
Contribution à la connaissance phytosociologique de la xérosère des dunes littorales non boisées d’Aquitaine (France)
Title
Contribution to the phytosociological knowledge of the xeriscape of the non-wooded coastal dunes of Aquitaine (France)
Résumé
Une étude phytosociologique menée en 2014 et 2015 et basée sur un échantillonnage systématique a permis de dresser un état de référence sur la composition et la répartition des communautés végétales mésophiles à xérophiles des dunes littorales non boisées d’Aquitaine. De nombreux syntaxons décrits des Ammophiletea australis et de l’Euphorbio portlandicae-Helichrysion stoechadis ont pu être confortés dans leur définition, typifiés ou validés. Enfin, plusieurs syntaxons originaux sont proposés ici. La forte pression d’observation a aussi permis de dresser une image fine et actualisée de la répartition de ces syntaxons du nord au sud de la dition et latéralement dans la dune.
Abstract
A reference state on the composition and distribution of the plant communities of the non-wooded coastal dunes of Aquitaine region is established following a phytosociological study based on a systematic sampling carried out in 2014 and 2015. Numerous syntaxons described in Ammophiletea australis and Euphorbio portlandicae-Helichrysion stoechadis have been confirmed in their definition, typified or validated. Finally, several original syntaxons are proposed here. The strong observation pressure also allows us to draw up a fine and updated picture of the distribution of these syntaxons from the north to the south of the dune and laterally in the dune.
1. Introduction
Depuis 2014, le Conservatoire botanique national Sud-Atlantique (CBNSA), avec l’appui financier de la direction régionale de l’Environnement, de l’Aménagement et du Logement (DREAL) Nouvelle-Aquitaine, réalise des études régionales de synthèse et d’amélioration des connaissances phytosociologiques des végétations dans le cadre de la mise en œuvre de la politique Natura 2000. Ainsi chaque année, un grand système écologique est étudié sur la base d’une synthèse bibliographique et d’une campagne de relevés de terrain.
C’est dans ce cadre qu’ont été étudiées les végétations des 230 km de dunes littorales non boisées d’Aquitaine, situées entre l’embouchure de l’Adour au sud des Landes et la pointe du Médoc en Gironde, en 2014 et 2015.
Le littoral aquitain a fait l’objet de nombreuses études phytosociologiques spécifiques (Géhu, 2008 ; Géhu & Géhu, 1975 ; Géhu & Géhu-Franck, 1975 ; Géhu et al. 1995 ; Lahondère, 1980a, 1980b ; Lazare, 2017 ; Pavillard, 1928 ; Vanden Berghen, 1964a, 1964b, 1971) ou a contribué à des synthèses nationales (Géhu, 1968b, 1968c, 1975,1982, 1994, 1995 ; Géhu & de Foucault, 1982 ; Géhu & Géhu, 1969 ; Géhu & Tüxen, 1972…). Toutefois, ces études sont pour la plupart assez anciennes et certaines végétations observées durant nos prospections ne peuvent pas être rattachées aux syntaxons qui y sont définis. Une étude phytosociologique fine, basée sur la collecte de données inédites et sur la bibliographie existante, a donc été entreprise afin de recenser de manière exhaustive les végétations présentes en précisant leur composition floristique, leur écologie, leur répartition ainsi que leur valeur patrimoniale (Lafon et al., 2015).
Ce travail se veut donc être une synthèse et un état des lieux des communautés végétales du littoral aquitain qui sont particulièrement sensibles à l’érosion naturelle, aux changements globaux et aux perturbations anthropiques. Cette étude a également permis de préciser et d’actualiser la répartition et l’écologie de nombreuses espèces remarquables (Hieracium eriophorum, Alyssum loiseleurii, Astragalus baionensis, Linaria thymifolia, Silene uniflora subsp. thorei, Solidago virgaurea subsp. macrorhiza…).
Nous présentons dans cet article uniquement nos apports à la synsystématique des végétations mésophiles à xérophiles. Les autres végétations étudiées (hygrosère, landes, fourrés…) ainsi que la géosynsystématique ont fait l’objet d’autres études (Dufay, 2014 ; Lafon et al., 2015) et ne seront donc pas abordées dans cet arcticle.
2. Méthodes
L’intégralité des 230 km de dunes littorales d’Aquitaine (Landes et Gironde) a été prospectée (carte 1). Un premier secteur de 42 km, de Tarnos à Vieux-Boucau (Landes), a été exploré en 2014 dans l’objectif de mettre au point la méthode (Dufay, 2014), mais la majeure partie des prospections a été réalisée en 2015 (Lafon et al., 2015).
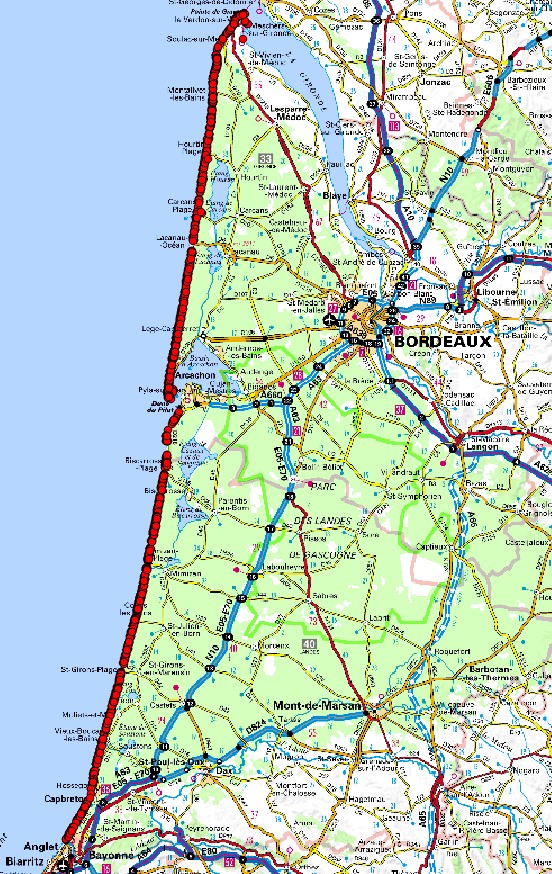
Une première période assez courte dans le temps a été programmée au début du mois d’avril 2015 spécifiquement sur les végétations prévernales. Dans ce cadre, l’ensemble du littoral de Vieux-Boucau-les-Bains (40) à la pointe du Médoc (33) a été échantillonné en différents points afin d’appréhender la diversité floristique et phytocénotique selon un gradient nord-sud. L’essentiel des prospections a ensuite été réalisé en mai, juin et début juillet.
Les dunes relictuelles d’Anglet (Pyrénées-Atlantiques), qui ont déjà fait l’objet de plusieurs études floristiques (Jeandenand & Blanchard, 2009) et phytosociologiques antérieures (Lahondère, 1980b ; Géhu, 2008) n’ont pas fait l’objet de nouvelles prospections dans ce cadre et ces données phytosocioloques ont été intégrées à cette étude.
Afin d’obtenir la chorologie précise et complète des espèces et des communautés végétales, le littoral aquitain a été subdivisé en 160 mailles de 4 km². Au sein de chacune de ces mailles, des relevés phytosociologiques ont été réalisés selon la méthode sigmatiste (Guinochet, 1973 ; Géhu & Rivas-Martínez, 1981) sur toutes les communautés végétales paraissant différentes rencontrées dans la maille.
Près de deux mille relevés phytosociologiques géoréférencés ont ainsi été effectués auxquels s’ajoutent plus de cent-cinquante relevés bibliographiques réalisés historiquement dans la dition. Une centaine de colonnes synthétiques de syntaxons présents ou potentiellement présents sur ce territoire et des régions limitrophes a également été saisie. Ces relevés ont été intégrés à la base de données de l’Observatoire de la biodiversité végétale (OBV) de Nouvelle-Aquitaine (www.obv-na.fr), base publique pour la gestion et la diffusion des données sur les habitats naturels en Nouvelle-Aquitaine.
Le jeu de données a été traité une première fois statistiquement à l’aide du module modified Twinspan (Rolececk et al., 2009) du logiciel Juice en présence-absence. En plus des groupes de végétations mésohygrophiles et aquatiques, trois grands groupes ont été différenciés : dune mobile, dune fixée, végétations forestières et pré-forestières. Chaque groupe a ensuite été analysé individuellement, tout d’abord statistiquement puis manuellement afin d’individualiser des groupes de relevés homogènes. Une colonne synthétique a été calculée pour chacun de ces syntaxons élémentaires afin de les comparer entre eux et aux syntaxons issus de la bibliographie. Au cours de cette analyse, certains relevés, dont la composition floristique paraissait comme peu typique, intermédiaire, aberrante ou considérée comme appartenant à des communautés basales ou dérivées (Kopecký & Hejný, 1974), ont été écartés.
Pour les associations composées en partie d’une synusie bryolichénique, les taxons les plus fréquents ont été rajoutés à partir de la consultation de la bibliographie (Jun, 2005 notamment). Une étude des communautés bryolichéniques est en cours sur le littoral de Nouvelle-Aquitaine et permettra de mieux faire le lien entre ces végétations.
La taxonomie suit le référentiel taxonomique national pour la faune, la flore et la fonge de France métropolitaine et d’outre-mer (Gargominy et al., 2020) ), élaboré et diffusé par le Muséum national d’histoire naturelle dans le cadre de la mise en œuvre du Système d’information sur la nature et les paysages. Seule différence, nous retenons Solidago virgaurea subsp. macrorhiza (Lange) Nyman, 1879 intégré dans la sous-espèce type dans ce référentiel. De par sa morphologie et son écologie (dunes oligotrophiles à sables non stabilisés), cette sous-espèce nous semble bien différente de la sous-espèce type et nous semble mériter le rang de sous-espèce dans les référentiels taxonomiques.
Le système phytosociologique suit le catalogue des végétations de Nouvelle-Aquitaine (Lafon et al., 2020) qui intègre les travaux récents de phytosociologie sigmatiste français et européens et pour lesquels nous proposons des modifications. Nous avons également actualisé les autorités synomenclaturales et les noms de syntaxons en suivant le code de nomenclature international (Theurillat et al., 2020).
Nous présentons uniquement les associations nouvelles et celles que nous validons. Elles sont présentées et discutées selon le gradient latéral de la plage vers la forêt, c’est-à-dire de la dune blanche (Ammophiletea…) jusqu’aux dunes fixées (Koelerio-Corynephoretea…).
3. Les pelouses de la dune blanche des Ammophiletea australis
3.1. La pelouse à Linaire à feuille de Thym et Fétuque à feuille de jonc : Linario thymifoliae-Festucetum juncifoliae P. Lafon & Le Fouler ass. nov. hoc loco [syn. : groupement à Linaria thymifolia et Festuca juncifolia P. Lafon, Le Fouler, Dufay & F. Hardy 2015]
Typus nominis hoc loco : relevé 08 du tableau 1 hoc loco.
Cette pelouse très ouverte est structurée par Festuca juncifolia et Galium arenarium. Elle se caractérise par la présence de ces deux espèces, ainsi que celle de Linaria thymifolia (photo 2) qui y trouve son optimum sur les dunes du Sud-Ouest. Cette végétation se différencie de son synvicariant septentrional, le Festuco-Galietum arenarii, par l’apparition de Linaria thymifolia et surtout par la disparition des taxons plus neutrophiles et basiphiles comme Orobanche caryophyllacea, Eryngium campestre ou encore Medicago marina (tableau 10). Elle se distingue également de son synvicariant méridional des climats à pluviométrie plus élevée, le Galio arenarii-Hieracietum eriophori, par l’absence des taxons thermophiles calcicoles et notamment Hieracium eriophorum et Astragalus baionensis (tableau 10).

