
Contribution à une monographie phytosociologique du domaine de Peyremale (commune de Montolieu, Aude)
Title
Contribution to a phytosociological monograph of the Peyremale domain (Montolieu, Aude)
Résumé
Cet article apporte des données phytosociologiques sur la végétation d’un site à diversité végétale remarquable dans l’Aude (France), le domaine de Peyremale, à Montolieu, et par extension de quelques sites voisins. Cinq associations nouvelles y sont définies : le Dioscoreo communis-Pyretum spinosae, l’Anacamptido laxiflorae-Serapiadetum linguae, le Ranunculo paludosi-Serapiadetum linguae, le Ranunculo paludosi-Isoetetum duriei, le Junco acutiflori-Bellevalietum romanae.
Abstract
This article provides some phytosociological data on the vegetation of a site with a remarkable plant diversity in the Aude departement (France), the Peyremale area, in Montolieu, and by extension some neighboring sites. Five new associations are defined: Dioscoreo communis-Pyretum spinosae, Anacamptido laxiflorae-Serapiadetum linguae, Ranunculo paludosi-Serapiadetum linguae, Ranunculo paludosi-Isoetetum duriei, Junco acutiflori-Bellevalietum romanae.
Le domaine de Peyremale (commune de Montolieu, Aude) est un des hauts lieux de la diversité végétale de ce département, mais il n’a à notre connaissance fait l’objet d’aucune étude fine des habitats élementaires. La présente monographie tente de remédier partiellement à ce défaut.
1. Généralités sur le domaine de Peyremale
1.1. Les exploitations agricoles
Organisation
Le domaine de Peyremale (photo 1) est situé sur la commune de Montolieu, de part et d’autre de la RD 629, à 20 km au nord-ouest de Carcassonne, dans le secteur du Cabardès-Montagne noire. Le site totalise environ 78 ha et abrite deux exploitations agricoles :
- l’élevage ovin viande géré par Annie et Jean-Pierre Pautou, installés depuis 1981 (72 hectares) ;
- un maraîchage bio créé par leur fils Émile en 2000 (6 hectares).
Les surfaces utilisées par le troupeau sont constituées de :
- 15 ha de zones boisées,
- 36 ha de prairies naturelles,
- 21 ha de pelouses.

Le troupeau compte actuellement 260 brebis allaitantes d’une race locale devenue très rare, la montagne noire (photo 2). Cette race ancienne d’origine audoise et tarnaise est connue pour sa productivité et la facilité de conduite de ses troupeaux. Un seul agnelage est réalisé annuellement, au printemps. Le troupeau est mené en parcs fixes sur le domaine quasiment toute l’année, sauf durant la période la plus froide de l’hiver où un affouragement lui est apporté en bergerie.

L’ensemble des parcelles fauchées représente 36 ha. Il s’agit majoritairement de prairies à graminées qui produisent un excellent foin, mais aussi de prairies hygrophiles à Juncus acutiflorus de moindre qualité fourragère (photo 3). Depuis 1981, ces prairies sont utilisées selon le cahier des charges de l’Agriculture biologique et n’ont fait l’objet d’aucun retournement. À l’automne, les parcelles prairiales les plus productives sont fertilisées en compost de fumier ovin issu de la bergerie. La productivité de ces herbages permet, outre le pâturage pendant la majeure partie de l’année, de réaliser une coupe par an. La période de récolte est assez variable en fonction des conditions météorologiques, se situant en général au début du mois de juin.

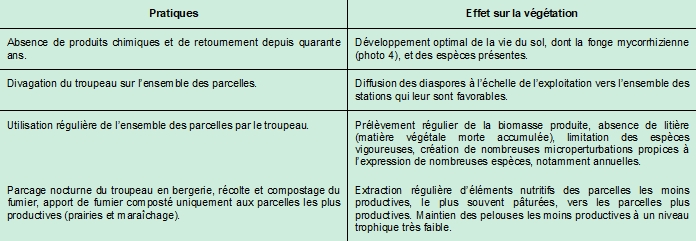
Aucun engrais chimique ni produit phytosanitaire n’y est utilisé. Ces pratiques contribuent à préserver une bonne qualité de l’eau des ruisseaux pour laquelle l’exploitation, située en tête de bassin versant, porte une responsabilité particulière. Le mode de gestion agricole peu intensif, avec très peu d’intrants notamment, a permis de maintenir et de développer une biodiversité exceptionnelle à l’échelle régionale, voire nationale.
Importance des pratiques agricoles pour la diversité végétale
Le tableau 1 présente quelques éléments clés permettant l’expression de la diversité floristique exceptionnelle du site.
Démarches de soutien à la gestion conservatoire du domaine
Depuis 2002, Mme et M. Pautou ont signé une convention de gestion avec le Conservatoire d’espaces naturels (CEN) du Languedoc-Roussillon, devenu récemment le CEN Occitanie. Dans le cadre de cette convention, les deux parties s’engagent à œuvrer pour maintenir et améliorer l’état de conservation du patrimoine naturel présent dans le site par une gestion agropastorale adaptée. Au cours des trois premières années de cette convention, le CEN a versé une contrepartie financière pour le manque à gagner dû à la baisse de récolte fourragère sur les parcelles conventionnées. En ce début des années 2000, la mise en place du réseau Natura 2000 démarrait à travers la désignation des premiers sites. Le porter à connaissance, par le CEN, des enjeux de biodiversité exceptionnels du domaine auprès des services de l’État a permis l’intégration de la majeure partie du site, côté ouest de la route départementale, dans le site Natura 2000 FR9101446 « Vallée du Lampy ».

