
Après un voyage à Chypre du Nord, nouveautés sur la végétation chypriote
Title
After a trip to Northern Cyprus, news about Cypriot vegetation
Résumé
À la suite d’un petit voyage à Chypre du Nord et sur la base d’un article antérieur consacré à la végétation forestière, divers types de végétation sont présentés et illustrés : végétation des murs, pelouses aérohalophiles, pelouse à géophytes vernales, boisements, friche rudérale à thérophytes.
Abstract
Following a short trip to North Cyprus and based on an earlier article devoted to forest vegetation, various types of vegetation are presented and illustrated: wall vegetation, aerohalophilous lawns, lawns with vernal geophytes, woodlands, ruderal wasteland with therophytes.
1. Introduction
Politiquement, l’île est-méditerranéenne de Chypre, au sud-est de la Turquie et à l’ouest du Liban, est bien connue pour sa partition en deux territoires depuis 1974 : une partie sud grecque (Chypre du Sud) et une partie nord turque (Chypre du Nord), la capitale ancienne, Nicosie, étant alors partagée en Lefkosía grecque et Lefkoşa turque. Physiquement, deux massifs montagneux se répartissent de part et d’autre de la Ligne verte séparant les deux territoires, le Troodos à Chypre du Sud (culminant à 1 952 m au mont Olympe) et le Pentadactylos à Chypre du Nord (culminant à 1 024 m au mont Kyparissovouno). Durant le mois de mars 2025, j’ai eu l’occasion de faire quelques observations floristiques et phytosociologiques à Chypre du Nord dont le compte rendu fait l’objet du présent article.
L’étude de la flore chypriote a donné lieu à de nombreux articles dont on trouvera une liste significative dans l’onglet Bibliography de Flora of Cyprus (Hand et al., 2011). On peut notamment citer l’ouvrage fondamental en deux volumes Flora of Cyprus (Meikle, 1977 et 1985) et en français les publications de G. Alziar (dont celle de 1999 sur Chypre du Sud et le compte rendu du voyage de la Société botanique de France édité en 2004), ainsi que celle de Coulot (2000).
La végétation est bien moins connue que la flore ; elle a particulièrement été décrite par Barbero & Quézel (1979) pour la végétation forestière (surtout à Chypre du Sud), puis Costa et al. (1984) et Géhu et al. (1984, 1990) pour la végétation littorale. On présentera ici le résultat de quelques observations réalisées lors du court séjour à Chypre du Nord en 2025. La nomenclature suit Plants of the World online à l’exception de Calicotome villosa nommé ici Cytisus laniger.
2. Quelques végétations ponctuelles
On commencera par présenter quelques végétations ponctuelles et peu documentées. Ainsi la végétation des murs des remparts historiques a été illustrée notamment par Brandes (2020) qui malheureusement n’en présente aucun relevé. Elle est surtout caractérisée par Hyoscyamus aureus (photo 1) et Parietaria judaica et doit se rattacher au Parietario judaicae-Hyoscyamion aurei Brullo & Guarino 1998. Mais Brullo et Guarino ont publié des associations de Crète, Rhodes, Jérusalem et Betlem (Bethléem) et aucune d’elles ne semble pouvoir correspondre à la végétation observée à Chypre du Nord où Capparis orientalis (= C. spinosa subsp. rupestris) et C. aegyptiaca manquent (de Foucault, 2014). En revanche, l’Adianto capilli-veneris-Parietarietum judaicae Segal 1969, des murs humides à suintants anthropisés, a été rencontré à Bellapaïs (photo 2).


La végétation vraiment littorale n’a guère été analysée en 2025, à l’exception d’une pelouse aérohalophile décrite par Géhu et al. (1990), le Frankenio hirsutae-Limonietum cyprii Géhu, M.J. Costa & Uslu 1990 rattaché au Frankenio hirsutae-Limonion cyprii B. Foucault 2023 (de Foucault, 2023) et revu à Alsancak, notamment caractérisé par le Limonium éponyme (photo 3), L. sinuatum (photo 4), Andrachne telephiifolia (photo 5) et Cichorium spinosum (photo 6). Le relevé suivant en offre un nouvel exemple
Alsancak (ouest de Kyrenia), Dumlupınar sokak, N 35° 21’ 02,7’’, E 33° 13’ 37,1’’, 35 %, 40 m², 10 taxons
Cichorium spinosum 2, Limonium cyprium 2, Andrachne telephiifolia 2, Frankenia hirsuta 2, Reichardia picroides 2, Phagnalon rupestre 2, Crithmum maritimum 1, Lotus cytisoides 1, cf. Crocodilium creticum 1, Pistacia lentiscus (anémomorphosé) +.




