
Compléments aux synthèses des Thero-Suaedetea splendentis, Sarcocornietea fruticosae et Juncetea maritimi
Title
Supplements to the overviews of the Thero-Suaedetea splendentis, Sarcocornietea fruticosae and Juncetea maritimi
Résumé
À la suite de synthèses récentes des Thero-Suaedetea splendentis, Sarcocornietea fruticosae et Juncetea maritimi, cet article apporte des compléments synsystématiques pour ces végétations halophiles vers la Roumanie, la Turquie, l’Ukraine et la Russie. Deux ordres et sept alliances sont nouvellement décrits ou validés.
Abstract
Following recent overviews of the Thero-Suaedetea splendentis, Sarcocornietea fruticosae and Juncetea maritimi, this article provides additional syntaxonomical information for the halophytic vegetations on Romania, Turkey, Ukraine and Russia. Two orders and seven alliances are newly described or validated.
Dans les synthèses récentes des Sarcocornietea fruticosae, Juncetea maritimi et Thero-Suaedetea splendentis (de Foucault, 2021a, 2021b ; de Foucault & Guitton, 2021), plusieurs syntaxons décrits de Roumanie, d’Anatolie et de Russie n’avaient pas été pris en compte. Leur originalité vis-à-vis des syntaxons ouest-européens justifie qu’ils soient considérés et examinés sous l’angle de la phytosociologie moderne.
La nomenclature suit Euro+Med PlantBase, et Catalogue of Life pour les taxons absents du premier référentiel ; dans les tableaux, le signe * remplace ‘subsp.’ ou ‘var.’ ; les sous-espèces autonymes ont leur épithète subspécifique réduite à leur initiale. Les types biologiques des genres d’Amaranthaceae absents d’Europe occidentale ont été vérifiés dans la monographie de Townsend (1993).
1. Les prés halophiles thérophytiques
1.1. Les prés halophiles thérophytiques de Roumanie
Il faut d’abord compléter le synsystème ouest- et centre-européen avec les données suivantes de Ţopa (1939) synthétisées dans le tableau 1 :
- Camphorosmetum annuae Ţopa 1939 (Ţopa, 1939, tableau p. 39).
- Composante thérophytique de l’Obionetum verruciferae Ţopa 1939 (Ţopa, 1939, tableau p. 45).
- Composante thérophytique du Camphorosmetum monspeliacae Ţopa 1939 (Ţopa, 1939, tableau p. 47, sub Camphorosmetum pilosae).
- Composante thérophytique de l’Halocnemetum strobilacei Ţopa 1939 (Ţopa, 1939, tableau p. 49).
- Crypsietum schoenoidis sensu Ţopa 1939 (Ţopa, 1939, tableau p. 51).
- Suaedo maritimae-Kochietum hirsutae sensu Ţopa 1939 (Ţopa, 1939, tableau p. 53).
Les colonnes 1 à 3 semblent décrire un même Camphorosmetum annuae Ţopa 1939, plus fragmentaire dans la colonne 3, ce nom est toutefois un homonyme illégitime du Camphorosmetum annuae Rapaics ex Soó 1933 à en croire le tableau 9 in Vicherek (1973, sub Camphorosmetum ovatae), association à Matricaria chamomilla, Plantago tenuiflora, Cerastium dubium ; on peut le renommer Suaedo maritimae–Camphorosmetum annuae (Ţopa 1939) nom. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 5 du tableau in Ţopa (1939, Bul. Făc. Şti. Cernăuţi XIII : 39). Le Camphorosmetum annuae décrit de Slovaquie par Dítĕ et al. (2008) est en fait la composante thérophytique d‘un pré halophile vivace à Puccinellia distans (V sur les 32 relevés des tableaux 1 et 2 des auteurs), Artemisia santonicum subsp. patens (V), Plantago maritima subsp. m. (IV), Festuca valesiaca subsp. parviflora (III), Limonium gmelinii (II), Tripolium pannonicum (II), Achillea collina (I), Cynodon dactylon (I)…, qui correspond sans doute à une association originale du Festucion pseudovinae Soó in Máthé 1933 à muter en Festucion parviflorae nom. mut. hoc loco.
La colonne 4 est basée sur seulement quatre relevés et deux taxons, alors que la colonne 5 décrit un syntaxon déjà placé dans les Crypsietea aculeatae (de Foucault, 2021c). Quant au syntaxon 6, que son auteur rapporte au Suaedo maritimae-Kochietum hirsutae Braun-Blanq. 1933 ouest-méditerranéen muté en Suaedo maritimae-Spirobassietum hirsutae par de Foucault & Guitton (2021), la comparaison montre que cette identification n’est pas tenable : ici, Halimione pedunculata et Salicornia europaea sont significativement présents, alors que manquent Salicornia procumbens subsp. p., S. perennans subsp. p. et Salsola soda ; une association originale vicariante peut être définie sous le nom de Halimiono pedunculatae-Spirobassietum hirsutae (Ţopa 1939) nom. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 10 du tableau in Ţopa (1939, Bul. Făc. Şti. Cernăuţi XIII : 53).
1.2. Des prés halophiles thérophytiques anatoliens
Le tableau 2 synthétise les données sur la végétation halophile thérophytique anatolienne publiées par Yurdakulol et al. (1996) puis Hamzaoğlu & Aksoy (2006, 2009).
- Composante thérophytique de l’Halocnemetum strobilacei Ţopa 1939 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 1).
- Composante thérophytique de l’Inulo aucherianae-Elymetum salsi Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 7).
- Composante thérophytique du Lepidio caespitosi-Limonietum iconici Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 2).
- Composante thérophytique du Limonio tamaricoidis-Puccinellietum convolutae Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 3).
- Composante thérophytique du Sphenopodo divaricati-Halocnemetum strobilacei Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 4).
- Suaedo prostratae-Salsoletum nitrariae Aydoğdu, Hamzaoğlu & Kurt 2002 nom. mut. hoc loco (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 5, sub Suaedo anatolicae-Salsoletum nitrariae).
- Composante thérophytique du Lepidio caespitosi-Limonietum iconici Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2006, tableau 3 : relevés 359 à 390) ; réapparaît sous des fomes plus fragmentaires dans les Halocnemetum strobilacei et Inulo aucherianae-Elymetum salsi de la même référence.
- Composante thérophytique du Bupleuro gracilis-Limonietum gmelinii Yurdakulol ex Hamzaoğlu in Hamzaoğlu & Aksoy 2009 (‘…gracili-…‘) (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 6), nom invalide selon l’article 7 de l’ICPN.
- Petrosimonietum nigdeensi-brachiatae Yurdakulol, Öncel, Demirörs, Yildiz & Keles 1996 (Yurdakulol et al., 1996, tableau 5, ‘… nigdeenso-…’).
- Composante thérophytique du Tamaricetum parvifloro-tetrandrae Hamzaoğlu & Aksoy 2006 (Hamzaoğlu & Aksoy, 2006, tableau 3 : relevés 370 à 389).
- Composante thérophytique de l’Eragrostio collinae-Puccinellietum anatolicae Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 7), surtout à Linum seljukorum, Blackstonia perfoliata et Polypogon monspeliensis.
On laissera tout d’abord de côté les colonnes 10, trop fragmentaire, et 11, assez originale mais ne relevant sans doute pas des Thero-Suaedetea splendentis (description à approfondir). Les colonnes 1 et 2 relèvent bien des Thero-Suaedetea splendentis mais correspondent à une communauté très basale réduite à Salicornia europaea. Les colonnes 3 et 4 semblent correspondre à un même Salicornietum centre-anatolien à Salsola tetraptera : Salsolo tetrapterae-Salicornietum europaeae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 22 du tableau 2 in Aydoğdu, Hamzaoğlu & Kurt (2002, Israel J. Pl. Sci. 50 : 316). La colonne 5 corespond à une association originale à Microcnemum coralloides à séparer d’un Halocnemetum strobilacei local, par ailleurs bien différente du Microcnemetum coralloidis Rivas Mart. in Rivas Mart. & M.J. Costa 1976 ibérique : Sphenopodo divaricati-Microcnemetum coralloidis ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 4 du tableau 2 in Aydoğdu, Hamzaoğlu & Kurt (2002, Israel J. Pl. Sci. 50 : 316). La colonne 7 peut être distinguée sous le nom de Salsolo stenopterae-Petrosimonietum nigdeensis ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 359 du tableau 3 in Hamzaoğlu & Aksoy (2006, Ekoloji 15 (60) : 12). La colonne 8 rappelle le Bupleuro gracilis-Petrosimonietum oppositifoliae Oberd. 1952 nord-égéen (de Foucault & Guitton, 2021, syntaxon TSs8) ; on pourrait le distinguer sous le nom de Petrosimonio brachiatae-Bupleuretum gracilis mais aucun holotype publié ne paraît disponible pour valider ce nom.
