8 : colonnes 9 et 10).
HAm6. Astragalo monspessulanii-Ampelodesmetum mauritanici Minissale 1995 (Brullo et al., 2010, tableau 8 : colonne 11).
HAm7. Arrhenathero nebrodensis-Helictotrichetum convoluti Brullo, Scelsi, Siracusa & Tomaselli 1998 (Brullo et al., 2010, tableau 8 : colonne 12).
HAm8. Helictochloo cincinnatae-Brachypodietum phoenicoidis Brullo, Minissale & Spampinato in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 8 : colonne 13, sub Avenulo cincinnatae-Brachypodietum phoenicoidis).
HAm9. Helictochloo cincinnatae-Stipetum siculae Brullo, Minissale & Spampinato in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 8 : colonnes 14 et 15, sub Avenulo cincinnatae-Stipetum siculae).
HAm10. Helictochloo cincinnatae-Stipetum barbatae Brullo, Giusso & Scuderi in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 8 : colonne 16, sub Avenulo cincinnatae-Stipetum barbatae).
HAm11. Diantho graminifolii-Brachypodietum retusi Brullo, Giusso & Scuderi in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 15 : relevés 6 à 12).
HAm12. Cachryo siculae-Brachypodietum retusi Brullo, Giusso & Scuderi in C. Brullo, Brullo, Giusso, Guarino, Minissale, Scuderi, Siracusa, Sciandrello & Spampinato 2010 (Brullo et al., 2010, tableau 15 : relevés 13 à 20).
HAm13. Helminthotheco aculeatae-Brachypodietum retusi C. Brullo, Brullo, Giusso & Tomaselli 2006 (Brullo et al., 2010, tableau 14 : colonnes 4 et 5).


Alliance Pt. Polygonion tenorei Brullo, De Marco & Signorello 1990 (Boll. Accad. Gioenia Sci. Nat. 23 : 568, sub Polygonion tenoreani) nom. mut. hoc loco (tableau 1 : colonne Pt)
Typus allianciae : Hordeo secalini-Polygonetum tenorei Brullo, De Marco & Signorello 1990 (Boll. Accad. Gioenia Sci. Nat. 23 : 568, sub Hordeo secalini-Polygonetum tenoreani) nom. mut. hoc loco désigné par les auteurs (1990) à la même page.
Steppes à Lygeum spartum des badlands de la Basilicate et du nord de la Calabre, caractérisées par Atriplex halimus, Camphorosma monspeliaca subsp. m. (photo 9), Polygonum tenorei (= P. tenoreanum), Allium paniculatum, vicariantes, sous bioclimat mésoméditerranéen subhumide, du Moricandio arvensis-Lygeion sparti plus xérophile ; cartographie in Marcenò et al. (2019, figure 2).
Synthèse de la colonne 6 de l’Appendix 5 in Marcenò et al. (2019), incluant les Camphorosmo monspeliacae-Lygeetum sparti Brullo, De Marco & Signorello 1990, Hordeo secalini-Polygonetum tenorei Brullo, De Marco & Signorello 1990 nom. mut. hoc loco et Cardopatio corymbosi-Lygeetum sparti Brullo, De Marco & Signorello 1990.

Alliance MLs. Moricandio arvensis-Lygeion sparti Brullo, De Marco & Signorello 1990 (Boll. Accad. Gioenia Sci. Nat. 23 : 569) (tableau 1 : colonne MLs ; tableau 9)
Typus allianciae : Loto cytisoidis-Lygeetum sparti Brullo, De Marco & Signorello 1990 (Boll. Accad. Gioenia Sci. Nat. 23 : 569) désigné par les auteurs (1990, Boll. Accad. Gioenia Sci. Nat. 23 : 568).
Steppes à Lygeum spartum généralement confinées aux pentes abruptes ravinées fortement affectées par l’érosion, sous bioclimat sec thermoméditerranéen, du niveau de la mer jusqu’à 400 m, avec Moricandia arvensis (photo 10), Capparis sicula, Eryngium dichotomum, E. triquetrum ; alliance endémique du sud de la Calabre, de la Sicile, de la Sardaigne et de Malte ; cartographie in Marcenò et al. (2019, figure 2).
MLs1. Eryngio dichotomi-Lygeetum sparti Gentile & Di Benedetto 1961 (Brullo et al., 2010, tableau 20 : colonnes 1 à 3).
MLs2. Malvo agrigentinae-Lygeetum sparti Brullo 1985 (Brullo et al., 2010, tableau 20 : colonnes 8 et 9, sub Lavatero agrigentinae-Lygeetum sparti).
MLs3. Tripolietum sorrentinoi Brullo 1985 (Brullo et al., 2010, tableau 20 : colonnes 10 à 12, sub Asteretum sorrentini).
MLs4. Phagnalo annotici-Lygeetum sparti Biondi & Mossa 1992 (Brullo et al., 2010, tableau 20 : colonnes 4 à 7).
MLs5. Loto cytisoidis-Lygeetum sparti Brullo, De Marco & Signorello 1990 (Brullo et al., 1990, tableau 4).

