
Miscellanées phytosociologiques IX : syntaxons nouveaux, validés ou complétés
Title
Phytosociological miscellanées IX: new, validated or completed syntaxa
Résumé
Ce neuvième numéro de Miscellanées phytosociologiques apporte de nouvelles données sur des forêts, notamment de Crète, de Syrie, de Slovaquie et du Japon (Honshu), la végétation chasmophile, halophile, amphibie thérophytique ; une grande partie est consacrée aux bas-marais oligotrophiles (du Québec, du Kamchatka, à Rhynchospora alba).
Abstract
This ninth issue of Miscellanées phytosociologiques provides new data on forests, particularly from Crete, Syria, Slovakia and Japan (Honshu), chasmophilous, halophilous and amphibious therophytic vegetation; a large part is devoted to oligotrophic fens (from Quebec, Kamchatka, with Rhynchospora alba).
Cette neuvième livraison de Miscellanées phytosociologiques apporte quelques précisions syntaxonomiques, quelques corrections et validations à propos de végétations forestières, chasmophytiques, halophiles, amphibies, de bas-marais (dont surtout ceux à Rhynchospora alba).
1. Une précision préalable
Dans ma publication sur la végétation de la Colombie (de Foucault, 2025a), un taxon était resté en suspens suite à son absence des référentiels usuels. Ph. Julve vient de m’en donner le nom actuellement valide : Rhynchospora daweana Britton doit être appelé Rh. aristata Boeckeler ; un grand merci à lui.
2. Quelques associations forestières
2.1. De France
Le « Carici elongatae-Alnetum glutinosae » est une association de forêt hygrophile mésotrophile classique en Europe, mais dont la définition demande une révision selon le paradigme synusial. L’association strictement arborescente ressortit au Betulo pubescentis-Alnetum glutinosae Julve & Gillet ex B. Foucault 2018 (de Foucault, 2018a, 2020). Compte tenu de l’hétérogénéité fréquente du sol de cette aulnaie, avec des mosaïques de zones nettement inondées et de zones plus atterries, on ne peut parler d’une seule association herbacée vivace, mais de plusieurs. Parmi celles-ci, il y a donc une association hygrophile à Carex elongata, mais on ne peut utiliser la plupart des tableaux publiés (Bodeux, 1955 ; Döring, 1987 ; Mast, 2007…) pour la mettre en évidence. Des adeptes du paradigme synusial ont décrit de telles associations à C. elongata :
- Julve et Gillet (1994 : 56) définissent le Thelypterido palustris-Caricetum elongatae prov. ;
- Decocq (1999 : 194) définit le Carici elongatae-Deschampsietum cespitosae; ce nom reste toutefois invalide : sa thèse de 1997 à laquelle il fait référence est un mémoire inédit et elongata manque dans le tableau 3 (p. 198 à 201) de cet article de 1999.
Il semble que le premier des deux soit plus continental (avec Thysselinum palustre, Carex canescens ; décrit de Pologne, on le retrouve en Basse-Saxe allemande, Döring, 1987) que le second, on peut donc valider ici le Carici elongatae-Deschampsietum cespitosae Decocq ex ass. nov. hoc loco, typus nominis hoc loco : relevé in de Foucault (1994, Bull. Soc. Bot. N. France 47 (2) : 18, bas de page).
Le Mercuriali perennis-Hyacinthoidetum non-scriptae a été défini dans les Hauts-de-France (de Foucault et al., 1996 ; de Foucault, 1997), mais est resté invalide faute de désignation d’un type nomenclatural ; on le valide ici : le typus nominis hoc loco est désigné comme étant le relevé 12 du tableau 3 in de Foucault (1997, Nat. Mosana 50 (3) : 72-73).
En forêt domaniale de Crécy-en-Ponthieu (Somme), au nord-ouest de la maison forestière des Vieux-Chênes, le long d’un sentier inondable (N 50° 13’ 37,5’’, E 1° 50’ 12,6’’, 61 m), le 27 juin 2025, a été observé l’ourlet thérophytique hygrophile à Impatiens noli-tangere (photo 1) et Persicaria hydropiper initialement décrit des Ardennes françaises sous le nom de Persicario hydropiperis-Impatientetum noli-tangere (de Foucault, 2011 ; photo 2) et revu ultérieurement dans les Pyrénées audoises (de Foucault, 2019).


2.2. Une microphorbiaie à Saxifraga hirsuta
Dans ma synthèse des Montio-Cardaminetea amarae (de Foucault, 2018b), j’ai été amené à définir l’alliance du Lysimachio nemorum-Saxifragion hirsutae pour le Pays basque. En étudiant la structure synfloristique de l’« Hyperico androsaemi-Alnetum glutinosae », je suis amené à décrire ici une quatrième association sur la base des tableaux 40 de Loidi (1983) et 6 de Rivas-Martínez et al. (1984), le Myosotido lamottianae-Saxifragetum hirsutae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 1 du tableau 6 in Rivas-Martínez et al. (1984, Lazaroa 6 : 145).
2.3. D’Albanie
De massifs d’Albanie, entre 650 et 1 100 m d’altitude, Hoda (1993) décrit une forêt à Pinus nigra et Forsythia europaea, où l’on peut distinguer de façon plus moderne :
- une association arborescente (sur 12 relevés hors relevé 8), Pinus nigra *n. V, Fraxinus ornus III, Quercus petraea III, dalechampii II, Abies alba I, Acer pseudoplatanus I, A. platanoides +, le Fraxino orni-Pinetum nigrae ass. nov. hoc loco, typus nominis hoc loco: composante arborescente (Trees) du relevé 5 du tableau 2 in Hoda (1993, Annali di botanica LI : 84), à rapprocher du Quercion congesto-dalechampii B. Foucault & Julve ex B. Foucault 2020 (de Foucault, 2020) ;
- une association arbustive (sur l’ensemble des 13 relevés) : Forsythia europaea V, Juniperus oxycedrus *o. III, Rubus ulmifolius III, idaeus II, Sorbus graeca I, Prunus mahaleb I, soit le Junipero oxycedri-Forsythietum europaeae ass. nov. hoc loco, typus nominis hoc loco: composante arbustive (Shrubs) du relevé 11 du tableau 2 in Hoda (1993, Annali di botanica LI : 84).
Quant à l’association herbacée intraforestière, on peut la décrire comme Pteridio aquilini-Betonicetum scardicae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée (Herbs) du relevé 4 du tableau 2 in Hoda (1993, Annali di botanica LI : 84, en changeant Stachys scardica en Betonica s.).
2.4. De Slovaquie
En 2022, Kučera a décrit des pessières de Slovaquie selon le paradigme phytosociologique classique. On les reprend ici selon le paradigme synusial.
Ainsi, le tableau 1 hoc loco en décrit la végétation arborescente ; les colonnes 1 à 3, les plus significatives, décrivent le Sorbo aucupariae-Piceetum abietis ass. nov. hoc loco, déjà entrevu par Gallandat et al. (1995), typus nominis hoc loco : composante arborescente (E3 + E2) du relevé 1 du tableau 1 in Fajmonová (1986, Preslia 58 : 44-49).
Le tableau 2 est dédié à la végétation arbustive du sous-bois, guère caractérisée, y compris dans les colonnes 1 à 4, les plus significatives avec Lonicera nigra, Daphne mezereum, Clematis alpina, Rosa pendulina, Ribes petraeum, Pinus mugo, mais pouvant être rapprochée du Sorbo mougeotii-Lonicerion nigrae B. Foucault ex B. Foucault 2024.
Le tableau 3 développe la végétation herbacée vivace intraforestière en ajoutant aux données de Kučera celles de Fajmonová (1986). Bien caractérisée, la colonne 1 pourrait être décrite comme Crepido jacquinii-Seslerietum caeruleae, mais il faudrait avoir accès à la publication de Fajmonová (1978) pour la valider. Les colonnes 2 à 6 décrivent le Seslerio caeruleae-Cardaminetum enneaphylli ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 1 du tableau 1 in Fajmonová (1986, Preslia 58 : 44-49). Il en est des colonnes 7 à 9 comme de la colonne 1, c’est-à-dire sans données aisément disponibles pour les valider ; la colonne 7 paraît représenter une végétation de forêt de ravin avec Actaea spicata, Polystichum aculeatum, Lunaria rediviva, vicariante des végétations ouest-européennes de l’Actaeo spicatae-Mercurialion perennis Gillet ex Gillet & B. Foucault in B. Foucault 2020. La colonne 10 peut en revanche être validée comme Calamagrostietum arundinaceo-villosae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée du relevé 1 du tableau 3 in Kučera (2022, Hacquetia 21 (1) : 135).