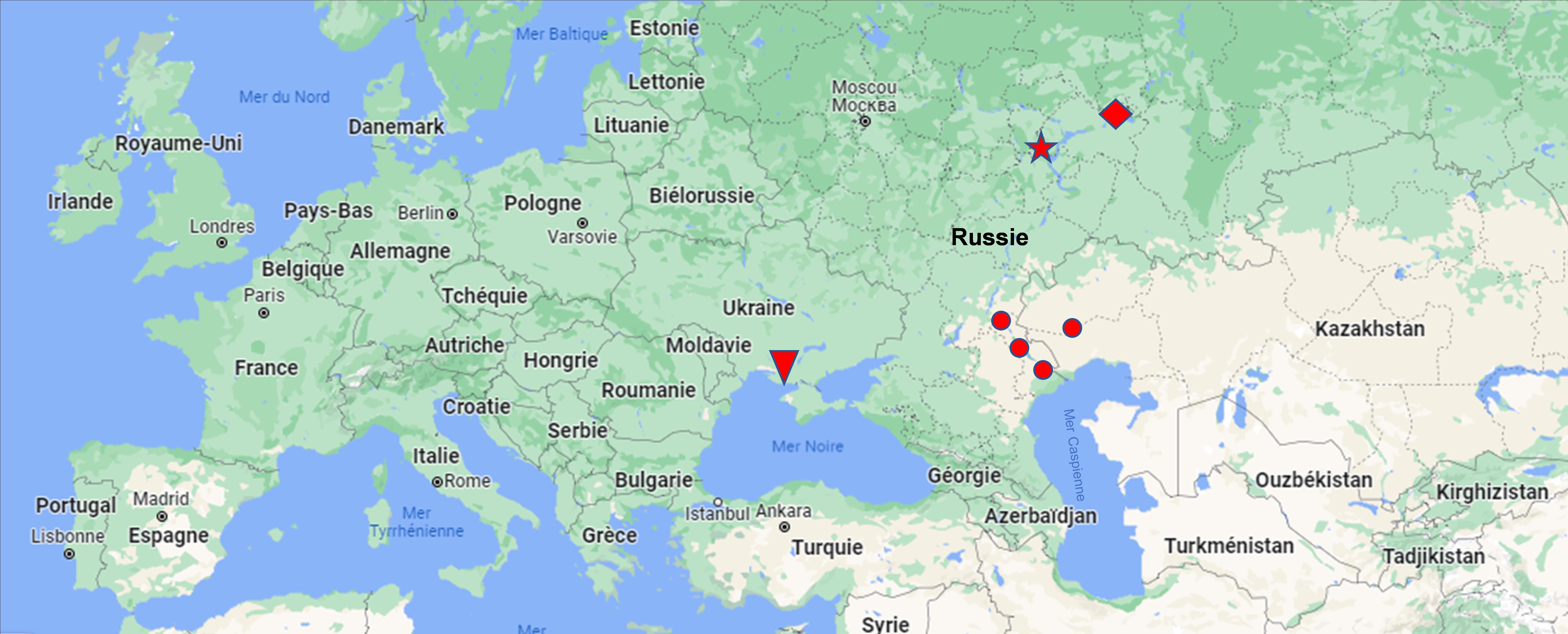
On peut rapprocher les syntaxons correspondant aux colonnes 3 à 9 dans un même ensemble centre-anatolien que l’on peut au moins provisoirement rattacher aux Thero-Suaedetalia splendentis mais qui ne correspond à aucune des sept alliances déjà connues (de Foucault & Guitton, 2021) ; on peut donc définir ici le Salsolo stenopterae-Petrosimonion brachiatae all. nov. hoc loco, rassemblant des prés halophiles thérophytiques colonisant les rives de lacs centre-anatoliens (carte 1), alliance caractérisée par Petrosimonia brachiata, P. nigdeensis, Thesium compressum, Salsola stenoptera, S. inermis, Eremopyrum orientale, typus nominis hoc loco : Suaedo anatolicae-Salsoletum nitrariae Aydoğdu, Hamzaoğlu & Kurt 2002 (Israel J. Pl. Sci. 50 : 315), muté ici en Suaedo prostratae-Salsoletum nitrariae.
À cette alliance on pourrait encore ajouter les composantes thérophytiques des Artemisietum santonici, Cardopatio corymbosi-Juncetum orientalis, Lepidio caespitosi-Halimionetum portulacoidis et Cresso creticae-Halocnemetum strobilacei décrits par Yurdakulol et al. (1996, tableaux 2, 3, 6, 7) pas toujours bien caractérisées.

1.3. Les prés halophiles thérophytiques de Russie et d’Ukraine méridionales
Le tableau 3 synthétise les données sur la végétation halophile thérophytique du sud de la Russie (basse vallée de la Volga, nord de la mer Caspienne) publiées par Golub & Mirkin (1986), Golub & Čorbadze (1989), Freitag et al. (2001).
- Composante thérophytique du Suaedo salsae-Halocnemetum strobilacei V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 2, en changeant Suaeda salsa en S. acuminata selon Lysenko & Mucina, 2015).
- Composante thérophytique du Limonietum suffruticosi V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 3, en changeant Suaeda salsa en S. acuminata).
- Composante thérophytique du Kalidietum foliati V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 4, en changeant Suaeda salsa en S. acuminata).
- Composante thérophytique du Suaedo salsae-Frankenietum hirsutae V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 5, en changeant Suaeda salsa en S. acuminata).
- Suaedo acuminatae-Petrosimonietum oppositifoliae V.B. Golub 1986 nom. mut. hoc loco (Golub & Čorbadze, 1989, tableau 6, sub Suaedo salsae-Petrosimonietum oppositifoliae), homonyme du syntaxon 8 ci-après et pourtant globalement assez distinct ; signé du seul nom de Golub, alors que la publication originale est signée de Golub & Mirkin (cf. syntaxon 8).
- Groupement à Petrosimonia oppositifolia-Suaeda acuminata (Freitag et al., 2001, tableau 10 : relevés 1 à 4).
- Groupement à Suaeda altissima-Suaeda acuminata (Freitag et al., 2001, tableau 10 : relevés 5 à 8).
- Suaedo acuminatae-Petrosimonietum oppositifoliae V.B. Golub & Mirkin 1986 (Golub & Mirkin, 1986, tableau 11).
- Groupement à Salsola tragus-Suaeda acuminata (Freitag et al., 2001, tableau 10 : relevés 9 à 13).
Les syntaxons d’origine sont assez souvent biologiquement hétérogènes, les seuls réellement thérophytiques étant les syntaxons 5 à 9. On peut considérer qu’ils relèvent du Climacoptero crassae-Suaedion acuminatae V.B. Golub & Čorbadze 1989 (Folia Geobot. Phytotax. 24 : 120) corr. Lysenko & Mucina 2015 (Hacquetia 14 (2) : 304) (Golub & Čorbadze, 1989 ; Lysenko & Mucina, 2015), dont la chorologie est reprise en carte 2.
Le tableau 4 synthétise des données sur les environs de la mer Caspienne (Golub & Mirkin, 1986 ; Golub & Čorbadze, 1989 ; Freitag et al., 2001) et de l’Ukraine méridionale (Dubyna & Neuhäuslová, 2003 ; Dubyna et al., 2020).
- Salicornietum prostratae sensu Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 1), probablement une forme fragmentaire du suivant.
- Salicornio perennantis-Suaedetum salsae Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 2).
- Suaedetum eltonicae Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 3).
- Groupement à Suaeda acuminata-Salicornia perennans (Freitag et al., 2001, tableau 4).
- Salsoletum acutifoliae Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 6).
- Groupement à Bassia hyssopifolia-Suaeda salsa (Freitag et al., 2001, tableau 7).
- Ofaisto monandri-Salicornietum perennantis Dubyna & Neuhäuslová 2003 (Dubyna & Neuhäuslová, 2003, tableau 4).
- Salicornietum prostratae Soó 1927 (Dubyna & Neuhäuslová, 2003, tableau 1), à réinterpréter car l’association des auteurs ukrainiens est sans doute distincte de celle de Soó; elle semble d’ailleurs à inclure dans l’association suivante.
- Suaedetum salsae V.B. Golub & Čorbadze 1995 (Dubyna & Neuhäuslová, 2003, tableau 6) ; on peut aussi y rattacher la communauté thérophytique du Limonio caspii-Salicornietum prostratae Koržhenĕvskij & Kljukin 1991 (Dubyna & Neuhäuslová, 2003, tableau 2), ainsi que le tableau 1 in Grechushkina et al. (2010, sub Salicornietum prostratae et Salicornio perennantis-Suaedetum salsae).
- Petrosimonio oppositifoliae-Salicornietum perennantis Koržhenĕvskij & Kljukin 1991 (Dubyna & Neuhäuslová, 2003, tableau 3).
- Halimionetum pedunculatae Şerbănescu 1965 (Dubyna et al., 2020, tableau 4 : colonne 5).
- Suaedetum salso-prostratae Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 5).
- Salicornio perennantis-Suaedetum salsae V.B. Golub & Čorbadze 1995 (Dubyna et al., 2020, tableau 4 : colonne 6).