Alliance PrHh. Panico repentis-Hyparrhenion hirtae Brullo & Siracusa 2000 (Arch. Geobot. 4 (1) : 75) (tableau 1 : colonne PrHh)
Typus allianciae : Imperato cylindricae-Hyparrhenietum hirtae Brullo & Siracusa 2000 (Arch. Geobot. 4 (1) : 76) désigné par les auteurs (1998, Arch. Geobot. 4 (1) : 75).
Pelouses mésohygrophiles du sud de l’Italie et de la Sicile à Hyparrhenia hirta, Panicum repens, Imperata cylindrica et Dichanthium annulatum. Cette alliance pourrait atteindre le sud-est de la France et la Corse où est connu P. repens ; toutefois les populations connues de Panicum repens en France continentale ne présentent aucun lien avec les Lygeo-Macrochloetea, étant plutôt liées à des sables rudéralisés en arrière-dune avec Sonchus bulbosus, Lobularia maritima, Daucus carota s. l., Pancratium maritimum, Phragmites australis et divers espèces issues des Euphorbio paraliae-Ammophiletea australis et des Juncetea maritimi.
Synthèse de trois syntaxons in Brullo et al. (2010, tableau 1 : colonnes 19 à 21), les Imperato cylindricae-Hyparrhenietum hirtae Brullo & Siracusa 2000, Dichanthio annulati-Hyparrhenietum hirtae Brullo & Siracusa 2000 et Phalarido coerulescentis-Hyparrhenietum hirtae Brullo & Siracusa 2000.
Alliance SLs. Scorzonero creticae-Lygeion sparti Brullo, Giusso & Guarino 2002 (Lazaroa 23 : 70) (tableau 1 : colonne SLs ; tableau 10)
Typus allianciae : Erodio crassifolii-Lygeetum sparti Brullo, Giusso & Guarino 2002 (Lazaroa 23 : 70) désigné par les auteurs (2002 : 70).
Steppes plutôt est-méditerranéennes (Italie, Libye, Grèce) à Hyparrhenia hirta, Thymbra capitata, Phagnalon graecum, Carlina graeca, Helichrysum stoechas subsp. barrelieri, Piptatherum holciforme, surtout décrites par Brullo et al. (1997), puis Brullo et al. (2002) ; cartographie in Brullo et al. (2002, figure 4) et Marcenò et al. (2019, figure 2). Deux groupes d’associations.
* Groupe d’associations à Aristida adscensionis subsp. coerulescens
SLs1. Thymbro capitatae-Hyparrhenietum hirtae ass. nov. hoc loco ; ce syntaxon italien (Brullo et al., 1997, tableau 6 : relevés 9 à 19, sub Oryzopsio coerulescentis–Hyparrhenietum hirtae) a été initialement rattaché au Piptathero coerulescentis-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 nom. mut. hoc loco (cf. supra, syntaxon AHh2), lequel doit garder le nom initial, étant typifié par le relevé 3 du tableau 6 des auteurs ; ce syntaxon SLs1 doit alors en être séparé ; typus nominis hoc loco : relevé 16 du tableau 6 in Brullo, Scelsi & Spampinato (1997, Fitosociologia 32 : 200).
SLs2. Piptathero holciformis-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 nom. mut. hoc loco ; a – race de Cyrénaïque (Brullo et al., 1997, tableau 7 : relevés 1 à 6, sub Oryzopsio holciformis–Hyparrhenietum hirtae) ; b – race de Crète (Brullo et al., 1997, tableau 7 : relevés 7 à 11) ; ces deux races pourraient être distinguées comme deux associations.
SLs3. Diantho criniti-Hyparrhenietum hirtae Brullo, Scelsi & Spampinato 1997 (Brullo et al., 1997, tableau 9).
* Groupe d’associations à Lygeum spartum et Scorzonera cretica, incluant le type
SLs4. Limonio hyssopifolii-Lygeetum sparti Brullo, Giusso & Guarino 2002 (Brullo et al., 2002, tableau 1 : relevés 1 à 15).
SLs5. Erodio crassifolii-Lygeetum sparti Brullo, Giusso & Guarino 2002 (Brullo et al., 2002, tableau 1 : relevés 16 à 24).
SLs6. Convolvulo oleifolii-Lygeetum sparti Brullo, Giusso & Guarino 2002 (Brullo et al., 2002, tableau 1 : relevés 25 à 34).
La photo 11 illustre une steppe crétoise qui pourrait se rattacher à cette alliance.

Alliance HBr. Hyparrhenio hirtae-Brachypodion retusi Horvatić 1957 ex 1962 (Acta Bot. Croatica XX-XXI : 250, sub Cymbopogo-Brachypodion ramosi) (tableau 1 : colonne HBr ; tableau 11)
[syn. : Cymbopogono-Brachypodion ramosi Horvatić 1957 (Acta Bot. Croatica XVI : 35) nom. inval. ; Cymbopogono-Brachypodion ramosi Horvatić 1958 (Acta Bot. Croatica XVII : 15) nom. inval.]
Typus allianciae : Brachypodio ramosi-Hyparrhenietum hirtae Horvatić 1962 (Horvatić, 1961-62, Acta Bot. Croatica XX-XXI : 250, sub Brachypodio retusi-Cymbopogonetum hirti).