L’ensemble des colonnes 1 à 6 a la valeur d’une alliance de végétation herbacée intraforestière centre-européenne vicariante du Seslerio caeruleae-Mercurialion perennis Gillet ex Gillet & B. Foucault in B. Foucault 2020 ouest-européen, essentiellement caractérisée par Sesleria caerulea, Cardamine enneaphyllos, Bellidiastrum michelii, Arabidopsis arenosa, Rubus saxatilis, Carduus defloratus subsp. glaucus, Gymnocarpium robertianum, Poa stiriaca, Mercurialis perennis, le Seslerio caeruleae-Cardaminion enneaphylli all. nov. hoc loco, typus nominis hoc loco : le Seslerio caeruleae-Cardaminetum enneaphylli nov.
2.5. De Crète et Syrie
Sur la base des données publiées par Barbero & Quézel (1980), la végétation forestière crétoise avait été abordée dans un article antérieur (de Foucault, 2022), mais sans interprétations définitives ; on va donc la reprendre ici, en y ajoutant des données sur des forêts syriennes issues de Barbero & Quézel (1976).
Le tableau 4 synthétise les données sur la végétation arborescente. En premier lieu, les colonnes 1 à 3 décrivent une forêt thermoméditerranéenne de très basse altitude à Quercus coccifera arborescent, Olea europaea et Ceratonia siliqua, Querco cocciferae-Oleetum europaeae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 1 du tableau I in Barbero & Quézel (1980, Ecol. Medit. 5 : 181). Les colonnes 4 et 5, correspondant à des altitudes un peu supérieures, décrivent une forêt caractérisée par Prunus webbii et Quercus pseudococcifera, le Pruno webbii-Quercetum pseudococciferae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 8 du tableau IV in Barbero & Quézel (1980, Ecol. Medit. 5 : 189, en changeant Amygdalus webbii en Prunus w.). Les colonnes 9 à 11 peuvent se rattacher au Pino brutiae-Ceratonietum siliquae B. Foucault 2025 (de Foucault, 2025d). On propose de rattacher tout l’ensemble formé par les colonnes 1 à 12 à l’Oleo europaeae-Ceratonion siliquae Braun-Blanq. ex Guin. & Drouineau 1944.
Dans l’ensemble des colonnes 13 à 17, notamment la colonne 16 peut s’isoler comme Querco cocciferae-Castaneetum sativae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 14 du tableau VI in Barbero & Quézel (1980, Ecol. Medit. 5 : 195).
Les colonnes 19 à 21 correspondent à une association arborescente antérieurement définie comme Aceri sempervirentis-Cupressetum sempervirentis (Barbero & Quézel 1980) B. Foucault ex B. Foucault 2020 (de Foucault, 2020).
Provenant de Syrie littorale, les colonnes 22 et 23 sont à rattacher au Querco pubescentis-Pinetum halepensis B. Foucault 2020. La colonne 26 est une variation à Pinus brutia du Quercetum veneris-cerridis B. Foucault 2025 décrit d’Anatolie orientale (de Foucault, 2025b).
Les associations arbustives intraforestières crétoises ont déjà été abordées dans une publication précédente (de Foucault, 2022, tableau 5). On avait alors défini des associations et une alliance à Cytisus creticus, le Cytision cretici B. Foucault 2022, typifié par le Cytiso cretici-Ericetum arboreae B. Foucault 2022. Mais le grand ensemble Ia à VIIa de ce tableau 5 de 2022 avait été quelque peu laissé de côté, il est repris ici dans le tableau 5 ; on peut y mettre en exergue :
- la colonne 1 rappelant l’Euphorbio dendroidis-Prasietum majoris Zaffran & B. Foucault in Foucault 2001, aussi décrit de Crète mais pourtant floristiquement bien plus riche ici (de Foucault, 2021a), se rattachant plutôt à l’Asparago aphylli-Pistacietum lentisci (Géhu, Apostolidès, Géhu-Franck & Arnold 1992) B. Foucault 2021 ;
- la colonne 8 comme Cisto parviflori-Juniperetum turbinatae ass. nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 6 du tableau III in Barbero & Quézel (1980, Medit. 5 : 185, en remplaçant Juniperus phoenicea subsp. lycia par J. ph. subsp. turbinata).
L’interprétation de ce grand ensemble 1 à 13, qui relève manifestement des Rubio tenuifoliae-Pistacietalia terebinthi B. Foucault 2021 (de Foucault, 2021b), est l’occasion de reprendre ici tout ce qui est connu sur cet ordre, grâce au tableau synthétique 6, venant ainsi compléter une synthèse récente (de Foucault, 2025d, tableau 3). Tout d’abord, le Styraco officinalis-Pistacietum terebinthi B. Foucault 2025 décrit de Palestine (de Foucault, 2025c) n’entre bien dans aucune de ces alliances. On voit par ailleurs que la colonne ARo, qui synthétise cet ensemble 1 à 13 du tableau 5 hoc loco, est originale au sein de l’ordre des Rubio tenuifoliae-Pistacietalia terebinthi et a valeur d’alliance, ne pouvant se rattacher à aucune des alliances déjà connues, caractérisée par Rhamnus lycioides subsp. oleoides, Euphorbia characias subsp. c., Salvia fruticosa, Asparagus aphyllus, Cistus creticus, Dioscorea cretica : Asparago aphylli-Rhamnion oleoidis all. nov. hoc loco, typus nominis hoc loco : le Cisto parviflori-Juniperetum turbinatae nov.
La végétation des sous-bois herbacés crétois a fait l’objet d’une synthèse antérieure (de Foucault, 2012, tableau 7), mais sans interprétation définitive, à l’exception du Phlomido creticae-Crepidion fraasii. On poursuit donc ici cette synthèse restée inachevée. Le tableau 7 hoc loco reprend le tableau 7 de 2022 tout en déplaçant la colonne IIa dans le second ensemble où il devient la colonne 14.
Dans l’ensemble des colonnes 1 à 12, on peut mettre en évidence
- en colonne 5, l’Ebeno creticae-Gelasietum creticae ass. nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 2 du tableau III in Barbero & Quézel (1980, Medit. 5 : 185, en changeant Scorzonera cretica en Gelasia c.) ;
- en colonne 6, l’Helichryso orientalis-Iridetum cretensis nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 12 du tableau III in Barbero & Quézel (1980, Ecol. Medit. 5 : 185) ;
- en colonne 8, le Bunio ferulacei-Aristolochietum creticae nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 12 du tableau IV in Barbero & Quézel (1980, Ecol. Medit. 5 : 189) ;
- en colonne 10, le Teucrio divaricati-Iridetum cretensis nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 18 du tableau IV in Barbero & Quézel (1980, Ecol. Medit. 5 : 189).
On peut doter cet ensemble de douze colonnes d’un statut d’alliance caractérisée ou différenciée des unités voisines par Phlomis lanata, Thymbra capitata, Galium fruticosum, Carex distachya, Teucrium divaricatum, T. flavum, Phagnalon graecum, Melissa officinalis subsp. o., Selaginella denticulata, Lamyropsis cynaroides (Asteraceae), Fumana arabica, Satureja thymbra, Aristolochia cretica, Iris unguiculata subsp. cretensis, le Galio fruticosi-Phlomidion lanatae all. nov. hoc loco, typus nominis hoc loco : le Bunio ferulacei-Aristolochietum creticae nov.