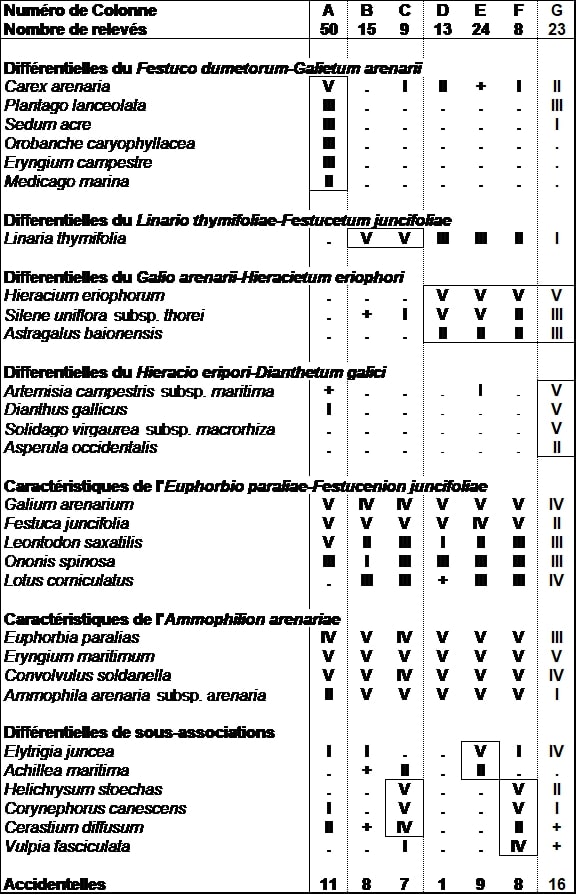
Cette végétation xérophile et neutroclinophile se développe sur des sables plus oligotrophiles et moins riches en bases que ses synvicariants. Elle se développe au niveau des zones de transit sableux où le substrat reste très meuble, du centre de l’Aquitaine (photo 1). Par son cortège original, cette pelouse que nous proposons de nommer Linario thymifoliae-Festucetum juncifoliae se positionne au sein des végétations des dunes semi-fixées de l’Euphorbio paraliae-Festucenion juncifoliae. Cette végétation synendémique de la région se répartit sur une section de littoral où aucune dune semi-fixée n’avait été identifiée jusqu’alors : du nord de la Gironde (hors pointe de Grave) au nord des Landes.

En plus du typicum, nous pouvons mettre en évidence une variation plus stabilisée, faisant transition écologiquement avec les dunes grises, et différenciée par Helichrysum stoechas, Corynephorus canescens et Cerastium diffusum. Nous proposons d’élever cette variation au rang de sous-association sous le nom d’helichrysetosum stoechadis P. Lafon & Le Fouler subass. nov. hoc loco (typus nominis hoc loco : relevé 20 du tableau 1 hoc loco).

3.2. La pelouse à Gaillet des sables et Épervière laineuse : Galio arenarii-Hieracietum eriophori Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy ass. nov. hoc loco [syn. : Galio arenarii-Hieracietum eriophori Géhu 1969 nom. nud. (art. 2b, 7)]
Typus nominis hoc loco : relevé 5 du tableau 2 hoc loco.
Cette pelouse basse et assez ouverte (photo 3) se caractérise par la présence de Hieracium eriophorum (photo 4) et Astragalus baionensis, deux espèces endémiques des dunes du sud-ouest de la France, ainsi que de Galium arenarium, Festuca juncifolia et d’autres espèces de l’Euphorbio paraliae-Ammophilion arenariae telles qu’Euphorbia paralias, Eryngium maritimum, Convolvulus soldanella, Ammophila arenaria ou encore Silene uniflora subsp. thorei. Lotus corniculatus et surtout Ononis spinosa y sont assez fréquentes et peuvent être particulièrement recouvrantes, mais Hieracium eriophorum, Galium arenarium et Festuca juncifolia restent généralement les espèces les plus structurantes et permettent une différenciation nette sur le plan physionomique de cette végétation avec la dune vive à Ammophila arenaria. Elle se différencie du Hieracio eriophori-Dianthetum gallici par l’absence des espèces plus calcicoles telles qu’Artemisia campestris subsp. maritima, Dianthus gallicus ou encore Solidago virgaurea subsp. macrorhiza.


Cette végétation xérophile, basiphile à neutrophile se développe sur des sables oligo-mésotrophes encore riches en bases. Ces sables sont moins mobiles qu’au niveau de la dune vive et sont d’une granulométrie plus importante. Cette végétation se développe du nord des Landes jusqu’à l’embouchure de l’Adour.
Cette pelouse a été mise en évidence pour la première fois par Géhu (1969) avec pour seules informations une écologie (dune blanche semi-fixée) et une chorologie (landaise, au sud d’Arachon) et pour seule caractéristique Hieracium eriophorum. À notre connaissance, il faut attendre 1979 (Lahondère) et 1982 (Géhu) pour que soient présentées des colonnes synthétiques sans holotype. Par l’apport de relevés inédits, nous proposons de valider cette association (invalide au titre des articles 2b, 7 de l’ICPN ; Theurillat et al., 2020) sous son nom d’origine de Galio arenarii-Hieracietum eriophori Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy. Du fait de son riche cortège d’espèces des dunes semi-fixées, cette pelouse se positionne clairement au sein de l’Euphorbio paraliae-Festucenion juncifoliae.
Plusieurs sous-associations avaient été mises en évidence par Géhu (1982) ; nous proposons de valider ici sur la base des nombreux relevés accumulés lors de nos prospections. La première, elytrigietosum boreoatlanticae Géhu ex subass. nov. hoc loco (typus nominis hoc loco : relevé 31 du tableau 2 hoc loco), se localise sur les zones où les mouvements de sable sont encore importants et fait la transition écologique avec le Sileno thorei-Ammophiletum arenariae voire directement avec la dune embryonnaire. Cette sous-association se différencie par la présence d’Elytrigia juncea (dont seule la sous-espèce boreoatlantica a été observée dans la région) et parfois par Achillea maritima qui peut former des buttes dans les secteurs de forte accrétion. La seconde, helichrysetosum stoechadis Géhu ex subass. nov. hoc loco (typus nominis hoc loco : relevé 44 du tableau 2 hoc loco) fait la transition avec les végétations de l’Euphorbio portlandicae-Helichrysion stoechadis et se différencie par le taxon éponyme, ainsi que par Corynephorus canescens, Vulpia fasciculata et Cerastium diffusum.