Pelouses croates à Hyparrhenia hirta avec Petrorhagia saxifraga, Teucrium polium subsp. p., Convolvulus althaeoides subsp. tenuissimus, Carlina corymbosa, Aethionema saxatile, Allium subhirsutum, Helichrysum italicum, Fumana ericoides.
Les données initiales de Horvatić (1961-62, tableau V) sont assez réduites (seulement deux relevés) et ne permettent guère de cerner l’identité précise du Brachypodio ramosi-Hyparrhenietum hirtae ; les deux autres associations de cette alliance paraissent assez distinctes de ce dernier, surtout HBr3 qui sera donc réinterprété.
HBr1. Brachypodio retusi-Hyparrhenietum hirtae Horvatić 1957 ex 1962 (Horvatić, 1961-62, tableau V, sub Brachypodio ramosi-Cymbopogonetum hirti).
HBr2. Cymbopogono hirti-Brachypodietum ramosi sensu Trinajstić & Kamenjarin 2001 (Trinajstić & Kamenjarin, 2001, tableau 1).
HBr3. Cymbopogono hirti-Brachypodietum ramosi sensu Hećimović 1984 (Hećimović, 1984, tableau 2) nom. illeg. (non Brachypodio ramosi-Hyparrhenietum hirtae Horvatić 1957 ex 1962), renommé Koelerio lobatae-Hyparrhenietum hirtae (Hećimović 1984) nom. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 2 in Hećimović (1984, Acta Bot. Croatica 43 hors texte).
On peut rapprocher de ces deux alliances le Micromerio graecae-Hyparrhenietum hirtae de Céphalonie (une des îles Ioniennes grecques) décrit par de Bolòs et al. (1996, tableau 11), qui montre des liens avec les Cisto-Micromerietea julianae Oberd. 1954 (notamment par Sarcopoterium spinosum).
Ordre 3. STIPELLULO PARVIFLORAE-LYGEETALIA SPARTI ord. nov. hoc loco
[syn. : Stipetalia tenacissimae Djebaili 1990 (Ecol. Medit. 16 : 238) nom. inval.]
Typus nominis hoc loco : Noaeo mucronatae-Lygeion sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Appl. Veg. Sci. X : 9).
Steppes nord-africaines à Lygeum spartum, Stipellula parviflora, Stipa barbata ; unité de transition entre les Lygeo sparti-Macrochloetea tenacissimae nord- et eu-méditerranéens et les Pergulario tomentosae-Pulicarietea crispae Quézel 1965 sahariens comme le montre l’apparition d’Helianthemum lippii et quelques autres taxons (Quézel, 1965 ; de Foucault & Lazare, 2020).
Alliance NLs. Noaeo mucronatae-Lygeion sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Appl. Veg. Sci. X : 9) (tableau 1 : colonne NLs ; tableau 12)
[syn. : Noaeo mucronatae-Artemision herbae-albae Aidoud 1990 (Doc. Phytosoc., NS, XII : 112) nom. inval. (art. 8) ; ? Macrochloo tenacissimae-Launaeion laniferae Djebaili 1990 (Ecol. Medit. 16 : 238) nom. inval., sub Stipo-Launaeion acanthocladae]
Typus allianciae : Noaeo mucronatae-Lygeetum sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Appl. Veg. Sci. X : 9).
Steppes surtout à Macrochloa tenacissima caractérisées par Noaea mucronata subsp. m. (photo 12), Nitrosalsola vermiculata (photo 13), Echium pycnanthum, Paronychia arabica, Helianthemum virgatum, Atractylis humilis, Marrubium deserti ; cartographie in Marcenò et al. (2019, figure 2) ; photo 14.
NLs1. Noaeo mucronatae-Lygeetum sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Aidoud, 1990, tableau I, sub Noaeo mucronatae-Artemisietum herbae-albae nom. inval.) ; reconnaissable aussi chez Wojterski (1985 : 127).
NLs2. Groupement à Thymelaea microphylla-Stipellula parviflora (Aidoud, 1990, tableau I, « Hippocrepido-Hedysaretum spinosissimi ») ; forme du précédent.