Trois belles associations peuvent être mises en évidence dans l’ensemble formé par les colonnes 13 à 16 :
- en 13, l’Origano onitae-Pulicarietum odorae nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 3 du tableau VI in Barbero & Quézel (1980, Ecol. Medit. 5 : 195, en remplaçant Origanum heracleoticum par O. onites ; photo 3) ;
- en 15, le Cyclamini cretici-Ranunculetum paludosi nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 13 du tableau VI in Barbero & Quézel (1980, Ecol. Medit. 5 : 195) ; cette association vivace accompagne une composante annuelle à Lathyrus stenolobus, Geranium purpureum, plus rarement Cynosurus elegans à valeur d’ourlet thérophytique ;
- en 16, l’Hyperico empetrifolii-Oenanthetum pimpinelloidis nov. hoc loco, typus nominis hoc loco: composante herbacée vivace du relevé 23 du tableau VI in Barbero & Quézel (1980, Ecol. Medit. 5 : 195).
Ce petit ensemble est peu caractérisé quoique bien différencié de ses voisins par Oenanthe pimpinelloides, Pteridium aquilinum, Brachypodium sylvaticum, Clinopodium vulgare ; il paraît pour l’instant prématuré de le dénommer.
Il est difficile de classer ces trois unités (dont deux nommées : Phlomido creticae-Crepidion fraasii et Galio fruticosi-Phlomidion lanatae) au niveau supérieur (ordre et classe).

2.6. De Honshu (Japon)
On propose de reprendre ici les données phytosociologiques sur des forêts des monts Chugoku, sud-ouest d’Honshu, au Japon, apportées par Ohno (1982). Dans ses tableaux 3 et 4, il décrit respectivement des « ptérocaryaies » et des frênaies dont la différenciation écologique n’apparaît pas clairement.
En premier lieu, le tableau 8 rassemble l’information sur la végétation arborescente. La colonne 1 peut être définie comme Lindero umbellatae-Pterocaryetum rhoifoliae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé de la colonne 4 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314). La colonne 2 est l’Aceri carpinifolii–Pterocaryetum rhoifoliae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé de la colonne 17 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314). La colonne 3 peut recevoir le nom de Torreyo nuciferae-Zelkovetum serratae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé de la colonne 6 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314), la colonne 4 comme Querco salicinae–Pterocaryetum rhoifoliae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé de la colonne 23 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314), la colonne 6 comme Aesculo turbinatae-Fraxinetum platypodae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé de la colonne 7 du tableau 4 in Ohno (Jap. J. Ecol. 32 : 315-317). Pour tester la place synsystématique de cet ensemble de six colonnes, on va s’appuyer sur une synthèse antérieure des forêts d’Asie nord-orientale (de Foucault, 2023b), dans laquelle trois classes de végétation arborescente avaient été reconnues : les Betulo ermanii-Piceetea jezoensis, les Fago crenatae-Fraxinetea sieboldianae et les Machilo thunbergii-Camellietea japonicae. Il est clair que le présent ensemble relève de la deuxième classe. Le tableau 9 reprend donc les colonnes Td, MsAj, A et B du tableau 11 de la synthèse de 2023b et ajoute en PrAt celle issue du tableau 8 hoc loco. On voit la forte originalité de cette végétation de Honshu : si elle relève bien de la classe citée, elle ne peut relever du seul ordre reconnu jusqu’ici (les Lindero umbellatae-Fagetalia crenatae) ; on doit d’abord y reconnaître la valeur d’une alliance nouvelle richement caractérisée par Aesculus turbinata, Pterocarya rhoifolia, Cornus controversa (photo 4), Fraxinus lanuginosa, F. platypoda, Acer carpinifolium, A. argutum, Pterostyrax hispidus (Styracaceae), Zelkova serrata (Ulmaceae ; photo 5), Cryptomeria japonica, Ulmus laciniata, Stachyurus praecox (Stachyuraceae), Cercidiphyllum japonicum (Cercidiphyllaceae), Styrax obassia (photo 6), Torreya nucifera (Taxaceae), Lindera sericea subsp. glabrata (Lauraceae), Meliosma tenuis (Sabiaceae)…, le Pterocaryo rhoifoliae-Aesculion turbinatae all. nov. hoc loco, typus nominis hoc loco : l’Aesculo turbinatae-Fraxinetum platypodae nov. Par suite on peut définir un second ordre dans la classe, les Pterocaryo rhoifoliae-Aesculetalia turbinatae ord. nov. hoc loco, ordre caractérisé par les mêmes taxons que son typus nominis hoc loco, le Pterocaryo rhoifoliae-Aesculion turbinatae nov.



Le tableau 10 est dédié à la végétation arbustive intraforestière de ces boisements. Deux associations se dégagent :
- les colonnes 1 à 4 décrivent diverses variations du Cephalotaxo nanae-Hydrangeetum petiolaris nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé de la colonne 6 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314, en remplaçant Cephalotaxus harringtonia var. nana par C. nana) ;
- les colonnes 5 et 6 correspondent à deux variations de l’Hydrangeo acuminatae-Euonymetum oxyphylli nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé de la colonne 1 du tableau 4 in Ohno (Jap. J. Ecol. 32 : 315-317).
On renonce à placer ces deux associations dans le synsystème actuellement connu des fourrés est-asiatiques, tout en reconnaissant leur rapprochement dans une alliance caractérisée par Alangium platanifolium (Cornaceae), Toxicodendron orientale subsp. o., Deutzia crenata (Hydrangeaceae), Euonymus alatus, Hydrangea acuminata (Hydrangeaceae), Ligustrum tschonoskii, Sambucus sieboldiana, Viburnum plicatum subsp. p., Helwingia japonica (Helwingiaceae), Weigela hortensis (Caprifoliaceae), le Ligustro tschonoskii-Hydrangeion acuminatae all. nov. hoc loco, typus nominis hoc loco : le Cephalotaxo nanae-Hydrangeetum petiolaris nov.
Dans le tableau 11, rassemblant la végétation vivace intraforestière de ces boisements, deux ensembles se séparent, d’ailleurs selon une logique différente de celle des végétations arborescente et arbustive. Les colonnes 1 à 4 caractérisées par le groupe sociologique à Persicaria filiformis se séparent en
- Hydrangeo alternifoliae-Laporteetum cuspidatae ass. nov. hoc loco (colonne 1), typus nominis hoc loco: composante herbacée vivace du relevé de la colonne 4 du tableau 3 in Ohno ( J. Ecol. 32 : 311-314, en remplaçant Cardiandra alternifolia par Hydrangea a. et Laportea macrostachya par L. cuspidata) ;
- Symplocarpo renifolii-Dryopteridetum crassirhizomae ass. nov. hoc loco (colonne 3), typus nominis hoc loco: composante herbacée vivace du relevé de la colonne 6 du tableau 4 in Ohno ( J. Ecol. 32 : 315-317) ;
- Cardiocrino cordati-Cirsietum buergeri ass. nov. hoc loco (colonne 4), typus nominis hoc loco: composante herbacée vivace du relevé de la colonne 12 du tableau 3 in Ohno ( J. Ecol. 32 : 311-314, en remplaçant Lilium cordatum par Cardiocrinum c.).
Les colonnes 5 et 6 sont moins nettement caractérisées, on peut au moins distinguer la colonne 6 comme Carici glaberrimae-Rubetum palmati ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé de la colonne 23 du tableau 3 in Ohno (Jap. J. Ecol. 32 : 311-314). Sans pouvoir rattacher ces syntaxons à des unités supérieures bien définies, on peut au moins attribuer à l’ensemble des colonnes 1 à 4 la valeur d’une alliance caractérisée par Persicaria filiformis, Laportea cuspidata, Elatostema laetevirens, E. involucratum (deux genres d’Urticaceae), Angelica pubescens, Viola vaginata, Sanicula chinensis, Galium trifloriforme, G. japonicum, Spuriopimpinella brachycarpa (Apiaceae), Asimitellaria pauciflora (Saxifragaceae), Petasites japonicus, Oxalis griffithii, Carex foliosissima, C. dolichostachya subsp. glaberrima, ainsi que des fougères (Polystichum retrosopaleaceum, P. tripteron, Arachniodes standishii, Thelypteris mollissima), le Laporteo cuspidatae-Persicarion filiformis all. nov. hoc loco, typus nominis hoc loco : le Symplocarpo renifolii-Dryopteridetum crassirhizomae nov. Par ailleurs, remarquons la bonne représentation du genre Chrysosplenium avec Ch. macrostemon, Ch. kiotense, Ch. japonicum, Ch. album, qui laisse présager l’existence de végétations de sources (« crénophiles ») à décrire.
