
Contribution à la description de forêts nord-américaines selon le paradigme synusial
Title
Contribution to the description of some North American forests according to the synusial paradigm
Résumé
Sur une base bibliographique, l’auteur dresse un système hiérarchique pour la végétation arborescente (deux ordres, avec six et deux alliances), la végétation arbustive (deux classes avec respectivement deux ordres, l’un à quatre alliances et l’autre à une alliance, et deux ordres, l’un à deux alliances, l’autre à une alliance) et la végétation basse vivace (une classe avec trois ordres, respectivement à cinq, quatre et quatre alliances) de forêts nord-américaines (Canada et États-Unis).
Abstract
Based on bibliographic sources, the author establishes a hierarchical system for tree vegetation (two orders, with six and two alliances), shrub vegetation (two classes with two orders each, one with four alliances and the other with one alliance, and two orders, one with two alliances and the other with one alliance) and low perennial vegetation (one class with three orders, with five, four and four alliances respectively) in North American forests (Canada and the United States).
1. Introduction
Pendant plusieurs années, je me suis intéressé à l’application du paradigme synusial (de Foucault, 1986 ; Gillet et al., 1991) aux forêts eurasiatiques, africaines et sud-américaines (de Foucault, 2020, 2022, 2023a, b, 2024a, b, 2025a, b). Faute d’avoir réalisé un voyage de découverte en Amérique du Nord, les forêts de cette partie du monde avaient alors été délaissées. L’objet de cette contribution est justement d’apporter une nouvelle vision de cette végétation sur la base de la bibliographie, susceptible de stimuler de nouvelles recherches sur ce thème.
Quelques conventions préalables sont nécessaires : la nomenclature taxonomique suit Plants of the World Online (POWO), même si ce référentiel est quelque peu en retard pour certains taxons à la date actuelle (par exemple Myrica californica devrait être nommé Morella californica, Blechnum spicant devrait être Struthiopteris spicant) ; pour économiser de la place dans les tableaux et les listes isolées, les rangs ‘subsp.’ ou ‘var.’ sont indiqués par *. À titre d’information, dans le texte, les familles sont indiquées pour les genres absents de la flore française. Le symbole ’j’ appliqué à un taxon potentiellement arbustif ou arborescent ligneux indique une forme juvénile. Parfois un nom ancien recouvre actuellement deux noms, le défaut d’indication des autorités dans les sources utilisées ne permet pas toujours de préciser quel taxon est concerné ; dans ce cas le nom sera alors indiqué selon l’exemple suivant : dans Rivas-Martínez et al. (1999), on rencontre Artemisia arctica qui, selon POWO, doit être actualisé soit en A. richardsoniana, soit en A. norvegica subsp. saxatilis ; devant l’impossibilité de choisir, on notera ce taxon comme Artemisia richardsoniana/norvegica *saxatilis. Pour éviter de longues listes de taxons, on a aussi utilisé la notion de « groupe sociologique » (GS) pour représenter un ensemble de taxons à affinités sociologiques déduit des tableaux de synthèse présentés.
Contrairement à des choix précédents, j’ai retenu les noms de taxons initiaux lorsqu’ils respectent l’homogénéité biologique inhérente au paradigme synusial ; par exemple Géhu et Géhu-Franck (1984) ont décrit le Pino pinastri-Quercetum ilicis selon le paradigme classique, nom repris sous la même forme dans ma publication de 2020 (p. 36) en y mettant mon autorité faute de pouvoir le dénommer autrement car seules les deux essences éponymes sont présentes dans la végétation arborescente ; cette position n’est guère tenable, d’où ce choix de reprendre le nom initial.
Parmi les travaux phytosociologiques antérieurs, je n’ai pu tenir compte par exemple du travail de Ansseau & Grandtner (1988) car beaucoup de syntaxons empruntés à des mémoires inédits sont invalides. De même le catalogue de Dansereau (1959) est inutilisable dans une telle synthèse. Faute d’avoir voyagé en Amérique du Nord, pour illustrer cette synthèse j’ai utilisé des illustrations empruntées à la Flore laurentienne de Marie-Victorin (1935).
Dans la suite, on abordera donc successivement la végétation arborescente, les végétations arbustives et herbacées. La végétation bryophytique a été laissée de côté, car, compte tenu de l’espace vital des taxons concernés, son homogénéité physionomique est loin d’être assurée ; il y aura sans aucun doute nécessité de séparer dans l’avenir des associations plutôt pionnières à petites hépatiques et mousses acrocarpes, et des bryoclimax à grandes pleurocarpes, éventuellement dendroïdes, reproduisant alors à une petite échelle la canopée arborescente qui les surmonte.
2. La végétation arborescente
Les forêts nord-américaines sont caractérisées par de nombreux taxons arborescents relevant surtout des Pinaceae (Abies, Larix, Picea, Pinus, Pseudotsuga, Tsuga), Betulaceae (Alnus, Betula), Sapindaceae (Acer), Cupressaceae (Thuja, Sequoia, parfois Juniperus), Rosaceae (Prunus, Sorbus), Salicaceae (Populus, Salix)…
2.1. De Colombie-Britannique
Pour celles de Colombie-Britannique, des données sont disponibles dans Wali (1969, selon une méthodologie et une nomenclature aujourd’hui inadéquates, mais ses données sont utiles et donc reprises selon le paradigme suivi ici), puis Krestov et al. (2000), repris par Klinka et al. (2002) et synthétisées dans le tableau 1. En dépit du titre de leur article, dans le tableau 2 de Krestov et al. (2000), Picea mariana (figure 1) est complétement absent, alors qu’il est noté avec une présence de V (une fois IV) dans leur tableau différentiel 3 ; en conséquence on a repris les présences de ce tableau 3 pour ce taxon.
Les colonnes 1 à 9 décrivent un unique Piceo laxae-Pinetum contortae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (A2 + A3) du relevé 1 du tableau 2 in Wali (1969, Vegetation-environment relationships… : 18, en remplaçant Picea glauca par P. laxa), avec une variation à Picea mariana dans les colonnes 6 à 9. Les colonnes 10 et 11 (et 12 ?) correspondent plutôt à un Betulo papyriferae-Piceetum laxae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente (A1 + A2 + A3) du relevé 6 du tableau 4 in Wali (1969, Vegetation-environment relationships… : 27, en remplaçant Picea glauca par P. laxa). La colonne 15 décrit un boisement à Pinus contorta-Larix laricina-Picea mariana qui n’est sans doute qu’une variation territoriale du Larici laricinae-Piceetum marianae défini plus bas (cf. 2.3).
2.2. Du Québec
Gagnon et Grandtner (1973) apportent des données sur les « tremblaies » à Populus tremuloides du Québec méridional synthétisées dans le tableau 2. Les colonnes 1 à 6 peuvent être rapportées à l’Abieti balsameae-Populetum tremuloidis Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 180) (incl. le Pino strobi-Populetum tremuloidis Gagnon & Grandtner 1973 et le Pino divaricatae-Populetum tremuloidis Gagnon & Grandtner 1973), lectotypifié en 2.3, avec une variation à Pinus banksiana (= P. divaricata ; figure 2) en colonne 4. Les colonnes 8 et 9 correspondent au Betulo alleghanensis-Populetum tremuloidis ass. nov. hoc loco [syn. : Abieti balsameae-Populetum tremuloidis Blouin 1970 apud Gagnon & Grandtner 1973 p. p., excluant son lectotype], typus nominis hoc loco : composante arborescente (A) du relevé 250 du tableau 43 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et des Forêts du Québec 14 hors texte), avec une variation à Thuja occidentalis en colonne 9. Les colonnes 10 à 13 décrivent l’Aceri sacchari–Populetum tremuloidis Blouin 1970 apud Gagnon & Grandtner 1973 (Mémoires du ministère des Terres et des Forêts du Québec 14 : 43), neotypus nominis hoc loco : composante arborescente (A) du relevé 441 du tableau 40 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et des Forêts du Québec 14 hors texte), avec une variation à Tilia americana en colonne 12. Enfin les colonnes 14 et 15 correspondent à l’Alno rugosae–Populetum tremuloidis Grandtner 1960 apud Gagnon & Grandtner 1973 (Mémoires du ministère des Terres et des Forêts du Québec 14 : 90), neotypus nominis hoc loco : composante arborescente (A) du relevé 254 du tableau 44 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et des Forêts du Québec 14 hors texte, en remplaçant Alnus rugosa var. americana par A. incana subsp. rugosa), avec une variation à Fraxinus nigra en colonne 15. Faute d’accès aux mémoires correspondants, on n’a pas pu vérifier la validité des syntaxons attribués par les auteurs à Blouin (1970, sans doute sa thèse non publiée, voir bibliographie) et Grandtner (1960).
2.3. D’autres régions, dont États-Unis
Les soixante-dix-neuf colonnes du tableau 3 synthétisent de nombreuses données émanant de Blouin & Grandtner (1971) et surtout de Rivas-Martínez et al. (1999), se répartissant en dix ensembles codés de I à X.
L’ensemble I (colonnes 1 à 10) est surtout marqué par Pinus banksiana dans les colonnes 1 à 5 : Populo tremuloidis-Pinetum banksianae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 3 du tableau 3 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 52) ; on retrouve cette association dans la fiche préparée par Baldwin & Chapman (2017). La colonne 6 est le Larici laricinae-Piceetum marianae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 7 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 63), reconnaissable aussi chez Gauthier et al. (1986, tableau 1 : colonne 4). Les colonnes 7 à 10 sont définies un peu négativement comme Piceo marianae-Abietetum balsameae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 1 du tableau 8 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 65). Cet ensemble est aussi présent à Terre-Neuve (Damman, 1964, tableaux 1 pour le Larici laricinae-Piceetum marianae et 2 pour le Piceo marianae-Abietetum balsameae).
L’ensemble II (colonnes 11 à 26) est différencié par le GS à Acer spicatum. La colonne 11 peut être affectée à l’Abieti balsameae-Piceetum rubentis Marcotte & Grandtner 1974, la colonne 12 au Sorbo americanae-Betuletum cordifoliae Sardinero in Rivas Mart. et al. (1999, Itinera Geobot. 12 : 63), les colonnes 13 à 16 au Betulo alleghaniensis-Abietetum balsameae Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 175), lectotypus nominis hoc loco : composante arborescente du relevé 6533 du tableau 143 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), les colonnes 17 et 18 à l’Abieti balsameae-Populetum tremuloidis Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 180, sous une race à Sorbus decora), lectotypus nominis hoc loco : composante arborescente du relevé 5999 du tableau 144 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), les colonnes 19 à 22 au Betulo alleghaniensis-Aceretum sacchari Jurdant & Roberge 1965 apud Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 180), incluant le Fago-Aceretum sacchari Lemieux 1963 apud Blouin & Grandtner 1971, le Piceo glaucae-Abietetum Blouin & Grandtner 1971 et l’Aceri-Populetum tremuloidis Blouin & Grandtner 1971, neotypus nominis hoc loco : composante arborescente du relevé 6596 du tableau 148 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), la colonne 23 à l’Abietetum balsameae Damman 1964, la colonne 24 au Betulo cordifoliae-Populetum tremuloidis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 60 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 191). Cet ensemble II possède la valeur d’une alliance de végétation arborescente boréo-américaine caractérisée par Abies balsamea (figure 3), Sorbus decora, Picea rubens, Acer spicatum, A. saccharum, A. rubrum, A. pensylvanicum, Betula alleghaniensis, B. cordifolia, Thuja occidentalis (figure 4), Fagus grandifolia (figure 5), Tsuga canadensis (figure 6), l’Aceri spicati-Abietion balsameae all. nov. hoc loco, typus nominis hoc loco : le Betulo alleghaniensis-Abietetum balsameae Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 175). Plusieurs de ces unités sont illustrées par Elliot-Fisk (1988 : 38, 39, 44, 45, 48) et se retrouve aussi au Québec (Ansseau & Grandtner, 1988 ; Desponts et al., 2004, tableau A1) ainsi qu’à Saint-Pierre-et-Miquelon (Bosseaux, 1965 ; Muller, 2006, 2025).