3.3. La pelouse à Épervière laineuse et Œillet de France : Hieracio eriophori-Dianthetum gallici Dufay & F. Hardy ass. nov. hoc loco [syn. : Groupement à Hieracium eriophorum et Artemisia campestris subsp. maritima P. Lafon, Le Fouler, Dufay & F. Hardy 2015, Alysso loiseleuri-Helichrysetum stoechadis Géhu 1975 nom. nud. (art. 2b, 7) p. min p.]
Typus nominis hoc loco : relevé 19 du tableau 3 hoc loco.
Cette pelouse rase et semi-fermée est structurée par la co-dominance de trois espèces vivaces neutroclinophiles à basiphiles : Artemisia campestris subsp. maritima, Dianthus gallicus et Solidago virgaurea subsp. macrorhiza. En plus de ces taxons, elle se caractérise par la présence d’espèces de dune semi-fixée telles que Hieracium eriophorum, Galium arenarium et Astragalus baionensis. Cette végétation est floristiquement proche du Galio arenarii-Hieracietum eriophori dont elle se différencie essentiellement par la présence d’Artemisia campestris subsp. maritima, Dianthus gallicus, Solidago virgaurea et Asperula occidentalis. Ainsi, dans plusieurs relevés de Lahondère (1979) qu’il rattache au Galio arenarii-Hieracietum eriophori, il est possible de reconnaître notre association. Cette association diffère de la dune grise de l’Alysso loiseleuri-Helichrysetum stoechadis par l’abondance d’espèces de dunes semi-fixées, l’absence de tapis bryo-lichénique, de Sedum acre, de Corynephorus canescens, d’Alyssum loiseleurii et la rareté d’Helichrysum stoechas.


Cette pelouse colonise le revers interne (exposé à l’est) des dunes non calibrées de type étalé du Sud landais, sur des sables oligotrophes particulièrement calcarifères. Cette végétation se développe sur des sables semi-fixés, peu stabilisés où le transit sableux reste présent.
Cette pelouse, que nous proposons de nommer Hieracio eriophori-Dianthetum gallicii Dufay & F. Hardy, est à la charnière entre les végétations de dune semi-fixée et fixée. Néanmoins, compte tenu de l’absence de mousses et de lichens, de la grande ouverture du tapis végétal, de l’absence de Corynephorus canescens et la rareté d’Helichrysum stoechas, nous la positionnons au sein des végétations de l’Euphorbio paraliae-Festucenion juncifoliae. Cette végétation est synendémique du littoral sud-landais et se répartit sur quelques kilomètres de dune au sud du département des Landes.
Certains relevés (relevés 1 à 9 du tableau 3) semblent se différencier par la présence et l’abondance plus marquée de Festuca juncifolia, Astragalus baionensis et Asperula occidentalis. Des études complémentaires seraient à mener sur ces communautés pour comprendre l’écologie de cette variation et déterminer son statut phytosociologique.
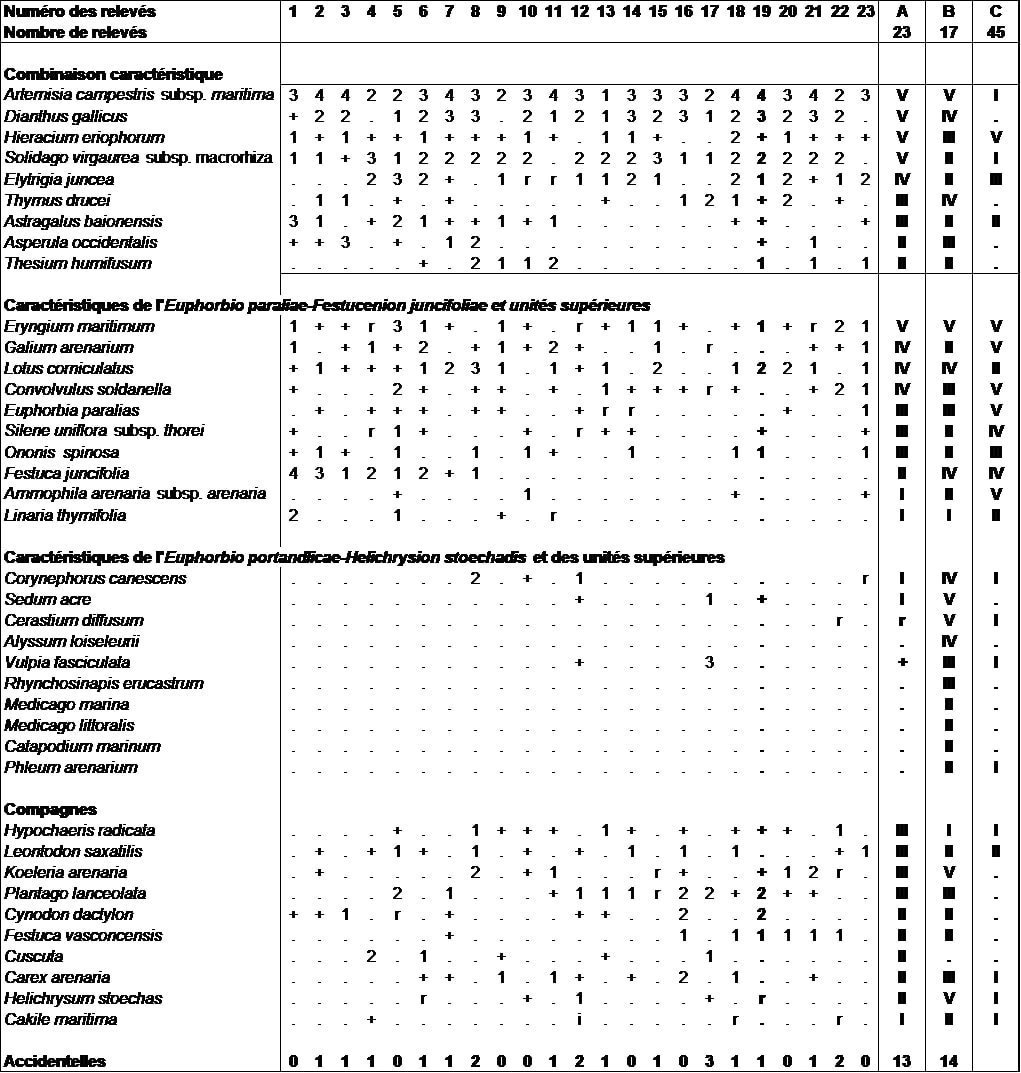
4. Les pelouses de la dune grise des Koelerio-Corynephoretea canescentis
En Aquitaine, il semble possible de distinguer deux grands types de végétation de dunes grises suivant le niveau de fixation des sables et l’éloignement à l’océan : une dune grise que l’on pourrait qualifier de pionnière et une dune grise fixée avec des sables plus décalcifiés et une strate bryolichénique bien développée qui permet un début d’enrichissement trophique (Esteve, 1980 ; Jun, 2005). Ces différences, si elles s’observent au sein de l’aire de répartition de l’Euphorbio portlandicae-Helichrysion stoechadis, pourraient donner lieu à la création de deux sous-alliances.
4.1. La pelouse à Armoise maritime et Immortelle des dunes : Artemisio maritimae-Helichrysetum stoechadis P. Lafon & Le Fouler ass. nov. hoc loco [syn. : Groupement à Saxifraga tridactylites et Koeleria arenaria P. Lafon, Le Fouler, Dufay & F. Hardy 2015, Artemisio lIoydii-Helichrysetum stoechadis Géhu 1995 nom. ined. (art. 1) et nom. nud. (2b, 3o, 7, 5)]
Typus nominis hoc loco : relevé 7 du tableau 4 hoc loco.
Cette pelouse basse et fermée où s’exprime un grand nombre d’espèces calcicoles est marquée physionomiquement par Artemisia campestris subsp. maritima (photo 7), Helichrysum stoechas et Koeleria arenaria ainsi que par un tapis de mousses et de lichens (Cladonia mediterranea, C. foliacea, Racomitrium canescens, Polytrichum juniperinum, Campylopus introflexus, Dicranum scoparium, etc.). Elle est relativement pauvre en espèces annuelles comparée aux autres végétations de dunes fixées du Sud-Ouest.

Cette communauté est proche floristiquement et géographiquement de l‘Artemisio maritimae-Ephedretum distachyae mais l’absence d’Eryngium campestre, Sonchus bulbosus, Poterium sanguisorba, Silene otites, Bupleurum baldense et surtout d’Ephedra distachya nous semble rendre impossible ce rapprochement.
Cette végétation xérophile thermo-atlantique des dunes littorales aux sables fixés et non décalcifiés est observée en Aquitaine uniquement à l’extrême pointe du Médoc, dans un secteur du littoral particulièrement soumis aux perturbations (érosion marine, fréquentation humaine…) et parfois relativement jeune comme c’est le cas des dunes du phare de Saint-Nicolas. Ce contexte est défavorable à l’installation des taxons des dunes les plus matures à Ephedra distachya. Abritant également un cortège non négligeable d’espèces de l’Euphorbio paraliae-Ammophilion arenariae, elle pourrait constituer une forme pionnière ou des sables légèrement plus décalcifiés de l’Artemisio maritimae-Ephedretum distachyae. D’ailleurs, Géhu (1995) définit, sans caractérisation précise, un Artemisio maritimae-Helichrysetum stoechadis comme une forme de cicatrisation ou de jeunesse de l’Artemisio maritimae-Ephedretum distachyae qui semble correspondre à notre communauté. Pour ces raisons nous proposons de nommer cette association Artemisio maritimi-Helichrysetum stoechadis P. Lafon & Le Fouler.
Il est intéressant de voir que les relevés classés dans l’Artemisio maritimae-Ephedretum distachyae de Lahondère (1979, 1980a) réalisés sur le littoral du Centre-Ouest correspondent très fortement à la communauté mise en évidence ici. Elle pourrait alors constituer un syntaxon présent dans toute l’aire de l’Artemisio maritimae-Ephedretum distachyae et même du Roso spinosissimae-Ephedretum distachyae (Kühnholtz-Lordat 1928) Vanden-Berghen 1958 dans le Morbihan et en Vendée (E. Glemarec & H. Guitton, comm. pers.), les liens entre ces végétations devraient être étudiés sur ce grand secteur géographique.
En plus du typicum différencié par la présence de Corynephorus canescens, il est possible de reconnaître une variation riche en taxons plus nitrophiles (Centaurea aspera, Oenothera glazioviana, etc.) ou des sols plus stabilisés ou tassés (Sedum acre, Sedum rupestre). Cette variation semble conditionnée par une pression touristique, ici très importante, qui favorise l’enrichissement trophique et le développement de taxons adaptés au piétinement répété. Le nombre non négligeable de relevés ainsi que la cohérence et l’étendue géographique de cette variation nous permettent de proposer de l’élever au rang de sous-association pouvant être nommée sedetosum acris subass. nov. hoc loco (typus nominis hoc loco : relevé 20 du tableau 4 hoc loco).