- Suaedetum acuminatae V.B. Golub & Čorbadze in V.B. Golub 1995 corr. Lysenko & Mucina 2015 (Dubyna et al., 2020, tableau 4 : colonne 8, sub Suaedetum confusae).
- Composante thérophytique du Petrosimonio brachiatae-Artemisietum santonici Koržhenĕvskij & Kljukin 1991 (Dubyna et al., 2020, tableau 4 : colonne 10), à reconsidérer sous le nom de Atriplici calothecae-Petrosimonietum brachiatae ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 2 du tableau 2 in Koržhenĕvskij & Kljukin (1991, Feddes Repert. 102 (1-2) : 141).
Plusieurs des auteurs placent ces syntaxons dans le Salicornion prostratae Géhu 1992, synonyme du Salicornion patulae Géhu & Géhu-Franck 1984 muté en Salicornion perennantis par de Foucault & Guitton (2021). En comparant les colonnes Spe et Sa du tableau synthétique 6, on voit qu’effectivement ces deux syntaxons partagent Salicornia perennans, Salsola soda, parfois Spirobassia hirsuta, Bassia hyssopifolia, mais bien d’autres taxons les différencient (Suaeda maritima et Atriplex prostrata côté Spe, Suaeda acuminata, S. salsa, S. eltonica et Salsola acutifolia côté Sa), ce qui justifie la distinction d’une alliance originale, qui doit correspondre au Suaedion salsae V.B. Golub & Čorbadze in V.B. Golub 1995 (Halophytic, desert and semidesert plant communities… : 1) corrigé en Suaedion acuminatae par Lysenko & Mucina (2015), partiellement sympatrique du Climacoptero-Suaedion acuminatae (carte 2) quoique floristiquement bien différencié.
Le tableau 5 synthétise des données de Russie centro-occidentale (Freitag et al., 2001 ; Karpov et al., 2003 ; Lysenko et al., 2003).
- Suaedetum corniculatae sensu Freitag, V.B. Golub & Yuritsyna 2001 (Freitag et al., 2001, tableau 8) ; voir ci-dessous syntaxon 4.
- Groupement basal à Suaeda corniculata (Karpov et al., 2003, tableau 2 : relevés 6 à 14, tableau 3 : relevés 1 à 10) ; peut-être à rattacher au précédent.
- Composante thérophytique de l‘Atriplici laevis-Elymetum repentis V.B. Golub, Rukhlenko & Sokolov 2001 (Lysenko et al., 2003, tableau 3), qui peut être distinguée sous le nom de Atriplici laevis-Camphorosmetum songoricae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 39 du tableau 3 in Lysenko et al. (2003, Vegetation of Russia 4 : 45).
Ces syntaxons peuvent se placer dans le Camphorosmo songoricae-Suaedion corniculatae Freitag, V.B. Golub & Yuritsyna 2001 (Phytocoenologia 31 (1) : 103), typifié par le Suaedo corniculatae-Camphorosmetum songoricae Korolyuk 1999 qu’il n’a pas été possible de consulter pour vérifier la typification. La carte 2 précise la chorologie connue de cette alliance.
À droite de ce tableau 5, on a aussi inséré des données sur des communautés thérophytiques à Suaeda corniculata de la vallée de la Lena, au nord et au nord-est du lac Baïkal, en Sibérie :
- Suaedetum corniculatae Burtseva in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992 (Mirkin et al., 1992, tableau 17) ; il nous semble que ce syntaxon sibérien est distinct du Suaedetum corniculatae au sens de Freitag et al. (2001, cf. supra, syntaxon 1), qui doit être alors renommé : Suaedetum salso-corniculatae (Freitag, V.B. Golub & Yuritsyna 2001) nom. nov. hoc loco, typus nominis hoc loco: composante thérophytique du relevé 2 du tableau 8 in Freitag et al. (2001, Phytocoenologia 31 (1) : 89).
- Composante thérophytique du Suaedo corniculatae-Puccinellietum tenuiflorae Mirkin, Maniazar, Alimbekova & Oniščenko 1986 (Mirkin et al., 1992, tableau 18).
- Composante thérophytique de l‘Artemisio jacuticae-Suaedetum corniculatae Burtseva in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992 (Mirkin et al., 1992, tableau 19).
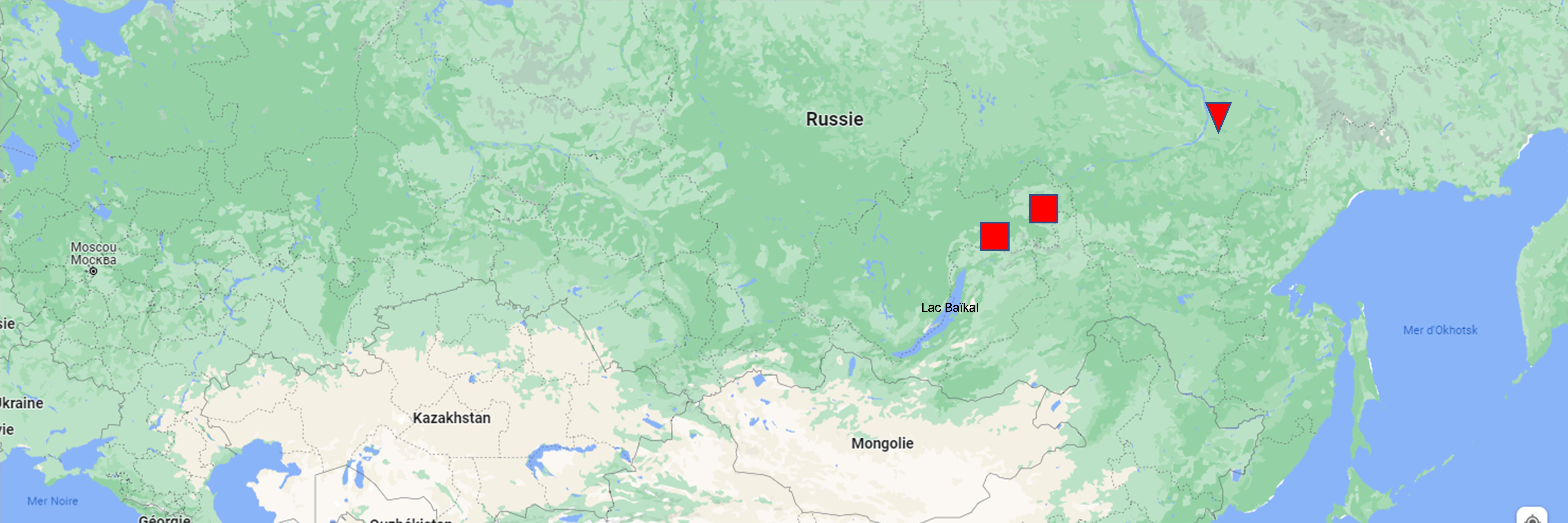
Ces deux derniers syntaxons se réduisent à des formes, dont l’une notable à Eutrema salsugineum, du Suaedetum corniculatae Burtseva in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992. Les auteurs russes rangent deux de ces trois syntaxons dans le Suaedo corniculatae-Puccinellion tenuiflorae Mirkin in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992 nom. inval. car non typifié, qu’on ne peut d’ailleurs conserver selon nos choix méthodologiques ; on peut plutôt définir le Salicornio europaeae-Suaedion corniculatae all. nov. hoc loco, typus nominis hoc loco : Suaedetum corniculatae Burtseva in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992 (Folia Geobot. Phytotax. 27 (3) : 290) (carte 3).
1.4. Synthèse actualisée des Thero-Suaedetea splendentis
Avec ces nouvelles données, une synthèse actualisée des Thero-Suaedetea splendentis doit être tentée ; elle sera basée sur le tableau 1 publié par de Foucault & Guitton (2021), dans lequel on injectera les colonnes synthétiques des tableaux 1 à 5 pour structurer le tableau 6. Au sein des Thero-Suaedetalia splendentis, les alliances déjà considérés en 2021 sont les Salicornion procumbentis (colonne Spr du tableau 6), Salicornion disarticulato-europaeae (Sde), Salicornion perennantis (Spe), Thero-Suaedion splendentis (TSs), Microcnemion coralloidis (Mc), Salicornion grandtneri (Sg) et Atriplici gmelinii-Suaedion japonicae (ASj).
La colonne CSm (synthèse du tableau 1) correspond à une alliance originale au sein de cet ensemble, le Camphorosmo annuae-Suaedion maritimae all. nov. hoc loco, caractérisée surtout par Camphorosma annua, Petrosimonia triandra et Plantago tenuiflora en plus des taxons caractéristiques de classe, typus nominis hoc loco : Suaedo maritimae-Camphorosmetum annuae (Ţopa 1939) nov. La colonne SPb synthétise le Salsolo tetrapterae-Petrosimonion brachiatae déjà défini et validé (tableau 2). La colonne SSc synthétise le Salicornio europaeae-Suaedion corniculatae sibérien (tableau 5). On peut rattacher ces trois alliances aux Thero-Suaedetalia splendentis, le Salsolo-Petrosimonion brachiatae et le Salicornio-Suaedion corniculatae plutôt en limite d’ordre et de classe.