L’ensemble III (colonnes 27 à 32) est assez peu caractérisé du point de vue arborescent, en dehors éventuellement de Betula pendula subsp. mandshurica, souvent nommé B. neoalaskana dans la publication espagnole (caractéristique d’alliance). À côté du Piceo glaucae-Betuletum neoalaskanae Peinado, Aguirre & Cruz 1998 (colonne 29), muté ici en Piceo laxae-Betuletum mandshuricae nom. mut. hoc loco, on peut mettre en exergue les colonnes 30 à 32 comme Populo tremuloidis-Piceetum laxae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 3 du tableau 54 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 179, en remplaçant Picea glauca par P. laxa). On peut alors définir cet ensemble III comme Betulo mandshuricae-Piceion laxae all. nov. hoc loco, typus nominis hoc loco : le Populo tremuloidis-Piceetum laxae nov.
L’ensemble IV (colonnes 33 à 46) est différencié ou caractérisé par le GS à Pseudotsuga menziesii var. glauca. Les colonnes 34 à 36 sont autant de variations d’un unique Abieti lasiocarpae-Piceetum engelmannii ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 42 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 149, en remplaçant Abies bifolia par A. lasiocarpa). La colonne 37 est le Piceo engelmannii-Pinetum flexilis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 41 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 147). Les colonnes 38 et 39 correspondent au Piceetum engelmannii-albertianae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 152, … engelmannio-…), la colonne 40 au Piceetum pungentis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 155). La colonne 41 est le Pinetum engelmannii-aristatae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 1 du tableau 46 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 158). La colonne 42 est le Larici occidentalis-Pseudotsugetum glaucae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 164). Les colonnes 43 et 44 peuvent être considérées comme deux variations du seul Junipero scopulorum-Pinetum scopulorum ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 51 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 169). La colonne 45 est le Pseudotsugo glaucae-Pinetum flexilis Rivas Mart. et al. (1999, Itinera Geobot. 12 : 170). Tout cet ensemble IV a la valeur d’une alliance caractérisée ou différenciée par Pseudotsuga menziesii subsp. glauca, Pinus flexilis, P. scopulorum, P. engelmannii, Abies lasiocarpa var. l., Picea engelmannii, Juniperus scopulorum, Prunus virginiana var. demissa, le Pino engelmannii-Pseudotsugion glaucae all. nov. hoc loco, typus nominis hoc loco : le Piceo engelmannii-Pinetum flexilis nov. Cette alliance est illustrée par Peet (1988 : 87, 90, 94).
Le superensemble formé par les ensembles I à IV possède alors la valeur d’un ordre caractérisé par Populus tremuloides, P. balsamifera (figure 7), Picea laxa (figure 8) et P. mariana, les Populo tremuloidis-Piceetalia laxae ord. nov. hoc loco, typus nominis hoc loco : l’Aceri spicati-Abietion balsameae nov. Avec seulement trois relevés, il semble que le « Vaccinio scoparii-Pinetum albicaulis » Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 161) puisse se placer dans le Pino engelmannii-Pseudotsugion glaucae.
L’ensemble V (colonnes 47 à 56) rassemble des boisements à Fraxinus nigra et Alnus incana subsp. rugosa. Les colonnes 48 et 49 correspondent au Thujo occidentalis-Abietetum balsameae Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 : 137), lectotypus nominis hoc loco : composante arborescente du relevé 5850 du tableau 142 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), les 50 et 51 au Fraxino nigrae-Populetum balsamiferae Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 : 318), lectotypus nominis hoc loco : composante arborescente du relevé 5824 du tableau 149 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), la 52 à l’Ulmo americanae-Fraxinetum nigrae Lemieux 1964 apud Blouin & Grandtner 1971 (Mémoires du ministère des Terres et des Forêts du Québec 6 : 314), neotypus nominis hoc loco : composante arborescente du relevé 5637 du tableau 149 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte), les 53 à 56 à l’Alnetum rugosae Dansereau 1959 (Contributions de l’Institut botanique de l’Université de Montréal 75 : 41), neotypus nominis hoc loco : composante arborescente du relevé 5825 du tableau 149 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte). Caractérisé par Fraxinus nigra, Alnus incana subsp. rugosa, Ulmus americana, Thuja occidentalis, cet ensemble V peut être interprété comme alliance, Fraxino nigrae-Alnion rugosae all. nov. hoc loco, vicariante américaine de plusieurs alliances européennes des Fraxino excelsioris-Alnetalia glutinosae B. Foucault & Julve in B. Foucault 2020 (de Foucault, 2020), typus nominis hoc loco : le Fraxino nigrae-Populetum balsamiferae Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 : 318) ; il est évident qu’on peut y rattacher aussi l’Alno rugosae–Populetum tremuloidis Grandtner 1960 apud Gagnon & Grandtner 1973 évoqué supra et dans le tableau 2 (colonnes 14 et 15). Cette alliance doit sans doute inclure aussi la saulaie arborescente québécoise à Salix fragilis, Fraxinus nigra et F. pennsylvanica citée par Lacoursière et Grandtner (1971, 1972). Du tableau XXXV de Lacoursière & Grandtner (1972), on peut extraire la liste suivante (sur cinq relevés) : Salix fragilis V, Prunus virginiana II, Fraxinus nigra II, F. pennsylvanica I, soit le Fraxino nigrae-Salicetum fragilis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau XXXV in Lacoursière & Grandtner (1972, Nat. Can. 99 (5) : 503).
L’ensemble VI (colonnes 57 à 67) est assez homogène, caractérisé par le GS à Tsuga heterophylla ; les colonnes 57 à 59 (plutôt appauvries en 59) sont à considérer comme de petites variations du Tsugo heterophyllae-Piceetum sitchensis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 15 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 86). La colonne 60 est le Tsugo heterophyllae-Sequoietum sempervirentis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 91). La colonne 61 est un Aceri macrophylli-Pseudotsugetum menziesii ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 6 du tableau 19 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 94). La colonne 62 est l’Abieti grandis–Pseudotsugetum menziesii Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 96), la colonne 63 l’Abieti amabilis-Tsugetum heterophyllae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 99), la colonne 64 le Piceo engelmannii-Tsugetum heterophyllae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 102), la colonne 65 le Tsugo mertensianae-Abietetum procerae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 110), la colonne 66 est le Piceo sitchensis-Pinetum contortae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 30 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 120), la colonne 67 étant l’Aceri macrophylli-Alnetum rubrae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 127). Caractérisé par Tsuga heterophylla, Pseudotsuga menziesii subsp. m., Thuja plicata (photo 1), Picea sitchensis, Alnus rubra, Acer circinatum, A. macrophyllum, Pinus monticola, P. contorta var. murrayana, Taxus brevifolia (essence par ailleurs connue pour ses molécules anticancéreuses), tout cet ensemble VI correspond à une alliance originale de l’Ouest américain, le Thujo plicatae-Tsugion heterophyllae all. nov. hoc loco, typus nominis hoc loco : le Tsugo heterophyllae-Piceetum sitchensis nov. Cette alliance est illustrée par Franklin (1988 : 108, 112, 117).

Passons à l’ensemble VII (colonnes 68 à 73) caractérisé par le GS à Tsuga mertensiana, avec
- en 68, le Tsugo mertensianae-Piceetum sitchensis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 113) ;
- en 69, l’Abieti amabilis-Tsugetum mertensianae Brooke, Peterson & Krajina 1970 ;
- en 70, l’Abieti lasiocarpae-Tsugetum mertensianae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 108) ;
- en 71, l’Abieti lasiocarpae-Pinetum albicaulis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 115) ;
- en 72, l’Abieti grandis-Laricetum lyalli nov. hoc loco, typus nominis hoc loco: composante arborescente du relevé 3 du tableau 28 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 117).
Cet ensemble correspond au Tsugion mertensianae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 104), caractérisé par Tsuga mertensiana, Callitropsis nootkatensis, Sorbus sitchensis, Abies grandis. Il est illustré par Franklin (1988 : 120, 122) et Barbour (1988 : 154).
L’ensemble VIII (colonnes 74 à 76) rapproche les syntaxons suivants :
- en 74, l’Aceri douglasii-Pinetum contortae nov. hoc loco, typus nominis hoc loco: composante arborescente du relevé 2 du tableau 31 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 122) ;
- en 75 et 76, l’Aceri douglasii-Populetum tristis nov. hoc loco, typus nominis hoc loco: composante arborescente du relevé 2 du tableau 35 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 132, en changeant Populus trichocarpa en P. tristis).
Caractérisé par Acer glabrum var. douglasii, Pinus contorta subsp. c. et Populus tristis, cet ensemble a valeur d’alliance : le Populo tristis-Acerion douglasii all. nov. hoc loco, typus nominis hoc loco : l’Aceri douglasii-Populetum tristis nov.
L’ensemble IX rapproche deux ripisylves à Populus angustifolia, en colonne 77 le Populetum angustifolio-moniliferae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 197) et en colonne 78 le Betulo occidentalis-Populetum angustifoliae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 197) ; elles relèvent du Populion angustifolio-moniliferae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 196) et des Populetalia angustifolio-deltoidis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 194), dont l’aire est tracée sur la carte 1, avec celle du Platano occidentalis-Populion deltoidis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 201) prov. Peet (1988 : 77) en donne une illustration du Wyoming.
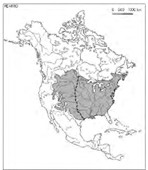
L’ensemble X est réduit à la colonne 79, le Platano racemosae-Populetum fremontii Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 203), seule association connue du Populion fremontii Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 202) et des Populetalia fremontii Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 201) sud-ouest-nord-américains (carte 2).
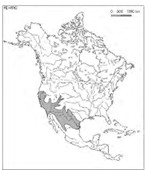
2.4. Synthèse des associations arborescentes
Le tableau 4 dresse une synthèse plus condensée de ces unités (hors alliances IX et X déjà bien placées) afin d’en avoir une vision plus globale. Il montre notamment que les deux associations de Colombie-Britannique (colonne PAl) se placent bien dans les Populo tremuloidis-Piceetalia laxae, mais dans une alliance distincte des cinq premières, le Piceo laxae-Abietion lasiocarpae all. nov. hoc loco, caractérisée par Abies lasiocarpa subsp. l., Betula papyrifera, Pinus contorta var. c. et l’absence de nombreux taxons caractérisant les autres alliances, typus nominis hoc loco : le Piceo laxae-Pinetum contortae nov. Warder (1970, tableau 2) dresse une liste synthétique quantifiée d’un groupement canadien à Abies balsamea (présence V), Betula papyrifera (IV), Larix laricina (III ; figure 9), Acer rubrum (III), Thuja occidentalis (III), Picea mariana (III), P. laxa (III), Populus tremuloides (III), Picea rubens (II), Betula alleghaniensis (II), avec de nouveaux taxons : Pinus strobus (III ; figure 10), Prunus pennsylvanica (II)… qui se rattache encore à cet ordre. Bouchard (1970) rapporte aussi des données du sud du Québec. Béguin et al. (1995, tableau 1) décrivent un « Diervillo lonicerae-Quercetum rubrae », dont la végétation arborescente entre bien dans les Populo tremuloidis-Piceetalia laxae.
L’unité I ne rentre dans aucune des alliances déjà définies ; il convient de la définir sous le nom de Piceion banksiano-marianae all. nov. hoc loco, caractérisée par Pinus banksiana, P. resinosa, Larix laricina, ainsi que par l’absence de taxons caractérisant les alliances voisines, typus nominis hoc loco : le Larici laricinae-Piceetum marianae nov.