Suite à cette première phase de soutien, le CEN a inscrit le domaine de Peyremale dans un volet du programme Semclimed financé par le fonds européen Feder. Ce cadre a permis la publication d’un outil de communication pertinent car visuel, une chemise de six fiches-habitats illustrées (Klesczewski & Bosca, 2008). À l’aide de cet outil et afin de trouver un soutien financier à cette agriculture très favorable à la biodiversité mais économiquement fragile, une discussion a été entamée avec les services de la Région Languedoc-Roussillon et du Département de l’Aude sur la possibilité de créer une réserve naturelle régionale sur le domaine de Peyremale. Cette réflexion n’a finalement pas abouti, mais l’idée d’un soutien financier à l’exploitation agricole a été retenue par les services du Département (voir aussi Klesczewski, 2012). Ces subventions ont en effet été attribuées pendant cinq ans, entre 2011 et 2015, et ont constitué un élément clé pour le maintien des pratiques très peu intensives et donc favorables à la biodiversité sur le domaine en cette période préalable à la mise en place des contrats Natura 2000. Le Département a également financé le suivi scientifique permettant de documenter l’atteinte des objectifs de gestion conservatoire (Klesczewski, 2011-2015).
Depuis 2017, la partie « élevage » du domaine bénéficie de « mesures agro-environnementales et climatiques » (MAEC), signées dans l’objectif de gestion conservatoire des habitats et espèces d’intérêt communautaire présents dans la partie du domaine située au sein du site Natura 2000.
1.2. Géologie et hydrologie
La diversité géologique du site explique une grande partie de la diversité de la végétation qui s’y développe (d’après Berger et al., 1993). Dans la partie sud, des formations de calcaires lacustres de Montolieu (Thanétien inférieur) jouxtent la vallée dans laquelle se sont accumulés des limons argileux fluviatiles (Dano-Montien ou Montien, ère tertiaire) formant des sols lourds et peu perméables. La partie nord, en revanche, est entièrement assise sur des roches siliceuses de type granite et gneiss (orthogneiss indifférenciés de Saissac, granites de Brousse) sur lesquelles se forment des sols acides. De manière générale, les sols sur les coteaux sont peu profonds, voire superficiels, d’où la présence de nombreux affleurements rocheux calcaires ou granitiques. À noter également que le domaine se situe sur une zone de contact géologique entre gneiss et granite. Dans cette zone de contact, ces roches ancestrales ont subi une recristallisation (« mylonitisation ») dynamique avec pour conséquences une texture très finement litée et une importante réduction de taille de grain (source : https://fr.wikipedia.org/wiki/mylonite). Notamment sur ce type de substrat géologique sont localisés de nombreux suintements temporaires parfois assez étendus.
Le domaine est situé sur deux bassins versants, celui du Fresquel (du Lampy à la Rougeanne) et celui du Lampy. La Vernassonnelle, petit cours d’eau temporaire en tête du bassin versant du Lampy, prend sa source sur le domaine et rejoint la Vernassonne au nord de l’abbaye de Villelongue ; la Vernassonnelle est alimentée par deux sources naturelles, qui forment des petits bassins relativement visibles, mais aussi par de nombreux suintements qui ne sont apparents qu’en période de pluie. À l’image des différents substrats géologiques, l’hydromorphie des sols très variable contribue à l’exceptionnelle diversité des milieux.
1.3. Climat et phytogéographie
La commune de Montolieu est située dans une zone de transition qu’on peut qualifier de « carrefour climatique ». On y constate des influences climatiques à la fois de type méditerranéen (hivers doux, étés chauds et secs, pluies parfois diluviennes) et atlantique (hivers doux, brouillards fréquents, précipitations plutôt régulières). La végétation reflète de façon très nette ces différentes influences : dans le domaine se côtoient des groupements typiquement méditerranéens (chênaies vertes, mares temporaires à Isoetes duriei) et atlantiques telles que les prairies à Juncus acutiflorus et Trocdaris verticillatum. Le site constitue donc pour certains types de végétation l’extrême limite d’aire de répartition vers le sud, pour d’autres vers le nord.
Selon le découpage de l’Aude en régions biogéographiques (Barreau et al., 2016), ce domaine se situe à la limite entre Montagne noire méditerranéenne occidentale et Montagne noire atlantique occidentale.
1.4. Valeur patrimoniale de la flore
La combinaison de tous les facteurs stationnels (géologie, climat) et des pratiques de l’éleveur respectueuses de la biodiversité a permis le maintien et le développement d’un nombre très important d’espèces végétales et animales. Plus de 630 taxons végétaux vasculaires ont été relevés sur le domaine. Trois sont protégés au niveau supra-départemental (annexe II de la directive Habitats-Faune-Flore, protection nationale, protection au niveau de l’ex-région Languedoc-Roussillon) : Bellevalia romana, Isoetes duriei, Kickxia commutata subsp. c. ; deux sont considérés comme menacés, donc inscrits sur la Liste rouge nationale : Bellevalia romana (NT), Ophrys bombyliflora (NT). Huit sont considérés comme d’intérêt patrimonial dans l’Aude (Plassart et al., 2016) : Chamaemelum nobile, Anacamptis coriophora, Bellevalia romana, Exaculum pusillum, Isoetes duriei, Kickxia commutata subsp. c., Neotinea lactea, Trifolium micranthum (photos 5 à 12). Serapias cordigera y a été introduit en provenance d’un site voisin où le taxon était menacé de disparition (photo 13, prise à Verdun-en-Lauragais).










1.5. Inclusion au moins partielle dans des zonages naturalistes
Le domaine est identifié comme espace naturel sensible par le Conseil départemental de l’Aude et inscrit comme ZNIEFF 910030412 (Prairies du domaine de Peyremale). Les deux tiers de l’exploitation sont situés sur le site Natura 2000 FR9101446 (Vallée du Lampy). À l’appui de cette dernière labellisation, il faut préciser la présence de plusieurs habitats d’intérêt communautaire (les prioritaires notés *), dont neuf sur Peyremale (nomenclature Corine-biotopes) :
- 3170* – « Mares temporaires méditerranéennes » ;
- 6210(*) – « Formations herbeuses sèches semi-naturelles et faciès d’embuissonnement sur calcaires (Festuco-Brometalia) » ;
- 6220* – « Parcours substeppiques de graminées et annuelles des Thero-Brachypodietea » ;
- 6410 – « Prairies à molinie sur calcaire et argile (Eu-Molinion) » ;
- 6420 – « Prairies méditerranéennes à hautes herbes et joncs (Molinio-Holoschoenion) » ;
- 6430 – « Mégaphorbiaies eutrophes » ;
- 6510 – « Prairies maigres de fauche de basse altitude (Alopecurus pratensis, Sanguisorba officinalis) » ;
- 91E0* – « Forêts alluviales à Alnus glutinosa et Fraxinus excelsior (Alno-Padion, Alnion incanae, Salicion albae» ;
- 9340 – « Forêts à Quercus ilex».
L’habitat 7220* [« Sources pétrifiantes avec formation de tuf (Cratoneurion) »] est signalé dans le site, mais ne s’y trouve que sous forme très fragmentaire.
2. Méthodes d’étude de la végétation
La reconnaissance de ces habitats d’intérêt communautaire en reste toutefois aux grandes catégories de l’annexe I de la directive européenne Habitats-Faune-Flore, c’est-à-dire aux habitats qualifiés habituellement de « génériques ». Il faut maintenant aller plus loin et aborder les habitats élémentaires, comment ces habitats génériques se concrétisent sur un site possédant des caractéristiques édapho-climatiques précises. Pour cela, il faut mettre en place l’outil phytosociologique.
L’étude de la végétation du domaine de Peyremale suit donc les grands principes méthodologiques de la phytosociologie sigmatiste, toutefois affinée par l’approche synusiale (de Foucault, 1986a ; Gillet et al., 1991). Le document d’objectifs du site Natura 2000 propose vingt-deux relevés phytosociologiques réalisés sur l’ensemble de son périmètre (Ivanez, 2012, annexe 5). Les relevés récents ont été acquis par l’un de nous (BdF) lors de deux journées de terrain sur le domaine, les 10 et 13 mai 2019 ; des relevés plus anciens réalisés par le second auteur (MK) sur plusieurs années ont été ajoutés pour aboutir à la typologie présentée ci-après. Parfois des relevés hors Peyremale ont été inclus ; d’une part c’est une occasion de les publier, d’autre part c’est une garantie sur la répétitivité des combinaisons floristiques qui sous-tend la définition des syntaxons, surtout du rang association. Sauf exceptions (surtout des relevés anciens), les surfaces relevées (en m²) et les recouvrements (en %) de la végétation sont indiqués, ainsi que l’altitude (m), la latitude et la longitude (coordonnées sexagésimales).
La nomenclature des taxons suit Flora Gallica (Tison & de Foucault, 2014). Les épithètes subspécifiques des taxons autonymes sont réduites à leur initiale pour gagner de la place (exemple : Rubia peregrina subsp. p. au lieu de Rubia peregrina subsp. peregrina) ; pour la même raison, dans les tableaux, le signe * remplace ‘subsp.’ ou ‘var.’. Le signe ‘j’ qualifie un taxon arborescent ou arbustif représenté par des individus juvéniles dans les synusies inférieures, ‘!’ désigne un taxon représenté par des individus plus vigoureux que la moyenne.
Parfois l’approche systémique a été introduite pour représenter les relations spatio-temporelles entre syntaxons, avec les symboles suivants :