Dans les ouvertures de cette pelouse aérohalophile vivace, peut se développer une pelouse thérophytique caractérisée par Sagina maritima et Anthemis rigida, une petite Asteraceae à capitules discoïdes jaunes (photo 7) ; elle rappelle des pelouses homologues à A. rigida décrites de Grèce au sein du Sileno sedoidis-Catapodion marini B. Foucault & Bioret 2010 (Géhu et al., 1992 ; de Foucault & Bioret, 2010).

En arrière de ces végétations aérohalophiles, on peut aussi parfois observer une pelouse vernale à géophytes (photo 8) caractérisée par Moraea sisyrinchium (photo 9), Bellevalia nivalis (endémique chyprio-syrienne ; photo 10), Ornithogalum pedicellare (endémique chypriote ; photo 11), Asphodelus ramosus (photo 12). Cette combinaison de taxons rappelle beaucoup la végétation des Charybdido pancratii-Asphodeletea ramosi Biondi in Biondi et al. 2016 surtout étudiée en Méditerranée occidentale et centrale (de Foucault, 2024).





Enfin la végétation rudérale des Dittrichetea viscosae Trinajstić et al. in B. Foucault & Jasprica 2019 et l’ordre des Hyparrhenietalia hirtae Braun-Blanq. et al. 1956, surtout étudiée, elle aussi, en Méditerranée occidentale et centrale, peut s’étendre à l’île de Chypre où elle est caractérisée par Dittrichia viscosa, Verbascum sinuatum, Hyparrhenia hirta, Oloptum miliaceum et Mandragora officinalis (photo 13).
On va aborder maintenant quelques types de végétation plus précis, notamment la végétation forestière sur la base de la publication de Barbero & Quézel (1979).

3. La végétation arborescente
Le tableau 1 dresse une synthèse des associations arborescentes observées à Chypre, d’après Barbero & Quézel (1979) ; pour comparaisons on y a inclus des données sur des associations décrites dans d’autres régions d’Europe.
Les colonnes 1 à 3 peuvent se rattacher à trois variations (type, à Cupressus sempervirens, à Quercus coccifera) d’un unique Pino brutiae-Ceratonietum siliquae ass. nov. hoc loco de l’étage thermoméditerranéen (altitude souvent inférieure à 200 m). Lors de mon séjour dans l’ile, j’ai eu l’occasion d’en faire un relevé :
Kani Akansı caddesi, Alsancak, N 35° 20’ 28,7’’, E 33° 13’ 43,5’’, 51 m, 14/03/2025, 5 taxons (photo 15)
Ceratonia siliqua 3, Pinus brutia (photo 14) 3, Olea europaea 1, Quercus ilex 1, Smilax aspera ! 1 ;
typus nominis hoc loco : le relevé précédent. Localement cette association paraît aussi caractérisée par Acer obtusifolium (photos 16 et 17) absent du tableau et du relevé présentés ici.