Il existe enfin quelques éléments d’une végétation herbacée annuelle à valeur d’ourlet thérophytique sciaphile, caractérisée par Laportea bulbifera, Impatiens textorii, I. noli-tangere, Persicaria thunbergii, P. debilis, l’Impatiento noli-tangere-Laporteetum bulbiferae ass. nov. hoc loco, typus nominis hoc loco : composante herbacée annuelle du relevé de la colonne 1 du tableau 4 in Ohno (Jap. J. Ecol. 32 : 315-317), apparaissant vicariant d’autres ourlets à I. noli-tangere tels que le Persicario hydropiperis-Impatientetum noli-tangere B. Foucault 2011 (photo 2), le Cardamino flexuosae-Impatientetum noli-tangere (Gruber 1978) B. Foucault 2020, le Lapsano grandiflorae-Impatientetum noli-tangere B. Foucault 2020, le Geranio robertiani-Impatientetum noli-tangere B. Foucault 2023.
3. Un fourré alluvial
En 1991, j’ai décrit le Salici incanae-Cornetum sanguineae, muté ici en Salici eleagni-Cornetum sanguineae nom. mut. hoc loco, des zones alluviales de niveau topographique moyen en zones méditerranéennes. Une lecture récente m’a montré que Nègre (1984 : 181) en avait déjà publié un relevé de la vallée de l’Ouvèze, affluent du Rhône, non loin du mont Ventoux.
4. Quelques associations chasmophytiques
Dans la synthèse des Parietarietea judaicae Rivas-Mart. in Rivas Goday 1964 est cité le Cymbalario muralis-Trachelietum caerulei Rivas Mart. 1969 décrit de la péninsule Ibérique, puis reconnu en Italie ; on vient de l’observer dans le jardin des plantes de Montpellier, quoiqu’il soit difficile de préciser si Trachelium caeruleum (photo 7) provient des introductions au sein du jardin ou est arrivé seul :
Montpellier (34), muret du jardin des plantes, N 43° 36’ 51,3’’, E 3° 52’ 21’’, 36 m, 30 %, 10 m², le 3 juin 2025, 7 taxons, photo 8
- Parietarietea judaicae: Trachelium caeruleum 2, Parietaria judaica 2, Cymbalaria muralis 1, Centranthus ruber 1, Asplenium trichomanes *quadrivalens +,
- autres taxons : Plantago lanceolata +, Hedera helix


Toujours dans la région de Montpellier, à Castelnau-le-Lez, entre la clinique du Parc et le parc Vincent-Sablé (N 43° 38’ 16,4’’, E 3° 53’ 35,6’’, 33 m, le 3 septembre 2025), le long du Lez, sur des murets anthropiques soumis à une humidité atmosphérique suffisante (bord des cours d’eau, murs suintants), on peut aussi observer l’Adianto capilli-veneris-Parietarietum judaicae Segal 1969 (photo 9).

L’Asplenietum quadrivalenti-scolopendrii B. Foucault ex B. Foucault 2018 est une association chasmophytique riche en fougères liée à des ambiances climatiques océaniques (de Foucault, 1995, 2008) :
Saint-Valéry-sur-Somme (80), muret de l’hôtel du Cap-Hornu, N 50° 11’ 25,3’’, E 1° 26’ 41,4’’, 5 m, exp. nord, 30 %, 10 m², le 22 juin 2025, 10 taxons, photo 10
- Asplenietea trichomanis: Asplenium scolopendrium 3, adiantum-nigrum *a. 2, A. trichomanes *quadrivalens 2, Dryopteris filix-mas 2, Asplenium ruta-muraria var. r. +, Geranium robertianum +,
- autres taxons : Convolvulus arvensis 1, Festuca rubra 1, Trifolium repens+, Epilobium +.

5. Des prés halophiles du Québec
En annexe d’une conséquente étude de forêts du Québec, Blouin et Grandtner (1971) apportent quelques données sur la végétation herbacée halophile de la rive sud du fleuve Saint-Laurent. Le tableau 12 en synthétise l’essentiel en distinguant la végétation vivace, dominante, et la végétation annuelle, ponctuelle, qui ne sera pas discutée ici, renvoyant à de Foucault & Guitton (2021) à propos du Salicornion grandtneri Géhu & Géhu-Franck 1992, dont le statut reste problématique.
Pour la végétation vivace, deux associations sont bien séparées :
- celle de la colonne 1 rattachée par les auteurs au Spartinetum alterniflorae Dansereau 1959 (en fait le Spartinetum alterniflorae Conard 1935, prioritaire, muté ici en Sporoboletum alterniflori Conard 1935 mut. hoc loco), mais qui, compte tenu de l’importance du GS à Lysimachia maritima, s’avère plus une association des Puccinellietalia phryganodis que des Sporoboletalia alterniflori Conard 1935 nom. mut. hoc loco; au sein des Puccinellietalia phryganodis, c’est du Limonio carolinianae–Puccinellion phryganodis B. Foucault 2021 (de Foucault, 2021c, tableau 4 : colonne LPp) qu’elle se rapproche le plus malgré l’absence du second taxon éponyme d’alliance ; elle est bien distincte de la seule association connue jusqu’à maintenant, le Limonio caroliniani-Puccinellietum phryganodis (Grandtner 1984) B. Foucault 2021, ce qui justifie le nouveau Sporobolo alterniflori-Plantaginetum maritimae ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 5731 du tableau 150 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte, en remplaçant Spartina alterniflora par Sporobolus alterniflorus et Plantago juncoides par P. maritima subsp. m.) ;
- celle de la colonne 2, notamment caractérisée par Halerpestes cymbalaria (= Ranunculus c. ; figure 1) et dénommée Festucetum rubrae par les auteurs, un nom illégitime redéfini ici comme Sporobolo michauxiani-Lysimachietum maritimae (Blouin & Grandtner 1971) nov. hoc loco, typus nominis hoc loco: composante vivace du relevé 5741 du tableau 150 in Blouin & Grandtner (1971, Mémoires du ministère des Terres et des Forêts du Québec 6 hors texte, en remplaçant Spartina pectinata par Sporobolus michauxianus et Glaux maritima par Lysimachia m.), toujours à rattacher au Limonio caroliniani-Puccinellion phryganodis.
6. Des pelouses thérophytiques
Dans un des tout derniers Carnets botaniques (de Foucault, 2025b) j’avais discuté du statut syntaxonomique du Lindernio dubiae-Limoselletum subulatae décrit par Béguin et al. (1995) du Québec. Je n’avais alors pas en tête le fait que ce nom s’avère illégitime face au Limosello subulatae-Cyperetum rivularis Géhu & Géhu-Franck 1985 (Colloq. Phytosoc. XII, séminaire Mégaphorbaies : 170), muté ici en Limosello subulatae-Cyperetum bipartiti Géhu & Géhu-Franck 1985 nom. mut. hoc loco, l’identité entre ces deux noms étant montrée dans le tableau 13. Cette association présente une variation significative plus eutrophile (relevés 7 à 12) différenciée par des Bidens, surtout Bidens hyperborea. Le petit problème est que le type de l’association et donc du typicum appartient justement à cette variation à Bidens (colonne 12 du tableau 13), de sorte qu’il est difficile de nommer l’autre variation dans laquelle il n’y a guère de taxons vraiment différentiels (variation inops). Quoi qu’il en soit, cette variation typicum incite à poser l’hypothèse d’une association encore plus eutrophile qui serait caractérisée par B. hyperborea et ne semblant pas encore décrite.