Les alliances VI à VIII, restées non rattachées à un ordre selon le paradigme suivi ici, sont reliées statistiquement par Picea sitchensis et P. engelmannii, auxquels on peut ajouter Sorbus sitchensis et Abies amabilis, de sorte que l’on peut définir les Piceetalia sitchensi-engelmannii ord. nov. hoc loco, typus nominis hoc loco : le Thujo plicatae-Tsugion heterophyllae nov.
Enfin les colonnes Qu1 à Qu15 du tableau 4 reprennent les données du tableau 2 du Québec ; cette synthèse montre l’appartenance des Qu1 à Qu13 aux Populo tremuloidis-Piceetalia laxae et à l’Aceri spicati-Abietion balsameae, ajoutant d’ailleurs Prunus pennsylvanica et P. virginiana aux taxons caractéristiques de l’alliance, alors que les colonnes Qu14 et Qu15 se placent bien dans le Fraxino nigrae-Alnion rugosae.
Ce même tableau 4 montre enfin que les deux ordres nouvellement décrits ici ne partagent aucun taxon et qu’ils ne peuvent donc en principe être rattachés à la même classe phytosociologique. Si on suit Julve (in e-veg.net [29.07.2025]) on peut rattacher les Populo tremuloidis-Piceetalia laxae aux Abieti balsameae-Piceetea glaucae Knapp ex Grandtner 1966, muté en Abieti balsameae-Piceetea laxae nom. mut. hoc loco, les Piceetalia sitchensi-engelmannii aux Tsugo heterophyllae-Thujetea plicatae Knapp 1957. Il faudrait cependant vérifier la validité de ces noms de classe.
En 1938, Cain et Penfound décrivent l’« Aceretum rubri » de Long Island (près de New York) sur la base de cinq relevés. L’association arborescente (tableau II des auteurs) correspond à la liste synthétique suivante :
Acer rubrum V, Nyssa sylvatica (Nyssaceae) III, Amelanchier canadensis II, Betula populifolia II, Quercus alba II, Sassafras albidum (Lauraceae) II, Fraxinus americana I, Pinus rigida I,
qui n’a pas d’équivalent dans ce qui est décrit ici, Nysso sylvaticae-Aceretum rubri ass. nov. hoc loco, typus nominis hoc loco : relevé de la colonne Masapequa du tableau II in Cain & Penfound (1938, Am. Midl. Nat. 19 : 394). Cette association n’entre bien dans aucune des unités du tableau 4.
On ne possède que fort peu de données sur les forêts de Louisiane. Penfound et Howard (1940) apportent cependant quelques relevés d’une forêt à Quercus nigra (V sur quinze relevés de leur tableau 1), Q. virginiana (IV), Celtis laevigata (III), Ulmus americana (III), Fraxinus americana (II), soit le Quercetum virginiano-nigrae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du quadrat 9 du tableau in Penfound & Howard (1940, Am. Midl. Nat. 23 : 168). Dans leur tableau 3, Hall et Penfound (1939) dressent un inventaire synthétique de forêts hygrophiles à cyprès chauve :
- en strate dominante : Taxodium distichum V, Nyssa biflora V, aquatica V, Acer rubrum *drummondii V, Fraxinus profunda V, Nekemias arborea (Vitaceae) II, Fraxinus caroliniana I,
- en strate arbustive : Gelsemium sempervirens (Gelsemiaceae) V, Toxicodendron radicans V, Myrica cerifera IV, Itea virginica (Iteaceae) IV, Smilax walteri IV, Cephalanthus occidentalis (Rubiaceae) III, Wisteria frutescens *macrostachya (Fabaceae), Styrax americanus II, Smilax laurifolia I, plus des taxons de présence +,
deux associations potentielles à valider avec des relevés précis.
3. La végétation arbustive intraforestière
Cette végétation rassemble des taxons relevant des Ericaceae (Gaultheria, Rhododendron, Vaccinium), Rosaceae (Amelanchier, Holodiscus, Rosa, Rubus, Spiraea), Caprifoliaceae (Lonicera, Symphoricarpos), Betulaceae arbustives (Alnus, Betula, Corylus), Viburnaceae (Sambucus, Viburnum), Anacardiaceae (Rhus, Toxicodendron), Cornaceae (Cornus), Grossulariaceae (Ribes), Salicaceae arbustives (Salix)…
3.1. De Colombie-Britannique
En ce qui concerne la végétation arbustive intraforestière, le tableau 5 porte d’abord sur la végétation arbustive de Colombie-Britannique. Les colonnes 3 à 6 décrivent le Ribeso lacustris-Alnetum sinuatae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 4 in Wali (1969, Vegetation-environment relationships… : 27). La colonne 7 est le Ribeso hudsoniani-Viburnetum edulis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 4 du tableau 5 in Wali (1969, Vegetation-environment relationships… : 33). Les colonnes 11 à 13 peuvent être interprétées comme Shepherdio canadensis-Rosetum acicularis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 89 de l’appendice 7 in Krestov et al. (2000, Scientia Silvica 26 : 72-73). Tout cet ensemble peut être considéré comme une alliance nord-ouest-américaine bien caractérisée par Lonicera involucrata subsp. i., Rubus pedatus, Viburnum edule, Rosa acicularis, R. gymnocarpa, Ribes lacustre, R. triste, R. hudsonianum, Alnus alnobetula subsp. sinuata, Amelanchier alnifolia, Salix scouleriana, Shepherdia canadensis (Elaeagnaceae), Spiraea betulifolia, le Viburno edulis-Lonicerion involucratae all. nov. hoc loco, typus nominis hoc loco : le Ribeso hudsoniani-Viburnetum edulis nov.
3.2. Du Québec
Béguin et al. (1995) décrivent plusieurs syntaxons arbustifs du Québec, rapportés dans les colonnes 1 à 4 du tableau 6 ; ici on a conservé les formes juvéniles des arbres car ces noms sont utilisés pour nommer des associations. On aurait aimé tester la position synsystématique des groupements périforestiers arbustifs du Québec cités par Ansseau & Grandtner (1986), mais, comme pour les groupements arborescents, ceux-ci sont souvent déduits de thèses inédites et difficilement accessibles ; en outre il n’y a pas de tableaux attachés aux syntaxons cités. Les colonnes 1 et 2 de ce tableau 6 se rapportent au Querco rubrae-Cornetum rugosae Béguin et al. 1995, dont la colonne 3 paraît n’être qu’une variation. La colonne 4 est la composante arbustive d’une association que les auteurs nomment le Betulo papyriferae-Populetum tremuloidis, que l’on peut dénommer plutôt Populo tremuloidis-Rhoetum typhinae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 6 in Béguin et al. (1995, Doc. Phytosoc., n. s., XV : 191). Ces deux associations relèvent d’une alliance caractérisée par Diervilla lonicera (Caprifoliaceae), Rhus typhina (photo 2), Toxicodendron radicans, Cornus rugosa, Amelanchier laevis, Corylus cornuta, le Rhoo typhinae-Diervillion lonicerae all. nov. hoc loco, typus nominis hoc loco : le Querco rubrae-Cornetum rugosae Béguin et al. 1995 (Doc. Phytosoc., n. s., XV : 188).

Les colonnes 5 à 8 décrivent une végétation différente, mais finalement assez peu caractérisée, au contraire de l’ensemble formé par les colonnes 9 à 19, décrivant le Ribeso glandulosae-Loniceretum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 298 du tableau 42 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et Forêts du Québec 14 hors texte). Cet ensemble 9 à 19 décrit alors une alliance originale, l’Hydrangeo arborescentis-Lonicerion canadensis all. nov. hoc loco, caractérisée par Hydrangea arborescens (Hydrangeaceae), Amelanchier bartramiana, Ribes cynosbati, R. americanum (photo 11), Lonicera canadensis, Viburnum cassinoides, Cornus sericea et les taxons du typus nominis hoc loco : le Ribeso glandulosae-Loniceretum canadensis nov.
L’ensemble formé par les colonnes 20 à 26 se démarque par le GS à Salix lucida. Les colonnes 20 à 22 (et sans doute 23) sont des variations d’un unique Salici lucidae-Cornetum sericeae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 6588 du tableau 142 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Cornus stolonifera par C. sericea). Les colonnes 24 à 26 décrivent le Salici pyrifoliae-Cornetum sericeae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 6546 du tableau 141 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Cornus stolonifera par C. sericea). Cet ensemble de sept colonnes a la valeur d’une alliance caractérisée par Salix lucida, S. discolor (photo 12), S. pyrifolia, S. eriocephala, Ribes hirtellum, Spiraea alba var. latifolia, Cornus rugosa, Viburnum trilobum, le Salicion discolori-lucidae all. nov. hoc loco, typus nominis hoc loco : le Salici lucidae-Cornetum sericeae nov.
Les colonnes 27 et 28 décrivent le Viburno edulis-Ribesetum lacustris ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5839 du tableau 142 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte). Les colonnes 29 à 33 paraissent se rattacher à un unique Ribeso lacustris-Loniceretum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5930 du tableau 143 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte). Les colonnes 34 à 38 sont marquées par Kalmia angustifolia (Ericaceae), mais restent difficiles à interpréter.
Les colonnes 39 à 47 partagent Rubus idaeus ; l’on peut séparer
- la colonne 39 comme Alno crispae-Viburnetum cassinoidis nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 6532 du tableau 145 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte) ;
- la colonne 41 comme Corylo cornutae-Viburnetum trilobi nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 5775 du tableau 149 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte) ;
- les colonnes 44 à 47 comme Sambuco pubentis-Coryletum cornutae nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 5871 du tableau 148 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte).
Le large ensemble formé par les colonnes 27 à 47 possède clairement la valeur d’une alliance, quoique définie un peu négativement relativement aux précédentes, faiblement différenciée par Alnus alnobetula subsp. crispa, Kalmia angustifolia, Lonicero canadensis-Corylion cornutae all. nov. hoc loco, typus nominis hoc loco : le Viburno edulis-Ribesetum lacustris nov.
Avant de passer aux syntaxons de Rivas-Martínez et al. (1999), il reste le petit ensemble formé par les colonnes 48 à 52, marqué par le GS à Kalmia angustifolia. On peut surtout mettre en exergue les colonnes 49 à 51, le Rhododendro canadensis-Kalmietum angustifoliae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5599 du tableau 140 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Ledum groenlandicum par Rhododendron canadensis). On peut alors élever cet ensemble au rang d’alliance originale, caractérisée par Kalmia angustifolia, K. polifolia, Rhododendron canadense, Aronia melanocarpa (Rosaceae), Myrica gale, l’Ilici mucronatae-Kalmion angustifoliae all. nov. hoc loco, typus nominis hoc loco : le Rhododendro canadensis-Kalmietum angustifoliae nov. On retrouve cette unité dans le tableau 1 de Damman (1964) de Terre-Neuve, mais floristiquement assez pauvre, ainsi que dans la fiche préparée par Baldwin & Chapman (2017) ; elle est aussi reconnaissable dans les listes de Muller & Etcheberry (in Muler, 2006).