3. Résultats phytosociologiques
3.1. Le système de niveau supérieur
Dans les niveaux topographiques supérieurs, le système comporte des boisements, des fourrés, diverses prairies à Serapias et des pelouses thérophytiques.
Les boisements
Les boisements n’occupent qu’une assez faible surface sur le domaine ; ils n’ont guère d’usage pastoral, quoique parfois ouverts au pâturage extensif.
L’association arborescente combine essentiellement Quercus ilex, Q. pubescens, Fraxinus angustifolia subsp. a. (tableau 2 ; tous les relevés viennent de Peyremale ; photo 14) et peut donc être rattachée au Quercetum pubescenti-ilicis dans une forme hygrophile à F. angustifolia subsp. a.
Nous passons sur la végétation arbustive intraforestière, mal caractérisée, pour aborder la végétation herbacée intraforestière. Les types biologiques y sont diversifiés, de sorte que nous pouvons séparer une végétation vivace et une végétation plutôt annuelle.

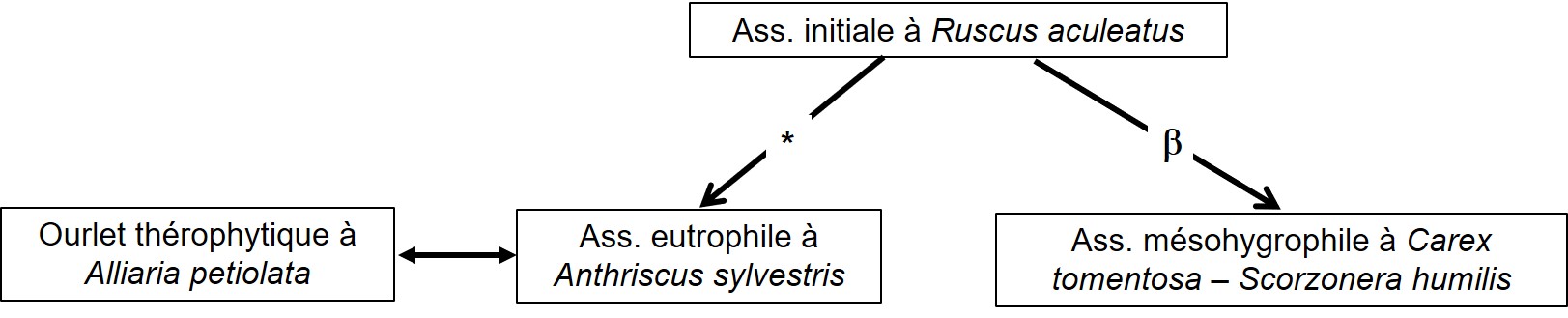
La végétation vivace est illustrée par les six relevés du tableau 3 (tous viennent de Peyremale). On peut y séparer deux syntaxons partageant essentiellement Arum italicum, Ruscus aculeatus et Dioscorea communis, plus rarement Euphorbia amygdaloides subsp. a. et Oenanthe pimpinelloides. Le premier syntaxon (relevés 1 à 4) décrit une association plutôt naturelle à Ficaria verna subsp. ficariiformis, Brachypodium sylvaticum, Ajuga reptans, Hedera helix, avec une variante à Luzula forsteri et une variante plus remarquable à Carex tomentosa, C. leporina var. l., C. pallescens, C. umbrosa subsp. u., C. flacca subsp. f., Scorzonera humilis…, développée en léger contrebas du relevé 1, qui évoque le résultat de l’humidification d’une synusie initialement mésophile et qui s’apparente à un groupement du Molinion caeruleae acidiphile.
Le deuxième syntaxon (relevés 5 et 6), plus eutrophile, est caractérisée par Geum urbanum, Rumex sanguineus, Anthriscus sylvestris subsp. s. ; il paraît dériver de la synusie initiale par eutrophisation consécutive au pâturage sous forêt.
La végétation thérophytique est décrite par les trois relevés du tableau 4 (provenant tous de Peyremale). Elle est surtout associée au syntaxon vivace eutrophile : en effet, l’eutrophisation induite par le pâturage sous forêt, en plus de modifier la végétation vivace, ouvre celle-ci et permet la structuration d’une mosaïque vivace/annuelle, la végétation thérophytique restant encore fragmentaire pour les relevés 1 et 2, mieux caractérisée dans le cas du relevé 3, caractérisant alors un ourlet interne en nappe à Alliaria petiolata, Geranium purpureum, G. rotundifolium, Anisantha rigida, Lapsana communis subsp. c., Veronica hederifolia, Torilis africana var. heterophylla, intermédiaire entre Valantio-Galion muralis thermophile et Drabo-Cardaminion hirsutae plus psychrophile. Le graphe de la figure 1 illustre cette dynamique intraforestière ; malheureusement, le nombre de relevés est trop réduit pour envisager la définition d’associations originales.
Il faudrait poursuivre les investigations dans les boisements autour de Montolieu, si possible dépourvus de perturbations d’origine biotique, pour statuer sur cette végétation.
Les fourrés
Çà et là des fourrés héliophiles se développent à l’extérieur des boisements ou forment des haies limitant des parcelles. Le tableau 5 en rapporte sept relevés de Peyremale (nos 1 à 4, 6, 8 et 9) plus trois de Villardonnel (« Les Combals », nos 5, 7 et 10, réalisés le 26 mai 2020). Globalement, ils partagent Pyrus spinosa, Crataegus monogyna, Prunus spinosa, Rosa canina, Dioscorea communis, Rubus ulmifolius et de jeunes arbres de la végétation arborescente déjà présentée (Fraxinus angustifolia subsp. a., Quercus ilex, Q. pubescens), le taxon le plus original étant le premier de cette liste. Un tel fourré paraît inédit ; il est assez distinct du Lonicero etruscae-Pyretum spinosae décrit des Rougiers de Camarès (de Foucault, 2016a ; Kessler et al., 2019) et peut être distingué sous le nom de Dioscoreo communis-Pyretum spinosae ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 5 hoc loco. Il accueille en moyenne près de 10 taxons par relevé et présente deux variations, en plus du typicum (relevés 1 à 5) : une provisoire plus eutrophile à Sambucus nigra et Bryonia dioica (relevés 6 et 7), l’autre plus oligotrophile (relevés 8 à 10) différenciée par Erica scoparia subsp. s., E. arborea et Cytisus scoparius subsp. s. (ericetosum scopariae subass. nov. hoc loco, typus nominis hoc loco : relevé 8 du tableau 5 hoc loco ; photo 15). Le Dioscoreo communis-Pyretum spinosae peut se ranger dans le Pruno-Rubion ulmifolii O. Bolòs 1954, comme le Lonicero etruscae-Pyretum spinosae (de Foucault & Royer, 2016).