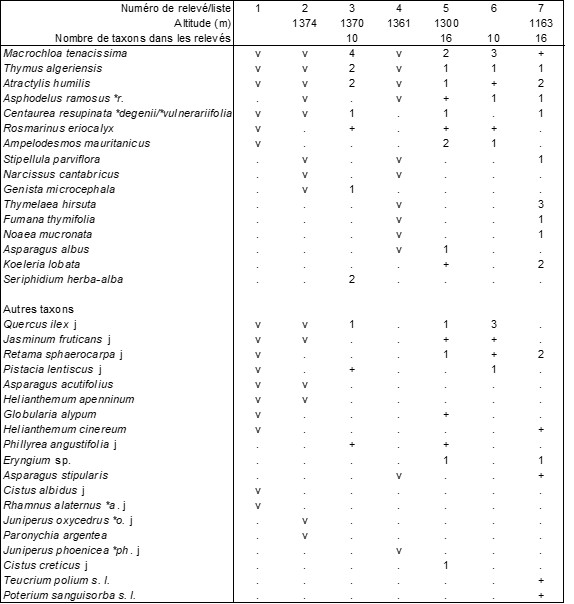
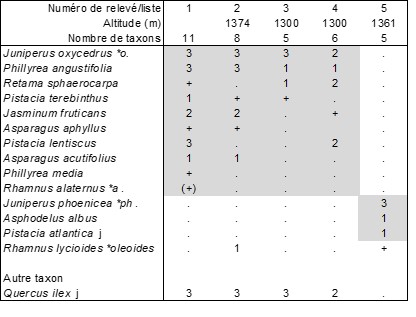
NLs3. Thymo algeriensis-Macrochloetum tenacissimae B. Foucault ass. nov. hoc loco (tableau 13, ci-dessus ; photo 15), association définie à partir de quatre relevés et trois listes provenant du massif du Nador, au sud de Tiaret (Algérie) réalisés entre le 4 et le 7 novembre 2012 à une altitude variant de 1 100 à 1 400 m ; typus nominis hoc loco : relevé 7 du tableau 13 hoc loco. La présence de jeunes arbustes ou arbres dans ce Thymo algeriensis-Macrochloetum tenacissimae autorise à poser l’hypothèse d’une dynamique progressive vers des fourrés, à étudier au voisinage de la steppe ; le tableau 14 ci-dessus en rapporte effectivement cinq relevés ; laissant à part le relevé 5, unique représentant d’un syntaxon à confirmer, les relevés 1 à 4 décrivent bien un fourré original des Pistacio lentisci-Rhamnetea alaterni, le Retamo sphaerocarpae-Juniperetum oxycedri B. Foucault ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 14 hoc loco (photo 16) ; cette association semble se rattacher à l’Asparago albi-Rhamnion oleoidis Rivas Goday ex Rivas Mart. 1975 d’optimum ibérique (de Foucault, 2021).
NLs4. Groupement à Seriphidium herba-alba et Macrochloa tenacissima (Benabadji & Bouazza, 2002, tableau 1).
On peut rapprocher de cette alliance la « steppe à Stipa tenacissima » algérienne étudiée par Lazare & Roux (1979, tableau floristique : syntaxon IV). Une autre végétation algérienne relevant de cette alliance est aussi rapportée par Henni & Mehdadi (2012, tableau 2), mais dans une publication à orientation plus phytoécologique que phytosociologique.





Alliance LlLs. Launaeo laniferae-Lygeion sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Appl. Veg. Sci. X : 7) (tableau 1 : colonne LlLs)
Typus allianciae : Convolvulo trabutiani‐Lygeetum sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019 (Appl. Veg. Sci. X : 7) désigné par les auteurs à la même page.
Steppes surtout marocaines à Launaea lanifera, Paronychia chlorothyrsa, Convolvulus trabutianus, Teucrium luteum subsp. flavovirens, Catananche caerulea, Thymus willdenowii, unité déduite des données inédites de Peltier (1982) ; cartographie in Marcenò et al. (2019, figure 2).
Synthèse de la colonne 2 de l’Appendix 5 in Marcenò et al. (2019).
Alliance LaLs. Launaeo angustifoliae-Lygeion sparti Le Houérou 1969 (Ann. Inst. Natl. Rech. Agron. Tunisie 42 non consulté) (tableau 1 : colonne LaLs)
Lectotypus nominis hoc loco : Aristido ciliatae-Lygeetum sparti Le Houérou 1969 (Ann. Inst. Natl. Rech. Agron. Tunisie 42 : 234).
Steppes surtout tunisiennes à Gymnocarpos decander (photo 17), Deverra tortuosa, D. denudata, Helianthemum lippii, Echiochilon fruticosum, Reaumuria vermiculata (photo 18), Echium humile, Thymelaea hirsuta, Artemisia campestris subsp. glutinosa, Ajuga iva, Launaea angustifolia ; cartographie in Marcenò et al. (2019, figure 2).
Synthèse de la colonne 4 (liste incomplètement reprise ici) de l’Appendix 5 in Marcenò et al. (2019).
La photo 19 illustre une steppe à Macrochloa tenacissima et Seriphidium herba-alba des environs de Douiret, au sud-est de la Tunisie.



4. Conclusion
En définitive, le schéma synsystématique de la classe obtenu s’établit ainsi (en grisé, l’alliance présente en France) :
LYGEO SPARTI-MACROCHLOETEA TENACISSIMAE Rivas Mart. 1978
LYGEO SPARTI-MACROCHLOETALIA TENACISSIMAE Braun-Blanq. & O. Bolòs 1954
Agropyro pectinati-Lygeion sparti Braun-Blanq. & O. Bolòs 1954 nom. mut. Rivas Mart., Fern. Gonz. & Loidi 1999
Macrochloion tenacissimae Rivas Mart. 1978
Lapiedro martinezii-Macrochloion tenacissimae nov.
Stipellulion parviflorae De la Torre, Alcaraz & Vicedo 1996
HYPARRHENIO HIRTAE-BRACHYPODIETALIA RETUSI Horvatić 1957 ex 1962
Hyparrhenion sinaico-hirtae nov.
Aristido coerulescentis-Hyparrhenion hirtae Brullo, Scelsi & Spampinato 1997
Phagnalo saxatilis-Hyparrhenion hirtae nov.