En revanche les colonnes CSa, CSc et Sa paraissent ne plus se rattacher aux Thero-Suaedetalia splendentis ; il faut au moins définir un nouvel ordre pour ces syntaxons : Suaedetalia corniculatae V.B. Golub ex ord. nov. hoc loco [syn. : Suaedetalia corniculatae V.B. Golub 1994 (Folia Geobot. Phytotax. 29 : 46) nom. inval.], caractérisé par Suaeda salsa, S. altissima, S. acuminata, S. corniculata, Salicornia perennans, Atriplex calotheca, Bassia hyssopifolia, typus nominis hoc loco : Climacoptero crassae-Suaedion acuminatae V.B. Golub & Čorbadze 1989 (Folia Geobot. Phytotax. 24 : 120) corr. Lysenko & Mucina 2015. Provisoirement on propose de placer cet ordre encore dans les Thero-Suaedetea splendentis, quelques taxons reliant faiblement les deux ordres : Atriplex tatarica, Petrosimonia oppositifolia, Halimione pedunculata, Salicornia perennans.
Le système hiérarchique de la classe est actualisé comme suit (en grisé, les alliances présentes en France) :
THERO-SUAEDETEA SPLENDENTIS Rivas Mart. 1972
Thero-Suaedetalia splendentis Braun-Blanq. & O. Bolòs 1958
Salicornion procumbentis Tüxen ex Géhu 1976
Salicornion disarticulato-europaeae (Géhu & Géhu-Franck 1984) B. Foucault & Guitton 2021
Salicornion perennantis Géhu & Géhu-Franck 1984 nom. mut. B. Foucault & Guitton 2021
Thero-Suaedion splendentis Braun-Blanq. ex Braun-Blanq. in Braun-Blanq., Roussine & Nègre 1952
Microcnemion coralloidis Rivas Mart. 1984
Camphorosmo annuae-Suaedion maritimae nov.
Salicornion grandtneri Géhu & Géhu-Franck 1992
Atriplici gmelinii-Suaedion japonicae B. Foucault & Guitton 2021
Salsolo tetrapterae-Petrosimonion brachiatae nov. (cate 1)
Salicornio europaeae-Suaedion corniculatae nov.(carte 3)
Suaedetalia corniculatae V.B. Golub ex nov.
Climatoptero crassae-Suaedion acuminatae V.B. Golub & Čorbadze 1989 corr. Lysenko & Mucina 2015 (carte 2)
Camphorosmo songoricae-Suaedion corniculatae Freitag, V.B. Golub & Yuritsyna 2001 (carte 2)
Suaedion acuminatae V.B. Golub & Čorbadze in V.B. Golub 1995 corr. Lysenko & Mucina 2015 (carte 2)
Le Tamarici ramosissimae-Salsolion australis V.B. Golub 1994 est un complexe biologiquement hétérogène dont les composantes thérophytiques n’offrent que des liens ténus avec les Thero-Suaedetea splendentis (Golub, 1994b). En particulier, le premier taxon éponyme devrait caractériser des fourrés, éventuellement associé à Tamarix parviflora et T. tetrandra, Alhagi maurorum, Calligonum aphyllum… Ainsi, le Tamaricetum parvifloro-tetrandrae Hamzaoğlu & Aksoy 2006 (‘… parviflorae-…’ ; Hamzaoğlu & Aksoy, 2006, tableau 3 : relevés 370 à 389) gagnerait à être considéré, au moins potentiellement, comme un fourré halophile à rapprocher des Nerio oleandri-Tamaricetea africanae, mais peut-être pas des Nerio oleandri-Tamaricetalia africanae qui semblent plutôt ouest-méditerranéo-atlantiques, la composante herbacée vivace restant alors à interpréter et valider.
Les composantes thérophytiques des associations décrites par Shelyag-Sosonko et al. (1989, tableaux 2 à 6, Russie d’Europe) possèdent en commun Salicornia perennans subsp. p., Salsola soda, Petrosimonia oppositifolia, Suaeda maritima, Halimione pedunculata, Bassia sedoides, Pholiurus pannonicus, Myosurus minimus ; elles relèvent plus des Thero-Suaedetalia splendentis que des Suaedetalia corniculatae, se rapprochant du Thero-Suaedion splendentis.
Enfin, plusieurs des associations décrites de Crimée par Korženĕvskij et Kljukin (1991) ne se placent pas bien dans ce système tout en se rattachant nettement aux Suaedetalia corniculatae.
1.5. Une association des Crypsietea aculeatae
La composante thérophytique de l’Argusio sibiricae-Phragmitetum australis V.B. Golub & Mirkin 1986 de la Volga inférieure peut se définir par la liste synthétique suivante (d’après Golub & Mirkin, 1986, tableaux 26/27, sur 10/10 relevés) : Crypsis schoenoides V/IV, Atriplex calotheca IV/V, Crypsis aculeata II/III, Polygonum arenarium *pulchellum II/IV, Suaeda acuminata II/IV, Atriplex littoralis I/III, Echinochloa crus-galli +/-, Xanthium strumarium +/II, Salicornia europaea -/V ; il s’agit clairement d’une association originale des Crypsietea aculeatae, l’Atriplici calothecae-Crypsietum schoenoidis ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 2 du tableau 26 in Golub & Mirkin (1986, Folia Geobot. Phytotax. 21 : 388), avec une variation à Salicornia europaea.
2. Les prés halophiles vivaces
2.1. Les prés halophiles vivaces de Roumanie
Il faut d’abord compléter le synsystème ouest- et centre-européen déjà considéré avec les données suivantes de Ţopa (1939) synthétisées dans le tableau 7.
- Rhapontico serratuloidis-Oenanthetum silaifoliae Ţopa 1939 nom. mut. hoc loco (Ţopa, 1939 : 34, sub Leuzeo-Oenanthetum silaifoliae).
- Limonio gmelinii-Artemisietum santonici Ţopa 1939 nom. mut. hoc loco (Ţopa, 1939 : 36, sub Statico-Artemisietum monogynae).
- Composante vivace du Camphrosmetum annuae Ţopa 1939 (Ţopa, 1939 : 39).
- Triglochino maritimae-Tripolietum pannonici Ţopa 1939 nom. mut. hoc loco (Ţopa, 1939 : 41, sub Triglochino-Asteretum tripolii).
- Lepidietum crassifolii Ţopa 1939 (Ţopa, 1939 : 44).
- Camphorosmetum monspeliacae Ţopa 1939 (Ţopa, 1939 : 47, sub Camphorosmetum pilosae).
- Composante vivace du Crypsietum schoenoidis Ţopa 1939 (Ţopa, 1939 : 51).
- Composante vivace du Suaedo-Kochietum hirsutae sensu Ţopa 1939 (Ţopa, 1939 : 53).
- Halimionetum verruciferae Ţopa 1939 nom. mut. hoc loco (Ţopa, 1939 : 45, sub Obionetum verruciferae).
- Halocnemetum strobilacei Ţopa 1939 (Ţopa, 1939 : 49).
Dans la publication de 1939 (p. 34), l’auteur roumain définit validement le Puccinellio limosae-Limonion gmelinii où il place les syntaxons 1, 2 et 3, alliance que l’on peut lectotypifier par le Limonio gmelinii-Artemisietum santonici Ţopa 1939 (Bul. Făc. Şti. Cernăuţi XIII : 34) ; on peut aussi y rattacher les syntaxons 4 à 8. Par contre les syntaxons 9 et 10, caractérisés par les chaméphytes Halimione verrucifera et Halocnemum strobilaceum, sont distingués à part : Limonio gmelinii-Halimionion verruciferae all. nov. hoc loco, typus nominis hoc loco : Halimionetum verruciferae Ţopa 1939 (Bul. Făc. Şti. Cernăuţi XIII : 65), alliance caractérisée ou différenciée par Halimione verrucifera, Puccinellia distans subsp. limosa, Plantago schwarzenbergiana, Podospermum canum, Limonium bellidifolium.
L’existence et la validité de l’Halocnemetum strobilacei Ţopa 1939 rendent illégitime l’Halocnemetum strobilacei Quézel 1965 saharien (Quézel, 1965, tableau 5), qu’il faut alors renommer : Limoniastro guyoniani–Halocnemetum strobilacei (Quézel 1965) nom. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 5 in Quézel (1965, Geobot. Selecta II : 61).
2.2. Les prés halophiles vivaces anatoliens
Le tableau 8 synthétise les données sur les prés halophiles vivaces d’Anatolie rapportés par Yurdakulol et al. (1996), Aydoğdu et al. (2002) et Hamzaoğlu & Aksoy (2006, 2009.