On peut finir ce thème en évoquant un ourlet thérophytique vernal externe assez distinct de l’ourlet interne à Alliaria petiolata car caractérisé par Calepina irregularis, Anthriscus caucalis var. c., Geranium molle, G. purpureum, G. rotundifolium, Galium aparine, Scandix pecten-veneris subsp. p., Anisantha sterilis, qui évoque le Valerianello carinatae-Calepinetum irregularis (de Foucault, 1988a, 2016a).
Les pelouses vivaces à Serapias
Des pelouses à Serapias, dans les tableaux 6 et 7, nous avons rassemblé 17 relevés, dont sept réalisés en dehors du domaine de Peyremale (Villardonnel, Labécède-Lauragais, Verdun-en-Lauragais, Saissac ; voir annexes des tableaux 6 et 7). Nos analyses statistiques ont permis de séparer deux syntaxons.
Le premier syntaxon (tableau 6 ; tous les relevés viennent de Peyremale, sauf nos 5 à 9) rassemble neuf relevés. Outre Serapias lingua, ils partagent Anacamptis morio subsp. m., A. laxiflora, Linum usitatissimum subsp. angustifolium, Eryngium campestre, Poterium sanguisorba s. l., Galium verum subsp. v., Carex flacca subsp. f., C. caryophyllea, Oenanthe pimpinelloides, Gaudinia fragilis, Avenula pubescens subsp. p., Luzula campestris subsp. c., Urospermum dalechampii, Ranunculus bulbosus… Le tableau comparatif 8 montre que le présent syntaxon (colonnes ASl1 et ASl2) se sépare bien du suivant (colonnes RSl) par la présence d’Anacamptis morio subsp. m., Lotus corniculatus… et l’absence de Ranunculus paludosus, Rumex acetosella, Cynodon dactylon… Ce premier point acquis, il nous faut encore tester l’identité éventuelle de notre syntaxon avec l’Anacamptido morionis-Serapiadetum linguae décrit de régions moins méditerranéennes : Armagnac (de Foucault, 1986b, tableau 5, sub Orchido morionis-Serapiadetum linguae, synthétisé ici en Arm.), basse Auvergne (Billy, 2000, sub Scorzonero humilis-Serapiadetum linguae, tableau VI : syntaxon II-6, Auv.), Limousin (Mady et al., 2018, tableau 2, sub Orchido morionis-Serapiadetum linguae, Lim.). La comparaison (tableau 6, colonnes ASl1 et ASl2 vs colonnes Arm., Auv., Lim.) montre que l’Anacamptido–Serapiadetum linguae se différencie positivement par Leucanthemum ircutianum, Pilosella officinarum, Daucus carota subsp. c., Briza media subsp. m., Polygala vulgaris subsp. v., Rumex acetosa subsp. a., Lathyrus pratensis, Ajuga reptans, Primula veris subsp. v., Neotinea ustulata…, alors que notre pelouse se distingue par Eryngium campestre, Urospermum dalechampii, Avenula pubescens subsp. p., Anacamptis laxiflora, plus rarement Neotinea maculata, Ophrys exaltata. L’Anacamptido–Serapiadetum linguae se différencie alors en races de l’Armagnac (Arm., à Galium verum subsp. v., Plantago media subsp. m., Coeloglossum viride, Scabiosa columbaria…), de basse Auvergne (Auv., à Nardus stricta, Centaurea jacea…), du Limousin (Lim.) ; plusieurs taxons hygrophiles oligotrophiles comme Succisa pratensis et Scorzonera humilis en Arm. et Auv. semblent plutôt liés à une dérive par humidification édaphique de pelouses à Anacamptis morio-Serapias lingua initialement plus mésophiles. De toutes ces comparaisons, nous pouvons tirer la conclusion de l’originalité de notre pelouse que l’on peut décrire sous le nom de Anacamptido laxiflorae-Serapiadetum linguae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 6 hoc loco. Cette pelouse se développe sur des substrats acidiclines assez profonds sous climat méditerranéen atténué. À côté de la variation typicum (tableau 6 : relevés 1 à 5), accueillant en moyenne 26 taxons par relevé, on peut dégager une variation significative mais statistiquement et écologiquement encore mal définie (relevés 6 à 9) à Achillea millefolium, Malva moschata, Arrhenatherum elatius subsp. e., Poa trivialis subsp. t., Stellaria graminea, Rumex acetosa subsp. a., Filipendula vulgaris, Ophioglossum vulgatum, Rorippa pyrenaica…, floristiquement très riche (près de 48 taxons par relevé en moyenne). Outre à Peyremale, cette association a été observée à Villardonnel, Labécède-Lauragais, Verdun-en-Lauragais et Saissac. Divers aspects restent encore à préciser sur cette belle association audoise.
La seconde pelouse à Serapias lingua est décrite par les huit relevés du tableau 7 (tous les relevés viennent de Peyremale, sauf les nos 2 et 5). Sur la base du tableau comparatif 8, dans le paragraphe précédent, nous avons déjà évoqué la bonne différenciation entre ce qui a été maintenant reconnu comme Anacamptido laxiflorae-Serapiadetum linguae et ce second syntaxon, statistiquement lui-même bien défini. Cette pelouse peut être décrite sous le nom nouveau de Ranunculo paludosi-Serapiadetum linguae ass. nov. hoc loco, typus nominis hoc loco : relevé 4 du tableau 7 hoc loco (photo 16). Cette association se développe sur des substrats acidiclines nettement plus minces que ceux colonisés par l’Anacamptido l.-Serapiadetum linguae. Outre à Peyremale, cette pelouse a aussi été observée à Villardonnel et Labécède-Lauragais.