Helictochloo cincinnatae-Ampelodesmion mauritanici Minissale ex B. Foucault & Noble
Polygonion tenorei Brullo, De Marco & Signorello 1990
Moricandio arvensis-Lygeion sparti Brullo, De Marco & Signorello 1990
Panico repentis-Hyparrhenion hirtae Brullo & Siracusa 2000
Scorzonero creticae-Lygeion sparti Brullo, Giusso & Guarino 2002
Hyparrhenio hirtae-Brachypodion retusi Horvatić 1957 ex 1962
STIPELLULO PARVIFLORAE-LYGEETALIA SPARTI B. Foucault & Noble nov.
Noaeo mucronatae-Lygeion sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019
Launaeo laniferae-Lygeion sparti Marcenò, Guarino, Mucina, Biurrun, Deil, Shaltout, Finckh, Font & Loidi 2019
Launaeo angustifoliae-Lygeion sparti Le Houérou 1969
Il est certain que la partie maghrébine de cette synthèse est insuffisamment représentée ; plusieurs thèses restées inédites sont difficilement accessibles. Sans doute aussi ces steppes sont-elles sous-étudiées au plan phytosociologique.
Comme dit plus haut, vers le sud, ces steppes à Andropogoneae passent progressivement aux Pergulario tomentosae-Pulicarietea crispae Quézel 1965 surtout chaméphytiques (Quézel, 1965 ; de Foucault & Lazare, 2020). Mais au-delà de la zone saharienne, les savanes sahélo-soudaniennes, malgaches… (photos 20 et 21) sont aussi de hautes fomations végétales vivaces elles-mêmes caractérisées par des Andropogoneae, avec à nouveau les genres Andropogon, Bothriochloa, Chrysopogon, Dichanthium, Heteropogon, Hyparrhenia, mais aussi en plus Anadelphia, Capillipedium, Cymbopogon, Diheteropogon, Elionurus, Elymandra, Eremopogon, Eriochrysis, Eulalia, Hemarthria, Heteropholis, Homozeugos, Hyperthelia, Hypogynium, Ischaemum, Jardinea, Miscanthus, Monocymbium, Ophiuros, Oxyrhachis, Parahyparrhenia, Phacelurus, Rhytachne, Rottboellia, Schizachyrium, Sehima, Sorghum, Themeda, Trachypogon, Urelytrum (incl. Brachiaria)… (Lebrun, 1947 ; Mullenders, 1954 ; Morat, 1973 ; White, 1986 ; Kellogg, 2015…). Notamment, en termes d’association, Lebrun (1947) décrit validement les Themedo triandrae-Heteropogonetum contorti, Eragrostietum paniciformis, Bothriochloetum insculptae et Cymbopogonetum afronardi (sub Afronardetum). Duvigneaud (1953) définit validement quatre alliances pour l’ex-Congo belge : Crossopterygo febrifugae-Andropogonion pseudaprici, Landolphio lanceolatae-Trachypogonion spicati, Loudetion simplicis et Hymenocardio acidae-Loudetion arundinaceae (noms mutés selon la nomenclature actuelle). De même, en Côte-d’Ivoire centrale, Adjanohoun (1964) décrit validement l’Urochloetum serratae (sub Brachiarietum brachylophae) et le Loudetietum phragmitoidis. De son côté, Mullenders (1954) décrit le Loudetio kagerensis-Fimbristylidetum gabonicae, le Sclerio lagoensis-Elymandretum androphilae, l’Ochno leptocladae-Loudetietum arundinaceae, le Sopubio densiflorae-Andropogonetum schirensis ; l’« Acacio campylacanthae-Beckeriopsietum unisetae » offre un autre exemple d’association où l’on peut séparer
- une association arborescente, le Senegalio campylacanthae-Sclerocaryetum caffrae Foucault ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (types biologiques Pma et Pme de Mullenders) du relevé 1 du tableau XXIV in Mullenders (1954, Publ. Inst. Natl. Étude Agron. Congo belge, série sci., 61, hors texte) ;
- une association arbustive, l’Annono senegalensis-Piliostigmatetum thonningii Foucault ass. nov. hoc loco, typus nominis hoc loco: composante arbustive (types biologiques surtout Pmi et Pn de Mullenders) du relevé 1 du tableau XXIV in Mullenders (1954, Publ. Inst. Natl. Étude Agron. Congo belge, série sci., 61, hors texte) ;
- une savane vivace, l’Acalypho petiolaris-Cenchretum uniseti Foucault ass. nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 1 du tableau XXIV in Mullenders (1954, Publ. Inst. Natl. Étude Agron. Congo belge, série sci., 61, hors texte) ; une unique Andropogoneae annuelle y est aussi abondante et constante, Hyparrhenia confinis.


Est-il nécessaire de rappeler le rôle écosystémique fondamental de ces savanes pour le maintien de la grande faune herbivore africaine et par suite de la faune carnivore qui lui est liée (respectivement Connochaeto taurini-Hippotigridetum bohmi et Pantheretum pardo-leonis en termes zoosociologiques ; de Foucault, 1990).