Le tableau synthétique 7 facilite la hiérarchisation et la perception d’ensemble de ces six unités québécoises codées de I à VI dès le tableau 6. Les alliances II à V partagent des liens floristiques significatifs qui permettent de les rapprocher dans un ordre caractérisé par Lonicera canadensis, Salix bebbiana, Sambucus racemosa subsp. pubens, Cornus alternifolia, C. sericea, Ribes triste, les Corno alternifoliae-Loniceretalia canadensis ord. nov. hoc loco, typus nominis hoc loco : le Lonicero canadensis-Corylion cornutae nov. La colonne VI (Kalmion polifolio-angustifoliae) est reliée aux précédentes par Viburnum cassinoides et Ilex mucronata, mais ne se rattache pas à l’ordre qui vient d’être défini. Enfin, pour intégrer la colonne I, partageant avec les précédentes Corylus cornuta et Diervilla lonicera, il convient de définir la classe des Corylo cornutae-Diervilletea lonicerae classis nov. hoc loco, typus nominis hoc loco : les Corno alternifoliae-Loniceretalia canadensis nov., la colonne VI, dans laquelle manquent les deux taxons éponymes de classe, s’y rattachant par enchaînement.
3.2. Autres, notamment des États-Unis
Le tableau 8 synthétise la végétation arbustive intraforestière déduit de 337 relevés publiés par Rivas-Martínez et al. (1999). Six ensembles s’y séparent bien, codés de I à VI.
L’ensemble I (colonnes 1 à 7) est marqué par le GS à Gaultheria shallon. La colonne 1 est un Sambuco pubentis-Franguletum purshianae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 15 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 86, en remplaçant Sambucus arborescens par S. racemosa subsp. pubens), la colonne 2 le Rhododendro macrophylli-Gaultherietum shallon ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 18 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 92), la colonne 3 le Gaultherio shallon-Berberidetum nervosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 19 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 94, en remplaçant Mahonia nervosa par Berberis n.), la colonne 4 le Roso gymnocarpae-Berberidetum nervosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 6 du tableau 20 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 97, en remplaçant Mahonia nervosa par Berberis n.), la colonne 5 le Rhododendro menziesii-Vaccinietum ovalifolii ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 7 du tableau 21 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 100). Avec notamment Myrica californica, la colonne 7 est bien caractérisée : Lonicero ledebourii-Myricetum californicae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 30 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 120). Tout cet ensemble I est bien caractérisé par Gaultheria shallon (Ericaceae), Frangula purshiana, Berberis nervosa, Corylus cornuta subsp. californica, Rhododendron macrophyllum, Holodiscus discolor (Rosaceae), Rubus macropetalus, R. leucodermis pour justifier la définition d’une alliance, le Frangulo purshianae-Gaultherion shallon all. nov. hoc loco, typus nominis hoc loco : le Lonicero ledebourii-Myricetum californicae nov.
L’ensemble II (colonnes 8 à 11) est plutôt marqué par Rhododendron albiflorum. En particulier, les colonnes 9 et 10 peuvent être interprétés comme Rhododendro albiflori-Vaccinietum ovalifolii ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 7 du tableau 23 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 105). On peut en rapprocher la colonne 8 définie comme Roso gymnocarpae-Amelanchieretum sinuatae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 22 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 102). Il paraît prématuré de reconnaître une alliance dans cet ensemble II.
L’ensemble III (colonnes 12 à 19) est caractérisé par le GS à Alnus alnobetula subsp. sinuata, ses éléments semblant représenter des syntaxons vicariants de ceux à Alnus alnobetula subsp. a. eurosibériens. Les colonnes 12 et 13 peuvent constituer deux variations de l’Oplopanaco horridi-Vaccinietum ovalifolii ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 1 du tableau 17 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 90). La colonne 16 est le Vaccinio membranacei-Alnetum sinuatae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 135) et la colonne 18 le Salici acutifoliae-Alnetum sinuatae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 55 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 181). Cet ensemble III a alors valeur d’une alliance caractérisée surtout par Alnus alnobetula subsp. sinuata, Ribes laxiflorum/hudsonianum, Spiraea stevenii, Salix glauca var. acutifolia, le Rhododendro menziesii-Alnion sinuatae all. nov. hoc loco, typus nominis hoc loco : l’Oplopanaco horridi-Vaccinietum ovalifolii nov.
Le superensemble formé par les syntaxons I à III possède alors la valeur d’un ordre caractérisé par Rhododendron menziesii, Vaccinium ovalifolium, Rubus spectabilis, R. pedatus, Oplopanax horridus (Araliaceae), Sambucus racemosa subsp. pubens, les Oplopanaco horridi-Rhododendretalia menziesii ord. nov. hoc loco, typus nominis hoc loco : le Frangulo purshianae-Gaultherion shallon nov.
La large ensemble IV allant des colonnes 20 à 38 est bien caractérisé par le GS à Juniperus communis var. depressa. La colonne 23 est le Junipero depressae-Ribesetum montigeni ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 40 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 145). La colonne 26 représente le Spiraeo lucidae-Shepherdietum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 2 du tableau 42 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 149). La colonne 27 est le Roso sayi-Ribesetum lacustris ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 4 du tableau 44 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 153), la colonne 28 le Symphoricarpo occidentalis-Rosetum sayi ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 57 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 184), la colonne 29 le Symphoricarpo rotundifolii-Rosetum ultramontanae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 7 du tableau 45 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 156 en changeant Symphoricarpos utahensis en S. rotundifolius), la colonne 30 l’Amelanchiero alnifoliae-Symphoricarpetum albi ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 6 du tableau 49 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 165). Les colonnes 31 et 32 peuvent être rassemblées comme Shepherdio canadensis-Juniperetum depressae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 47 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 160). La colonne 37 est le Rhoo aromaticae-Symphoricarpetum oreophili ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 4 du tableau 51 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 169 en changeant Rhus trilobata en Rh. aromatica subsp. a.). Cet ensemble IV possède sans conteste la valeur d’une belle alliance caractérisée par Juniperus communis var. depressa, Shepherdia canadensis (Elaeagnaceae), Amelanchier alnifolia, Ribes montigenum, Rosa woodsii subsp. ultramontana, Lonicera involucrata subsp. i., L. utahensis, Physocarpus malvaceus (Rosaceae), Spiraea lucida, le Shepherdio canadensis–Juniperion depressae all. nov. hoc loco, typus nominis hoc loco : le Symphoricarpo rotundifolii-Rosetum ultramontanae nov.
Dans l’ensemble V (colonnes 39 à 43), surtout différencié négativement vis-à-vis des trois alliances qui viennent d’être définies, seule la colonne 41 est significative et définie comme Corno sericeae-Rosetum woodsii ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du tableau 63 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 198).
Enfin, l’ensemble VI (colonnes 44 à 46) rassemble des boulaies-saulaies arbustives paucispécifiques, apparaissant homologues des saulaies du Salicion waldsteinianae Oberd. 1978 des montagnes européennes : la 44 est le Salici barclayi-Betuletum glandulosae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 209), la 45 le Salici acutifoliae-Betuletum glandulosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 4 du tableau 68 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 211), la 46 le Salici pulchrae-Betuletum exilis Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 212) muté ici en Salici pulchrae-Betuletum glandulosae nom. mut. hoc loco. Ces trois associations relèvent du Salici pulchrae-Betulion glandulosae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 207). Betula glandulosa apparaît aussi à l’est de la baie d’Hudson (Canada) dans le tableau I de Payette & Filion (1975) avec Salix glauca et S. planifolia : Salici planifoliae-Betuletum glandulosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive (strate arbustive basse p. p.) du relevé 220 du tableau I in Payette & Filion (1975, Nat. Can. 102 : 790), qui ne relève cependant pas du Salici pulchrae-Betulion glandulosae. Dans leur monographie de marais de l’Alberta, Vitt et al. (1975) citent aussi B. glandulosa, mais sans guère d’autres taxons arbustifs associés.
Bliss et Cantlon (1957 : 455) ont rapporté d’Alaska des données floristiques et dynamiques sur de tels fourrés arctiques : un fourré pionnier à S. alaxensis, suivi dans le temps d’un fourré un peu plus mûr à S. alaxensis (IV), S. arbusculoides (III), S. glauca *acutifolia (II), S. hastata *h. (II), Alnus alnobetula *crispa (+), enfin un fourré mûr à Alnus alnobetula *crispa (V), Salix lanata *richardsonii (IV), S. pulchra (III), S. arbusculoides (II), S. glauca *acutifolia (II). Ces deux derniers stades dynamiques (colonnes 47 et 48 du tableau 8 hoc loco) mériteraient le rang d’association, mais elles ne pourraient être validées, car les données sources sont synthétiques. Par Salix hastata, S. arbusculoides, S. pulchra, S. alaxensis, il existe des liens floristiques avec le Salici pulchrae-Betulion glandulosae, mais ces fourrés devraient s’intégrer dans une alliance distincte inédite, illustrée par Bliss (1988 : 10, où la var. acutifolia de S. glauca est dénommée var. desertorum), qui devrait rejoindre le Salici-Betulion dans un ordre original caractérisé par Salix glauca var. acutifolia, S. alaxensis var. a., S. pulchra, S. hastata, les Salicetalia acutifolio-alaxensis ord. nov. hoc loco, typus nominis hoc loco : le Salici pulchrae-Betulion glandulosae Rivas Mart. et al. 1999 (Itinera Geobot. 12 : 207).
Dans le Manitoba (Canada central), au voisinage de boisements à Larix laricina et Picea mariana, Locky et al. (2005) ont analysé une végétation où apparaissent aussi des fourrés à Saules et Bouleau, avec Lonicera villosa, mais il s’agit ici de Betula pumila et les Salix sont très différents des précédents ; le tableau 17 présente cette végétation qui ne peut toutefois donner lieu à validation. On en reconnaît aussi des éléments dans les données de Nicholson (1993) provenant de l’Alberta.
On retrouve L. villosa et B. pumila dans des tourbières de Terre-Neuve étudiées par Wells (1976), avec des relevés précis (sur 49 relevés) : Lonicera villosa IV, Betula michauxii IV, Myrica gale IV, Aronia ´prunifolia III, Rosa nitida II, Juniperus communis *c. II, Betula pumila II, Kalmia angustifolia I et des taxons de présence + ou r (Alnus alnobetula subsp. crispa, Ilex mucronata, Viburnum cassinoides), Betulo michauxii-Loniceretum villosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 89-E du tableau 3-7 in Wells (1976, A classification of peatlands in eastern Newfoundland : 109) repris ci-après :
Terre-Neuve orientale, env. N 47° 20’, E -52° 57’, altitude 450 m, 8 taxons
Lonicera villosa 1.1, Myrica gale 2.1, Dasiphora fruticosa 1.1, Betula michauxii 1.1, B. pumila +, Rosa nitida +, Juniperus communis *c. +, Ilex mucronata +.
Le tableau 9 dresse une synthèse plus condensée de ces unités permettant d’en avoir une vision plus globale. L’unité IV peut être rapprochée de V dans un ordre caractérisé par Amelanchier alnifolia et Rosa sayi, les Roso sayi-Amelanchieretalia alnifoliae ord. nov. hoc loco, typus nominis hoc loco : le Shepherdio canadensis–Juniperion depressae nov. L’ensemble supraordinal des unités I à V a alors valeur de classe, bien distincte de celle des Corylo cornutae-Diervilletea lonicerae plus septentrionale, caractérisée par Amelanchier alnifolia, Symphoricarpos albus var. laevigatus, Corylus cornuta subsp. californica, Ribes lacustre, Viburnum edule, Salix scouleriana, Rosa sayi, les Roso sayi-Amelanchieretea alnifoliae classis nov. hoc loco, typus nominis hoc loco : les Oplopanaco horridi-Rhododendretalia menziesii nov. Comme vu au § précédent, l’unité d’Alaska (colonne Al) est rapprochée de l’unité VI dans les Salicetalia acutifolio-alaxensis, mais la classe reste indéterminée.