À ces deux associations à Serapias lingua, il faut ajouter la rencontre sur des substrats plus basiques (partie sud du domaine, sur les calcaires lacustres) de l’Urospermo dalechampii-Serapiadetum vomeraceae récemment décrit (de Foucault, 2018) :
N 43° 19’ 22,4’’, E 2° 11’ 39,6’’, 275 m, le 13 mai 2019, 35 taxons :
- Anacamptido p.-Serapiadion vomeraceae Foucault 2019 et Festuco-Brometea erecti: Serapias vomeracea 2, Ophrys apifera +, O. aranifera *a. +, Poa bulbosa *b. 2, Leontodon hispidus *h. 1, Anacamptis pyramidalis *p. +, Urospermum dalechampii 2, Eryngium campestre 1, Carex caryophyllea 2, C. flacca *f. 3, Galium verum *v. 1, Ononis spinosa *procurrens 1, Cirsium acaulon 1,
- autres taxons : Bellis perennis 2, Gaudinia fragilis 2, Trifolium dubium 2, repens *r. 2, T. pratense *p. 2, Avenula pubescens *p. 1, Luzula campestris *c. 2, Leucanthemum ircutianum 1, Serapias lingua 2, Anthoxanthum odoratum 2, Anacamptis laxiflora 1, A. morio *m. +, Hypochaeris radicata 1, Daucus carota *c. 2, Schedonorus pratensis *p. 1, Briza media *m. 1, Poa trivialis *t. 1, P. pratensis *p. 1, Plantago lanceolata 2, Rhinanthus minor +, Juniperus communis *c. j +, Prunus spinosa j +.
La présence de Serapias lingua et Anacamptis morio différencie une variation sur substrat plus acidicline (déjà évoquée lors de la description initiale de l’association). En outre, dans le second ensemble, plusieurs taxons prairiaux témoignent d’influences biotiques significatives (pâturage extensif). L’intensification de ce facteur biotique aboutit à la dérive de l’Urospermo d.-Serapiadetum vomeraceae vers une prairie à Trisetum flavescens, le Serapiado vomeraceae-Trisetetum flavescentis B. Foucault 2019 (de Foucault, 2018), mais celle-ci n’a pas été rencontrée à Peyremale.
En complément de cette présentation des pelouses à Serapias, ajoutons que les landes sont fort rares sur le domaine étudié, se réduisant souvent à des fragments à Lavandula stoechas subsp. s., Calluna vulgaris et Cistus salviifolius. Potentiellement, elle devrait correspondre au Lavandulo stoechadis-Ericetum cinereae Géhu ex B. Foucault 2021, au sein du Cisto salviifolii-Ericion cinereae Géhu in Bardat et al. 2004, alliance des Calluno-Ulicetea minoris faisant le passage vers les Cisto-Lavanduletea stoechadis (de Foucault, 2020, 2021).
Annexe au tableau 6 – rel. 5 : Villardonnel, « Les Combals », 20/05/2018 ; rel. 6 : Labécède-Lauragais, au-dessus du ruisseau des Glandes, 13/05/2017 ; rel. 7 : Verdun-en-Lauragais, « Jean-Raymond » près de « Puget-Haut », 17/05/2015 ; rel. 8 et 9 : Saissac, près de « La Garrigue », 14/05/2014.
Annexe du tableau 7 – rel. 2 : Villardonnel « Les Combals », 20/05/2018 ; rel. 5 : Labécède-Lauragais, 13/05/2017.
La pelouse vivace mésohygrophile à Isoetes duriei
Sur les substrats retenant de l’eau en hiver et en début de printemps, des taxons, dont de plus hygrophiles, s’introduisent dans la pelouse initiale à S. lingua : Isoetes duriei, Festuca rubra subsp. r., Anacamptis laxiflora, Danthonia decumbens subsp. d., Carex caryophyllea, Bellis perennis, Oenanthe pimpinelloides, Kickxia commutata subsp. c., Fraxinus angustifolia subsp. a. j, Ophioglossum vulgatum… ; nous la décrivons sous le nom nouveau de Ranunculo paludosi–Isoetetum duriei ass. nov. hoc loco (tableau 9, avec seize relevés de Peyremale), typus nominis hoc loco : relevé 7 du tableau 9 hoc loco.
L’hétérogénéité apparente de cet ensemble floristique différentiel semble liée à l’influence conjointe d’au moins deux facteurs écologiques, l’un étant une épaisseur plus élevée du substrat (indiquée par Ranunculus bulbosus, Galium verum subsp. v., Anacamptis laxiflora, Carex caryophyllea, Oenanthe pimpinelloides…, transition vers l’Anacamptido l.–Serapiadetum linguae), l’autre étant une humidification édaphique (surtout indiquée par Isoetes duriei, Ophioglossum vulgatum, Kickxia commutata subsp. c.). On peut y séparer une variante optimale (relevés 1 à 12), avec 22,6 taxons par relevé en moyenne, et une variante appauvrie (relevés 13 à 16), avec 15 taxons par relevé en moyenne. Le tableau synthétique 10 décrit la série d’humidification édaphique menant du Ranunculo paludosi-Serapiadetum linguae (colonne RSl) au Ranunculo paludosi-Isoetetum duriei (colonnes RId1 et RId2).
Notons que cette association de Isoetes duriei et Ophioglossum vulgatum offre un nouvel exemple de la structure formelle discutée par l’un de nous (de Foucault, 1988b : 54, § Le rôle des Ophioglosses). Dans cette publication, on a cité plusieurs cas d’association entre des Ophioglossum (en France : surtout O. lusitanicum et O. azoricum ; en Côte-d’Ivoire : O. gomezianum, O. costatum ; en Guyane française : O. nudicaule ; en Afrique du Sud : O. bergianum, O. nudicaule) et des Isoetes de niveau topographique moyen (en France : I. histrix, I. duriei ; en Côte-d’Ivoire : I. nigritiana ; en Afrique du Sud : I. stellenbossiensis). Une publication récente (Patel & Reddy, 2019) vient de révéler un nouveau cas pour l’Inde (État du Gujarat), avec O. hitkishorei et un Isoetes malheureusement non nommé, associés à Pogostemon deccanensis et un taxon dénommé « Euriculon sp. » (à notre connaissance ce genre n’existe pas, sans doute une erreur d’écriture pour Eriocaulon).
Place synsystématique des pelouses vivaces à Serapias lingua et à Isoetes duriei
Le tableau 11 permet de discuter de la place synsystématique des pelouses vivaces à Serapias lingua. On peut en effet hésiter à les placer dans le Danthonio decumbentis-Serapiadion linguae B. Foucault 2012, au sein des Nardetea strictae (de Foucault, 2012) ; dans ce tableau 11, la colonne DSl synhétise les trois associations connues de cette alliance (de Foucault, 2012, tableau 1 : colonne A.1.9), qui partage plusieurs taxons avec le reste des Nardetea strictae (Veronica officinalis, Pilosella officinarum, Coeloglossum viride, Carex pilulifera, Luzula multiflora, Potentilla erecta, Agrostis capillaris), peu ou pas représentés ici. On comprend ainsi que le Danthonio-Serapiadion constitue bien une des fins des Nardetea strictae vers le sud.
Il nous reste alors la possibilité de les placer dans les Serapiadetea cordigero-linguae B. Foucault 2012 et les Serapiadetalia cordigero-linguae B. Foucault 2012, au sein de la seule alliance actuellement définie, le Serapiadion cordigero-neglectae B. Foucault 2012 (le premier des taxons éponymes d’alliance est présent, mais rare, en Montagne noire audoise ; Plassart et al., 2016 : 338). Dans cette unité, seulement deux associations sont véritablement bien définies (d’autres groupements sont évoqués, à consolider dans le futur), le Simethido mattiazzii-Serapiadetum linguae B. Foucault, Paradis & Pozzo di Borgo in B. Foucault 2012 (Corse méridionale) et l’Agrostio castellanae-Nardetum strictae Laribi, Acherar, B. Foucault, Mathez & Derridj 2017 (massif de l’Akfadou, Kabylie algérienne ; Laribi et al., 2017, tableau 1), respectivement synthétisés ici en SSl et ANs. L’altitude de la nardaie kabyle (entre 1 300 m et 1 400 m) a pour effet le retour de Nardus stricta dans une pelouse acidiphile. Encore plus haut dans les massifs nord-africains (Atlas, Djurdjura, Aurès, au-dessus de 2 500 m), les pelouses à Nardus stricta se généralisent au point d’avoir suscité la création des Udo-Nardetalia strictae Quézel 1953 (Mem. Soc. Brot. 9 : 49) nom. illeg. (art. 34), qui incluent le Plantaginion nivalis Quézel 1953 de la Sierra Nevada ibérique et le Trifolion thalii Quézel 1952 d’Afrique du Nord.
Quant au Ranunculo paludosi-Isoetetum duriei, il paraît difficile de le rattacher à l’Ophioglosso lusitanici-Isoetion histricis B. Foucault 1988, qui réunit des associations très spécialisées et souvent paucispécifiques de mares temporaires méditerranéo-atlantiques (de Foucault 1988b, 2013) ; on exprime ainsi un désaccord avec l’interprétation de cet habitat sous le code 3170* (cf. supra § 1.5). Le plus simple est sans doute de placer cette pelouse nettement plus riche en espèces aussi dans le Serapiadion cordigero-neglectae (tableau 11 : colonne RId), en attendant que de nouveaux travaux de terrain améliorent la définition et la structuration synsystématique des Serapiadetea cordigero-linguae. II est probable que les trois associations de Peyremale et la nardaie kabyle se placeront à l’avenir dans des alliances distinctes du Serapiadion cordigero-neglectae, l’une de transition vers les Nardetalia strictae, l’autre de transition vers les Udo-Nardetalia strictae.
Les pelouses thérophytiques
Pour les pelouses à thérophytes, qui entrent en superposition des pelouses vivaces précédentes dans leurs ouvertures, 32 relevés ont été réunis dans le tableau 12. Ils partagent globalement Aira caryophyllea, Vulpia bromoides, Sherardia arvensis, Myosotis discolor, Moenchia erecta, Briza minor, Trifolium subterraneum… Le traitement statistique permet de dégager plusieurs groupements.
Les relevés 1 et 2, à Crassula tillaea et Polycarpon tetraphyllum subsp. t., ont été réalisés sur les auréoles entourant les affleurements rocheux granitiques qui transpercent les pelouses développées sur sol plus profond à Peyremale ; ils accueillent une dizaine de taxons en moyenne.
Les relevés 3 à 6 (3 et 4 de Peyremale ; 5 de Labécède-Lauragais, le 17/05/2017 ; 6 de la forêt communale de Cailhau, vers « À Jordanac », le 18/05/2014) décrivent une pelouse caractérisée par la combinaison de Tuberaria guttata, Logfia gallica, Filago germanica, Ornithopus compressus, Aphanes australis, Hypochaeris glabra…
Les relevés 7 à 16 (tous de Peyremale) décrivent plutôt une pelouse à Trifolium incarnatum subsp. molinerii, T. striatum, T. strictum, Silene gallica, Aphanes australis, Hypochaeris glabra, Parentucellia latifolia, Aira cupaniana… (photo 17), que l’on retrouve aussi parmi les données de Billy (2000, tableau II : syntaxon 1-2-3 pour B1 ; tableau IV : syntaxon 1-2-7 pour B2 ; Auvergne) et Klesczewski (2000, tableau 3 pour K ; mont Aigoual). Statistiquement bien défini, ce syntaxon mérite sans aucun doute le rang d’association, mais nous préférons attendre une synthèse moderne des Tuberarietea guttatae pour statuer définitivement. Les relevés 12 à 16 correspondent à une petite variation mésohygrophile à Juncus bufonius subsp. b. et J. capitatus. Ils accueillent une quinzaine de taxons en moyenne. Un relevé voisin mais réalisé à une altitude bien supérieure (Pyrénées audoises, 1 124 m) a été publié par l’un de nous (de Foucault, 2017 : 193).