Bibliographie
Adjanohoun E., 1964. Végétation des savanes et des rochers découverts en Côte-d’Ivoire centrale. Mémoires ORSTOM 7 : 1-178 et annexes.
Aidoud F., 1990. Analyse syntaxonomique des groupements steppiques du complexe Lygeum spartum- Artemisia herba-alba-Stipa tenacissima du Sud-Ouest algérien. Documents phytosociologiques, NS, XII : 103-121.
Aidoud-Lounis F., Celles J.-C. & Loisel R., 1999. Les communautés thérophytiques des steppes arides d’Algérie. Documents phytosociologiques, NS, XIX : 15-45.
Alcaraz F., 1984. Flora y vegetación del NE de Murcia. Murcia Secretariado de Publicaciones de la Universidad de Murcia, 404 p.
Asensi A., Díez-Garretas B. & Martín Osorio M.V., 1993. The structure and dynamics of Festuca scariosa communities in southern Spain. Ecologia Mediterranea 19 (3-4) : 1-7.
Aubert G. & Loisel R., 1971. Contribution à l’étude des groupements des Isoeto-Nanojuncetea et des Helianthemetea annua dans le Sud-Est méditerranéen français. Annales de l’Université de Provence XLV : 203-241.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Patrimoines naturels 61 : 1-171.
Benabadji N. & Bouazza M., 2002. Contribution à l’étude du cortège floristique de la steppe au sud d’El-Aricha (Oranie, Algérie). Sciences & Technologie (Constantine) n° sp. D : 11-19.
Biondi E., Pesaresi S., Galdenzi D., Gasparri R., Biscotti N., Del Viscio G. & Casavecchia S., 2016. Post-abandonment dynamic on Mediterranean and sub-Mediterranean perennial grasslands: the edge vegetation of the new class Charybdido pancratii-Asphodeletea ramosi. Plant Sociology 53 (2) : 3-18.
Biondi E., Pesaresi S., Gasparri R., Biscotti N., del Viscio G., Bonsanto D. & Casavecchia S., 2017 New contributions to the class Charybdido pancratii-Asphodeletea ramosi Biondi 2016. Plant Sociology 54 (1) : 137-144.
Bolòs O. (de), 1962. El paisaje vegetal barcelonés. Cátedra Ciudad de Barcelona, 192 p.
Bolòs O. (de), Masalles R.M., Ninot J.M. & Vigo J., 1996. A survey on the vegetation of Cephalonia (Ionian islands). Phytocoenologia 26 (1) : 81-123.
Braun-Blanquet J. ,1925. Die Brachypodium ramosum-Phlomis lychnitis-Assoziation der Roterdeböden Südfrankreichs. Veröffentlichungen des Geobotanischen Institutes der E.T.H. Stiftung Rübel 3 : 304-320.
Braun-Blanquet J., 1936. L’excursion de la SIGMA en Catalogne (Pâques 1934). Cavanillesia VII : 153-167.
Braun-Blanquet J., Font Quer P., Braun-Blanquet G., Frey E., Jansen P. & Moor M., 1936. L’excursion de la Sigma en Catalogne (Pâques 1934). Cavanillesia VII : 89-110.
Braun-Blanquet J., Roussine N. & Nègre R., 1952. Les groupements végétaux de la France méditerranéenne. CNRS, Paris. 297 p.
Braun-Blanquet J. & de Bolòs O., 1954. Datos sobre las comunidades terofíticas de las llanuras del Ebro medio. Collectanea Botanica (Barcelona) 4 (2) : 235-242.
Braun-Blanquet J. & de Bolòs O., 1957 (publié 1958). Les groupements végétaux du bassin moyen de l’Ebre et leur dynamisme. Anales de la Estacion experimental Aula Dei 5 (1-4) : 1-266.
Brullo C., Brullo S., Giusso del Galdo G., Guarino R., Minissale P., Scuderi L., Siracusa G., Sciandrello S. & Spampinato G., 2010. The Lygeo‐Stipetea class in Sicily. Annali di Botanica (Roma) 0 : 1-31.
Brullo S., 1985. Sur la syntaxonomie des pelouses thérophytiques des territoires steppiques de l’Europe sud-occidentale. Documents phytosociologiques, NS, IX : 1-17.
Brullo S., De Marco G. & Signorello P., 1990. Studio fitosociologico delle praterie a Lygeum spartum dell’Italia meridionale. Bollettino dell’Accademia Gioenia di Scienze Naturali in Catania 23 : 561-579.
Brullo S. & Siracusa G., 1996. Studio fitosociologico dell’Isola di Linosa. Documents phytosociologiques, NS, XVI : 123-174.
Brullo S., Scelsi F. & Spampinato G., 1997. Aristido caerulescentis-Hyparrhenion hirtae, alleanza nuova della classe Lygeo-Stipetea a distribuzione sud-mediterraneo-macaronesica. Fitosociologia 32 : 189-206.
Brullo S. & Siracusa G., 1998 (publié 2000). Indagine fitosociologica di un’area umida del versante sud-occidentale dell’Etna di notevole interesse naturalistico. Archivio Geobotanico 4 (1) : 71-90.
Brullo S., Giusso del Galdo G., & Guarino R., 2002. Phytosociological notes on the Lygeum spartum grasslands from Crete. Lazaroa 23 : 65-72.