- Composante vivace du Petrosimonietum nigdeensi-brachiatae Yurdakulol, Öncel, Demirörs, Yildiz & Keles 1996 (Yurdakulol et al., 1996, tableau 5).
- Lepidio caespitosi-Halimionetum portulacoidis Yurdakulol, Öncel, Demirörs, Yildiz & Keles 1996 (Yurdakulol et al., 1996, tableau 6).
- Cresso creticae-Halocnemetum strobilacei Yurdakulol, Öncel, Demirörs, Yildiz & Keles 1996 (Yurdakulol et al., 1996, tableau 7).
- Halocnemetum strobilacei Ţopa 1939 (Hamzaoğlu & Aksoy, 2006, tableau 3 : Halocnemetum strobilacei ; Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 1) ; à quelques détails près (absence de Limonium gmelinii, L. bellidifolium, présence de Camphorosma monspeliaca, Puccinellia festuciformis), on peut accepter le rattachement au syntaxon roumain de Ţopa en tant que race anatolienne.
- Lepidio caespitosi-Limonietum iconii Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2006, tableau 3 : Lepidio caespitosi-Limonietum iconici ; Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 2).
- Limonio tamaricoidis-Puccinellietum festuciformis Aydoğdu, Hamzaoğlu & Kurt 2002 nom. mut. hoc loco (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 3, sub Limonio tamaricoidis-Puccinellietum convolutae).
- Composante vivace du Suaedo anatolicae-Salsoletum nitrariae Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 5).
- Composante vivace du Sphenopodo divaricati-Halocnemetum strobilacei Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 4), qu’on peut distinguer en tant que Lepidio caespitosi-Halocnemetum strobilacei ass. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 8 du tableau 2 in Aydoğdu et al. (2002, Israel J. Pl. Sci. 50 : 316).
- Composante vivace du Bupleuro gracilis-Limonietum gmelinii Yurdakulol ex Hamzaoğlu in Hamzaoğlu & Aksoy 2009 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 6).
- Inulo aucherianae-Elytrigietum salsae Aydoğdu, Hamzaoğlu & Kurt 2002 nom. mut. hoc loco (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 7, sub Inulo aucheranae-Elymetum salsi).
- Eragrostio collinae-Puccinellietum anatolicae Aydoğdu, Hamzaoğlu & Kurt 2002 (Hamzaoğlu & Aksoy, 2009, tableau 2 : colonne 8).
- Inulo aucherianae-Elytrigietum salsae Aydoğdu, Hamzaoğlu & Kurt 2002 nom. mut. hoc loco (Hamzaoğlu & Aksoy, 2006, tableau 3 : Inulo aucheranae-Elymetum salsi et Tamaricetum parviflorae-tetrandrae), suffisamment distinct du syntaxon 10 pour justifier un nouveau nom : Inulo aucherianae-Aeluropodetum littoralis (Hamzaoğlu & Aksoy 2006) nom. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 371 du tableau 3 in Hamzaoğlu & Aksoy (2006, Ekoloji 15 (60) : 12).
- Puccinellio distantis-Juncetum maritimi Yurdakulol, Öncel, Demirörs, Yildiz & Keles 1996 (Yurdakulol et al., 1996, tableau 4).
Les syntaxons 1 à 9 relèvent du Lepidio caespitosi-Limonion iconii Aydoğdu, Hamzaoğlu & Kurt 2002 (Israel J. Pl. Sci. 50 : 315), typifié par le Lepidio caespitosi-Limonietum iconii Aydoğdu, Hamzaoğlu & Kurt 2002 (syntaxon 5), alors que les syntaxons 10 à 13 relèvent de l’Inulo aucherianae-Elytrigion salsae Aydoğdu, Hamzaoğlu & Kurt 2002 (Israel J. Pl. Sci. 50 : 319), typifié par l’Inulo aucherianae-Elytrigietum salsae Aydoğdu, Hamzaoğlu & Kurt 2002 (syntaxon 10). L’importance des chaméphytes dans la première alliance incite à la rattacher aux Sarcocornietea fruticosae, alors que la seconde se rapproche des Juncetea maritimi. L’aire de ces deux alliances est représentée en carte 1.
2.3. Les prés halophiles vivaces de Russie et d’Ukraine méridionales
Faute de précisions, ici on a été obligé de conserver le nom de “Puccinellia fominii” utilisé par les auteurs qui recouvre trois taxons d’après Euro+Med PlantBase : P. dolicholepis subsp. fominii, P. festuciformis subsp. f. et P. festuciformis subsp. lagascanae.
Le tableau 9 synthétise les données suivantes.
- Composante vivace du Suaedo-Petrosimonietum oppositifoliae V.B. Golub & Mirkin 1986 (Golub & Mirkin, 1986, tableau 11), isolée ici comme Limonio gmelinii-Aeluropodetum pungentis ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 4 du tableau 11 in Golub & Mirkin (1986, Folia Geobot. Phytotax. 21 : 363).
- Composante vivace du Suaedo salsae-Halocnemetum strobilacei V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 2) ; voir aussi le tableau 3 in Grechushkina et al. (2010, sub Salicornio prostratae–Halocnemetum strobilacei).
- Limonietum suffruticosi V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 3).
- Kalidietum foliati V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 4).
- Composante vivace du Suaedo salsae-Frankenietum hirsutae V.B. Golub & Čorbadze 1989 (Golub & Čorbadze, 1989, tableau 5).
- Composante vivace du Suaedo salsae-Petrosimonietum oppositifoliae V.B. Golub 1986 (Golub & Čorbadze, 1989, tableau 6).
Cet ensemble possède la valeur d’une alliance originale de prés halophiles vivaces, le Limonio gmelinii-Aeluropodion pungentis all. nov. hoc loco, caractérisée par Aeluropus pungens, Limonium gmelinii, L. suffruticosum, Halocnemum strobilaceum, Kalidium foliatum, typus nominis hoc loco : Limonio gmelinii-Aeluropodetum pungentis nov. (carte 2).
Le tableau 10 synthétise les données suivantes.
- Composante vivace du Salicornietum prostratae Soó ex Vicherek 1973 selon Dubyna & Neuhäuslová (2003, tableau 1).
- Composante vivace du Limonio caspii-Salicornietum prostratae Korženĕvskij & Kljukin 1991 (Dubyna & Neuhäuslová, 2003, tableau 2), que l’on propose de distinguer en Aeluropodo littoralis-Limonietum caspii ass. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 25 du tableau 2 in Dubyna & Neuhäuslová (2003, Thaiszia J. Bot. 13 (1) : 18).
- Composante vivace du Petrosimonio oppositifoliae–Salicornietum prostratae Korženĕvskij & Kljukin 1991 (Dubyna & Neuhäuslová, 2003, tableau 3).
- Composante vivace du Suaedo maritimae–Salicornietum prostratae Solomakha & Shelyag-Sosonko 1984 (Dubyna & Neuhäuslová, 2003, tableau 5).
- Composante vivace de l’Ofaisto monandri–Salicornietum prostratae Dubyna & Neuhäuslová 2003 (Dubyna & Neuhäuslová, 2003, tableau 4).
- Composante vivace du Suaedetum salsae V.B. Golub & Čorbadze 1995 (Dubyna & Neuhäuslová, 2003, tableau 6), que l’on propose de distinguer en Puccinellio syvaschicae-Limonietum meyeri ass. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 72 du tableau 6 in Dubyna & Neuhäuslová (2003, Thaiszia J. Bot. 13 (1) : 28).
- Puccinellio fominii-Aeluropodetum littoralis Shelyag-Sosonko, V.B. Golub & Solomakha 1989 (Shelyag-Sosonko et al., 1989, tableau 3, ‘…-Aeluropetum…’).
- Puccinellio fominii-Halimionetum verruciferae Shelyag-Sosonko, V.B. Golub & Solomakha 1989 (Shelyag-Sosonko et al., 1989, tableaux 4 et 5).
- Puccinellio fominii-Halocnemetum strobilacei Shelyag-Sosonko, V.B. Golub & Solomakha 1989 (Shelyag-Sosonko et al., 1989, tableau 6).
Ces syntaxons relèvent de l’Artemisio santonici-Puccinellion fominii Shelyag-Sosonko, V.B. Golub & Solomakha 1989 (Ukrayins’k. Bot. Z. 3 : 8), caractérisé notamment par Halocnemum strobilaceum, Puccinellia fominii s. l., Frankenia hirsuta, Artemisia santonicum, plusieurs Limonium (L. suffruticosum, L. gmelinii, L. meyeri, L. caspium)… et typifié par le Puccinellio fominii-Halimionetum verruciferae Shelyag-Sosonko, V.B. Golub & Solomakha 1989 (syntaxon 8).
2.4. Les prés halophiles vivaces de Russie centro-occidentale
Le tableau 11 synthétise les données suivantes.
- Poo bulbosae-Artemisietum pauciflorae Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 2).
- Puccinellio tenuissimae-Limonietum suffruticosi Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 3 : relevés 1 à 10).
- Leymo ramosi-Artemisietum austriacae Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 3 : relevés 11 à 18).