Les relevés 17 à 31 (de Peyremale) se différencient surtout par Leontodon saxatilis subsp. s., Trifolium micranthum et Medicago polymorpha. La moyenne du nombre de taxons par relevé tombe ici à 9,5. Comme précédemment, les relevés 30 et 31 correspondent à une petite variation mésohygrophile à Juncus bufonius subsp. b. et J. capitatus.
Enfin, dans le relevé 32, de Villardonnelle (« Les Combals », le 20/05/2018), le lot des taxons xérophiles diminue bien, alors que s’étoffe le groupe des taxons plus hygrophiles (Hypericum humifusum, Montia arvensis, Lysimachia minima, Juncus capitatus) ; ce groupement relève plutôt des Nanocyperetalia flavescentis (de Foucault, 2013).
La garrigue à Genista hispanica
Vers le sud-est du domaine, sur un substrat plutôt basique (calcaires lacustres) et quelque peu marneux, on peut observer une garrigue à Genista hispanica subsp. h., dont trois relevés sont réunis dans le tableau 13 (tous provenant de Peyremale), qui accueillent en moyenne 25 taxons. Outre le genêt éponyme, ils partagent Carlina hispanica subsp. h., Bromopsis erecta subsp. e., Thymus vulgaris subsp. v., Eryngium campestre, Poterium sanguisorba s. l., Poa bulbosa subsp. b., Lavandula latifolia, Ophrys exaltata, Polygala vulgaris subsp. v., Pilosella officinarum, Catananche caerulea, Carex flacca subsp. f., Cirsium acaulon, Urospermum dalechampii, Aphyllanthes monspeliensis, Globularia bisnagarica (photo 18). Ce syntaxon se rapproche un peu du Koelerio macranthae-Thymetum vulgaris décrit de la Malepère (de Foucault, 2018), mais s’en distingue par la présence significative de G. hispanica ; de nouveaux relevés dans la région de Montolieu sont nécessaires pour statuer sur cette garrigue qui semble bien se ranger dans l’Helianthemo–Aphyllanthion monspeliensis.

En mosaïque avec le relevé 2, il existe une pelouse thérophytique assez bien structurée (9 taxons) : Brachypodium distachyon 2, Vulpia unilateralis 1, V. ciliata *c. 1, Trifolium campestre 1, Catapodium rigidum 1, Veronica arvensis 1, Euphorbia exigua +, Helianthemum salicifolium +, Cerastium pumilum +, à rattacher au Brachypodion distachyi.
Cette garrigue à G. hispanica semble en relation avec un pré oligotrophile à Schoenus nigricans (N 43° 19’ 22,8’’, E 2° 11’ 11,3’’, 268 m, 22 taxons ; photo 19) :
- Molinion caeruleae Koch 1926 et unités supérieures : Molinia caerulea 3, Lotus maritimus 2, Carex panicea 2, Ophioglossum vulgatum 2, Scorzonera humilis 2, Schoenus nigricans 1, Juncus acutiflorus 1, J. subnodulosus 2, Agrostis canina *c. 2, Lysimachia tenella 1, Galium debile +,
- autres taxons : Anacamptis laxiflora 1, Anthoxanthum odoratum 2, Ranunculus acris 1, Pulicaria dysenterica +, Briza media *m. +, Carex flacca *f. 2, Galium verum *v. 1, Bellis perennis +, Lathyrus pratensis 2, Serapias lingua 2, Fraxinus angustifolia *a. j +, Salix atrocinerea j +.
Elle est originale par son mélange de flore plutôt basiphile et de flore plutôt acidiphile.