L’association arbustive subordonnée au Nysso sylvaticae-Aceretum rubri décrit en 2.3 à partir des données de Cain & Penfound (1938) est ainsi constituée (tableau III de ces auteurs) :
Clethra alnifolia (Clethraceae) V, Rhododendron viscosum V, Viburnum dentatum V, Vaccinium corymbosum V, Lindera benzoin j (Lauraceae) IV, Alnus incana *rugosa j IV, Smilax rotundifolia IV, Ilex verticillata III, Aronia melanocarpa III, A. ´prunifolia II, Lyonia ligustrina (Ericaceae) II, Eubotrys racemosa (Ericaceae) II, Toxicodendron radicans II, T. vernix I, Sambucus canadensis I, Parthenocissus quinquefolia I, Ilex glabra I, I. laevigata I, Viburnum prunifolium I, Rosa palustris I, Vitis labrusca I, Smilax glauca I,
décrivant le Rhododendro viscosi-Clethretum alnifoliae ass. nov. hoc loco, typus nominis hoc loco : relevé de la colonne Hauppauge du tableau III in Cain & Penfound (1938, Am. Midl. Nat. 19 : 395).
4. La végétation basse intraforestière
On aborde maintenant la végétation herbacée vivace inféodée aux boisements décrits dans les paragraphes précédents. Il existe bien de rares thérophytes, comme Impatiens capensis (malgré son nom ce taxon est d’origine nord-américaine), mais on n’a pas identifié de groupements thérophytiques significatifs à isoler, lesquels pourraient s’avérer vicariants des ourlets européens à Impatiens noli-tangere. Dans les tableaux de cette végétation, apparaît très souvent Epilobium angustifolium, taxon de répartition circumboréale ; en Europe, il caractérise surtout la classe des Epilobietea angustifolii, rassemblant la végétation pionnière vivace des coupes forestières (de Foucault & Catteau, 2015) ; ce type de végétation existe sans doute en Amérique du Nord, mais ne semble pas y avoir été étudié. Avant d’aller plus loin, signalons que Bournérias (1972) a publié quelques photos de taxons cités dans cette partie 4 (Cornus canadensis, Linnaea borealis subsp. americana, Rubus chamaemorus, Lycopodium annotinum).
4.1. De Colombie-Britannique
Le tableau 10 synthétise diverses données sur les sous-bois herbacés vivaces de Colombie-Britannique qui se répartissent en quatre ensembles assez inégaux, plusieurs à valeur d’alliance.
Dans le premier (colonnes 1 et 2), on peut surtout mettre en avant le Corno canadensis-Arctostaphyletum uvae-ursi ass. nov. hoc loco (colonne 2), typus nominis hoc loco : composante herbacée du relevé 5 du tableau 2 in Wali (1969, Vegetation-environment relationships… : 18). Cette unité I possède la valeur d’une alliance notamment caractérisée par Arctostaphylos uva-ursi, Piptatheropsis pungens (Poaceae), Solidago spathulata, Antennaria neglecta, A. rosea, Pedicularis labradorica, Eurybia conspicua (Asteraceae), partageant avec l’alliance suivante Vaccinium cespitosum et Oryzopsis asperifolia (Poaceae), le Vaccinio cespitosi-Arctostaphylion uvae-ursi all. nov. hoc loco, typus nominis hoc loco : le Corno canadensis-Arctostaphyletum uvae-ursi nov.
Le deuxième ensemble (colonnes 3 à 8) est bien différencié par le GS à Aralia nudicaulis. La colonne 4 est l’Oryzopsio asperifoliae-Linnaeetum americanae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 4 du tableau 3 in Wali (1969, Vegetation-environment relationships… : 22). Les colonnes 6 et 7 sont deux variations d’un Galio triflori-Gymnocarpietum dryopteridis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 1 du tableau 5 in Wali (1969, Vegetation-environment relationships… : 33). Là encore, cette unité II a valeur d’alliance, surtout caractérisée par Viola orbiculata, Prosartes hookeri (Liliaceae), Clintonia uniflora (Liliaceae), Maianthemum amplexicaule, M. canadense (figure 13), Tiarella trifoliata subsp. t. (Saxifragaceae), Platanthera orbiculata, Osmorhiza berteroi (Apiaceae), Aralia nudicaulis, Aconitum columbianum, Streptopus lanceolatus, partageant avec l’alliance précédente Vaccinium cespitosum et Oryzopsis asperifolia, et Equisetum pratense avec l’unité IV, le Violo obiculatae-Clintonion uniflorae all. nov. hoc loco, typus nominis hoc loco : le Galio triflori-Gymnocarpietum dryopteridis nov.
Le troisième ensemble (colonnes 9 à 15) est marqué par le GS à Vaccinium vitis-idaea et autres Ericaceae qui caractérisent des landes boréo-américaines. Les colonnes 10 à 12, synthétisant 73 relevés, peuvent être rattachées au Rhododendro groenlandici-Vaccinietum vitis-idaeae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 13 de l’appendice 4 in Krestov et al. (2000, Scientia Silvica 26 : 64, en remplaçant Ledum groenlandicum par Rhododendron g.). Caractérisée par Vaccinium vitis-idaea, Betula nana, Arnica cordifolia, Delphinium glaucum, Symphyotrichum ciliolatum, Osmorhiza berteroi, Neottia cordata, Empetrum nigrum subsp. hermaphroditum, cette unité III décrit le Rhododendro groenlandici-Vaccinion vitis-idaeae all. nov. hoc loco, typus nominis hoc loco : le Rhododendro groenlandici-Vaccinietum vitis-idaeae nov. Une végétation affine est décrite par Vitt et al. (1975, tableau 1 : relevés 26 à 37), quoique sans doute plus proche des tourbières du Kalmio-Sphagnion fusci.
Les colonnes 16 et 17 du dernier ensemble décrivent des végétations plus hygrophiles statistiquement encore mal définies.
4.2. Du Québec
Les tableaux 11 à 13 synthétisent diverses données sur les sous-bois herbacés vivaces du Québec qui se répartissent en sept unités assez inégales, plusieurs à valeur d’alliance, mais de telles unités ne seront pas définies avant la large synthèse finale (tableau 16).
Le tableau 11 est consacré à une première partie de la végétation intraforestière québécoise. Les colonnes 1 à 4 sont différenciées par le GS à Apocynum androsaemifolium. Les colonnes 1 et 2 peuvent être réunies dans l’Epigaeo repentis-Pteridietum latiusculi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 406 du tableau 40 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et Forêts du Québec 14 hors texte). Assez bien différenciée, la colonne 3 est le Solidagini canadensis-Pteridietum latiusculi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 415 du tableau 40 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et Forêts du Québec 14 hors texte).
Pour les colonnes 5 à 10, Blouin et Grandtner (1971) évoquent une végétation à Pteridium aquilinum, il semble cependant que ce soit plutôt à nouveau P. latiusculum, seul taxon de ce genre cité du Québec par Marie-Victorin (1935), ce qui est confirmé par POWO. Ces colonnes sont donc liées par le GS à Lycopodium complanatum. Les colonnes 6 à 8 correspondent au Solidagini rugosae–Pteridietum latiusculi ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 5988 du tableau 144 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Pteridium aquilinum par P. latiusculum). La colonne 10 peut être décrite comme Cypripedio acaulis–Maianthemetum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 5904 du tableau 146 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Pteridium aquilinum par P. latiusculum).
Le tableau 12 concerne une deuxième partie de la végétation intraforestière québécoise. Les colonnes 11 à 19 sont fortement caractérisées par le GS à Rubus pubescens. Les colonnes 11 et 12 peuvent être rattachées au Carici flavae-Calamagrostietum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 6576 du tableau 141 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte). La colonne 14 paraît décrire une mégaphorbiaie en lien avec les aulnaies-frênaie du Fraxino nigrae-Alnion rugosae, le Veratro viridis-Onocleetum struthiopteridis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 5831 du tableau 149 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Matteuccia struthiopteris par Onoclea s.). La colonne 15 est le Botrychio virginiani-Aralietum nudicaulis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 5907 du tableau 142 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte).
Dans tout cet ensemble des colonnes 11 à 19, apparaissent Sanguisorba canadensis, Chelone glabra (Lamiaceae), Iris versicolor (figure 14), Thalictrum pubescens… C’est l’occasion de rappeler que Géhu et Géhu-Franck (1985) ont décrit une mégaphorbiaie laurentienne, le Chelono glabrae-Sanguisorbetum canadensis, dans laquelle se naturalisent Lythrum salicaria, Lysimachia vulgaris, Filipendula ulmaria, Symphytum officinale, quatre éléments des mégaphorbiaies européennes.
En dépit de seulement trois colonnes marquées par le GS à Neottia cordata, l’ensemble des colonnes 20 à 22 ne se réduit pas aux ensembles voisins. C’est surtout la colonne 21 qui est bien caractérisée : Lycopodio annotini-Gaultherietum hispidulae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 826 du tableau 2 in Damman (1964, Forest Sci. Monographs 8 : 17).
Les colonnes 23 à 29 partagent le GS à Rhododendron groenlandicum et d’autres Ericaceae caractérisant les tourbières bombées et les landes boréo-américaines. Se détachent tout particulièrement
- la colonne 25 (et colonne 26 ?) comme Maianthemo trifolii-Gaultherietum hispidulae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 5931 du tableau 140 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Smilacina trifolia par Maianthemum trifolium et Chiogenes hispidula par Gaultheria h.) ;
- la colonne 28 comme Melampyro linearis-Chamaedaphnetum calyculatae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 5950 du tableau 140 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Cassandra calyculata par Chamaedaphne c.).
Le tableau 13 prolonge la présentation de la diversité des associations herbacées des sous-bois québécois. L’unité X (colonnes 30 à 46) est irrégulièrement caractérisée, un petit GS à Lycopodium complanatum en étant le marqueur. Peu de syntaxons floristiquement et statistiquement significatifs se détachent, en dehors peut-être des colonnes 36 à 39 différenciées par Corallorhiza trifida : Corallorhizo trifidae-Lysimachietum borealis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 5920 du tableau 143 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Trientalis borealis par Lysimachia b.).
L’unité XI (colonnes 47 à 56) est mieux caractérisée, notamment par le GS à Equisetum sylvaticum. Parmi les syntaxons bien définis, on peut retenir
- en colonne 47, le Botrychio virginiani-Maianthemetum canadensis ass. nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 260 du tableau 42 in Gagnon & Grandtner (1973, Mémoires du ministère des Terres et Forêts du Québec 14 hors texte) ;
- en colonne 52, le Maianthemo trifolii-Coptideteum trifoliae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 5819 du tableau 141 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et Forêts du Québec 6 hors texte, en remplaçant Smilacina trifolia par Maianthemum trifolium et Coptis groenlandica par C. trifolia ; figures 15 et 16).

4.3. D’autres données nord-américaines, notamment des États-Unis
Les tableaux 14 et 15 synthétisent la végétation basse (à chaméphytes et cryptophytes) intraforestière déduit de 386 relevés publiés par Rivas-Martínez et al. (1999). Tout d’abord le tableau 14 fait apparaître trois ensembles, codés de XII à XIV.
L’unité XII (colonnes 1 à 5) rassemble des syntaxons plutôt cryptophytiques caractérisé par le GS à Maianthemum canadense et Aralia nudicaulis. La colonne 2 peut être isolée comme Mertensio paniculatae-Galietum triflori ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 8 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 65), la 3 comme Pyrolo asarifoliae-Linnaeetum americanae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 2 du tableau 57 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 184), la 4 comme Lysimachio borealis-Cornetum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 9 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 67, en remplaçant Trientalis borealis par Lysimachia b.), et la 5 comme Dryopterido campylopterae-Oclemenetum acuminatae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 1 du tableau 59 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 188, en remplaçant Aster acuminatus par Oclomena acuminata). Tout cet ensemble I, caractérisé par Aralia nudicaulis (Araliaceae) et partageant avec l’unité XIII Maianthemum canadense, Lysimachia borealis (figure 17), Clintonia borealis (Liliaceae ; figure 18) et Actaea rubra (figure 19), possède la valeur d’une alliance, le Linnaeo americanae-Aralion nudicaulis all. nov. hoc loco, typus nominis hoc loco : le Pyrolo asarifoliae-Linnaeetum americanae nov. On reconnaît cette alliance dans des lisières québécoises citées par Ansseau & Grandtner (1986), dans des données du ministère québécois des Ressources naturelles et des Forêts (2023, par exemple p. 105) et de Warder (1970, tableau 7).