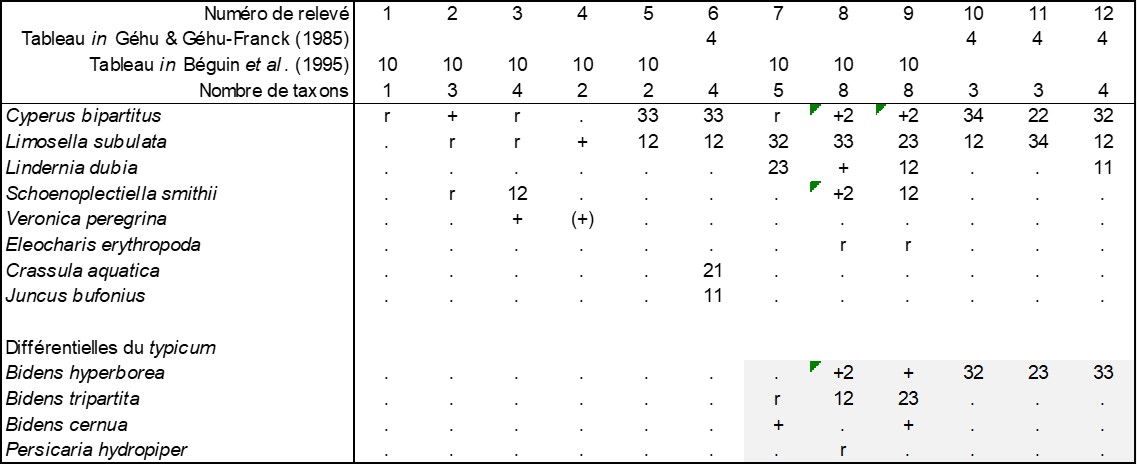
On peut reconnaître cette potentielle association à B. hyperborea dans des données de Lacoursière & Grandtner (1972, tableaux V : relevés 1 à 8 et 10, VIII : relevés 14 à 36 et 41, XXIV : relevés 2 à 7, soit 39 relevés) : Bidens hyperborea V, Zizania aquatica (Poaceae ; figure 2) V, Eleocharis geniculata II, Persicaria hydropiper I, Amphicarpaea bracteata (Fabaceae) +, Cyperus diandrus +, Limosella subulata r, Lindernia dubia r, Bidens eatonii r, soit le Zizanio aquaticae-Bidentetum hyperboreae ass. nov. hoc loco, association thérophytique eutrophile commensale de diverses roselières vivaces nord-américaines, où les taxons caractéristiques du Limosello subulatae-Cyperetum bipartiti sont très peu présents, typus nominis hoc loco : composante annuelle du relevé 1 du tableau V in Lacoursière & Grandtner (1972, Nat. Can. 99 (5) : 477).

On profite de cette précision pour reconnaître qu’une autre association thérophytique amphibie, le Gnaphalio uliginosi-Corrigioletum littoralis, décrit de Montagne Noire (de Foucault, 2023a), à une altitude moyenne de 720 m, peut inclure le petit groupement reconnu sur un relevé en Lozère (Le Malzieu-Ville, 48, vers 850 m ; de Foucault, 1987 : 339).
On peut achever cette partie en évoquant un groupement thérophytique plutôt mésophile (photo 11), qui pourrait se rattacher au Digitario sanguinalis-Polygonion avicularis B. Foucault 2010 au sein de la classe des Polygono-Poetea annuae (de Foucault, 2010a) :
Saint-André-de-la-Marche (49), la Petite Félixière, N 47° 05’ 12,3’’, E 01° 00’ 12,9’’, 101 m, le 3 août 2025, 40 %, 10 m², 10 taxons
- Polygono-Poetea annuae: Portulaca oleracea 3, Digitaria sanguinalis 2, Polygonum aviculare 2, Poa annua +,
- autres taxons : Chenopodium album 1, Capsella bursa-pastoris +, Erigeron canadensis +, Echinochloa crus-galli +, Erodium cicutarium *c. +, Amaranthus albus +,
à rapprocher de l’Eragrostio minoris-Polygonetum arenastri Oberd. 1954 corr. Mucina in Mucina et al. 1993. Cette alliance paraît constituer la limite septentrionale des Sagino apetalae-Polycarpetalia tetraphylli B. Foucault 2010 et tend à remonter vers le nord sous l’effet des changements climatiques (de Foucault, 2010b ; Matysiak, 2022).

7. Des bas-marais
7.1. Un Caricetum nigrae nord-américain
Du Québec, Doyon (1968, tableau IV) décrit un Caricetum nigrae, évidemment homonyme illégitime de divers Cariceta nigrae européens et plus anciens, correspondant à la liste suivante (sur huit relevés) :
- combinaison caractéristique : Carex nigra V, Lycopus uniflorus IV, Viola palustris IV, Carex crawfordii IV, echinata *e. III, Hydrocotyle americana III, Lycopus americanus II, Juncus filiformis II, Epilobium ciliatum *glandulosum II, E. leptophyllum II, Agrostis scabra II, Carex leptalea I, C. canescens I,
- autres taxons : Equisetum arvense III, Juncus tenuis III, Agrostis stolonifera II, Scirpus atrocinctus II, Glyceria canadensis II, Fragaria virginiana *v. II, Euthamia graminifolia I, plus des taxons naturalisés d’Europe et des taxons de présence I ;
ce bas-marais québécois peut être nouvellement décrit comme Caricetum crawfordii-nigrae (Doyon 1968) ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau IV in Doyon (1968, Nat. Can. 95 : 384).
7.2. Des bas-marais du Kamchatka
Neshatayeva et Neshatayev (2001) ont apporté des données sur des bas-marais du sud de la péninsule du Kamchatka (aussi écrit Kamtchatka) en utilisant une nomenclature incorrecte propre à une école russe de phytosociologie.
Il existe tout d’abord une association arborescente connue par trois relevés des auteurs (leur tableau 1 : relevés 13 à 15) : Salix pulchra *parallelinervis 3, Alnus hirsuta 2, Betula ermanii *e. 1, soit l’Alno hirsutae-Salicetum parallelinervis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 13 du tableau 1 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 61-62), que l’on peut rapprocher des Betulo ermanii-Piceetalia jezoensis B. Foucault 2023 (de Foucault, 2023b).
Il existe ensuite une association arbustive à Myrica tomentosa (d’après Neshatayeva & Neshatayev, 2001, tableau 2, soit 18 relevés) : Betula glandulosa V, Salix fuscescens V, Myrica tomentosa III, Dasiphora fruticosa II, Salix pulchra *p. j II, décrivant le Salici fuscescentis-Betuletum glandulosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 18 du tableau 2 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 64-66, en changeant Betula exilis en B. glandulosa). Les colonnes 13 à 15 du tableau 1 des auteurs russes décrivent une seconde association arbustive : Dasiphora fruticosa 3, Myrica tomentosa 2, Lonicera chamissoi 2, Spiraea betulifolia *aemiliana 1, le Lonicero chamissoi-Myricetum tomentosae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 14 du tableau 1 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 61-62).
La végétation herbacée de ces marais est reprise ici dans le tableau 14. Deux ensembles se partagent les onze colonnes de ce dernier, qui possèdent en commun Comarum palustre, Equisetum palustre, Carex lyngbyei, C. rariflora, C. aquatilis subsp. minor.
Les colonnes 1 à 7 forment un ensemble bien caractérisé par le groupe sociologique à Sanguisorba tenuifolia et correspondant à des marais sur substrat plutôt ferme. Trois associations suffisamment définies au plan statistique se dégagent :
- en colonne 2, le Trichophoro alpini-Caricetum chordorrhizae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 4 du tableau 2 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 64-66) ;
- en colonne 3, le Luzulo confusae-Sanguisorbetum tenuifoliae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 11 du tableau 2 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 64-66) ;
- en colonne 6, le Rubo chamaemori-Empetretum stenopetali ass. nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 8 du tableau 1 in Neshatayeva & Neshatayev (2001, of Russia 2 : 61-62).
Ce large ensemble caractérisé par Sanguisorba tenuifolia, Calamagrostis neglecta, Andromeda polifolia subsp. p., Bistorta vivipara, Empetrum nigrum subsp. stenopetalum, Lysimachia europaea, Rubus arcticus, Trichophorum alpinum, Vaccinium uliginosum, V. microcarpum, Iris setosa, Luzula confusa, Viola hultenii, Gentianella auriculata, Pedicularis resupinata possède la valeur d’une alliance, le Sanguisorbo tenuifoliae-Calamagrostion neglectae all. nov. hoc loco, typus nominis hoc loco : le Luzulo confusae-Sanguisorbetum tenuifoliae nov.