- Festuco beckeri-Artemisietum lerchianae Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 4 : relevés 1 à 6).
- Festuco valesiacae-Artemisietum austriacae Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 4 : relevés 7 à 16).
- Puccinellietum tenuissimae Karpov & Mirkin 1985 (Karpov et al., 2003, tableau 6 : relevés 1 à 4).
- Rhapontico serratuloidis-Puccinellietum dolicholepidis Karpov, Lysenko & V.B. Golub 2003 nom. mut. hoc loco (Karpov et al., 2003, tableau 6 : relevés 5 à 14, sub Stemmacantho serratuloidis-Puccinellietum dolicholepidis).
- Festuco pratensis-Artemisietum austriacae Karpov, Lysenko & V.B. Golub 2003 (Karpov et al., 2003, tableau 7).
- Composante vivace du Puccinellio tenuissimae-Camphorosmetum songoricae Lysenko, Karpov & V.B. Golub 2003 (Lysenko et al., 2003, tableau 2), réduite au premier taxon éponyme.
- Composante vivace de l’Atriplici laevis-Puccinellietum tenuissimae Golub et al. 2001 (Lysenko et al., 2003, tableau 3), que l’on propose de distinguer en Artemisio santonici-Puccinellietum tenuissimae ass. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 5 du tableau 3 in Lysenko et al. (2003, Vegetation of Russia: 45).
- Poo pratensis-Lepidietum latifolii Lysenko, Karpov & V.B. Golub 2003 (Lysenko et al., 2003, tableau 4).
- Composante vivace du Salicornio perennantis-Suaedetum salsae (Freitag et al., 2001, tableau 2 : relevés 13 à 24).
- Puccinellietum tenuiflorae Mirkin, Gogoleva & Kononov 1985 (Mirkin et al., 1985, tableau 11), incluant la composante vivace du Suaedo corniculatae-Puccinellietum tenuiflorae selon Mirkin et al. (1992, tableau 18).
- Knorringio sibiricae–Puccinellietum tenuiflorae Mirkin, Gogoleva & Kononov 1985 nom. mut. hoc loco (Mirkin et al., 1985, tableau 12, sub Polygono sibirici–Puccinellietum tenuiflorae), incluant la composante vivace du Suaedo corniculatae-Puccinellietum tenuiflorae selon Mirkin et al. (1985, tableau 13) ; peut se réduire à une sous-association à Knorringia sibirica du précédent.
- Composante vivace de l’Artemisio jacuticae-Suaedetum corniculatae Burtseva in Mirkin, Kononov, Gogoleva, Burtseva & Naumova 1992 (Mirkin et al., 1992, tableau 19), que l’on propose de distinguer en Artemisio jacuticae-Puccinellietum tenuiflorae ass. nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 2 du tableau 19 in Mirkin et al. (1992, Folia Geobot. Phytotax. 27 (3) : 290).
Les syntaxons 1 et 2 ont été placés dans le Poo bulbosae-Artemision pauciflorae Karpov, Lysenko & V.B. Golub 2003 (Vegetation of Russia 4 : 31) qui semble (publication en russe) typifié par le Poo bulbosae-Artemisietum pauciflorae Karpov, Lysenko & V.B. Golub 2003 (Vegetation of Russia 4 : 31). Les syntaxons 3 à 8 sont rassemblés au sein du Festuco valesiacae-Limonion gmelinii Mirkin in V.B. Golub & Solomakha 1988 (Бюл. Моск. O-ва ИспЫтателей ПриродЫ. Отд. Биол. 93 (6) : 90) qui semble (publication en russe) typifié par le Limonio gmelinii-Puccinellietum tenuissimae Karpov & Mirkin 1985, ce qui n’a pu être vérifié . Les syntaxons 9 à 12 peuvent être rapprochés dans une alliance caractérisée par Puccinellia tenuissima, Artemisia santonicum subsp. s., Limonium gmelinii, Tripolium pannonicum, le Tripolio pannonici-Puccinellion tenuissimae all. nov. hoc loco, typus nominis hoc loco : Artemisio santonici-Puccinellietum tenuissimae nov. (carte 2). Ces trois alliances partagent Limonium gmelinii.
Enfin les syntaxons 13 à 15 (de Russie nord-orientale : Yakoutie et vallée moyenne de la Lena) partagent surtout Puccinellia tenuiflora, Taraxacum ceratophorum et Saussurea amara : Saussureo amarae-Puccinellion tenuiflorae all. nov. hoc loco, typus nominis hoc loco : Polygono sibirici-Puccinellietum tenuiflorae Mirkin, Gogoleva & Kononov 1985 (Folia Geobot. Phytotax. 20 : 375) muté en Knorringio sibiricae–Puccinellietum tenuiflorae (carte 3).
Dans les paragraphes qui précèdent, ce sont donc dix alliances qui ont été mises en évidence, dont plusieurs déjà reconnues antérieurement et confirmées. Elles paraissent se répartir entre deux classes, les Sarcocornietea fruticosae et les Juncetea maritimi. On va maintenant reprendre ces alliances et les intégrer dans les tableaux synthétiques de ces classes (de Foucault, 2021a, 2021b).
2.5. Synthèse actualisée des Sarcocornietea fruticosae
Ainsi le tableau 12 concerne les Sarcocornietea fruticosae. Selon la synthèse de 2021a, réparties en trois ordres, les dix-huit premières alliances sont les Sarcocornion fruticosae Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952 (Sf), Suaedion braun-blanquetii Braun-Blanq. & O. Bolòs 1957 corr. Rivas Mart., Báscones, T.E. Díaz, Fern.Gonz. & Loidi 1991 (Sb), Halimionion portulacoidis Géhu 1975 (Hp), Suaedion verae (Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990) Rivas Mart., Fern.Gonz. & Loidi 1999 (Sv), Halimiono portulacoidis-Artemision santonici B. Foucault 2021 (HAs), Limonion confusi (Braun-Blanq. 1931) Rivas Mart. & M.J. Costa 1984 (Lc), Limonio compagnyonis-Arthrocnemion macrostachyi B. Foucault 2021 (LAm), Limonion lanceolato-algarvensis J.C. Costa, Neto, Monteiro-Henriques, Arsénio, E. Pereira, Caperta & Izco 2014 (Lla), Limoniastrion monopetali Pignatti 1953 (Lm), Triglochino barrelieri-Limonion glomerati Biondi, Diana, Farris & Filigheddu 2001 (TLg), Limonio virgati-Goniolimonion dalmatici (Horvatić 1934) B. Foucault 2021 (LGd), Limonion catalaunico-viciosoi Rivas Mart. & M.J. Costa 1976 (Lcv), Lygeo sparti-Lepidion cardaminis (Rivas Goday & Rivas Mart. 1963) Rivas Mart. & M.J. Costa 1976 (LLc), Lygeo sparti-Limonion angustebracteati Rigual ex Alcaraz, P. Sánchez & De la Torre 1988 (LLa), Limoniastrion guyoniani Quézel 1965 (Lg), Tetraeno albae-Arthrocnemion macrostachyi Géhu, Arnold, Géhu-Franck & Apostolidès ex B. Foucault 2021 (TAm) et Frankenio thymifoliae-Tetraenion albae B. Foucault 2021 (FTa). À l’extrême droite sont insérées les quatre colonnes LHv (Limonio gmelinii-Halimionion verruciferae ; tableau 7 : colonnes 9 et 10), LLi (Lepidio caespitosi-Limonion iconii ; tableau 8 : colonnes 1 à 9), LAp (Limonio gmelinii-Aeluropodion pungentis ; tableau 9) et APf (Artemisio santonici-Puccinellion fominii ; tableau 10). Ce tableau montre que ces quatre alliances s’intègrent mal dans les trois premiers ordres, tout en partageant Artemisia santonicum et Limonium gmelinii avec HAs, Frankenia hirsuta avec TAm, Halocnemum strobilaceum avec les Limoniastretalia guyoniani. Il faut donc définir un quatrième ordre.
Ne pouvant reprendre les Halostachyetalia belangerianae Ţopa 1939 (Bul. Făc. Şti. Cernăuţi XIII : 64), syntaxon invalide par défaut d’indication d’au moins une alliance (en p. 30 de sa publication, l’auteur roumain passe en effet directement de l’ordre à ses quatre associations). On ne peut pas suivre Golub et Čorbadze (1989) qui valident l’ordre chaméphytique par une alliance thérophytique considérée ici même au sein des Thero-Suaedetea splendentis, le Climacoptero-Suaedion acuminatae. On définit donc ici les Halimionetalia verruciferae ord. nov. hoc loco, ordre caractérisé par Halimione verrucifera, Limonium suffruticosum, L. gmelinii, Frankenia hirsuta, Artemisia santonicum et partageant Halocnemum strobilaceum avec les Limoniastretalia guyoniani, typus nominis hoc loco : Lepidio caespitosi-Limonion iconii Aydoğdu, Hamzaoğlu & Kurt 2002 (Israel J. Pl. Sci. 50 : 315).