L’ensemble XIII (colonnes 6 à 10) combine le GS à Maianthemum canadense et celui à Vaccinium angustifolium. La colonne 7 est le Lycopodietum obscuro-annotini ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 2 du tableau 5 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 59), la 9 l’Eriophoro brachyantheri-Vaccinietum oxycocci ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 1 du tableau 7 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 63), correspondant à une tourbière bombée à Sphagnum et Ericaceae dont l’assèchement favorise le développement d’une association arborescente, le Larici laricinae-Piceetum marianae (cf. § 2), où Eriophorum brachyanthemum est homologue d’E. vaginatum. La colonne 10 est l’Aralio hispidae-Vaccinietum angustifolii ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 60 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 191). Caractérisé par Coptis trifolia (Ranunculaceae), Gaultheria hispidula (Ericaceae), Epigaea repens (Ericaceae), Vaccinium angustifolium, Rhododendron groenlandicum, cet ensemble XIII peut être considéré comme le Gaultherio hispidulae-Vaccinion angustifolii all. nov. hoc loco, typus nominis hoc loco : le Lycopodietum obscuro-annotini nov. Les tourbières bombées évoquées à propos de l’Eriophoro brachyantheri-Vaccinietum oxycocci ont été étudiées par Damman (1977, 1980).
Le large ensemble XIV (colonnes 11 à 22) est plutôt chaméphytique et bien caractérisé par le GS à Vaccinium vitis-idaea (souvent appelé V. minus dans la monographie utilisée). La colonne 11 est interprétée ici comme Carici lugentis-Vaccinietum uliginosi ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 10 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 70), la 12 comme Corno canadensis-Rhododendretum groenlandici ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 2 du tableau 11 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 72), la 13 comme Mertensio paniculatae-Vaccinietum vitis-idaeae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 9 du tableau 12 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 75), la 15 comme Gymnocarpio dryopteridis-Cornetum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 26 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 114), la 16 comme Festuco altaicae-Lupinetum arctici ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 6 du tableau 54 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 179), la 17 comme Arctagrostio arundinaceae-Rhododendretum groenlandici ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 5 du tableau 55 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 181, en remplaçant Ledum groenlandicum par Rhododendron g.), la 19 comme Rubo arctici-Empetretum hermaphroditi ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 67 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 210) et la 18 comme Rhododendro tomentosi-Empetretum hermaphroditi ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 68 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 211, en remplaçant Ledum decumbens par Rhododendron tomentosum). La colonne 22 est le Phyllodoco glanduliflorae-Vaccinietum scoparii ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 43 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 151). Cet ensemble XIV est suffisamment caractérisé par Vaccinium vitis-idaea, Empetrum nigrum subsp. hermaphroditum, Rubus chamaemorus, Geocaulon lividum (Santalaceae), Mertensia paniculata (Boraginaceae ; figure 20), Lupinus arcticus, Arctagrostis arundinacea (Poaceae), Petasites frigidus subsp. f., Festuca altaica, Arctous rubra (Ericaceae), Equisetum pratense, Pedicularis labradorica, Athyrium cyclosorum, Anticlea elegans (Melanthiaceae ; figure 21), Cornus suecica (figure 22)… pour justifier la définition du Linnaeo americanae-Empetrion hermaphroditi all. nov. hoc loco, typus nominis hoc loco : le Festuco altaicae-Lupinetum arctici nov.

Le tableau 15 poursuit l’analyse de cette végétation basse intraforestière en rapprochant trente-cinq autres colonnes qui se répartissent en quatre ensembles codés de XV à XVIII. L’unité XV (colonnes 23 à 32) est caractérisée par le GS à Polystichum munitum. La colonne 23 peut être définie comme Maianthemo dilatati-Vaccinietum ovati ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 10 du tableau 15 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 86), la colonne 24 comme Vancouverio hexandrae-Polystichetum muniti ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 10 du tableau 5 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 92), la colonne 25 comme Achlydo triphyllae-Vaccinietum parvifolii ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 18 du tableau 19 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 94), la colonne 26 comme Maianthemo stellati-Chimaphiletum umbellatae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 4 du tableau 20 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 97, Chimaphila occidentalis étant inclus dans Ch. umbellata), la colonne 27 comme Galio trifidi-Polystichetum muniti ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 33 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 128), la colonne 29 comme Clintonio uniflorae-Gaultherietum ovatifoliae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 13 du tableau 25 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 111), la colonne 31 est proche du Maianthemo dilatati-Vaccinietum ovati précédent, mais floristiquement bien plus pauvre. Tout cet ensemble XV est hautement caractérisé par Polystichum munitum, Pteridium pubescens, Trillium ovatum (Melanthiaceae), Maianthemum dilatatum, M. stellatum (figure 23), Vaccinium parvifolium, V. ovatum, Lysimachia latifolia, Achlys triphylla (Berberidaceae), Adenocaulon bicolor (Asteraceae), Blechnum spicant, Gaultheria ovatifolia (Ericaceae), Asarum caudatum, Anemonastrum deltoideum (Ranunculaceae), Streptopus lanceolatus, Oxalis oregana, Stachys mexicana, Marah oregana (Cucurbitaceae), ce qui justifie la définition du Trillio ovati-Polystichion muniti all. nov. hoc loco, typus nominis hoc loco : l’Achlydis triphyllae-Vaccinietum parvifolii nov.
L’ensemble XVI (colonnes 33 à 42) est moins nettement caractérisé que le précédent, mais ne peut de toute façon pas y être inclus. La colonne 33 est le Streptopodo americani-Tiarelletum trifoliatae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 17 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 90), la colonne 34 est l’Aralio nudicaulis-Tiarelletum unifoliatae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 5 du tableau 22 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 102), la colonne 36 le Maianthemo stellati-Linnaeetum americanae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 49 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 165), la colonne 37 l’Arnico cordifoliae-Cornetum canadensis ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 3 du tableau 42 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 149), la colonne 38 le Luzulo piperi-Vaccinietum membranacei ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 8 du tableau 23 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 105), la colonne 39 le Luzulo hitchcockii-Valerianetum sitchensis ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 9 du tableau 24 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 109), la 40 l’Osmorhizae depauperatae-Pyroletum asarifoliae ass. nov. hoc loco, typus nominis hoc loco : composante basse du relevé 9 du tableau 44 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 153). Cet ensemble XVI peut être interprété comme une alliance caractérisée par Veratrum viride (figure 24), Actaea rubra, Valeriana sitchensis, Arnica cordifolia, A. latifolia, Streptopus streptopoides subsp. s., partageant en outre le GS à Vaccinium membranaceum avec l’alliance précédente, le Veratro viridis-Vaccinion membranacei all. nov. hoc loco, typus nominis hoc loco : l’Arnico cordifoliae-Cornetum canadensis nov.
L’unité XVII (colonnes 43 à 47) rassemble cinq syntaxons reliés plutôt négativement par l’absence des taxons différenciant les quatre autres ensembles du tableau 15. L’unité XVIII (colonnes 48 à 57), enfin, est bien caractérisée par le GS à Carex rossii. On peut y mettre en exergue
- la colonne 49 comme Doellingerio engelmannii-Arnicetum latifoliae ass. nov. hoc loco, typus nominis hoc loco: composante basse du relevé 5 du tableau 40 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 145, en remplaçant Aster engelmannii par Doellingeria e.) ;
- la colonne 50 comme Arctostaphylo patulae-Boecheretum lemmonii nov. hoc loco, typus nominis hoc loco: composante basse du relevé 3 du tableau 41 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 147, en remplaçant Arabis lemmonii par Boechera l.) ;
- la colonne 51 comme Poo stenanthae-Arctostaphyletum klamathensis ass. nov. hoc loco, typus nominis hoc loco: composante basse du relevé 3 du tableau 27 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 115) ;
- la colonne 52 comme Pyrolo chloranthae-Paxistimetum myrisinitae nov. hoc loco, typus nominis hoc loco: composante basse du relevé 4 du tableau 45 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 156) ;
- la colonne 54 comme Vaccinio scoparii-Arctostaphyletum uvae-ursi nov. hoc loco, typus nominis hoc loco: composante basse du relevé 9 du tableau 47 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 160, Arctostaphylos uva-ursi incluant A. adenotricha selon POWO) ;
- la colonne 56 comme Pulsatillo multifidae-Elymetum glauci nov. hoc loco, typus nominis hoc loco: composante basse du relevé 1 du tableau 52 in Rivas-Martínez et al. (1999, Itinera Geobot. 12 : 171).
Caractérisé par Carex rossii, Calamagrostis rubescens, Berberis repens, Antennaria rosea, Poa fendleriana, Thalictrum sparsiflorum, Penstemon whippleanus (Plantaginaceae), Pterospora andromedea (Ericaceae ; figure 25), Boechera lemmonii (Brassicaceae), Arnica cordifolia, Arctostaphylos uva-ursi, cet ensemble a la valeur d’une alliance, le Berberido repentis-Caricion rossii all. nov. hoc loco, typus nominis hoc loco : le Vaccinio scoparii-Arctostaphyletum uvae-ursi nov.
4.4. Synthèse de la végétation basse vivace
La description des syntaxons herbacés intraforestiers nord-américains a donc été menée sur la base des tableaux 10 à 15 et a donné lieu à la définition de neuf alliances sur les dix-huit synthétisées dans le tableau 16. Ce tableau tente une synthèse plus large afin de préciser les relations synsystématiques entre celles-ci en ne retenant que les colonnes synthétiques par alliance (unités I à XVIII).
Une coupure majeure passe entre les unités XIV et XVII. Dans l’ensemble formé des unités I à XIV une coupure secondaire sépare deux sous-ensembles (I à IV d’un côté, VII à XIV d’un autre). Le premier de ces deux sous-ensembles rassemble six unités à valeur d’alliance, dont cinq déjà définies plus haut ; il a alors la valeur d’un ordre caractérisé par Pyrola asarifolia, Geocaulon lividum (Santalaceae), Petasites frigidus subsp. f., Galium boreale, Clintonia uniflora (Liliaceae), Goodyera oblongifolia, Equisetum scirpoides, Mertensia paniculata (Boraginaceae), Rubus parviflorus, Gymnocarpium dryopteris, Tiarella trifoliata subsp. unifoliata (Saxifragaceae), Chimaphila umbellata (figure 26), les Pyrolo asarifoliae-Cornetalia canadensis ord. nov. hoc loco, typus nominis hoc loco : le Violo orbiculatae-Clintonion uniflorae nov.