4.2. La pelouse à Silène de Porto et Immortelle des dunes : Sileno portensis-Helichrysetum stoechadis Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy ass. nov. hoc loco [syn. : Sileno portensis-Helichrysetum stoechadis Géhu 1975 nom. nud. (art. 2b, 7) p. max p.]
Typus nominis hoc loco : relevé 12 du tableau 5 hoc loco.
Cette pelouse plus ou moins ouverte est structurée par Helichrysum stoechas et Corynephorus canescens (cette dernière curieusement absente des relevés de Géhu, 1982) au sein de laquelle se développent de nombreux thérophytes comme Silene portensis (photo 9), Vulpia fasciculata (inclus dans V. membranacea par Géhu, 1982), Phleum arenarium ou encore Cerastium diffusum. Les taxons des Ammophiletea australis y sont encore fréquents mais très faiblement recouvrants. Cette végétation des dunes grises pionnières se différencie du Micropyro tenelli-Helichrysetum stoechadis notamment par la rareté des taxons thérophytiques et une strate bryolichénique le plus souvent absente.

Cette végétation xérophile, neutroclinophile à acidiclinophile se développe sur des sables oligotrophes en partie lixiviés des systèmes dunaires du centre de l’Aquitaine (photo 8). Elle se rencontre au niveau des zones où les sables sont partiellement fixés. Cette végétation occupe classiquement une partie du plateau et le revers interne du cordon dunaire érigé par Goury ainsi que certains secteurs de lettes récemment saupoudrés.

Cette pelouse a été mise en évidence pour la première fois par Géhu (1975 puis 1978) avec sa chorologie comme seule information. Il ne nous a pas été possible de trouver la trace d’une publication de ce syntaxon par Géhu en 1974, auquel pourtant un bon nombre d’articles se réfèrent, et il faut attendre 1982 (Géhu) pour que soit présentées des colonnes synthétiques mais sans typification. Par l’apport de relevés inédits, nous proposons de valider cette association (invalide au titre des articles 2b, 7 de l’ICPN ; Theurillat et al., 2020) sous son nom d’origine du Sileno portensis-Helichrysetum stoechadis Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy. Les relevés rattachés à cette association par Géhu étant assez hétérogènes, nous précisons également son écologie ainsi que son cortège caractéristique.
En plus d’un typicum nous proposons de mettre en évidence une sous-association artemisietosum maritimae subass. nov. hoc loco (typus nominis hoc loco : relevé 42 du tableau 5 hoc loco) différenciée par le taxon éponyme et se localisant sur des sables contenant vraisemblablement encore des carbonates de calcium. Cette hypothèse reste à confirmer par des études physico-chimiques du sol. Cette végétation s’étend aux dunes recalibrées du nord de la Gironde (hors pointe de Grave) au centre des Landes, tandis que la sous-association artemisietosum se localise principalement aux alentours du Bassin d’Arcachon.

4.3. La pelouse à Catapode des graviers et Immortelle des dunes : Micropyro tenelli-Helichrysetum stoechadis P. Lafon & Le Fouler ass. nov. hoc loco [syn. : Groupement à Micropyrum tenellum et Helichrysum stoechas P. Lafon, Le Fouler, Dufay & F. Hardy 2015, Sileno portensis-Helichrysetum stoechadis Géhu 1975 nom. nud. (art. 2b, 7) p. min p.]
Typus nominis hoc loco : relevé 13 du tableau 6 hoc loco.
Cette pelouse se caractérise sur le plan physionomique par une codominance entre plantes vivaces, plantes annuelles, mousses et lichens dont résulte sa grande diversité spécifique (photo 10). Les plantes vasculaires les plus couvrantes sont Helichrysum stoechas (photo 11), Corynephorus canescens, Jasione maritima gr. (comprenant J. maritima et J. montana dont la différenciation sur les dunes d’Aquitaine s’est révélée complexe) et Carex arenaria. Les taxons thérophytiques sont diversifiés et peuvent présenter de forts recouvrements. La synusie bryolichénique est notamment composée de Cladonia mediterranea, C. arbuscula, Polytrichum juniperinum, Racomitrium canescens, Campylopus introflexus, etc.


Ce syntaxon nous semble nettement se différencier du Sileno portensis-Helichrysetum stoechadis, avec lequel il est en contact, par l’absence de taxons des Ammophiletea australis, par un recouvrement et une diversité importants de lichens, par un net enrichissement en plantes annuelles acidiphiles psammophiles non exclusivement littorales telles que Tuberaria guttata, Logfia minima, Aira praecox et par la présence de taxons vivaces des sols légèrement plus riches en nutriments comme Herniaria ciliolata subsp. robusta et Rumex acetosella. Il se distingue du Thymo drucei-Corynephoretum canescentis par la présence de Micropyrum tenellum et Teesdalia nudicaulis et l’absence d’espèces neutrocalcicoles (Thymus drucei et Koeleria arenaria notamment). Cette végétation des dunes littorales fixées depuis parfois plusieurs dizaines d’années se rencontre dans la lette dunaire, en net retrait du cordon dunaire et en contrebas de la frange forestière. Les sables ont été progressivement lixiviés dans les horizons supérieurs. Cette association couvre un linéaire important en Aquitaine car elle a été observée du nord de la Gironde (excepté l’extrême pointe Médoc) jusqu’au centre des Landes (Vielle-Saint-Girons). Cette végétation particulièrement riche en phytodiversité est menacée car prise en étau entre l’érosion marine et la forêt de production.
En plus du typicum, il existe deux variations que nous proposons d’élever au rang de sous-association. Une variation à Artemisia campestris subsp. maritima et Convolvulus soldanella (ainsi que Cladonia foliacea) est centrée sur le Bassin d’Arcachon sur des sables vraisemblablement plus riches en bases, nous proposons de nommer artemisietosum maritimae subass. nov. hoc loco (typus nominis hoc loco : relevé 29 du tableau 6 hoc loco). Le cistetosum salviifolii P. Lafon & Le Fouler subass. nov. hoc loco (typus nominis hoc loco : relevé 35 du tableau 6 hoc loco), piqueté de pieds isolés de Cistus salviifolius, constitue ainsi une transition floristique vers les ourlets psammophiles littoraux thermo-atlantiques (Rubio peregrinae-Cistetum salviifolii). Cette dernière sous-association ne peut être considérée comme une phase du Micropyro tenelli-Helichrysetum stoechadis car elle est stable dans le temps.

Par sa richesse en taxons des Corynephoretalia canescentis, cette végétation semble faire transition entre cet ordre non littoral et celui strictement littoral des Artemisio maritimae-Koelerietalia albescentis. Toutefois la présence d’espèces caractéristiques de l’Euphorbio portlandicae-Helichrysion stoechadis comme Helichrysum stoechas, Herniaria ciliolata subsp. robusta ou Phleum arenarium nous conduit à l’intégrer dans cette alliance.
4.4. La pelouse à Serpolet de Druce et Corynéphore blanchâtre : Thymo drucei-Corynephoretum canescentis Dufay & F. Hardy ass. nov. hoc loco [syn. : groupement à Thymus drucei et Helichrysum stoechas P. Lafon, Le Fouler, Dufay & F. Hardy 2015]
Typus nominis hoc loco : relevé 4 du tableau 7 hoc loco.
Cette pelouse de la dune grise (photo 12) à Helichrysum stoechas associe quelques espèces neutrophiles ou basiphiles, comme Thymus drucei ou Koeleria arenaria, à des espèces acidiphiles, notamment Rumex acetosella, avec plusieurs annuelles telles que Tuberaria guttata, Logfia minima, Vulpia fasciculata, Aira praecox et Cerastium semidecandrum. Festuca vasconcensis et Herniaria ciliolata subsp. robusta y sont également fréquentes. Généralement présente, la strate bryolichénique a un recouvrement moyen qui atteint près de 60%. Elle se distingue de l’Alysso loiseleuri-Helichrysetum stoechadis, avec lequel elle partage notamment Thymus drucei, Koeleria arenaria et Helichrysum stoechas, par l’absence ou la nette raréfaction de plusieurs espèces basiphiles qui caractérisent ou qui se rencontrent dans ce dernier (Alyssum loiseleuri, Sedum acre, Dianthus gallicus, Artemisia campestris subsp. maritima, plus rarement Asperula occidentalis) et par la fréquence plus marquée d’une partie du cortège d’espèces annuelles (Tuberaria guttata, Logfia minima, Aira praecox).