Les quatre dernières colonnes (8 à 11) décrivent plutôt des marais flottants à Menyanthes trifoliata :
- les colonnes 8 et 9 forment le Caricetum rarifloro-lyngbyei nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 10 du tableau 3 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 67, en remplaçant Carex cryptocarpa par C. lyngbyei) ;
- la colonne 11 est l’Utriculario macrorhizae-Menyanthetum trifoliatae nov. hoc loco, typus nominis hoc loco: composante herbacée du relevé 19 du tableau 3 in Neshatayeva & Neshatayev (2001, Veg. of Russia 2 : 67).
Ce petit ensemble de marais flottants mérite le rang d’alliance, étant caractérisé par Menyanthes trifoliata, Comarum palustre, Utricularia macrorhiza, Sparganium hyperboreum, Carex lasiocarpa, C. lyngbyei, C. rariflora, C. aquatilis subsp. minor, l’Utriculario macrorhizae-Caricion lyngbyei all. nov. hoc loco, typus nominis hoc loco : le Caricetum rarifloro-lyngbyei nov., alliance vicariante nord-est-asiatique du Caricion lasiocarpae Vanden Berghen in Lebrun et al. 1949 européen.
7.3. La végétation à Rhynchospora alba
Dans ses recherches sur les tourbières bombées du Canada, Damman (1977, tableau 2) décrit un Rhynchosporetum albae attribué à W. Koch 1926. Mais le syntaxon qu’il décrit est bien sûr assez différent de celui d’Europe centrale auquel il renvoie ; il est synthétisé dans la colonne 1 du tableau 15 hoc loco. On peut mettre en exergue ce groupement nord-américain comme Utriculario cornutae-Rhynchosporetum albae (Damman 1977) ass. nov. hoc loco, typus nominis hoc loco : relevé 75 du tableau 2 in Damman (1977, Vegetatio 35 (3) : 141). Dans son tableau 3-10, Wells (1976) en présente incidemment quatre relevés.
Ce dernier auteur présente par ailleurs dans son tableau 3-8 un groupement aquatique vivace : Nuphar variegata V, Eriocaulon aquaticum (Eriocaulaceae) III, Myrica gale j II et de présence I Chamaedaphne calyculata, Andromeda polifolia *latifolia, Utricularia intermedia, Carex limosa, Drosera intermedia…, l’Eriocaulo aquatici-Nupharetum variegatae ass. nov. hoc loco, typus nominis hoc loco : relevé 164-A du tableau 3-8 in Wells (1976, A classification of peatlands in eastern Newfoundland : 114) ; un groupement à N. variegata est aussi évoqué par Gauthier & Grandtner (1975) du Québec, toutefois pratiquement réduit à l’espèce éponyme, et par Muller & Etcheberry de Saint-Pierre-et-Miquelon (in Muller, 2006). Mais surtout, dans son tableau 3-9 (relevés 54-A à 65-E, soit six relevés), Wells décrit aussi une végétation de gouille à Sphagnum torreyanum reprise dans la colonne 2 du tableau 15 hoc loco, une association originale en lien dynamique avec une tourbière bombée du Kalmio-Sphagnion fusci Tüxen et al. 1972 (Tüxen et al., 1972), le Sphagno torreyani-Rhynchosporetum albae ass. nov. hoc loco, typus nominis hoc loco : relevé 163-B du tableau 3-9 in Wells (1976, A classification of peatlands in eastern Newfoundland : 118).
Plus globalement, le tableau 15 reprend en outre les syntaxons connus à Rhynchospora alba à travers l’hémisphère nord. Les colonnes 3 à 8, 13 et 14 sont reprises de ma thèse (de Foucault, 1984) avec le Rhynchosporion albae W. Koch 1926 s. l. (colonnes 3 à 8) [syn. : Molinio caeruleae-Rhynchosporion albae (W. Koch 1926) B. Foucault 1984 nom. ined. et illeg. repris par Julve (1993, Lejeunia, n. s., 140 : 108)], le Lycopodiello inundatae-Rhynchosporetum albae P. Allorge & Gaume ex Schaminée et al. in Schaminée et al. 1995 (colonne 9), le Drosero intermediae-Schoenetum nigricantis Braun-Blanq. & Tüxen 1952 (colonne 10), le Drosero intermediae-Rhynchosporetum albae (P. Allorge 1941) Fern. Prieto et al. 1987 (colonne 11), l’Eleocharito multicaulis-Rhynchosporetum albae C. Valle & F. Navarro ex Rivas Mart. et al. 2002 (colonne 12), le Moliniopsio japonicae-Rhynchosporion albae Tüxen, Suzuki & Fujiwara 1970 (colonne 13) et l’Eriocaulo–Rhynchosporion fujiianae Fujiwara 1979 (Bull. Yokohama Phytosoc. Soc. 16 : 329) nom. inval. (art. 3g) (colonne 14) validé ici en Rhynchosporion fauriei-fujiianae Fujiwara ex all. nov. hoc loco, caractérisé par Rhynchospora fujiiana, Rh. fauriei, Moliniopsis japonica, Sphagnum microporum, typus nominis hoc loco : le Rhynchosporo fauriei-Sphagnetum palustris Fujiwara 1979 (Bull. Yokohama Phytosoc. Soc. 16 : 329). Les colonnes 1 et 2 correspondent aux deux associations nord-américaines qui doivent alors se rattacher à une alliance originale vicariante des précédentes, caractérisée par des taxons issus des tourbières ombrotrophes du Kalmio-Sphagnion fusci : Kalmia polifolia, Andromeda polifolia var. latifolia, Vaccinium oxycoccos, V. macrocarpon, Chamaedaphne calyculata, Trichophorum cespitosum subsp. c., Sarracenia purpurea, le Kalmio polifoliae-Rhynchosporion albae all. nov. hoc loco, typus nominis hoc loco : l’Utriculario cornutae-Rhynchosporetum albae (Damman 1977) nov. L’ensemble des colonnes 3 à 12, correspondant au Rhynchosporion albae s. l. et partageant Rhynchospora fusca, Molinia caerulea, Carex panicea, Eriophorum angustifolium subsp. a., est assez bien différencié en
- un ensemble formé des colonnes 3 à 6, plutôt continental-montagnard et identifiable au strict Rhynchoporion albaeKoch 1926 (Jahrb. St. Gall. Naturw. Ges. 61 : 92), incl. Scheuchzerion palustris Nordh. 1937 ;
- un ensemble formé des colonnes 7 à 12, plutôt atlantique, étant bien différencié des alliances voisines par Erica tetralix et des taxons caractérisant par ailleurs les Eleocharitetalia multicaulis et le Juncion acutiflori, surtout Eleocharis multicaulis, Narthecium ossifragum, Pinguicula lusitanica, Juncus acutiflorus, Carex demissa, Lysimachia tenella, l’Eleocharito multicaulis-Rhynchosporion albae nov. hoc loco, typus nominis hoc loco: l’Anagallido tenellae-Rhynchosporetum albae Chouard 1924 (Bull. Soc. Bot. France 71 : 1151), muté en Lysimachio tenellae-Rhynchosporetum albae nom. mut. hoc loco, prioritaire sur le Rhynchosporetum albae P. Allorge & Denis ex P. Allorge 1926 (Bull. Mayenne-Sci. : 19, plusieurs relevés conformes à l’art. 2b de l’ICPN ; après vérification dans cet article, le nom souvent employé ultérieurement de « Drosero intermediae-Rhynchosporetum albae » en est bien absent), incl. le Rhynchosporetum albae sensu Diemont & Tüxen in Tüxen 1937 du nord-ouest de l’Allemagne (Tüxen, 1937). On peut encore rattacher à cette alliance atlantique
- le Drosero intermediae-Rhynchosporetum albae (P. Allorge 1941) Fern. Prieto et al. 1987 (colonne 11), difficilement identifiable au Lysimachio tenellae-Rhynchoporetum albae Chouard 1924 avec Erica mackayana qui y remplace tetralix; comme cela vient d’être dit, il faut préciser que le terme de Drosero intermediae-Rhynchosporetum albae attribué à Allorge n’existe pas dans l’article de 1926, il n’y a donc pas homonymie ;
- l’Eleocharito multicaulis-Rhynchosporetum albae Tüxen ex Rivas Mart. in Rivas Mart. et al. 2002 [colonne 12 ; syn. : Eleocharito multicaulis-Rhynchosporetum albae Tüxen in Tüxen & Oberd. 1958 (Veröff. Geobot. Inst. Rübel in Zürich 32 (2) : 165) inval.].