3.1. Le système hygrophile
La topographie diversifiée du domaine de Peyremale engendre un système de végétation hygrophile dans les bas niveaux.
Souvent, à la base des systèmes hygrophiles, il existe une mégaphorbiaie plus ou moins naturelle. À Peyremale, celle-ci est mal structurée et se réfugie à l’abri des influences biotiques, notamment en lisière des saulaies arbustives à Salix atrocinerea. Elle est surtout caractérisée par Cirsium monspessulanum subsp. m., Scirpoides holoschoenus subsp. h., Lythrum salicaria, Solanum dulcamara, Rumex conglomeratus, Scrophularia auriculata et relève du Loto recti-Rumicion conglomerati Gradstein & Smittenberg 1977 (de Foucault, 2011, sub Dorycnio recti-Rumicion conglomerati).
Les niveaux les plus bas, où l’eau affleure et même coule nettement, sont occupés par une prairie inondable à rattacher aux Deschampsietalia cespitosae (de Foucault & Catteau, 2012) ; le tableau 14 rapproche trois relevés de Peyremale. On y note l’importance du groupe des taxons oligotrophiles amphibies, notamment Galium debile, Agrostis canina, Veronica scutellata, Ranunculus flammula, Trocdaris verticillatum… (photo 20) qui témoignent d’une évolution possible vers un bas-marais oligotrophile amphibie ; il existe là une homologie certaine avec des systèmes amphibies plus septentrionaux (de Foucault, 1984 : 438) où apparaissent aussi Galium debile, Veronica scutellata, Ranunculus flammula…

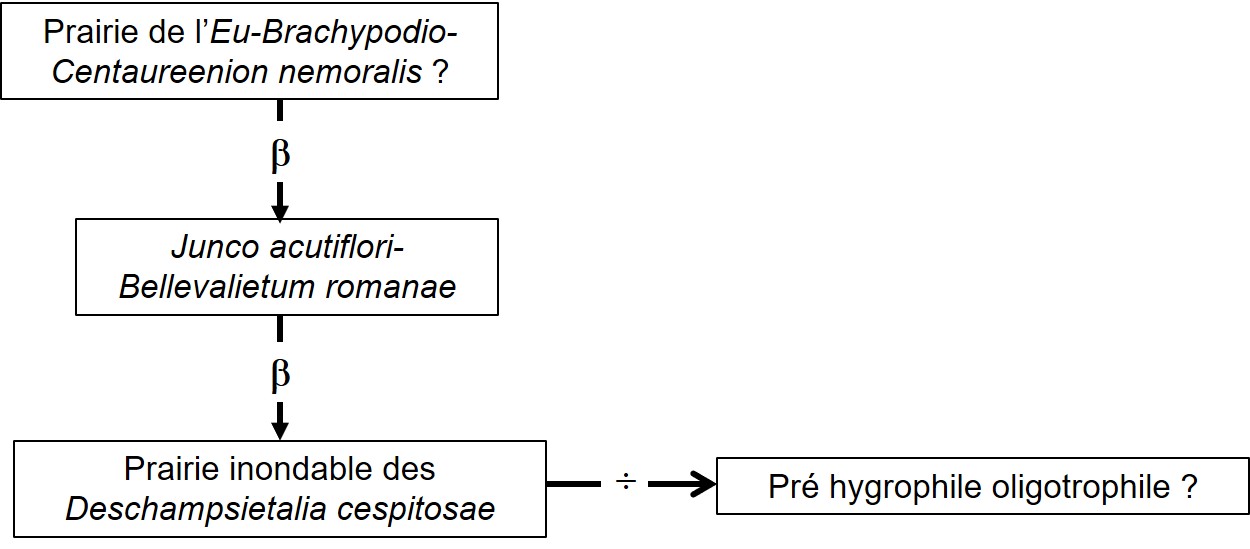
Les niveaux moyens, moins inondables car marqués par une eau faiblement affleurante, sont occupés par un autre type de prairie dont le tableau 15 rapporte deux relevés de Peyremale (nos 1 et 2) ; pour information, nous avons ajouté le relevé 3 réalisé à Fraisse-Cabardès, près des Bouriettes, au nord de la RD 203, dans la vallée du ruisseau de Laborde. Les relevés 1 et 2 décrivent une prairie combinant essentiellement Bromus racemosus, Trifolium squamosum, T. patens, Carex distans, Narcissus tazetta, Juncus acutiflorus, Bellevalia romana, Anacamptis laxiflora, Lychnis flos-cuculi subsp. f. Les deux premiers taxons apparaissent aussi dans la prairie de Fraisse-Cabardès, mais dans une tout autre combinaison, nettement en faveur des prairies mésohygrophiles du Brachypodio rupestris-Centaureion nemoralis et du Lino angustifolii-Oenanthenion pimpinelloidis (de Foucault, 2016b), sous-alliance maintenant à dénommer plutôt Eu-Brachypodio rupestris-Centaureenion nemoralis. Tout ceci paraît induire d’une part que la prairie du Bromion racemosi pourrait potentiellement exister dans le système du ruisseau de Laborde (d’autant plus que Bellevalia romana y est signalé ; Plassart et al., 2016) et d’autre part que la prairie mésohygrophile du Brachypodio-Centaureion nemoralis pourrait exister à Peyremale (mais nous ne l’avons pas observée). Ces remarques et le tableau 16, synthétisant 22 syntaxons du Bromion racemosi (d’après de Foucault & Catteau, 2012, tableau 10 actualisé ; les colonnes 91 à 108 renvoient aux syntaxons de cette alliance dans le tableau 10 cité ; la colonne JBr, entre les syntaxons 94b et 95, synthétise les deux relevés de notre tableau 15), nous incitent à créer une association végétale originale, une prairie hygrophile plutôt eutrophile au sein du Bromion racemosi, le Junco acutiflori-Bellevalietum romanae ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 15 hoc loco, accueillant en moyenne 28 taxons par relevé (photo 21). On remarque aussi quelques taxons caractérisant le Molinion caeruleae, qui incitent à poser l’hypothèse de l’existence d’une prairie hygrophile plus oligotrophile à Carex tomentosa de cette alliance. Rappelons qu’il a été observé une telle « prairie » à C. tomentosa dans un sous-bois de Peyremale (cf. supra, § 3.1).

Ce pré hygrophile oligotrophile est bien présent dans le système de Peyremale ; le tableau 17 en rapporte quatre relevés. Si Carex tomentosa n’y a pas été rencontré (ce taxon est rare dans la région), Carex panicea, Galium debile et Ophioglossum vulgatum y sont représentés, en association avec Scorzonera humilis, Agrostis canina subsp. c., Trodaris verticillatum, Carex leporina var. l., Juncus conglomeratus, Succisa pratensis (photo 22). Il s’agit certainement d’une association originale du Juncion acutiflori, mais les taxons prairiaux des Agrostienea stoloniferae restent encore trop bien représentés pour que ce syntaxon puisse se ranger dans cette dernière alliance. Il faudrait poursuivre l’étude de ce pré oligotrophile hygrophile.