Trois variations sont proposées pour être élevées au rang de sous-association. Ainsi, en plus du typicum, nous pouvons mettre en évidence un ammophiletosum arenariae Dufay & F. Hardy subass. nov. hoc loco différencié par des taxons de la dune blanche tels qu’Ammophila arenaria, Galium arenarium, plus rarement Eryngium maritimum, Hieracium eriophorum ou Euphorbia paralias (typus nominis hoc loco : relevé 14 du tableau 7 hoc loco) pouvant s’expliquer par un saupoudrage de la dune fixe. Le cistetosum salviifolii Dufay & F. Hardy subass. nov. hoc loco, qui occupe une position plus interne dans la dune, se différencie par la fréquence de taxons des ourlets psammophiles et notamment Cistus salviifolius (typus nominis hoc loco : relevé 29 du tableau 7 hoc loco).

Cette association qui relève clairement de l’Euphorbio portlandicae-Helichrysion stoechadis a été observée de l’Adour à Saint-Julien-en-Born (40). Il semble qu’elle soit synendémique de la partie méridionale du département des Landes. Cette végétation se retrouve également de manière sporadique, avec sa sous-association à Cistus salviifolius, dans les secteurs arrière-dunaires les plus décalcifiés dans l’aire de l’Alysso loiseleuri-Helichrysetum stoechadis.
4.5. La pelouse à Corbeille d’or des sables et Immortelle des dunes : Alysso loiseleurii-Helichrysetum stoechadis Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy ass. nov. hoc loco [syn. : Alysso loiseleurii-Helichrysetum stoechadis Géhu 1975 nom. nud. (art. 2b, 7) p. max p.]
Typus nominis hoc loco : relevé 13 du tableau 8 hoc loco.
Cette pelouse xérophile (photo 13) de dune grise, plus ou moins fermée, se structure par les touffes d’Helichrysum stoechas et d’Alyssum loiseleurii, entre lesquelles se développe un cortège d’espèces calcicoles caractéristiques telles que Sedum acre, Thymus drucei, Koeleria arenaria, Dianthus gallicus, ainsi que des taxons plus acidiclinophiles comme Rumex acetosella et Phleum arenarium. Une strate bryo-lichénique assez recouvrante est généralement présente et se compose de Tortula ruraliformis, Cladonia furcata, C. rangiformis, C. foliacea , etc. Cette végétation partage plusieurs espèces en commun avec son synvicariant septentrional le Thymo drucei-Corynephoretum canescentis (Thymus drucei, Koeleria arenaria, Helichrysum stoechas), mais en diffère par la présence de nombreuses espèces basiphiles et par un cortège d’annuelles moins riche et moins abondant.