Ces cinq alliances partagent Rhynchospora alba, des Drosera…, ce qui justifie l’ordre des Scheuchzerietala palustris Nordh. 1937. Il existe des associations de transition entre Rhynchosporion albae et Eleocharito-Rhynchosporion albae ; c’est par exemple le cas du « Rhynchosporetum albae » décrit de la haute Ardenne belge (Schumacker, 1980, tableau 2) sous climat atlantique montagnard. Conformément aux positions de Tüxen (1980) et Julve (1993), cet ordre est mieux placé dans les Vaccinio oxycocci-Sphagnetea magellanici que dans la classe des bas-marais.
Aux associations du Rhynchosporion fauriei-fujiianae décrites par Fujiwara (1979), il faut ajouter celle formée par les cinq relevés (6 à 10) du tableau 5 in Nishimoto (2006), le Rhynchosporetum fabrei-fujiianae ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 8 du tableau 5 in Nishimoto (2006, Bull. Okayama Pref. Nature Conserv. Center 14 : 30, en changeant Rhynchospora faberi en Rh. fabrei). Il existe en effet une communauté thérophytique associée : Dimeria ornithopoda (Poaceae) V, Eriocaulon miquelianum *m. (Eriocaulaceae) V, E. decemflorum III, Utricularia uliginosa III, Hypericum japonicum I, Eriocaulo miqueliani-Dimerietum ornithopodae ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 8 du tableau 5 in Nishimoto (2006, Bull. Okayama Pref. Nature Conserv. Center 14 : 30, en changeant Eriocaulon sikokianum en E. miquelianum). L’association vivace est présente aussi dans les tableaux 7 (relevés 12 à 17) et 9 (relevés 25 et 26) de cet auteur japonais. Les relevés 18 à 23 de ce même tableau 7 décrivent une autre association du Rhynchosporion fauriei-fujiianae, le Cirsio sieboldii-Rhynchosporetum fauriei ass. nov. hoc loco, typus nominis hoc loco : composante vivace du relevé 21 du tableau 7 in Nishimoto (2006, Bull. Okayama Pref. Nature Conserv. Center 14 : 35). Dans leur tableau 1, en colonnes J-a à J-b, Ohta et Hada (2012) citent encore des végétations à Rhynchospora fabrei, Rh. fujiiana, Rh. rugosa, Drosera rotundifolia, Utricularia caerulea, U. bifida qui doivent aussi y être incluses ; il peut s’y associer une composante thérophytique à Eriocaulon miquelianum *m., Schoenus apogon et Persicaria thunbergii ; ces unités ne peuvent cependant être mieux décrites, les données sources étant synthétiques.
Bibliographie
Allorge P., 1926. Remarques sur quelques associations du massif de Multonne. Bulletin de Mayenne-Sciences : 27-48.
Barbero M., Chalabi N., Nahal I. & Quézel P., 1976. Les formations à conifères méditerranéens en Syrie littorale. Ecologia Mediterranea 2 : 87-99.
Barbero M. & Quézel P., 1980. La végétation forestière de Crète. Ecologia Mediterranea 5 : 175-210.
Béguin C., Grandtner M.M. & Gervais C., 1995. Groupements végétaux côtiers du Saint-Laurent près de Cap-Rouge, Québec (Canada). Documents phytosociologiques, n. s., XV : 183-203.
Blouin J.-L. & Grandtner M.M., 1971. Étude écologique et cartographie de la végétation du comté de Rivière-du-Loup. Mémoires du ministère des Terres et des Forêts du Québec 6 : 1-371 et annexes.
Bodeux A., 1955. Alnetum glutinosae. Mitteilungen der Floristisch-Soziologischen Arbeitsgemeinschaft 5 : 114-137.
Braun-Blanquet J. & Tüxen R., 1952. Irische Pflanzengesellschaften. Veröffentlichungen der Geobotanischen Institutes Rübel in Zürich 25 : 1-421.
Chouard P., 1924. Monographies phytosociologiques, I – La région de Brigueil-l’Ainé (Confolentais). Bulletin de la Société botanique de France 71 : 1130-1158.
Damman A.W.H., 1977. Geographical changes in the vegetation pattern of raised bogs in the bay of Fundy region of Maine and New Brunswick. Vegetatio 35 (3) : 137-151.
Decocq G., 1999.Données phytosociologiques récentes sur les forêts de la Thiérache (Aisne, France) ; III – Les forêts hygrophiles. Acta Botanica Gallica 146 (3) : 187-205.
Döring U., 1987. Zur Freinstruktur amphibischer Erlenbruchwälder; Kleinstandörtliche Differenzierung in der Bodenvegetation des Carici elongatae-Alnetum im Hannoverschen Wendland. Tuexenia 7 : 347-366.
Doyon D., 1968. La végétation des pâturages naturels de Saint-Ferréol et de Saint-Tite-des-Caps, comté de Montmorency. Le Naturaliste canadien 95 (2) : 367-391.
Fajmonová E., 1978. K syntaxonómii spoločenstiev radu Athyrio-Piceetalia Hadač 62 v Západných Karpatoch. Biológia (Bratislava) 33 (7) : 551-563 (en tchèque).
Fajmonová E., 1986. K variabilite asociácie Cortuso-Piceetum na Slovensku. Preslia 58 : 43-54 (en tchèque).
Fernández Prieto J.A., Fernández Ordóñez M.C. & Collado Prieto M.A., 1987. Datos sobre la vegetación de las « turberas de esfagnos » galaico-asturianas y orocantábricas. Lazaroa 7 : 443-471.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’État, université de Rouen, 675 p.
Foucault B. (de), 1987. Données phytosociologiques sur la végétation observée lors de la treizième session de la S.B.C.O. en Aubrac et Margeride. Bulletin de la Société botanique du Centre-Ouest, n. s., 18 : 337-361.
Foucault B. (de), 1988. Phytosociologie et dynamique des landes de Lessay (Manche), plus particulièrement de la réserve de Mathon. Botanica Rhedonica, n. s., biologie végétale, 1 : 37-70.
Foucault B. (de), 1991. Introduction à une systémique des végétations arbustives. Documents phytosociologiques,
n. s., XIII : 63-104.
Foucault B. (de), 1994. Compte rendu de l’excursion de la S.B.N.F. en forêt d’Andigny (Aisne). Contribution à la connaissance phytosociologique des forêts du Nord. Bulletin de la Société de botanique du nord de la France 47 (2) : 5-20.
Foucault B. (de), 1995. Contribution à une monographie phytosociologique de la Hague (Manche, France). Bulletin de la Société botanique du nord de la France 48 (4) : 45-90.
Foucault B. (de), 1997. Étude phytosociologique de la vallée de l’Hogneau et de ses versants (département du Nord, France) (ZNIEFF 87-0, type I). Natura Mosana 50 (3) : 61-88.
Foucault B. (de), 2010a. Contribution au prodrome des végétations de France : les Polygono-Poetea annuae Rivas-Mart. 1975 corr. Rivas-Mart. et al. 1991. Journal de botanique de la Société botanique de France 49 : 55-72.
Foucault B. (de), 2010b. Note sur une association végétale des Polygono-Poetea annuae inattendue dans le département du Nord. Bulletin de la Société de botanique du nord de la France 63 (2-4) : 15-16.
Foucault B. (de), 2011. Synthèse phytosociologique sur la végétation observée lors de la 146e session de la SBF dans les Ardennes. Bulletin de la Société d’histoire naturelle des Ardennes 101 : 33-50.
Foucault B. (de), 2014. Contribution au prodrome des végétations de France : les Parietarietea judaicae Rivas-Mart. in Rivas Goday 1964. Acta Bot. Gallica: Botany Letters 161 (4) : 403-427.
Foucault B. (de), 2018a. Nouvelles validations de syntaxons invalides ou inédits. Journal de botanique de la Société botanique de la Société botanique de France 81 : 35-38.