La hiérarchie synsystématique des Sarcocornietea fruticosae est ainsi actualisée (en grisé les alliances présentes en France) :
SARCOCORNIETEA FRUTICOSAE Braun-Blanq. & Tüxen ex A. Bolòs & O. Bolòs in A. Bolòs 1950
Sarcocornietalia fruticosae Braun-Blanq. 1931 ex 1933
Sarcocornion fruticosae Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952
Suaedion braun-blanquetii Braun-Blanq. & O. Bolòs 1957 corr. Rivas Mart., Báscones, T.E. Díaz, Fern.Gonz. & Loidi 1991
Halimionion portulacoidis Géhu 1975
Suaedion verae (Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990) Rivas Mart., Fern.Gonz. & Loidi 1999
Halimiono portulacoidis-Artemision santonici B. Foucault 2021
Limonietalia Braun-Blanq. & O. Bolòs 1957
Eu-Limonienalia
Limonion confusi (Braun-Blanq. 1931) Rivas Mart. & M.J. Costa 1984
Limonio compagnyonis-Arthrocnemion macrostachyi B. Foucault 2021
Limonion lanceolato-algarvensis J.C. Costa, Neto, Monteiro-Henriques, Arsénio, E. Pereira, Caperta & Izco 2014
Limoniastrion monopetali Pignatti 1953 (en France ?)
Triglochino barrelieri-Limonion glomerati Biondi, Diana, Farris & Filigheddu 2001
Limonio virgati-Goniolimonion dalmatici (Horvatić 1934) B. Foucault 2021
Limonio delicatuli-Lygeenalia sparti B. Foucault 2021
Limonion catalaunico-viciosoi Rivas Mart. & M.J. Costa 1976
Lygeo sparti-Lepidion cardaminis (Rivas Goday & Rivas Mart. 1963) Rivas Mart. & M.J. Costa 1976
Lygeo sparti-Limonion angustebracteati Rigual ex Alcaraz, P. Sánchez & De la Torre 1988
Limoniastretalia guyoniani Guin. 1951
Limoniastrion guyoniani Quézel 1965
Tetraeno albae-Arthrocnemion macrostachyi Géhu, Arnold, Géhu-Franck & Apostolidès ex B. Foucault 2021
Frankenio thymifoliae-Tetraenion albae B. Foucault 2021
Halimionetalia verruciferae nov.
Limonio gmelinii-Halimionion verruciferae nov.
Lepidio caespitosi-Limonion iconii Aydoğdu, Hamzaoğlu & Kurt 2002 (carte 1)
Limonio gmelinii-Aeluropodion pungentis nov. (carte 2)
Artemisio santonici-Puccinellion fominii Shelyag-Sosonko, V.B. Golub & Solomakha 1989
2.5. Synthèse actualisée des Juncetea maritimi
Il s’agit maintenant de considérer le système hiérarchique des autres unités mises en évidence ici au sein des Juncetea maritimi. Le tableau 13 reprend le tableau synthétique 1 de la synthèse précédente (de Foucault, 2021b) avec les nouvelles unités. On rappelle les unités précédentes : Pph pour Puccinellion phryganodis, Cg pour Caricion glareosae, Ppu pour Puccinellion pumilae, SPd pour Stellario humifusae-Puccinellietum distantis, Sa pour Spartinion alterniflorae, Pm pour Puccinellion maritimae, PSs pour Puccinellio maritimae-Spergularion marinae, Am pour Armerion maritimae, LJm pour Lysimachio maritimae-Juncion maritimi, SPm pour Spergulario mediae-Plantaginion maritimae, Jm pour Juncion maritimi, Pca pour Puccinellion caespitosae, Pf pour Puccinellion festuciformis, Pcr pour Plantaginion crassifoliae, SJa pour Schizogyno sericeae-Juncion acuti, Pd pour Puccinellion distantis, Pi pour Puccinellion intermediae, Fp pour Festucion pseudovinae, LgJm pour Limonio gmelinii-Juncion maritimi, ITr pour Imperato cylindricae-Tripidion ravennae, EPm pour Elytrigio acutae-Plantaginion maritimae, AJr pour Aeluropodo lagopoidis-Juncion rigidi. Les nouvelles colonnes à intégrer sont PLg (Puccinellio limosae-Limonion gmelinii ; tableau 7 : colonnes 1 à 8), IEs (Inulo aucherianae-Elytrigion salsae ; tableau 8 : colonnes 10 à 13), PAp (Poo bulbosae-Artemision pauciflorae ; tableau 11 : colonnes 1 et 2), FLg (Festuco valesiacae-Limonion gmelinii ; tableau 11 : colonnes 3 à 8), TPt (Tripolio pannonici-Puccinellion tenuissimae ; tableau 11 : colonnes 9 à 12) et SPt (Saussureo amarae-Puccinellion tenuiflorae ; tableau 11 : colonnes 13 à 15). Ce tableau 12 montre finalement la bonne intégration de cinq de ces six alliances aux Festuco-Puccinellietalia [incluant donc les Festuco-Limonietalia gmelinii Mirkin in V.B. Golub & Solomakha 1988 (Бюл. Моск. O-ва ИспЫтателей ПриродЫ. Отд. Биол. 93 (6) : 80-92)], ordre dont de bonnes caractéristiques ou différentielles vis-à-vis des ordres voisins sont Plantago maritima subsp. ciliata, Taraxacum besarabicum, Artemisia santonicum, Puccinellia tenuissima, P. distans subsp. d., Lepidium cartilagineum, Limonium gmelinii, plus Halimione verrucifera issu des Halimionetalia verruciferae ; seul le Saussureo amarae-Puccinellion tenuiflorae reste marginal, cette alliance ne relève sans doute pas des Juncetea maritimi ; peut-être correspond-elle à une irradiation occidentale ou nord-occidentale d’une unité extrême-orientale non encore définie.
Le schéma synsystématique global de la classe devient donc (en grisé, les alliances et sous-alliances présentes en France).
JUNCETEA MARITIMI Tüxen & Oberd. 1958
Puccinellietalia phryganodis Hadač 1946
Puccinellion phryganodis Hadač 1946
Caricion glareosae Nordh. 1954
Puccinellion pumilae A. Miyaw. & Ohba 1965
Dupontion fischeri Hadač 1946
Limonio caroliniani-Puccinellion phryganodis B. Foucault 2021
Spartinetalia alterniflorae Conard 1935
Spartinion alterniflorae Conard 1935
Lysimachio maritimae-Puccinellietalia maritimae Beeftink & V. Westh. in Beeftink 1962
Puccinellion maritimae W.F. Christ. 1927
Puccinellio maritimae-Spergularion marinae Beeftink 1965
Armerion maritimae Braun-Blanq. & De Leeuw 1936
Eu-Armerienion maritimae
Frankenio laevis-Armerienion maritimae Géhu & Géhu-Franck in Géhu 1976
Lysimachio maritimae-Juncion maritimi Géhu & Géhu-Franck ex Géhu in Bardat et al. 2004
Spergulario mediae-Plantaginion maritimae B. Foucault 2021
Juncetalia maritimi Braun-Blanq. ex Horvatić 1934
Juncion maritimi Braun-Blanq. ex Horvatić 1934
Puccinellion caespitosae Rivas Mart. in Rivas Mart. & M.J. Costa 1976
Puccinellion festuciformis Micevski 1965
Plantaginion crassifoliae Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952
Eu-Plantaginion crassifoliae
Limonio virgati-Plantaginenion crassifoliae Mercadal 2020
Schizogyno sericeae-Juncion acuti B. Foucault 2021
Cression creticae B. Foucault 2021
Festuco-Puccinellietalia Soó 1968
Puccinellion distantis (Soó 1933) Wendelb. 1943
Puccinellion intermediae Wendelb. 1943
Festucion parviflorae Soó in Máthé 1933 nom. mut.
Limonio gmelinii-Juncion maritimi V.B. Golub & Solomacha ex Dubyna & Neuhäuslová 2000
Puccinellio limosae-Limonion gmelinii Ţopa 1939
Inulo aucherianae-Elytrigion salsae Aydoğdu, Hamzaoğlu & Kurt 2002 (carte 1)
Poo bulbosae-Artemision pauciflorae Karpov, Lysenko & V.B. Golub 2003
Festuco valesiacae-Limonion gmelinii Mirkin in V.B. Golub & Solomakha 1988
Tripolio pannonici-Puccinellion tenuissimae nov. (carte 2)
Tripidietalia ravennae Biondi, Blasi & Casavecchia in Biondi et al. 2014
Imperato cylindricae-Tripidion ravennae Braun-Blanq. & O. Bolòs 1958
Elytrigietalia acutae Géhu 1968
Elytrigio acutae-Plantaginion maritimae Horvatić 1934
Lysenko et Mucina (2015) définissent encore un Carici dilutae-Juncetum gerardii Lysenko & Mucina 2015, holotype du Carici dilutae-Juncion gerardii Lysenko & Mucina 2015 (Hacquetia 14 (2) : 302), d’après les relevés 1 à 4 du tableau 12 in Klotz & Köck (1984, Feddes Repert. 95 : 406, sub Junco-Glaucetum maritimae, où Carex diluta est dénommé C. karelinii) provenant de Bachkirie (carte 2). Cet État se situant entre les fleuves Volga à l’ouest et Oural à l’est, donc en Russie d’Europe, ces syntaxons devraient se rattacher aux Festuco-Puccinellietalia, mais les taxons strictement caractéristiques d’ordre sont rares dans les quatre relevés connus. Peut-être ce nombre réduit de relevés ne permet-il pas de décrire plus nettement cette alliance russe.