Cette pelouse xérophile, basiphile se développe sur des sables oligotrophes calcarifères (riches en dépôts coquilliers) du Sud landais qui présentent une granulométrie supérieure au reste du littoral aquitain. Cette végétation ne supporte pas le transit sableux et se retrouve sur des sables fixés en position de lette dunaire.
Cette végétation a été mise en évidence pour la première fois par Géhu (1975 puis 1978) sans informations floristiques ou écologiques. Tout comme pour le Sileno portensis-Helichrysetum stoechadis, la référence dans de nombreux articles à Géhu (1974) pour ce syntaxon semble erronée. C’est en 1982 que Géhu présente une colonne synthétique quelques années après celle publiée par Lahondère (1979). Par l’apport de relevés inédits, nous proposons de valider cette association (invalide au titre des articles 2b, 7 de l’ICPN ; Theurillat et al., 2020) sous son nom d’origine de l’Alysso loiseleurii-Helichrysetum stoechadis Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy.
La définition de Géhu semblant hétérogène, nous la précisons donc en la restreignant aux végétations de dunes grises stabilisées. En outre, nous proposons également de mettre en évidence une nouvelle association de dune plus mobile : le Hieracio eriophori-Dianthetum gallici qui semblait en partie intégrée dans l’Alysso loiseleurii-Helichrysetum stoechadis. Bien que ces deux associations partagent des espèces en commun (Thymus drucei, Dianthus gallicus, Artemisia campestris subsp. maritima), l’Alysso loiseleurii-Helichrysetum stoechadis se différencie de cette dernière par de nouvelles espèces basiphiles (Alyssum loiseleurii, Sedum acre), l’absence ou la quasi-absence des espèces de dune semi-fixée et a contrario un cortège d’espèces de dune fixée plus marqué avec l’apparition d’Helichrysum stoechas, de Corynephorus canescens, de Carex arenaria et de thérophytes.
Une variation peut être observée dans les relevés de Lahondère (1979) qui ont été intégrés à notre analyse. Elle se différencie par la présence d’espèces de dune mobile telles que Eryngium maritimum, Medicago marina, Convolvulus soldanella et Festuca juncifolia. Nous n’avons pas observé cette variation au cours de notre étude, ce qui peut s’expliquer probablement par le fait que les relevés de Lahondère ont été effectués sur les dunes relictuelles d’Anglet qui n’ont pas été prospectées au cours de notre étude. Nous maintenons donc cette variation simplement au rang de variante à Eryngium maritimum. Des études complémentaires sont nécessaires pour comprendre le déterminisme écologique et phytosociologique de cette variante et surtout le lien avec le Galio arenarii-Hieracietum eriophori.
Cette association est synendémique du Sud aquitain puisqu’elle a été observée seulement sur une vingtaine de kilomètres de côtes entre Tarnos et Soorts-Hossegor au sud du département des Landes.
5. Les pelouses thérophytiques des Tuberarietea guttatae
La pelouse à Sénéçon des dunes et Téesdalie à tige nue : Senecioni denticulati-Teesdalietum nudicaulis P. Lafon ass. nov. hoc loco
Typus nominis hoc loco : relevé 3 du tableau 9 hoc loco.
Cette pelouse très ouverte se caractérise par un cortège thérophytique classique du Thero-Airion comme Teesdalia nudicaulis, Cerastium semidecandrum, Micropyrum tenellum auquel s’ajoutent des taxons spécifiques des dunes littorales comme Senecio vulgaris subsp. denticulatus et dans une moindre mesure Cerastium diffusum ou Phleum arenarium. Ce dernier taxon, du fait de son léger décalage phénologique, a été sous-inventorié lors des inventaires trop précoces et devrait constituer un taxon caractéristique de fréquence plus importante que ce que présenté ici (tableau 9). Le cortège en taxons vivaces de l’Euphorbio portlandicae-Helichrysion stoechadis y est encore présent mais la plupart de ces individus ont été observés sous une forme juvénile de première année.
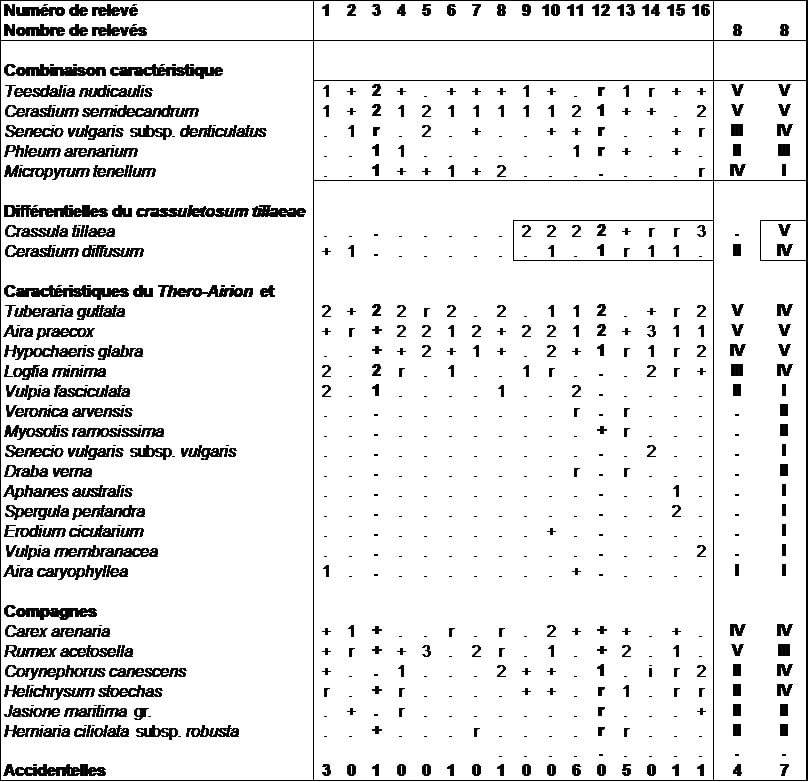
Cette végétation xérophile, neutroclinophile à acidiclinophile se développe sur des sables oligotrophes lixiviés des systèmes dunaires du centre de l’Aquitaine. Elle colonise des zones de réouverture de la dune grise par des activités anthropiques ou animales. Elle constitue ainsi une communauté de cicatrisation du Micropyro tenelli-Helichrysetum stoechadis.
Par son cortège, cette pelouse, que nous proposons de nommer Senecioni denticulati-Teesdalietum nudicaulis P. Lafon, se positionne au sein des végétations de pelouses thérophytiques du Thero-Airion. Cette végétation probablement synendémique d’Aquitaine se répartit du nord de la Gironde au nord des Landes.
Une variation, que l’on propose d’élever au rang de sous-association sous le nom de crassuletosum tillaeae P. Lafon subass. nov. hoc loco (typus nominis hoc loco : relevé 13 du tableau 9 hoc loco), se différencie par la présence de Crassula tillaea, Cerastium diffusum et la raréfaction de Micropyrum tenellum. Ce syntaxon se localise principalement au niveau des zones piétinées des sentiers.
Une autre végétation thérophytique a également pu être mise en évidence sans que le matériel phytosociologique soit toutefois suffisant pour la présenter ici. Ce groupement, nommé groupement à Senecio vulgaris subsp. denticulatus et Mibora minima, a ainsi pu être observé au nord du département de la Gironde. Il se compose d’espèces typiques des dunes (Viola kitaibeliana, Phleum arenarium, Senecio vulgaris subsp. denticulatus, Cerastium diffusum) et d’un cortège plus caractéristique du Thero-Airion comme Cerastium semidecandrum, Aphanes australis, Myosotis ramosissima, Spergula pentandra et Mibora minima qui trouve dans ce secteur ses seules populations littorales d’Aquitaine. Les relevés compilés (Lafon et al., 2015) apparaissent trop hétérogènes pour mettre en évidence un syntaxon typifiable. De plus, cette végétation semble posséder son expression optimale sur les côtes charentaises où des relevés devraient être réalisés afin d’en déceler toute son originalité.
6. Synsystème
En accord avec le code de nomenclature phytosociologique (Theurillat et al., 2020), plusieurs mutations de noms sont présentées pour les syntaxons formés par des noms de taxons qui ne sont plus reconnus actuellement dans les référentiels français (Gargominy et al., 2020 ; Tison & de Foucault, 2014).
Ammophiletea Braun-Blanq. & Tüxen ex V. Westh., J. Dijk, Passchier & G. Sissingh 1946
Ammophiletalia australis Braun-Blanq. 1933
Euphorbio paraliae-Ammophilion arenariae Géhu & Géhu-Franck 1969
Elytrigio boreoatlanticae-Honckenyenion peploidis (Tüxen in Braun-Blanq. & Tüxen 1952) Géhu 1988 nom. mut. nov. hoc loco P. Lafon (= Agropyro boreoatlantici-Minuartienion peploidis (Tüxen in Braun-Blanq. & Tüxen 1952) Géhu 1988)
- Euphorbio paraliae-Elytrigietum boreoatlanticae Tüxen in Braun-Blanq. & Tüxen 1952 mut. nov. hoc loco P. Lafon (= Euphorbio paraliae-Agropyretum junceiformis Tüxen in Braun-Blanq. & Tüxen 1952 corr. Darimont, J. Duvign. & Lambinon 1962)
- typicum
- race à Silene uniflora subsp. thorei Lafon, Le Fouler, Dufay & F. Hardy 2015
- Crithmo maritimi-Achilleetum maritimae (Pavillard 1928) Géhu 2009 mut. nov. hoc loco P. Lafon (Crithmo maritimi-Otanthetum maritimi (Pavillard 1928) Géhu 2009)
- race basque Géhu 2009
Ammophilenion arenariae Géhu 1988
- Sileno thorei-Ammophiletum arenariae (Géhu 1968) Géhu, Géhu-Franck & Bournique 1995
- typicum
- elytrigietosum boreoatlanticae Géhu, Géhu-Franck & Bournique 1995 mut. nov. hoc loco P. Lafon (elymetosum boreoatlanticae Géhu, Géhu-Franck & Bournique 1995)
- artemisietosum maritimae Géhu, Géhu-Franck & Bournique 1995
Euphorbio paraliae-Festucenion juncifoliae P. Lafon
- Festuco-Galietum arenarii Géhu 1964
- Linario thymifoliae-Festucetum juncifoliae Lafon & Le Fouler
- typicum
- helichrysetosum stoechadis Lafon & Le Fouler
- Galio arenarii-Hieracietum eriophori Géhu ex Lafon, Le Fouler, Dufay & F. Hardy
- typicum
- elytrigietosum boreoatlanticae Lafon, Le Fouler, Dufay & F. Hardy
- helichrysetosum stoechadis Lafon, Le Fouler, Dufay & F. Hardy
- Hieracio eriophori-Dianthetum gallici Dufay & F. Hardy
Nous retenons le nom d’Ammophiletea pour la classe car il s’agit de la première mention validement publiée par Westhoff et al. (1946) avant les Euphorbio paraliae-Ammophiletea australis habituellement retenus en France (Bardat et al., 2004). Nous suivons ainsi la vision de Mucina et al. (2016).
Seule l’alliance de l’Euphorbio paraliae-Ammophilion arenariae et ses trois sous-alliances sont présentes en Aquitaine. Nous ne retenons pas le nom d’Agropyro-Minuartion peploidis Tüxen in Braun-Blanq. & Tüxen 1952 pour l’alliance régionale comme le proposent Mucina et al. (2016), Marcenò et al. (2018) et de Foucault (2021) car elle est définie par ses auteurs (Braun-Blanquet & Tuxen, 1952) comme englobant les seules dunes embryonnaires. Nous suivons donc Géhu et Géhu-Franck (1969) qui lui donnent le rang de sous-alliance au sein de de l’Euphorbio paraliae-Ammophilion arenariae.
L’alliance de l’Euphorbio paraliae-Ammophilion arenariae, valide selon le code de nomenclature phytosociologique, n’a pas été typifiée par ses auteurs (Géhu & Géhu, 1969) et ne semble pas l’avoir été depuis. Nous proposons donc le Sileno thorei-Ammophiletum arenariae (Géhu 1968) Géhu, Géhu-Franck & Bournique 1995 comme association type (lectotypus nominis hoc loco) de cette alliance.
Selon de Foucault (2021) les différentes sous-alliances de l’Euphorbio paraliae-Ammophilion arenariae n’ont pas de valeur syntaxonomique. Si l’Elytrigio boreoatlanticae-Honckenyenion peploidis se différencie effectivement très faiblement de l’Ammophilenion arenariae par la quasi-absence d’Ammophila arenaria et la présence d’Elytrigia juncea subsp. boreoatlantica et Honckenya peploides, il en est tout autre pour l’Euphorbio paraliae-Festucenion juncifoliae. Ce dernier est ainsi bien individualisé par la présence de Galium arenarium, Festuca juncifolia, Leontodon saxatilis, Ononis spinosa, Lotus corniculatus et plus ponctuellement d’Artemisia campestris subsp. maritima.
En 1975, Géhu publiait l’Euphorbio-Festucion dumetorum Géhu 1975 (sans association type) qu’il attribuait au rang de sous-alliance par la suite (Géhu, 1994) sous le nom d’Euphorbio paraliae-Festucenion arenariae toujours sans préciser d’association type, ce qui la rend invalide selon le code de nomenclature (Theurillat et al., 2020). Ce nom ne peut être validé (art. 17f) pour les dunes semi-fixées puisque Festuca arenaria est absente du sud de la France et ne semble pas être une espèce de la dune semi-fixée (Catteau et al., 2021). Nous proposons comme association type (lectotypus nominis hoc loco) l’Euphorbio paraliae-Festucetum dumetorum Géhu 1963 incluse par Catteau et al. (2021) dans l’Euphorbio paraliae-Ammophiletum arenariae Tüxen in Braun-Blanq. & Tüxen 1952. De ce fait, l’Euphorbio-Festucion dumetorum devient un synonyme postérieur de l’Euphorbio paraliae-Ammophilion arenariae et l’Euphorbio paraliae-Festucenion arenariae de l’Ammophilenion arenariae .
Nous proposons donc la sous-alliance de l’Euphorbio paraliae-Festucenion juncifoliae P. Lafon suball. nov. hoc loco (typus nominis hoc loco : Galio arenarii-Hieracietum eriophori Géhu ex P. Lafon, Le Fouler, Dufay & F. Hardy) en la définissant comme une pelouse vivace des dunes semi-fixées à mouvement de sables encore important et caractérisée par Galium arenarium, Festuca juncifolia, Leontodon saxatilis, Ononis spinosa, Lotus corniculatus.
Koelerio-Corynephoretea canescentis Klika in Klika & V. Novák 1941
Artemisio maritimae-Koelerietalia arenariae G. Sissingh 1974 P. Lafon nom. mut. hoc loco (= Artemisio lloydii-Koelerietalia albescentis G. Sissingh 1974)
Euphorbio portlandicae-Helichrysion stoechadis Géhu & Tüxen ex G. Sissingh 1974
- Artemisio maritimae-Ephedretum distachyae Géhu & G. Sissingh in Sissingh 1974
- Artemisio maritimae-Helichrysetum stoechadis Lafon & Le Fouler
- typicum
- sedetosum acris Lafon & Le Fouler
- Sileno portensis-Helichrysetum stoechadis Géhu ex Lafon, Le Fouler, Dufay & F. Hardy
- typicum
- artemisietosum maritimae Lafon, Le Fouler, Dufay & F. Hardy
- Micropyro tenelli-Helichrysetum stoechadis Lafon & Le Fouler
- typicum
- artemisietosum maritimae Lafon & Le Fouler
- cistetosum salviifolii Lafon & Le Fouler
- Thymo drucei-Corynephoretum canescentis Dufay & F. Hardy
- typicum
- ammophiletosum arenariae Dufay & F. Hardy
- cistetosum salviifolii Dufay & F. Hardy
- Alysso loiseleurii-Helichrysetum stoechadis Géhu ex Lafon, Le Fouler, Dufay & F. Hardy