Foucault B. (de), 2018b. Contribution au prodrome des végétations de France : les Montio-Cardaminetea amarae Braun-Blanq. & Tüxen ex Klika & Hadač 1944. Documents phytosociologiques, 3e série, 7 : 1-86.
Foucault B. (de), 2019. Contribution à la connaissance phytosociologique de la végétation du pays de Sault (Aude, France). Evaxiana 6 : 29-129.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021a. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2021b. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de), 2021c. Contribution à une synthèse des Juncetea maritimi. Carnets botaniques 68 : 1-30 + 22 tableaux, https://doi.org/10.34971/nbat-mh09.
Foucault B. (de), 2022. Nouvelles considérations phytosociologiques sur la végétation de la Grèce. Carnets botaniques 109 : 1-27, https://doi.org/10.34971/Q6KD-W822.
Foucault B. (de), 2023a. Nouvelles données sur la végétation du versant audois de la Montagne Noire. Carnets botaniques 160 : 1-30, https://doi.org/10.34971/HTCT-WD37.
Foucault B. (de), 2023b. Contribution à la connaissance de la végétation forestière de l’Asie nord-orientale selon le paradigme synusial. Carnets botaniques 163 : 1-18, https://doi.org/10.34971/VHBV-Q853.
Foucault B. (de), 2025a. Compte rendu d’un voyage botanique et phytosociologique en Colombie. Carnets botaniques 259 : 1-23, https://doi.org/10.34971/4DEG-J010.
Foucault B. (de), 2025b. Miscellanées phytosociologiques VIII : syntaxons nouveaux, validés ou complétés. Carnets botaniques 264 : 1-13, https://doi.org/10.34971/AVAA-5634.
Foucault B. (de), 2025c. Nouvelles interprétations phytosociologiques de la végétation palestinienne. Carnets botaniques 241 : 1-8, https://doi.org/10.34971/233C-QE98.
Foucault B. (de), 2025d. Après un voyage à Chypre du Nord, nouveautés sur la végétation chypriote. Carnets botaniques 253 : 1-15, https://doi.org/10.34971/Q8PM-0Y84.
Foucault B. (de), Amat J.-P., Wattez J.-R. & Valcke M.-A., 1996. Observations sur la composition floristique actuelle des forêts de la région d’Albert (Somme), 80 ans après les combats. Mémoires de la Société linnéenne Nord-Picardie III, actes du colloque L’impact de la guerre sur la végétation et la régénération du tapis végétal, Albert, juin 1996 : 15-18.
Foucault B. (de) & Guitton H., 2021. Une déclinaison moderne des Thero-Suaedetea splendentis. Carnets botaniques 76 : 1-9 + 2 tableaux, https://doi.org/10.34971/3XK3-N640.
Fujiwara K., 1979. Moor vegetation in Japan with emphasis on Eriocaulo-Rhynchosporion fujiianae. Bulletin of the Yokohama Phytosociological Society 16 : 325-332.
Gallandat J.‑D., Gillet F., Havlicek E. & Perrenoud A., 1995. Typologie et systémique phyto‑écologiques des pâturages boisés du Jura suisse. Université de Neuchâtel, Institut de botanique.
Gauthier R. & Grandtner M.M., 1975. Étude phytosociologique des tourbières du bas Saint-Laurent, Québec. Le Naturaliste canadien 102 : 109-153.
Géhu J.-M. & Géhu-Franck J., 1985. La mégaphorbiaie à Sanguisorba canadensis, élément remarquable de la zonation végétale soumise aux marées d’eau douce des berges du fleuve Saint-Laurent (Québec, Canada). Colloques phytosociologiques XII, séminaire Mégaphorbiaies : 161-173.
Hoda P., 1993. A geobotanical survey of the black pine (Pinus nigra Arn.) forests in Albania. Annali di botanica LI : 77-92.
Julve Ph., 1993. Synopsis phytosociologique de la France (communautés de plantes vasculaires). Lejeunia, n. s., 140 : 1-160.
Julve Ph. & Gillet F., 1994. Conceptions and methods of the individual vegetation maps; III – Experiences of Frenc authors. Phytocenosis, n. s., 6, suppl. Cartographiae Geobotanicae 4 : 45-66.
Koch W., 1926. Die Vegetationseinheiten der Linthebene. Jahrbuch der St. Gallischen Naturwissenschaftlichen Gesellschaft 61 : 1-146.
Kučera P., 2022. Natural calcareous Norway spruce woodlands in Slovakia and their syntaxonomical classification. Hacquetia 21 (1) : 107-151.
Lacoursière E. & Grandtner M.M., 1972. Les groupements végétaux ripariens entre Sainte-Famille et la pointe d’Argentenaye, île d’Orléans, Québec. Le Naturaliste canadien 99 (5) : 469-507.
Loidi J., Estudio de la flora y la vegetación de las cuencas de los rios Deva y Urola en la provincia de Guipúzcoa. Thèse, Universidad Complutense de Madrid, 298 p.
Mast R., 2007. Vegetationsökologische Gliederung der Erlenbruchwälder (Alnion glutinosae) in Deutschland. Berichte der Reinhold-Tüxen-Gesellschaft 19 : 174-186.
Matysiak J.-P., 2022. Et nous basculâmes dans les Sagino apetalae-Polycarpetalia tetraphylli B. Foucault 2010… Bulletin de la Société de botanique du nord de la France 75 : 39-41.
Nègre R., 1984. Dynamisme végétal de l’Ouvèze. Colloques phytosociologiques IX, La végétation des zones alluviales : 171-190.
Neshatayeva V.Y. & Neshatayev V.Y., 2001. Mire vegetation of the southern Kamchatka state nature reserve. Vegetation of Russia (St. Petersburg) 2 : 58-70 (en russe).
Nishimoto T., 2006. Vegetational changes from the last forty years of four moors located in the Hiruzen area, Okayama Prefecture, Utsumitani moor, Simo-utsumitani moor, Orogatawa moor and Higashi moor. Bulletin of the Okayama Prefectural Nature Conservation Center 14 : 15-69 (en japonais).
Ohno K., 1982. A phytosociological study of the Vaijley forests in the Chugoku mountains, southwestern Honshu, Japan. Japanese Journal of Ecology 32 : 303-324.
Ohta K. & Hada Y., 2012. The vegetation on the granite rock area at Ashimori, Okayama City, S.W. Honshu, Japan. Naturalistae 16 : 19-27.
Rivas-Martínez S., Loidi J., Cantó P., Sancho L.G. & Sánchez Mata D., 1984. Datos sobre la vegetación del valle del río Bidasoa (España). Lazaroa 6 : 127-150.
Schumacker R., 1980. Groupements du Caricetum limosae (Paul 1910) Osv. 1923, du Rhynchosporetum albae Koch 1926, du Caricetum lasiocarpae Koch 1926 et à Carex rostrata-Sphagnum apiculatum en haute Ardenne nord-rientale. Colloques phytosociologiques VII, La végétation des sols tourbeux : 461-474.
Tüxen R., 1937. Die Pflanzengesellschaften Nordwestdeutschlands. Mitteilungen der Floristisch-soziologischen Arbeitsgemeinschaft in Niedersachsen 3 : 1-170.
Tüxen R., Miyawaki A. & Fujiwara K., 1972. Eine erweiterte Gliederung der Oxycocco-Sphagnetea. Bericht der internationale Symposion der Internationalen Vereinigung für Vegetationskunde 1970, Grundfragen und Methoden in der Pflanzensoziologie : 500-509.
Tüxen R. & Oberdorfer E., 1958. Eurosibirische Phanerogamen-Gesellschaften Spaniens. Veröffentlichungen der Geobotanischen Institutes Rübel in Zürich 32 (2) : 1-328.
Valle Guttiérrez C.J. & Navarro Andrés F., 1983. Sobre la vegetación y flora turfófila de la Sierra de la Culebra. Lazaroa 5 : 165-171.
Wells E.D., 1976. A classification of peatlands in eastern Newfoundland. Thèse, St. John’s, Newfoundland, 201 p.
Remerciements
Un grand merci à Hermann Guitton (Conservatoire botanique national armoricain de Brest) pour son aide bibliographique.