Capelo J.H., Costa J.C., Lousã M., Fontinha S., Jardim R., Sequeira M. & Rivas-Martínez S., 2000. Vegetação da Madeira (Portugal): Aproximação à tipologia fitosociologica. Silva Lusitana 7 (2) : 257-279.
Conesa J.A., 1990. Notes i addicions sobre la vegetació del territori Sicòric, I. Folia Botanica Miscellenea 7 : 87-97.
Djebaili S., 1990. Syntaxonomie des groupements préforestiers et steppiques de l’Algérie aride. Ecologia Mediterranea 16 : 231-244.
Duvigneaud P., 1953. Les savanes du Bas-Congo, essai de phytosociologie topographique. Lejeunia 10 : 1-192.
Foucault B. (de), 1990. Impressions biosociologiques après un voyage en Tanzanie. Documents phytosociologiques, NS, XII : 175-188.
Foucault B. (de), 1993. Remarques sur la végétation du Maroc et de l’Algérie. Bulletin de la Société botanique du Centre-Ouest, NS, 24 : 267-285
Foucault B. (de), 1999. Nouvelle contribution à une synsystématique des pelouses sèches à thérophytes. Documents phytosociologiques, NS, XIX : 47-105.
Foucault B. (de), 2013. Contribution au prodrome des végétations de France : les Isoëtetea velatae de Foucault 1988 et les Juncetea bufonii de Foucault 1988 (« Isoëto–Nanojuncetea bufonii »). Journal de botanique de la Société botanique de France 62 : 35-70, 63 : 63-109.
Foucault B. (de), 2020. Sur la synsystématique des Rosmarinetea officinalis ouest-méditerranéens. Carnets botaniques 4 : 1-27 + 26 tableaux, https://doi.org/10.34971/s9d8-qj43.
Foucault B. (de), 2021. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de) & Jasprica N., 2019. Sur une classe de végétation méconnue en France : les Dittrichietea viscosae Trinajstić, B. Foucault & Jasprica cl. nov. Botanique 5 : 155-170.
Foucault B. (de) & Lazare J.-J., 2020. Contribution à une synthèse phytosociologique des steppes d’Afrique du Nord (hors Lygeo-Stipetea tenacissimae). Botanique 6 : 53-116.
García‐Fuentes A., Salazar C., Torres J. A., Cano E. & Valle, F., 2001. Review of communities of Lygeum spartum L. in the south‐eastern Iberian Peninsula (western Mediterranean). Journal of Arid Environment 48 : 323-333.
Guinochet M., 1951 (publié 1952). Contribution à l’étude phytosociologique du Sud tunisien. Bulletin de la Société d’histoire naturelle d’Afrique du Nord 42 : 131-153.
Guinochet M., 1973. Phytosociologie. Masson, Paris, 227 p.
Guinochet M., 1978. Contribution à la synsystématique des pelouses thérophytiques du nord de la Tunisie et de l’Algérie. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 1-18.
Hećimović M., 1984. Grassland vegetation of the island of Šipan. Acta Botanica Croatica 43 : 161-166.
Henni M. & Mehdadi Z., 2012. Évaluation préliminaire des caractéristiques et floristiques des steppes à armoise blanche dégradées réhabilitées par la plantation d’Atriplex dans la région de Saïda (Algérie occidentale). Acta Botanica Gallica: Botany Letters 159 (1) : 43-52.
Horvatić S., 1957. Pflanzengeographische Gliederung des Karstes Kroatiens und der angrenzenden Gebiete Jugoslawiens. Acta Botanica Croatica XVI : 33-61 + 1 carte.
Horvatić S., 1958. Tipološko raščlanjenje primorske vegetacije gariga i borovih šuma. Acta Botanica Croatica XVII : 243-259.
Horvatić S., 1961-62. Novi prilog poznavanju primorske vegetacije gariga i kamenjarskih pašnjaka. Acta Botanica Croatica XX-XXI : 243-259.
Kaabèche M., 1990. Les groupements végétaux de la région de Bou Saada (Algérie). Essai de synthèse sur la végétation steppique du Maghreb. Thèse de doctorat de l’Université de Paris-Sud, Centre d’Orsay, 104 p.
Kaabèche M., 1995. Flore et végétation dans le chott El-Hodna (Algérie). Documents phytosociologiques, NS, XV : 393-402.
Kaabèche M., 1996. La végétation steppique du Maghreb (Algérie, Maroc, Tunisie). Essai de synthèse phytosociologique par application des techniques numériques d’analyse. Documents phytosociologiques, NS, XVI : 45-58.
Kellogg E.A., 2015. Poaceae. In K. Kubitzki (ed.), The families and genera of vascular plants XIII : 1-416.
Lazare J.-J. & Roux G., 1979. Quelques groupements végétaux des hauts plateaux au sud-ouest de Bou-Saada (Algérie). Documents phytosociologiques, NS, IV : 585-596.
Le Houérou H.-N., 1969. La végétation de la Tunisie steppique avec références aux végétations analogues de l’Algérie de la Libye et du Maroc. Annales de l’Institut national de recherches agronomiques de Tunisie 42 : 1-624.