On peut comparer cette association à d’autres précédemment décrites (colonnes 4 à 8 du tableau 1) dont les codes sont les suivants : COe pour le Ceratonio siliquae-Oleetum sylvestris B. Foucault 2021 (de Foucault, 2021a : 73), OPh pour l’Oleo sylvestris-Pinetum halepensis B. Foucault 2022 (de Foucault, 2022, tableau 2), BQ1a, BQ1b et BQ2 pour d’autres variations de l’Oleo sylvestris-Pinetum halepensis (de Foucault, 2022, tableau 1 : colonnes BQ1a à BQ2). On peut considérer que toutes ces associations arborescentes peuvent se rattacher à l’Oleo sylvestris-Ceratonion siliquae Braun-Blanq. ex Guin. & Drouineau 1944.
Les colonnes 9 et 10 décrivent plutôt le Pino brutiae-Quercetum cocciferae ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 5 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 21, en remplaçant Quercus calliprinos par Q. coccifera, ici potentiellement arborescent). Les colonnes 11 et 12 correspondent au Querco alnifoliae-Aceretum sempervirentis B. Foucault 2020, la colonne 13 au Querco alnifoliae-Cedretum brevifoliae B. Foucault 2020. Ces trois associations s’intègrent bien au Querco cocciferae-Pinion brutiae B. Foucault 2020 (sur ces syntaxons, voir de Foucault, 2020a), ici représenté entre 700 et 1 100 m d’altitude.
La colonne 14 est le Sorbo oblongifoliae–Juniperetum foetidissimae Barbero & Quézel 1979 nom. inval. (art. 3o, 5), validé ici comme Sorbo oblongifoliae–Juniperetum foetidissimae Barbero & Quézel ex Barbero, Quézel & B. Foucault ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 6 du tableau 11 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 30). Pour la Grèce, on en a rapproché les Abieti cephalonicae-Pinetum pallasianae B. Foucault 2022 (colonne 16), Abieti borisii-regis-Pinetum pallasianae B. Foucault 2022 (colonne 18) et Aceri obtusati-Pinetum pallasianae B. Foucault 2022 (colonnes 19 et 20). Provisoirement on intègre toutes ces associations arborescentes au Pinion pallasianae B. Foucault 2022, bien qu’une certaine différentiation apparaisse au sein de l’ensemble formé par les colonnes 14 à 20 (14 à 17 d’un côté, 18 à 20 d’un autre), ici développé entre 1 200 et 1 950 m.
Le tableau 9 de Barbero & Quézel (1979) correspond plutôt à une ripisylve à Platanus orientalis décrite antérieurement comme Alno orientalis-Platanetum orientalis (Barbero & Quézel 1979) B. Foucault 2021 (de Foucault, 2021c).
4. La végétation arbustive
Pour présenter la végétation arbustive, on va d’abord analyser quelques relevés effectués lors du séjour de mars 2025, rassemblés dans le tableau 2. Les relevés 1 et 2 tout d’abord présentent un fourré arrière-littoral dans lequel Asparagus stipularis s’insère dans des Lycium abondants (photo 18) ; la difficulté est de différencier les deux lyciets présents à Chypre, L. schweinfurthii de L. ferocissimum.

Les relevés 3 à 5 présentent un fourré associé au Pino brutiae-Ceratonietum siliquae, avec des représentants juvéniles de Ceratonia siliqua, associés à Cytisus laniger (photo 19), Crataegus azarolus (photo 20), Prasium majus (photo 21), Rhamnus lycioides subsp. graeca (photo 22). On peut aussi y rencontrer Bosea cypria (photo 23), une Amaranthaceae arbustive, représentant endémique chypriote d’un genre comportant deux autres espèces (une macaronésienne, B. yervamaura, et une himalayo-pakistanaise, B. amherstiana). Cet ensemble correspond à une association originale thermoméditerranéenne que l’on peut dénommer Ceratonio siliquae-Pistacietum lentisci ass. nov. hoc loco (photo 24), typus nominis hoc loco : relevé 5 du tableau 2 hoc loco.
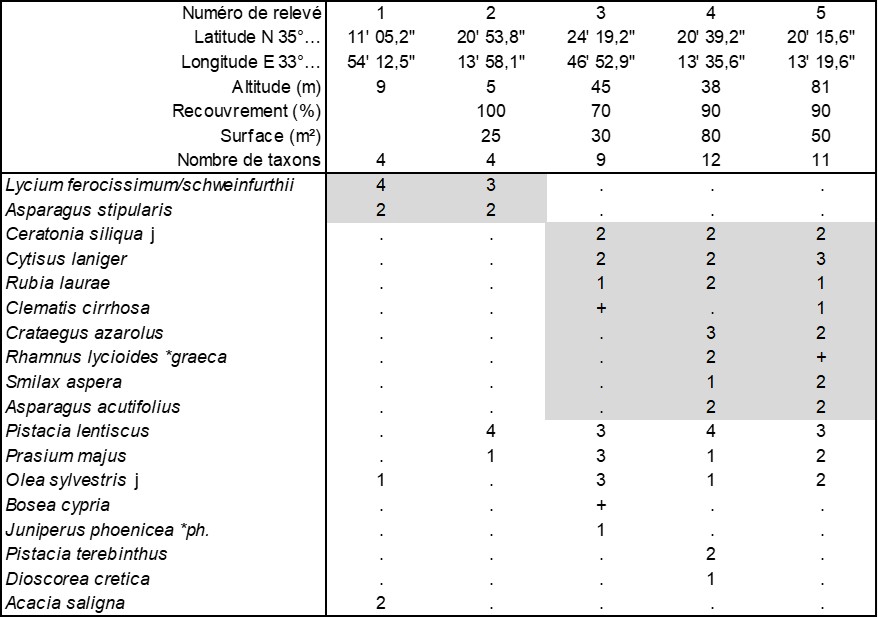
Annexe du tableau 2 – rel. 1 : site archéologique de Salamis ; rel. 2 : Alsancak, Şht. Salahi sokak ; rel. 3 : Tatlısu, D.10 ; rel. 4 : Alsancak, Uzunoğlu, Çağlayan Mahallesi, ; rel. 5 : Alsancak, Çağatay Özbirim caddesi.