3. Conclusion
Cette extension vers l’Europe orientale puis l’Asie septentrionale de la connaissance de la végétation halophile selon une méthodologie plus moderne a donc permis la définition de deux ordres (Suaedetalia corniculatae et Halimionetalia verruciferae), de sept alliances (Salsolo stenopterae-Petrosimonion brachiatae, Camphorosmo annuae-Suaedion maritimae, Salicornio europaeae-Suaedion corniculatae, Limonio gmelinii-Halimionion verruciferae, Limonio gmelinii-Aeluropodion pungentis, Tripolio pannonici-Puccinellion tenuissimae et Saussureo amarae-Puccinellion tenuiflorae) et de dix-sept associations. Il y aurait bien d’autres publications à consulter et analyser, mais leur accès est difficile, tant pour les acquérir que pour les comprendre (car souvent rédigées en langues slaves, surtout le russe).
Bibliographie
Aydoğdu M., Hamzaoğlu E. & Kurt L., 2002. New halophytic syntaxa from Central Anatolia (Turkey). Israel Journal of Plant Sciences 50 : 313-323.
Dítĕ D., Elias P. & Marek S., 2008. Camphorosmetum annuae Rapaics ex Soó 1933, vanishing plant community of saline habitats in Slovakia. Thaiszia Journal of Botany 18 (1) : 51-64.
Dubyna D.V. & Neuhäuslová Z., 2003. The vegetation of the Azov-Sivaš National Nature Park. Class Thero-Salicornietea (S. Pignatti 1953) R. Tx. in R. Tx. et Oberdorfer 1958. Thaiszia Journal of Botany 13 (1) : 1-30.
Dubyna D.V., Dziuba T.P., Iemelianova S.M. & Makhynia L.М., 2020. Syntaxonomy and ecological differentiation of the pioneer vegetation of Ukraine. 1 – Classes: Cakiletea maritimae, Ammophiletea, Crithmo-Staticetea, Crypsietea aculeatae, Therosalicornietea. Biosystems Diversity 28 (1) : 68-80.
Foucault B. (de), 2021a. Contribution à une synthèse des Sarcocornietea fruticosae. Carnets botaniques 59 : 1-19 + 17 tableaux, https://doi.org/10.34971/8h0c-0b94.
Foucault B. (de), 2021b. Contribution à une synthèse des Juncetea maritimi. Carnets botaniques 68 : 1-30 + 22 tableaux, https://doi.org/10.34971/nbat-mh09.
Foucault B. (de), 2021c. Contribution à la connaissance de la végétation des étangs asséchés et des canaux du Narbonnais. Carnets botaniques 38 : 1-15 + 3 tableaux, https://doi.org/10.34971/387K-6576.
Foucault B. (de) & Guitton H., 2021. Une déclinaison moderne des Thero-Suaedetea splendentis. Carnets botaniques 76 : 1-9 + 2 tableaux, https://doi.org/10.34971/3XK3-N640.
Freitag H., Golub V.B. & Yuritsyna N.A., 2001. Halophytic plant communities in the northern Caspian lowlands, I – Annual halophytic communities. Phytocoenologia 31 (1) : 63-108.
Golub V.B., 1994a. Class Asteretea tripolium on the territory of the former USSR and Mongolia. Folia Geobotanica et Phytotaxonomica 29 : 15-54.
Golub V.B., 1994b. The desert vegetation communities of the Lower Volga valley. Feddes Repertorium 105 (7-8) : 499-515.
Golub V.B., 1995. Halophytic, desert and semidesert plant communities on the territory of the former USSR. Russian Academy of Sciences, Institute of Ecology of the Volga River Basin, Togliatti, 32 p.
Golub V.B. & Mirkin B.M., 1986. Grasslands of the lower Volga valley. Folia Geobotanica et Phytotaxonomica 21 : 337-395.
Golub V.B. & Solomakha V.A., 1988. The highest units of the classification of the salt soil vegetation in the European part of the USSR. Бюл. Моск. O-ва ИспЫтателей ПриродЫ. Отд. Биол. 93 (6) : 80-92 (en russe).
Golub V.B. & Čorbadze N.B., 1989. The communities of the order Halostachyetalia Ţopa 1939 in the area of western substeppe ilmens of the Volga delta. Folia Geobotanica et Phytotaxonomica 24 : 113-130.
Grechushkina N.A., Sorokin A.N. & Golub V.B., 2010. Plant communities oft he Thero-Salicornietea and Salicornietea fruticosae classes along the Russian coast of the sea of Azov. Chornomors’k. Bot. Z. 6 (2) : 200-212 (en russe).
Hamzaoğlu E. & Aksoy A., 2006. Sultansazlığı bataklığı halofitik toplulukları Üzerine fitososyolojik Bir Çalışma (Iç Anadolu-Kayseri). Ekoloji 15 (60) : 8-15 (en turc).
Hamzaoğlu E. & Aksoy A., 2009. Phytosociological studies on the halophytic communities of Central Anatolia. Ekoloji 16 (71) : 1-14.
Karpov D.N., Lysenko T.M. & Golub V.B., 2003. Plant communities on solonetz and salinized soils oft he Southern Ural. In Vegetation of Russia, Saint-Pétersbourg 4 : 29-41 (en russe).
Klotz S. & Köck U.V., 1984. Vergleichende geobotanische Untersunchungen in der Baschkirischen ASSR, 3. Teil: Wasserpflanzen-, Flubufer- und Halophytenvegetation. Feddes Repertorium 95 (5-6) : 381-408.
Korženĕvskij V.V. & Kljukin A.A., 1991. Vegetation description of mud volcanoes of Cremia. Feddes Repertorium 102 (1-2) : 137-150.
Lysenko T.M., Karpov D.N. & Golub V.B., 2003. Halophytic plant communities oft he Stavropol’s depression (Samara Region). In Vegetation of Russia, Saint-Pétersbourg 4 : 42-50 (en russe).
Lysenko T. & Mucina L., 2015. Nomenclatural notes on some alliances of the halophytic vegetation of southern Ural and the Caspian lowlands. Hacquetia 14 (2) : 301-306.
Mirkin B.M., Gogoleva P.A. & Kononov K.E., 1985. The vegetation of Central Yacutian Alases. Folia Geobotanica et Phytotaxonomica 20 : 345-395.
Mirkin B.M., Kononov K.E., Gogoleva P.A., Burtseva E.I. & Naumova L.G., 1992. The floodplain grasslands of the Middle Lena-river, II – Classification. Folia Geobotanica et Phytotaxonomica 27 (3) : 247-300.
Quézel P., 1965. La végétation du Sahara, du Tchad à la Mauritanie. Geobotanica Selecta II : 1-333.
Shelyag-Sosonko Y.R., Golub V.B. & Solomakha V.A., 1989. The syntaxonomy of classis Salicornietea fruticosae of halophitic vegetation of European part of the USSR. Ukrayins’kyi botanichnyi zhurnal 3 : 5-10 (en russe).
Ţopa E., 1939. Vegetaţia halofitelor din Nordul României in legătură cu cea din restul ţării. Buletinul Făcultaţii de Ştiinţe din Cernăuţi XIII : 1-79 (en roumain, avec un large résumé en français).
Townsend C.C., 1993. Amaranthaeae. In K. Kubitzki, J.G. Rohwer & V. Bittrich, The families and genera of vascular plants II, Springer : 70-91.
Vicherek J., 1973. Die Pflanzengesellschaften der Halophyten- und Subhalophytenvegetation der Tschechoslowakei. Vegetace ČSSR A 5 : 1-200.
Yurdakulol E., Öncel I., Demirörs M., Yildiz A. & Keles Y., 1996. Ecological and syntaxonomic investigation of salt marshes vegetation in the vicinity of Burdur and Acigöl (Denizli/Turkey). Ecologia Mediterranea XXII (1-2) : 51-61.
Remerciements
De vifs remerciements s’adressent à ceux qui ont contribué à la recherche documentaire, la Bibliothèque botanique et phytosociologique de France (Conservatoire botanique national de Bailleul) et Vincent Gaudillat (cellule Habitats, UMS 2006 Patrimoine naturel, MNHN, Paris), ainsi qu’à Jean-Paul Theurillat pour un point dans l’application de l’ICPN v. 4.