Tuberarietea guttatae (Braun-Blanq. ex Rivas Goday 1958) Rivas Goday & Rivas Martinez 1963 P. Lafon nom. mut. nov. hoc loco (= Helianthemetea guttati (Braun-Blanq. ex Rivas Goday 1958) Rivas Goday & Rivas Mart. 1963)
Tuberarietalia guttatae Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940 P. Lafon nom. mut. nov. hoc loco (= Helianthemetalia guttati Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940)
Thero-Airion Tüxen ex Oberd. 1957
- Senecioni denticulati-Teesdalietum nudicaulis Lafon
- typicum
- crassuletosum tillaeae Lafon
- Groupement à Senecio vulgaris subsp. denticulatus et Mibora minima Lafon, Le Fouler, Dufay & F. Hardy 2015
Cisto-Lavanduletea stoechadis Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940
Lavanduletalia stoechadis Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940
Cistion laurifolii Rivas Goday in Rivas Goday, Borja, Monasterio, Galiano & Rivas Mart. 1956
- Rubio peregrinae-Cistetum salviifolii Botineau, Bouzillé & Lahondère 1988
- typicum
- ulicetosum europaei Botineau, Bouzillé & Lahondère 1988
- Artemisio maritimae-Cistetum salviifolii Lazare 2017 P. Lafon nom. nov. hoc loco (= Artemisio lloydii-Cistetum salviifolii Lazare 2017)
Le Rubio peregrinae-Cistetum salviifolii (photo 16) et l’Artemisio maritimae-Cistetum salviifolii sont intégrés par leurs auteurs respectivement dans le Teucrion scorodoniae, alors placé dans les Trifolio medii-Geranietea sanguinei (Botineau et al., 1988) mais correspondant actuellement aux Melampyro pratensis-Holcetea mollis, et le Cisto salviifolii-Ericion cinereae (Lazare, 2017) des Calluno vulgaris-Ulicetea minoris.