Lebrun J., 1947. La végétation de la plaine alluviale au sud du lac Édouard. Institut des parcs nationaux du Congo belge, Bruxelles, 800 p.
Lemée G., 1953. Contribution à la connaissance phytosociologique des confins saharo-marocains. Les associations à thérophytes des dépressions sableuses et limoneuses non salées et des rocailles aux environs de Beni-Ounif. Vegetatio 4 : 137-154.
Loisel R., 1970. Contribution à l’étude des pelouses à Andropogonées du littoral provençal. Annales de la Faculté des sciences de Marseille XLIII (8) : 197-213.
Loisel R., 1976. La végétation de l’étage méditerranéen dans le Sud-Est continental français. Thèse de doctorat de l’université d’Aix-Marseille. 384 p.
López G., 1976. Contribución al conocimiento fitosociológico de la Serranía de Cuenca. I – Comunidades fruticosas: bosques, matorrales, tomillares y tomillar-praderas. Anales del Instituto Botanico A.J. Cavanilles 33 : 5-87.
López Guadalupe M. & Esteve F., 1978. Adiciones al estudio de la vegetación y flora de la Penibetica. Anales del Instituto Botanico A.J. Cavanilles 34 (2) : 521-533.
Marcenò C., Guarino R., Mucina L., Biurrun I., Deil U., Shaltout K., Finckh M., Font X. & Loidi J., 2019. A formal classification of the Lygeum spartum vegetation of the Mediterranean Region. Applied Vegetation Science 0 : 1-16, https://doi.org/10.1111/avsc.12456.
Martínez-Parras J.M., Peinado M. & Alcaraz F., 1984. Estudio de la serie mesomediterránea basífila de la encina (Paeonio-Querceto rotundifoliae S.). Lazaroa 5 : 119-129.
Martínez-Parras J.M., Peinado M. & Alcaraz F., 1987. Algunas comunidades orófilas de Andalucía oriental. Lazaroa 7 : 49-53.
Mateo G., 1983. Estudio sobre la flora y vegetación de las sierras de Mira y Talayuelas. Monografia ICONA 31 : 1-290.
Minissale P., 1995. Studio fitosociologico delle praterie ad Ampelodesmos mauritanicus della Sicilia. Colloques phytosociologiques XXI, Ecologia del paesaggio e progettazione ambientale : 615-652.
Molinier R., 1954. Les climax côtiers de la Méditerranée occidentale. Vegetatio 4 : 284-308.
Morat Ph., 1973. Les savanes du sud-ouest de Madagascar. Mémoires ORSTOM 68 : 1-235.
Mullenders W., 1954. La végétation de Kaniama (Entre-Lubishi-Lubilash, Congo belge). Publications de l’Institut national pour l’étude agronomique du Congo belge, série scientifique, 61 : 1-499.
Nègre R., 1978. Données phytosociologiques sur les associations thérophytiques du Maroc aride. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 23-32.
Noble V. & Baret J., 2020. Catalogue des végétations du département des Bouches-du-Rhône. Conservatoire botanique national méditerranéen de Porquerolles, à paraître.
Peltier J.-P., 1982. La végétation du bassin versant de l’oued Sous (Maroc). Thèse de doctorat de l’université de Grenoble.
Quézel P., 1954. Contribution à l’étude de la flore et de la végétation du Hoggar. Monographies de l’Institut de recherches sahariennes 2 : 1-164.
Quézel P., 1965. La végétation du Sahara du Tchad à la Mauritanie. Geobotanica Selecta II : 1-333.
Rivas-Martínez S., 1978. Sur la syntaxonomie des pelouses thérophytiques de l’Europe occidentale. Colloques phytosoiologiques VI, La végétation des pelouses sèches à thérophytes : 55-69.
Rivas-Martínez S., Wildpret W., Del Arco M., Rodríguez O., Pérez de Paz P.L., García Gallo A., Acebes J.R., Díaz T.E. & Fernández-González F., 1993. Las comunidades vegetales de la Isla de Tenerife (Islas Canarias). Itinera Geobotanica 7 : 169-374.
Rivas-Martínez, S., Fernández-González F. & Loidi J., 1999. Checklist of plant communities of Iberian Peninsula, Balearic and Canary Islands to suballiance level. Itinera Geobotanica 13 : 353-451.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal; addenda to the syntaxonomical checklist of 2001. Itinera Geobotanica 15 : 5-922.
Sivim. Sistema de Información de la Vegetación Ibericá y Macaronesica, www.sivim.info.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.
Torre A. (de la), Alcaraz F. & Vicedo M., 1996. Stipion parviflorae all. nova, pastizales vivaces subnitrófilos y calcícolas mediterráneo-iberolevantinos. Acta Botanica Malacitana 21 : 297-302.
Trinajstić I. & Kamenjarin J., 2001. Fitocenološke značajke pašnjačke asocijacije Brachypodio-Cymbopogonetum hirti H-ić 1961. Agronomski Glasnik (4-5) : 181-188.
White F., 1986. La végétation de l’Afrique. Recherches sur les ressources naturelles XX : 1-384.
Wojterski T.W., 1985. Guide de l’excursion internationale de phytosociologie en Algérie du Nord. INA, El Harrach, 274 p.