Le graphe de la figure 2 synthétise les relations et les éléments, réellement observés ou hypothétiques, au sein de ce système prairial hygrophile.
4. Conclusion
Cette étude ne constitue qu’une première contribution à la description phytosociologique du domaine de Peyremale tant ce site est riche et diversifié. Elle a permis de décrire cinq associations végétales originales : le Dioscoreo communis-Pyretum spinosae (relevant du Pruno-Rubion ulmifolii), l’Anacamptido laxiflorae-Serapiadetum linguae, le Ranunculo paludosi-Serapiadetum linguae, le Ranunculo paludosi-Isoetetum duriei (trois associations que l’on propose de placer au voisinage du Serapiadion cordigero-neglectae) et le Junco acutiflori-Bellevalietum romanae (Bromion racemosi). L’Urospermo dalechampii-Serapiadetum vomeraceae a aussi été rencontré. D’autres syntaxons restent en attente de statut syntaxonomique définitif, surtout les pelouses à thérophytes et les prairies inondables à Galium debile. L’intégration de données extérieures au domaine permet de justifier la répétitivité des associations décrites et de penser que ce site pourrait s’avérer un modèle phytosociologique à étendre à des milieux naturels s’étendant sur le piémont du revers sud de la Montagne noire (Saissac, Villardonnel, Verdun-en-Lauragais, Labécède-Lauragais entre autres).
Bibliographie
Barreau D., Plassart C. & Vizcaïno D., 2016. Régions biogéographiques. In C. Plassart, D. Barreau & F. Andrieu (coord.), Atlas de la flore patrimoniale de l’Aude, Biotope, Mèze : 63-81.
Berger G.M., Debat P., Demange M., Issard H., Perrin M., Boyer F., Freytet P. & Mazéas H., 1993. Notice explicative de la feuille de Carcassonne (1037), Carte géologique de la France (1/50000). Orléans : BRGM, Orléans, 78 p.
Billy F., 2000. Prairies et pâturages en Basse-Auvergne. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 20 : 1-258.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’État, Rouen, 675 p.
Foucault B. (de), 1986a. La phytosociologie sigmatiste : une morpho-physique. Lille, 147 p.
Foucault B. (de), 1986b. Contribution à une étude phytosociologique des systèmes prairiaux hygrophile et mésophile de l’Armagnac méridional (Hautes-Pyrénées et Gers, France). Documents phytosociologiques, NS, X (1) : 221-254.
Foucault B. (de), 1988a. Contribution à la connaissance phytosociologique des corniches rocheuses de la vallée de l’Argenton, entre Argenton-Château et Massais (Deux-Sèvres). Bulletin de la Société botanique du Centre-Ouest, NS, 19 : 39-64.
Foucault B. (de), 1988b. Les végétations herbacées basses amphibies : systémique, structuralisme, synsystématique. Dissertationes Botanicae 121 : 1-150.
Foucault B. (de), 1994. Essai synsystématique sur les pelouses sèches acidophiles (Nardetea strictae, Caricetea curvulae). Colloques phytosocologiques XXII, Syntaxonomie typologique des habitats : 431-454.
Foucault B. (de), 2011. Contribution au prodrome des végétations de France : les Filipendulo ulmariae-Convolvuletea sepium Géhu & Géhu-Franck 1987. Journal de botanique de la Société botanique de France 53 : 73-137.
Foucault B., 2012 (de). Contribution au prodrome des végétations de France : les Nardetea strictae Rivas Goday in Rivas Goday & Rivas-Mart. 1963. Journal de botanique de la Société botanique de France 59 : 241-344.
Foucault B. (de), 2013. Contribution au prodrome des végétations de France : les Isoëtetea velatae de Foucault 1988 et les Juncetea bufonii de Foucault 1988 (« Isoëto–Nanojuncetea bufonii »). Journal de botanique de la Société botanique de France 62 : 35-70, 63 : 63-109.
Foucault B. (de), 2016a. Quelques données phytosociologiques prises lors de la minisession de la Société botanique de France « sur les traces de l’abbé Coste » (25‑27 avril 2014). Journal de botanique de la Société botanique de France 76 : 87-92.
Foucault B. (de), 2016b. Contribution au prodrome des végétations de France : les Arrhenatheretea elatioris Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952. Documents phytosociologiques, 3e série, 3 : 1-219.
Foucault B. (de), 2017. Données phytosociologiques collectées lors des sorties du groupe botanique de la SESA durant l’année 2016. Bulletin de la Société d’études scientifiques de l’Aude CXVI : 187-196.
Foucault B. (de), 2018. Contribution à la connaissance phytosociologique de pelouses, prairies et garrigues de la Malepère (département de l’Aude, France). Evaxiana 5 : 156-186.
Foucault B. (de), 2020. Données phytosociologiques sur les forêts de la Malepère (Aude, France) et leurs lisières. Carnets botaniques 9 : 1-15 et 15 tableaux. DOI : https://doi.org/10.34971/5nt2-zy92.
Foucault B. (de), 2021. Miscellanées phytosociologiques : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 44 : 1-23.
Foucault B. (de) & Catteau E., 2012. Contribution au prodrome des végétations de France : les Agrostietea stoloniferae Oberd. 1983. Journal de botanique de la Société botanique de France 59 : 5-131.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Ivanez D., 2012. Document d’objectifs Site Natura 2000 « Vallée du Lampy » FR9101446 ; 1 – Inventaires, enjeux et objectifs. Document Écologistes de l’Euzière, Communauté de communes du Cabardès au Canal du Midi, Communauté de communes du Cabardès-Montagne noire, Communauté de communes Lauragais-Montagne noire, DDTM 11, DREAL-LR, 219 p. & annexes.
Kessler F., Laigneau F., Infante M., Hannoire C., Sauter N. & Corriol G., 2019 (2015). Flore, fonge et végétation des Rougiers du Camarès (Aveyron, France). Le Monde des plantes 518 : 3-32.
Klesczewski M., 2000. Graslandgesellschaften im Bergmassiv des Mont Aigoual (Cevennen, Südfrankreich). Tuexenia 20 : 189-212.
Klesczewski M., 2011-2015. ENS de Peyremale (Montolieu, Aude) ; suivis de la richesse végétale des habitats prairiaux d’intérêt patrimonial, années 2011-2015. Rapports Conservatoire d’espaces naturels du Languedoc-Roussillon, Conseil départemental de l’Aude.
Klesczewski M., 2012. Quand l’agriculture garantit une richesse biologique exceptionnelle. Zones Humides Infos 75-76 : 17.
Klesczewski M. & Bosca F., 2008. Agriculture et Biodiversité – « Quand l’agriculture garantit une richesse biologique exceptionnelle » – Le domaine de Peyremale dans l’Aude. Plaquette de présentation Conservatoire d’espaces naturels du Languedoc-Roussillon, Fédération Aude Claire, SESA, LPO Aude, OPIE L-R, 6 p. & 6 fiches-habitat.
Laribi M., Acherar M., de Foucault B., Mathez J. & Derridj A., 2017. La nardaie marginale dans l’Akfadou (Grande Kabylie, Algérie) : essai de caractérisation et proposition de gestion conservatoire d’un habitat relictuel menacé. Documents phytosociologiques, 3e série, 6 : 172-181.
Mady M., de Foucault B. & Vergne Th., 2018. Analyse prospective d’une disparition inéluctable : les pelouses et prairies maigres à Anacamptis morio subsp. morio de la région de Rochechouart (Haute-Vienne). Bulletin de la Société botanique du Centre-Ouest, NS, 49 : 497-511.
Patel M. & Reddy M.N., 2019. Revealing a new species of Ophioglossum (Ophioglossaceae-Pteridophyta) from India with palynological and phylogenic implications. Botany Letters 166 (4) : 425-433.
Quézel P., 1953. Contribution à l’étude phytosociologique et géobotanique de la Sierra Nevada. Memórias da Sociedade Broteriana IX : 1-77.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Remerciements
Un grand merci à Annie et Jean-Pierre Pautou, pour leur accueil toujours chaleureux sur le domaine, à Elodie Klesczewski, pour le soin apporté à la relecture finale.