Un rattachement aux ourlets des Melampyro pratensis-Holcetea mollis est à exclure du fait de la trop faible représentation des espèces de cette classe (Rubia peregrina, Lonicera periclymenum), sauf en partie pour l’ulicetosum europaei, et de la dominance de taxons chaméphytiques frutescents. Un rattachement aux landes des Calluno vulgaris-Ulicetea minoris n’apparaît pas possible du fait de l’absence des espèces caractéristiques Erica cinerea et Calluna vulgaris.
Nous proposons d’intégrer ces végétations dans les chaméphytaies méditérranéennes des Cisto-Lavanduletea stoechadis et par défaut dans le Cistion laurifolii. Floristiquement si l’on suit les taxons différentiels de cette classe, ces associations contiennent les caractéristiques ou différentielles de la classe : Cistus salviifolius, Cytinus hypocistis (rare), Daphne gnidium et les différentielles des Lavanduletalia stoechadis et du Cistion laurifolii. Helichrysum stoechas, assez rare dans Botineau et al. (1988), est apparue plus fréquente dans les relevés effectués lors de cette étude. Chorologiquement, ces végétations sont positionnées dans des secteurs possédant une flore et des végétations méditerranéennes (Quercion ilicis).
Le rattachement de ces végétations à cette classe euryméditerranéenne est faite par défaut car la proximité floristique reste assez faible mais aucune autre classe ne semble pouvoir les intégrer. Ces végétations peuvent alors être considérées comme des associations d’appauvrissement floristique en limite chorologique (de Foucault, 1981) des Cisto-Lavanduletea stoechadis.
Calluno vulgaris-Ulicetea minoris Braun-Blanq. & Tüxen ex Klika in Klika & Hadač 1944
Ulicetalia minoris Quantin 1935
Cisto salviifolii-Ericion cinereae Géhu ex Bardat, Bioret, Botineau, Boullet, Delpech, Géhu, Haury, Lacoste, Rameau, J.-M. Royer, G. Roux & Touffet 2004
- Cisto salviifolii-Ericetum cinereae Guitton, Juhel & Julve 2018
- ulicetosum Botineau, Bouzillé & Lahondère 1988
- Festuco vasconcensis-Ericetum cinereae Géhu & Géhu-Franck 1975 corr. Glemarec, Delassus & Boullet
- typicum (= loniceretosum Géhu & Géhu-Franck 1975)
- ericetosum scopariae Géhu & Géhu-Franck 1975
7. Conclusion
Cette étude, basée sur une importante analyse bibliographique et sur la réalisation systématique de relevés phytosociologiques par maille sur l’ensemble du littoral aquitain, fournit une synthèse complète et actualisée sur les végétations des sables littoraux d’Aquitaine après celles de Géhu (1982, 1995), Géhu & de Foucault (1982) pour ne citer que les plus importantes. Elle aura permis de mettre encore en évidence cinq nouvelles associations, mettant ainsi une nouvelle fois en avant la richesse et l’originalité des végétations dunaires d’Aquitaine.
La méthode de prospection utilisée a permis d’actualiser la répartition des végétations littorales dunaires sur un vaste territoire, du nord au sud et latéralement dans la dune. Elle offre ainsi un état complet des végétations des dunes non boisées d’Aquitaine en 2015 qui permettra à l’avenir des comparaisons évolutives qualitatives (combinaisons de taxons) et quantitatives (synchorologie).
Ces connaissances typologiques, synchorologiques et écologiques synthétiques doivent maintenant être transmises aux propriétaires et gestionnaires de dunes afin que la gestion conservatoire de ces végétations hautement patrimoniales soit le plus possible adaptée à chaque communauté et à chaque localité.
Toutes ces informations sont consultables via la base de données en ligne de l’Observatoire de la biodiversité végétale (OBV) de Nouvelle-Aquitaine (www.obv-na.fr).
Bibliographie
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Collection Patrimoines naturels 61 : 1-171.
Botineau M., Bouzillé J.-B. & Lahondère Ch., 1988. Sur la présence d’un ourlet méditerranéo-atlantique dans le Centre-Ouest : le Rubio peregrinae-Cistetum salviifoliae ass. nov. Bulletin de la Société Botanique du Centre-Ouest, NS, 19 : 101-104.
Braun-Blanquet J. & Tüxen R., 1952. Irische Pflanzengesellschaften. Veröffentlichungen des Geobotanischen Institutes Rübel in Zürich 25 : 224-421.
Catteau E., Buchet J., Camart Ch., Coulombel R., Dardillac A., Delplanque S., Duhamel F., François R., Hauguel J.-C., Prey T. & Villejoubert G., 2021. Végétation du nord de la France, guide de détermination. Éditions Biotope, Mèze, 400 p.
Dufay J., 2014. Amélioration des connaissances sur la flore rare et menacée et les habitats naturels du littoral sud-landais. Conservatoire botanique national Sud-Atlantique, Audenge, 146 p.
Esteve G., 1980. Les dunes : leur formation et leur évolution. Bulletin de la Société Botanique du Centre-Ouest, NS, n° sp. 4 (La vie dans les dunes du Centre-Ouest) : 3-15.
Favennec J., 1997. Gestion conservatoire des dunes littorales non boisées. In Biodiversité et protection dunaire. Lavoisier, Tec et Doc : 165-171.
Foucault (de) B., 1981. Réflexions sur l’appauvrissement des syntaxons aux limites chorologiques des unités phytosociologiques supérieures et quelques-unes de leurs conséquences. Lazaroa 3 : 75-100.
Foucault (de) B., 2021. Contribution à une synthèse des classes des Honckenyo peploidis-Leymetea arenarii et des Euphorbio paraliae-Ammophiletea arundinaceae. Carnets botaniques 64: 1-22.
Foucault (de) B., Argagnon O. & Paradis G., 2012. Contribution au prodrome des végétations de France : les Cisto ladaniferi – Lavanduletea stoechadis Braun-Blanq. in Braun-Blanq., Molin. & Wagner 1940. Journal de botanique de la Société botanique de France 57 : 59-82.
Gargominy O., Tercerie S., Régnier C., Dupont P., Daszkiewicz P., Léotard G., Antonetti Ph., Ramage T., Vandel E., Petitteville M., Leblond S., Idczak L., Boullet V., Denys G., De Massary J.-C., Lévêque A., Jourdan H., Rome Q., Dusoulier F., Touroult J., Savouré-Soubelet A., Barbut J., Canard A., Simian G., Le Divelec R., Haffner P., Meyer C., Van Es J., Poncet R., Demerges D., Mehran B., Horellou A., Moulin N., Ah-Peng C., Bernard J.-F., Caesar M., Comolet-Tirman J., Courtecuisse R., Delfosse E., Dewynter M., Hugonnot V., Kondratyeva A., Lavocat Bernard E., Lebouvier M., Lebreton E., Malécot V., Moreau P.A., Muller S., Noblecourt T., Pellens R., Robbert Gradstein S., Rodrigues, C., Rouhan G. & Véron S., 2020. TAXREF v14.0, référentiel taxonomique pour la France. UMS PatriNat, Muséum national d’histoire naturelle, Paris. Archive de téléchargement contenant 8 fichiers, https://inpn.mnhn.fr/telechargement/referentielEspece/taxref/14.0/menu
Géhu J.-M., 1964. La végétation psammophile des îles de Houat et Hoëdic. Bulletin de la Société de botanique du nord de la France 17 (4) : 238-266.
Géhu J.-M., 1968. Sur la vicariance géographique des associations végétales des dunes mobiles de la côte atlantique française. Compte rendu hebdomadaire des séances de l’Académie des sciences, série D, 266 : 2422-2425.
Géhu J.-M., 1969. Application en phytosociologie de la cartographie en réseau. Bulletin de la Société de botanique du nord de la France 22 : 1-25.
Géhu J.-M., 1975. Essai systématique et chorologique sur les principales associations végétales du littoral atlantique français. Anales de la Real Academia de Farmacia 41 : 207-227.
Géhu J.-M., 1978. Les phytocoenoses endémiques des côtes françaises occidentales. Bulletin de la Société botanique de France 125 : 199-208.
Géhu J.-M., 1982. La végétation des plages de sable et des dunes des côtes françaises (aperçu synthétique). Université de Paris V, 60 p.
Géhu J.-M., 1994. Schéma synsystématique et typologie des milieux littoraux français atlantiques et méditerranéens. Colloques phytosociologiques XXII : 183-212.
Géhu J.-M., 1995. Inventaire typologique des milieux terrestres littoraux, 2. Centre régional de phytosociologie/Conservatoire botanique national de Bailleul, 223 p.
Géhu J.-M., 2008. Sur l’association à Crithmum maritimum et Otanthus maritimus de Pavillard 1928. Bulletin de la Société botanique du Centre-Ouest, NS, 39 : 163-168.
Géhu J.-M. & Géhu J., 1969. Les associations végétales des dunes mobiles et des bordures de plages de la côte atlantique française. Vegetatio 18 : 122-166.
Géhu J.-M. & Tüxen R., 1972. Essai de synthèse phytosociologique des dunes atlantiques européennes. Colloques phytosociologiques I : 61-70.
Géhu J.-M. & Géhu J., 1975. Les fourrés des sables littoraux du Sud-Ouest de la France. Beiträge zur naturkundlichen Forschung in Südwestdeutschland, 34 (Festschrift zum 70 Geburststag von Erich Oberdorfer) : 79-94.
Géhu J.-M. & Géhu-Franck J., 1975. Contribution à l’étude phytosociologique des landes du sud-ouest de la France. Colloques phytosociologiques II : 75-87.
Géhu J.-M. & Rivas-Martínez S., 1981. Notions fondamentales de phytosociologie. In H. Dierschke (ed.), Berichte über das Internationale Symposium Der Internationalen Vereinigung fur Vegetations, Syntaxonomie, Rinteln 1980 : 5-33.
Géhu J.-M. & de Foucault B., 1982. Analyse phytosociologique et essai de chorologie intégrée de l’hygrosère des dunes atlantiques françaises. Documents phytosociologiques, NS, VII : 387-398.
Géhu J.-M. & Géhu-Franck J., 1988. Variations floristiques et synchorologie des ammophilaies européo-africaines. Homenaje a P. Montserrat, Jaca y Huesca, 561-570.
Géhu J.-M., Franck J. & Bournique C.-P., 1995. Sur l’originalité syntaxonomique des ammophilaies du sud-ouest de la France (Sileno thorei–Ammophiletum arenariae) et leur positionnement dans le synsystème européen des végétations des dunes meubles. Bulletin de la Société botanique du Centre-Ouest, NS, 26 : 99-104.
Guinochet M., 1973. Phytosociologie. Masson, Paris, 227 p.
Guitton H., Juhel C. & Julve Ph., 2017. Une nouvelle association de lande thermo-atlantique à Cistus salviifolius et Erica cinerea en France. Colloques phytosociologiques VI : 203-211.
Jeandenand F. & Blanchard F., 2009. Bilan floristique et diagnostic phytoécologique des dunes d’Anglet (Pyrénées-Atlantiques). Conservatoire botanique national Sud-Atlantique, Conseil général des Pyrénées-Atlantiques, 26 p + annexes.
Jun R., 2005. Les mousses et lichens des dunes grises atlantiques : caractéristiques structurales, dynamique et typologie fonctionnelle des communautés. Ecology,environment. Université de Rennes 1, 168 p.
Kopecký K. & Hejný S.,1974. A new approach to the classification of anthropogenic plant communities. Vegetatio 29 :17-20.
Lafon P., Le Fouler A., Dufay J. & Hardy F., 2015. Les végétations des dunes littorales non boisées d’Aquitaine : synsystématique et synchorologie (Euphorbio paraliae-Ammophiletea australis, Koelerio-Corynephoretea canescentis p .p., Scheuchzerio palustris-Caricetea fuscae p. p.). Conservatoire botanique national Sud-Atlantique, DREAL Aquitaine, 119 p. + annexes.
Lafon P. (coord.), Mady M., Corriol G. & Belaud A., 2020. Catalogue des végétations de Nouvelle-Aquitaine. Classification, chorologie et correspondances avec les habitats européens. Conservatoire botanique national Sud-Atlantique / Conservatoire botanique national Massif central / Conservatoire botanique national des Pyrénées et de Midi-Pyrénées, 272 p., https://obv-na.fr/ofsa/ressources/7_habitats/CBN_2020-Catalogue_des_vegetations_de_Nouvelle-Aquitaine_v1.0.pdf
Lahondère Ch., 1980a. La flore et la végétation phanérogamiques. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 4 (La vie dans les dunes du Centre-Ouest) : 113-171.
Lahondère Ch., 1980b. La végétation des sables dunaires du littoral du golfe de Gascogne entre Capbreton et Hendaye. Bulletin de la Société botanique du Centre-Ouest, NS, 10 : 235-246.
Lahondère Ch., 1980c. Les dunes du Centre-Ouest : le milieu physique. Bulletin de la Société botanique du Centre-Ouest, NS, n° sp. 4 (La vie dans les dunes du Centre-Ouest) : 17-24.
Lazare J.-J., 2017. Contribution à l’étude phytosociologique et symphytosociologique du littoral aquitain (France). Botanique 2 : 41-71.
Marcenò C., Guarino R., Loidi J., Herrera M., Isermann M., Knollová I., Tichý L., Tzonev R. Acosta A., Fitzpatrick Ú., Iakushenko D., Janssen J.A.M.; Jiménez-Alfaro B., Kacki Z., Keizer-Sedláková I., Kolomiychuk V., Rodwell J., Schaminée J.H.J., Šilc U. & Chytrý M., 2018. Classification of European and Mediterranean coastal dune vegetation. Applied Vegetation Science 21 (3) : 533-559.
Mucina L., Bültmann H., Dierben K., Theurillat J.-P., Raus Th., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Y.P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S.M. et Tichý L., 2016. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 19 (1): 3-264.
Pavillard J., 1928. Le Crithmion maritimae autour de Biarritz. Bulletin de la Société botanique de France 75 : 795-799.
Rolecek J., Tichý L., Zeleny D. & Chytrý M., 2009. Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity. Journal of vegetation science 20 : 596-602.
Theurillat J.-P., Willner W., Fernández-González F., Bültmann H., Čarni A., Gigante D., Mucina L. & Weber H., 2020. International Code of Phytosociological Nomenclature, 4th edition. Applied Vegetation Science 24 (2) : 1-62.
Tison J.-M. & de Foucault B. (coord.), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Vanden Berghen C., 1963. L’association à Helichrysum stoechas dans les dunes du littoral du sud-ouest de la France (Roseto-Ephedretum Kuhnholz-Lordat). Vegetatio 11 : 317-324.
Vanden Berghen C., 1964a. Notes sur la végétation du sud-ouest de la France, I – La végétation herbacée des dunes maritimes fixées. Bulletin du Jardin botanique de l’État 35 (4) : 355-362.
Vanden Berghen C., 1964b. Notes sur la végétation du sud-ouest de la France, I – La végétation des dunes mobiles. Bulletin du Jardin botanique de l’État 34 : 519-525.
Vanden Berghen C., 1971. Notes sur la végétation du sud-ouest de la France, VIII – Les fourrés et les bois fangeux. Bulletin du Jardin botanique national de Belgique 41 : 383-395.
Westhoff, V., Dijk J.W., Passchier H. & Sissingh G., 1946. Overzicht der plantengemeenschappen in Nederland (2e druk). Uitg. KNNV, NJN Breughel, Amsterdam, 118 p.
Remerciements
Nous remercions la DREAL Aquitaine, puis Nouvelle-Aquitaine pour avoir soutenu cette étude et les programmes phytosociologiques régionaux. Nous tenons à remercier Pablo Massart (CBNSA) et Kevin Romeyer (CBNSA) pour leur participation à la phase de terrain. Merci également à toutes les personnes qui nous ont accompagnés sur le terrain : Gilles Granereau, David Rosebery, Émilie Sautret, Paul Tourneur de l’Office national des forêts, Didier Alard, Marie-Lise Bénot, Edith Reuzeau, Béatrice Sauzeau de l’UMR 1202 INRA – Biogeco de l’Université de Bordeaux et Vanessa Rispal de la DREAL Nouvelle-Aquitaine. Enfin, nous remercions vivement les personnes qui ont pris le temps de relire cet article et sa première version et de contribuer à son amélioration : Bruno de Foucault, Mickael Mady et Pierre-Marie Le Hénaff (CBN du Massif central), Erwan Glemarec et Hermann Guitton (CBN de Brest), Emmanuel Catteau (CBN de Bailleul), Pauline Delbosc (CBN Sud-Atlantique) et Laura Corredor. Ces travaux ont été menés sous la coordination de Grégory Caze, directeur scientifique du CBNSA, que nous remercions par ailleurs pour sa relecture. Le contenu de cet article a fait l’objet d’une présentation lors du septième symposium de la Société française de phytosociologie, colloque en hommage à Jean-Marie Géhu, Saint-Brieuc du 4-7 octobre 2016. Cet article rédigé et soumis il y a cinq ans est resté inédit, il a été donc été actualisé avec les publications parues entretemps et les nouvelles connaissances accumulées durant cette période.