En raison d’une forte différenciation floristique et biologique, on propose de subdiviser l’ensemble des unités VII à XIV en deux ordres. Le premier (unités VII à VIII) est caractérisé par de nombreux taxons et inclut diverses alliances :
- l’unité VII comme Thalictro pubescentis-Solidaginion rugosae nov. hoc loco, à valeur de mégaphorbiaie, caractérisée par Solidago rugosa, Galium palustre, G. asprellum, Carex stipata, C. crawfordii, C. echinata subsp. e., Thalictrum pubescens, Dryopteris cristata, Juncus effusus, Euthamia graminifolia, Epilobium leptophyllum, Lycopus uniflorus, L. americanus (figure 27), Sanguisorba canadensis, Chelone glabra (Lamiaceae), Myosotis laxa *l., Calamagrostis canadensis (figure 28), Glyceria striata, Trillium cernuum (figure 29), Ageratina altissima…, typus nominis hoc loco: le Veratro viridis-Onocleetum struthiopteridis nov. ;
- l’unité XI comme Gymnocarpio disjuncti-Dryopteridion carthusianae nov. hoc loco, à valeur de mégaphorbiaie, caractérisée par plusieurs fougères, certaines partagées avec l’alliance précédente, dont Gymnocarpium disjunctum, Osmundastrum cinnamomeum (figure 30), Thelypteris noveboracensis, Onoclea sensibilis (Aspleniaceae ; figure 31), typus nominis hoc loco: le Botrychio virginiani-Maianthemetum canadensis nov. ; une végétation relevant très probablement aussi de cette alliance est aussi rapportée par Desponts et al. (2004, tableau A1, Herbaceous plants) ;
- l’unité V comme Oryzopsio asperifoliae-Apocynion androsaemifolii nov. hoc loco, caractérisée par Apocynum androsaemifolium (Apocynaceae), Solidago nemoralis subsp. decemflora, Fragaria vesca subsp. americana, Gaultheria procumbens, Viola septentrionalis, Hypericum boreale, Pyrola americana…, typus nominis hoc loco: le Solidagini canadensis-Pteridietum latiusculi nov. ;
- l’unité VIII comme Neottio cordatae-Gaultherion hispidulae nov. hoc loco, caractérisée par Neottia cordata, Goodyera repens, Clintonia uniflora, typus nominis hoc loco: le Lycopodio annotini-Gaultherietum hispidulae nov.

Cet ensemble des unités VII à VIII est un ordre fortement caractérisé par Dryopteris carthusiana, Viola incognita s. l., V. renifolia, Eurybia macrophylla (Asteraceae), Lycopodium obscurum (figure 32), L. clavatum, Streptopus lanceolatus, Carex arctata, Solidago macrophylla, S. canadensis, Monotropa uniflora (figure 33), Huperzia lucidula (figure 34), Doellingeria umbellata (Asteraceae), Nabalus altissimus (Asteraceae), Trillium erectum, T. undulatum (Melianthaceae ; figures 35 et 36), Anaphalis margaritacea (Asteraceae), Agrostis scabra, Maianthemum racemosum (figure 37), Botrychium virginianum (figure 38), Malaxis unifolia (Orchidaceae ; figure 39), Osmunda claytoniana (figure 40), Tiarella cordifolia (Saxifragaceae ; figure 41), Mitchella repens (Rubiaceae, figure 42)…, les Streptopodo lanceolati-Eurybietalia macrophyllae ord. nov. hoc loco, typus nominis hoc loco : le Thalictro pubescentis-Solidaginion rugosae nov.
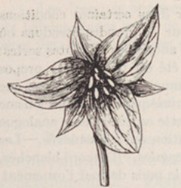



Le petit ensemble des unités IX à XIV comporte quatre alliances, dont trois déjà nommées plus haut ; seule l’unité IX reste à nommer en tant que Chamaedaphno calyculatae-Rhododendrion canadensis all. nov. hoc loco, rassemblant des landes dérivées de tourbières bombées par assèchement, caractérisée ou différenciée des alliances voisines par plusieurs Ericaceae (Rhododendron canadense, Chamaedaphne calyculata, Kalmia polifolia, Andromeda polifolia subsp. latifolia), ainsi que Drosera rotundifolia, Eriophorum vaginatum, E. tenellum, Sarracenia purpurea, Maianthemum trifolium. ; on reconnaît encore cette alliance chez Bubier (1991, tableau 1 : relevés 5 à 17) et dans la fiche dressée par Baldwin & Chapman (2017). Globalement, ces quatre unités forment un ordre caractérisé par Rhododendron groenlandicum, Calamagrostis purpurascens, Rubus chamaemorus, Vaccinium oxycoccos, les Corno canadensis-Rhododendretalia groenlandici ord. nov. hoc loco, typus nominis hoc loco : le Chamaedaphno calyculatae-Rhododendrion canadensis nov.
Finalement tout l’ensemble des unités I à XIV peut être assimilé à une grande classe de végétation vivace basse nord-américaine intraforestière, les Corno canadensis-Linnaeetea americanae classis nov. hoc loco, caractérisée par Cornus canadensis (figure 43), Linnaea borealis subsp. americana, Rubus pubescens, Galium triflorum, Lycopodium annotinum, L. complanatum, Mitella nuda (Saxifragaceae ; figure 44), Vaccinium myrtilloides, Fragaria virginiana subsp. v., Actaea rubra…, ainsi que des taxons s’étendant sur deux des trois ordres : Maianthemum canadense, Lysimachia borealis, Aralia nudicaulis (Araliaceae), Coptis trifolia (Ranunculaceae), Clintonia borealis, Oclemena acuminata (Asteraceae), Gaultheria hispidula (Ericaceae), Vaccinium angustifolium, Oxalis montana, Carex trisperma…, typus nominis hoc loco : les Streptopodo lanceolati-Eurybietalia macrophyllae nov.


Il est intéressant de rapprocher cette végétation à Cornus canadensis et Linnaea borealis subsp. americana de sa vicariante scandinave où C. suecica (figure 22) remplace C. canadensis et L. borealis subsp. b. remplace la sous-espèce americana. Dans son tableau 1, Regel (1927, tableau 1 : relevés 367 à 407) en publie trente relevés permettant de dresser la liste synthétique suivante :
C. suecica V, Vaccinium myrtillus V, V. vitis-idaea III, Gymnocarpium dryopteris III, Avenella flexuosa *f. III, Empetrum nigrum *hermaphroditum III, Solidago virgaurea *lapponica III, Equisetum sylvaticum II, Vaccinium uliginosum II, Epilobium angustifolium II, Lysimachia europaea II, Linnaea borealis *b. I, Lycopodium annotinum I, Pedicularis lapponica I, Melampyrum sylvaticum I, M. pratense I, Calluna vulgaris I et des taxons de présence + et r
et de définir le Vaccinio myrtilli-Cornetum suecicae ass. nov. hoc loco, typus nominis hoc loco : relevé 1112 du tableau 1 in Regel (1927, Veröff. Geobot. Inst. Rübel in Zürich 4 hors texte).
4.5. Quelques autres végétations herbacées vivaces intraforestières
La végétation vivace du sous-bois du Fraxino rubrae-Salicetum fragilis québécois cité en 2.3 est bien différente de celle abordée dans les paragraphes précédents (d’après Lacoursière & Grandtner, 1972, tableau XXXV, sur cinq relevés) :
- combinaison caractéristique : Onoclea sensibilis V, Stachys tenuifolia V, Thalictrum pubescens V, Amphicarpaea bracteata (Fabaceae) V, Cicuta maculata IV, Geum canadense IV, Circaea canadensis *quadrisulcata III, Heracleum maximum III, Agrimonia gryposepala III, Lysimachia terrestris III, Woodwardia areolata (figure 45) II, Carex deweyana II, crinita II, Symplocarpus foetidus (Araceae) II, Iris versicolor II, Chelone glabra II, Eutrochium maculatum (Asteraceae) II,
- autres taxons : Lythrum salicaria V, Mentha canadensis IV, Convolvulus sepium IV, Galium palustre IV, Lycopus uniflorus II, Equisetum palustre II, fluviatile II, Elymus virginicus II, Caltha palustris II, Epilobium angustifolium II, Carex rostrata II, C. stipata I, Myosotis laxa *l. I, Epipactis helleborine I, Arisaema triphyllum (Araceae) I, Euthamia graminifolia I.
Il s’agit d’une association très originale de mégaphorbiaie intraforestière, le Thalictro pubescentis-Onocleetum sensibilis ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 4 du tableau XXXV in Lacoursière & Grandtner (1972, Nat. Can. 99 (5) : 503). L’injection de cette colonne synthétique dans le tableau 16 montre que cette association possède des liens synfloristiques avec les alliances IV et VII, surtout la VII puisque la colonne IV est liée à la Colombie-Britannique, donc à l’opposé du Québec. Ces deux alliances hygrophiles paraissent vicariantes l’une de l’autre dans ces régions ; d’ailleurs elles partagent Onoclea struthiopteris, Geum macrophyllum, G. rivale, Mentha arvensis, Heracleum maximum, Platanthera dilatata. Mais ce qui les distingue floristiquement est nettement plus fort que ce qui les rapproche. De telles mégaphorbiaies sont citées aussi de Saint-Pierre-et-Miquelon (Muller & Etcheberry, in Muller, 2006).
L’association herbacée subordonnée au Nysso sylvaticae-Aceretum rubri décrit en 2.3 (tableau 5 des auteurs) est ainsi constituée :
Maianthemum canadense V, Symplocarpus foetidus V, Rubus hispidus IV, Carex retroflexa III, C. intumescens II, C. debilis *rudgei II, C. stricta II, Solidago caesia II, Thalictrum pubescens II, Dryopteris cristata II, D. intermedia II, Thelypteris palustris II, Athyrium angustum II, Platanthera lacera II, Arisaema triphyllum I, Aralia nudicaulis I, Juncus effusus I, Osmunda regalis I, Melampyrum lineare I et des taxons de présence +,
décrivant le Symplocarpo foetidi-Maianthemetum canadensis ass. nov. hoc loco, typus nominis hoc loco : relevé B de la Rubus union du tableau V in Cain & Penfound (1938, Am. Midl. Natur. 19 : 398, en remplaçant Unifolium canadense par Maianthemum c. et Spathyema foetida par Symplocarpus foetidus), à rapprocher du Thalictro pubescentis-Solidaginion rugosae (tableau 16 : unité VII).
5. Conclusion
Cette contribution à une ordination synsystématique des forêts nord-américaines a donc apporté de nombreuses unités nouvelles :
* pour la végétation arborescente
Populo tremuloidis-Piceetalia laxae avec six alliances (Piceion banksiano-marianae, Aceri spicati-Abietion balsameae, Fraxino nigrae-Alnion rugosae, Betulo mandshuricae-Piceion laxae, Pino engelmannii-Pseudotsugion glaucae, Piceo laxae-Abietion lasiocarpae), Piceetalia sitchensi-engelmannii avec deux alliances (Thujo plicatae-Tsugion heterophyllae et Populo tristis-Acerion douglasii) ;
* pour la végétation arbustive
Corylo cornutae-Diervilletea lonicerae avec Corno alternifoliae-Loniceretalia canadensis incluant quatre alliances (Hydrangeo arborescentis-Lonicerion canadensis, Salicion discolori-lucidae, Lonicero canadensis-Corylion cornutae, Ilici mucronatae–Kalmion angustifoliae), un ordre non défini pour le Rhoo typhinae-Diervillion lonicerae ; Roso sayi-Amelanchieretea alnifoliae avec Oplopanaco horridi-Rhododendretalia menziesii incluant deux alliances (Frangulo purshianae-Gaultherion shallon, Rhododendro menziesii-Alnion sinuatae), Roso sayi-Amelanchieretalia alnifoliae (incluant le Shepherdio canadensis-Juniperion depressae) ; dans une classe non définie Salicetalia acutifolio-alaxensis ; plus une alliance non placée, le Viburno edulis-Lonicerion involucratae ;
* pour la végétation vivace basse
Corno canadensis-Linnaeetea americanae avec Pyrolo asarifoliae-Cornetalia canadensis incluant cinq alliances (Vaccinio cespitosi-Arctostaphylion uvae-ursi, Violo orbiculatae-Clintonion uniflorae, Trillio ovati-Polystichion muniti, Veratro viridis-Vaccinion membranacei, Rhododendro groenlandici-Vaccinion vitis-idaeae), Streptopodo lanceolati-Eurybietalia macrophyllae incluant quatre alliances (Thalictro pubescentis-Solidaginion rugosae, Gymnocarpio disjuncti-Dryopteridion carthusianae, Oryzopsio asperifoliae-Apocynion androsaemifolii, Neottio cordatae-Gaultherion hispidulae), Corno canadensis-Rhododendretalia groenlandici incluant quatre alliances (Chamaedaphno calyculatae-Rhododendrion canadensis, Linnaeo americanae-Aralion nudicaulis, Gaultherio hispidulae-Vaccinion angustifolii, Linnaeo americanae-Empetrion hermaphroditi) ; une alliance non placée, le Berberido repentis-Caricion rossii.
Sans aucun doute, ce schéma évoluera après l’apport attendu de nouvelles données. Notamment un petit point sera à revoir, la place des landes (Vaccinio cespitosi-Arctostaphylion uvae-ursi, Rhododendro groenlandici-Vaccinion vitis-idaeae, Corno canadensis-Rhododendretalia groenlandici) au milieu d’un synsystème herbacé plutôt porté par les cryptophytes. En Europe, la tendance est en effet de séparer ces deux types de végétation dans des classes distinctes ; mais il est prématuré de faire de même en Amérique du Nord.
Il me reste à espérer découvrir un jour cette prodigieuse végétation nord-américaine assez peu étudiée par les écologues locaux, plus phytoécologues que phytosociologues.
Bibliographie
Ansseau C. & Grandtner M.M., 1986. Énumération des groupements végétaux de trois secteurs forestiers des cantons de l’Est, Québec ; 1 – Groupements périforestiers. Documents phytosociologiques, n. s., X (I) : 1-75.
Ansseau C. & Grandtner M.M., 1988. Énumération des groupements végétaux de trois secteurs forestiers des cantons de l’Est, Québec ; 2 – Groupements forestiers. Documents phytosociologiques, n. s., XI : 1-55.
Baldwin K. & K. Chapman K., 2017. Picea mariana / Rhododendron groenlandicum – Kalmia angustifolia / Sphagnum spp., Sault Ste. Marie (Ontario) Canada. In Classification nationale de la végétation du Canada produit le 12 décembre 2017, Association de la classification nationale de la végétation du Canada, CNVC00282, http://cnvc-cnvc/ca [26/07/2025].
Barbour M.G., 1988. Californian Upland Forests and Woodlands. In M.G. Barbour & W.D. Billings, North American Terrestrial Vegetation, Cambridge University Press : 131-164.
Béguin C., Grandtner M.M. & Gervais C., 1995. Groupements végétaux côtiers du Saint-Laurent près de Cap-Rouge, Québec (Canada). Documents phytosociologiques, n. s., XV : 183-203.
Bliss L.C., 1988. Arctic tundra and Polar Desert Biome. In M.G. Barbour & W.D. Billings, North American Terrestrial Vegetation, Cambridge University Press : 1-32.
Bliss L.C. & Cantlon J.E., 1957. Succession on river alluvion in northern Alaska. The American Midland naturalist 58 : 452-569.
Blouin J.-L., 1970. Étude des tremblaies du bas Saint-Laurent. Thèse Université Laval, Québec, non publiée.
Blouin J.-L. & Grandtner M.M., 1971. Étude écologique et cartographie de la végétation du comté de Rivière-du-Loup. Mémoires du ministère des Terres et Forêts du Québec 6 : 1-371.
Bosseaux M.E., 1965. Végétation et flore des îles Saint-Pierre-et-Miquelon, 2. Journal d’agriculture tropicale et de botanique appliquée 12 (4-5) : 194-210.
Bouchard A., 1970. The phytosociology of the northern conifer-hardwoods of the Appallachian foothills in southern Quebec. Ph D thesis, McGill University, Montréal, 111 p. et annexes.
Bournérias M., 1972. Voyage naturaliste au Nouveau Québec, I – La végétation au sud du 55e parallèle. Science et Nature 109 : 17-27.
Bubier J.L., 1991. Patterns of Picea mariana (Black Spruce) growth and raised bog development in Victory Basin, Vermont. Bulletin of the Torrey Botanical Club 118 (4) : 399-411.
Cain S.A. & Penfound W.T., 1938. Aceretum rubri: the red mapple swamp forest of central Long Island. The American Midland naturalist 19 : 390-416.
Damman A.W.H., 1964. Some forest types of central Newfoundland and their relation to environmental factors. Forest Science Monographs 8 : 1-62.
Damman A.W.H., 1977. Geographical changes in the vegetation pattern of raised bogs in the Bay of Fundy region of Maine and New Brunswick. Vegetatio 35 (3) : 137-151.
Damman A.W.H., 1980. Ecological and floristic trends in ombrotrophic peat bogs of eastern north America. Colloques phytosociologiques VII, La végétation des sols tourbeux : 61-77.
Dansereau P., 1959. Phytogeographia Laurentiana, II – The principal plant associations of the Saint Lawrence valley. Contributions de l’Institut botanique de l’Université de Montréal 75 : 1-147.
Desponts M., Brunet G., Bélanger L. & Bouchard M., 2004. The eastern boreal old-growth balsam fir forest: a distinct ecosystem. Canadian Journal of botany 82 : 830-849.
Elliot-Fisk D.L., 1988. The Boreal Forest. In M.G. Barbour & W.D. Billings, North American Terrestrial Vegetation, Cambridge University Press : 33-62.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2022. Nouvelles considérations phytosociologiques sur la végétation de la Grèce. Carnets botaniques 109 : 1-27, https://doi.org/10.34971/Q6KD-W822.
Foucault B. (de), 2023a. Un printemps botanique et phytosociologique au pays du Matin calme (fin mars et avril 2023). Carnets botaniques 156 : 1-34, https://doi.org/10.34971/spy7-vw87.
Foucault B. (de), 2023b. Contribution à la connaissance de la végétation forestière de l’Asie nord-orientale selon le paradigme synusial. Carnets botaniques 163 : 1-18, https://doi.org/10.34971/VHBV-Q853.
Foucault B. (de), 2024a. Nouveaux regards sur la végétation des forêts chiliennes et formations associées. Carnets botaniques 178 : 1-36, https://doi.org/10.34971/W983-T852.
Foucault B. (de), 2024b. La végétation hydrophile à hygrophile de l’Ader Doutchi (République du Niger). Carnets botaniques 213 : 1-16, https://doi.org/10.34971/ZRPA-8689.
Foucault B. (de), 2025a. Nouvelles approches phytosociologiques de la végétation africaine. Carnets botaniques 248 : 1-25, https://doi.org/10.34971/MRN1-NJ46.
Foucault B. (de), 2025b. Quelques nouvelles définitions de végétations forestières algériennes. Carnets botaniques 257 : 1-5, https://doi.org/10.34971/W5BC-PC73.
Foucault B. (de) & Catteau E., 2015. Contribution au prodrome des végétations de France : les Epilobietea angustifolii Tüxen & Preising in Tüxen 1950. Journal de botanique de la Société botanique de France 70 : 63-88.
Franklin J.F., 1988. Pacific Northwest Forests. In M.G. Barbour & W.D. Billings, North American Terrestrial Vegetation, Cambridge University Press : 103-130.
Gagnon G. & Grandtner M.M., 1973. Étude éco-dynamique des tremblaies de la section laurentienne. Mémoires du ministère des Terres et Forêts du Québec 14 : 1-131.
Gauthier L., Dumont M.-C. & Grandtner M.M., 1986. Soil-vegetation relationships on a cobby limestone floodplain, Anticosti island, Québec. Acta Societatis Botanicorum Poloniae 55 (3) : 385-402.
Géhu J.-M. & Géhu-Franck J., 1984. Sur les forêts sclérophylles de chêne et de pin maritime des dunes atlantiques françaises. Documents phytosociologiques, n. s., VIII : 219-231.
Géhu J.-M. & Géhu-Franck J., 1985. La mégaphorbiaie à Sanguisorba canadensis, élément remarquable de la zonation végétale soumise aux marées d’eau douce des berges du fleuve Saint-Laurent (Québec, Canada). Colloques phytosociologiques XII, séminaire Mégaphorbiaies : 161-173.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Hall Th.F. & Penfound W.T., 1939. A phytosociological study of à cypress-gum swamp in southeastern Louisiana. The American Midland naturalist 21 (3) : 378-395.
Klinka K., Krestov P.V. & Chourmouzis C., 2002. Classification and ecology of the mid-seral Picea mariana forests of British Columbia. Applied Vegetation Science 5 : 227-236.
Krestov P.V., Klinka K., Chourmouzis C. & Kahayara G., 2000 Classification of mid-seral black spruce forests ecosystems of northern British Columbia, full report. Scientia Silvica 26 : 1-88.
Lacoursière E. & Grandtner M.M., 1971. Contribution à l’étude écologique de la végétation riparienne de l’île d’Orléans Le Naturaliste canadien 98 (3) : 443-459.
Lacoursière E. & Grandtner M.M., 1972. Les groupements végétaux ripariens entre Sainte-Famille et la pointe d’Argentenaye, île d’Orléans, Québec. Le Naturaliste canadien 99 (5) : 469-507.
Locky D.A., Bayley S.E. & Vitt D.H., 2005. The vegetational ecology of black spruce swamps, fens, and bogs in southern boreal Manitoba, Canada. Wetlands 25 (3) : 564-582.
Marie-Victorin Frère, 1935. Flore laurentienne. Imprimerie de la Salle, Montréal, 917 p.
Ministère des Ressources naturelles et des Forêts, 2023. Étages de végétation du Québec méridional. Québec, 295 p.
Muller S., 2006. Conservation de la biodiversité à Saint-Pierre-et-Miquelon, rapport de mission dans l’archipel du 15 au 29 juillet 2006. Ministère de l’Écologie et du Développement durable et Université Paul-Verlaine (Metz), 33 p.
Muller S., 2025. Bilan des connaissances sur la flore, la fonge et les habitats naturels de l’archipel de Saint-Pierre-et-Miquelon. Naturae 2025 (6) : 71-94, https://doi.org/10.5852/naturae2025a6.
Nicholson B., 1993. The wetlands of Elk Island National Park: vegetation, development, and chemistry. Ph D thesis, University of Alberta, Edmonton, 166 p.
Payette S. & Filion L., 1975. Écologie de la limite septentrionale des forêts maritimes, baie d’Hudson, Nouveau-Québec. Le Naturaliste canadien 102 : 783-802.
Peet R.K., 1988. Forests of the Rocky Mountains. In M.G. Barbour & W.D. Billings, North American Terrestrial Vegetation, Cambridge University Press : 63-101.
Penfound W.T. & Howard J.A., 1940. A phytosociological study of evergreen oak forest in the vicinity of New Orleans, Louisiana. The American Midland naturalist 23 : 165-174.
Regel C., 1927. Die Cornus suecica-Assoziationen von Nordeuropa. Veröffentlichungen der Geobotanischen Institutes Rübel in Zürich 4 : 103-122.
Rivas-Martínez S., Sánchez Mata D. & Costa M., 1999. North American boreal and western temperate forest vegetation (Syntaxonomical synopsis of the potential natural plant communities of North America, II). Itinera Geobotanica 12 : 1-316.
Vitt D.H., Achuff P. & Andrus R.E., 1975. The vegetation and chemical properties of patterned fens in the Swan Hills, north central Alberta. Canadian Journal of botany 53 : 2776-2795.
Wali M.K., 1969. Vegetation-environment relationships of sub-boreal spruce zone ecosystems in British Columbia. Thèse, Université de Colombie-Britannique, Vancouver, 281 p.
Warder M., 1970. The phytosociology of boreal forest inclusions in Southern Ontario and Quebec. McGill University, Montreal.
Wells E.D., 1976. A classification of peatlands in eastern Newfoundland. Thèse, St. John’s, Newfoundland, 201 p.