La publication de Barbero et Quézel (1979) apporte bien d’autres données, notamment pour la végétation arbustive intraforestière ; elles sont synthétisées dans le tableau 3. Outre la colonne correspondant au Ceratonio-Pistacietum lentisci décrit dans le tableau 2 (en colonne 1), on peut notamment mettre en exergue :
- les colonnes 2 et 3 comme variations d’un unique Lithodoro versicoloris-Genistetum sphacelatae ass. nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 3 du tableau 5 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 20, en changeant Lithospermum hispidulum en Lithodora hispidula subsp. versicolor), lui-même thermoméditerranéen ; le relevé suivant en offre un nouvel exemple :
ouest Girne, Alsancak, Nargis sokak, N 35° 20’ 09,3’’, E 33° 13’ 44,6’’, 92 m, le 14/03/2025, 95 %, 50 m², 17 taxons
- Pistacia lentiscus 3, Cytisus laniger 2, Prasium majus 2, Rubia tenuifolia (photo 25) 2, Teucrium creticum 2, Genista sphacelata 1, Lithodora hispidula *versicolor (photo 26) 1, Smilax aspera 1, Asparagus acutifolius 1, Crataegus azarolus 1, Rhamnus lycioides *graeca 1, Cistus creticus (photo 27) 1, parviflorus (photo 28) +,
- autres taxons : Olea sylvestris j 2, Pinus brutia j 1, Salvia +, Cistus salviifolius ! + ;
- la colonne 5 comme Ephedro foemineae-Juniperetum turbinatae nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 6 du tableau 3 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 16, en changeant Ephedra campylopoda en E. foeminea et Juniperus phoenicea subsp. lycia en J. turbinata) ;
- la colonne 6 est le Vitici agni-casti-Myrtetum communis Barbero & Quézel 1979 inval. (art. 3o, 5), validé ici comme Vitici agni-casti-Myrtetum communis Barbero & Quézel ex Barbero, Quézel & B. Foucault ass. nov. hoc loco, typus nominis hoc loco: composante arbustive du relevé 3 du tableau 4 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 17) ; avec Arbutus andrachne et l’absence de Vitex agnus-castus, à une altitude de 1 100 m, le relevé 5 du tableau 4 des auteurs ne peut évidemment s’y rattacher.




Les colonnes 1 à 10 partagent Rhamnus lycioides subsp. graeca, Cytisus laniger, Ephedra foeminea, Prasium majus. D’après la synthèse de 2021b, avec en plus Rubia tenuifolia, tous ces fourrés sempervirents est-méditerranéens entrent dans les Rubio tenuifoliae-Pistacietalia terebinthi B. Foucault 2021, au sein desquels étaient alors reconnues cinq alliances (colonnes 7 à 10 et 20). Il est nécessaire de définir une alliance pour Chypre, caractérisée par Lithodora hispidula subsp. versicolor, Rubia laurae, Dioscorea cretica, Clematis cirrhosa, Crataegus azarolus, Aristolochia sempervirens, le Lithodoro versicoloris-Rhamnion graecae all. nov. hoc loco, typus nominis hoc loco : le Lithodoro versicoloris-Genistetum sphacelatae nov.
Les colonnes 11 à 16 partagent essentiellement Arbutus andrachne, Styrax officinalis, Lonicera etrusca, Dioscorea cretica. Parmi les colonnes les plus significatives, se distinguent les 13 et 14 comme Arbuto andrachnes-Pistacietum terebinthi ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 2 du tableau 7 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 23). La colonne 15 peut être considérée comme Rubio laurae-Arbutetum andrachnes ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 11 du tableau 8 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 26). Caractérisé ou différencié des autres unités du tableau 3 par Arbutus andrachne, Styrax officinalis, Lonicera etrusca, Dioscorea cretica, cet ensemble a la valeur d’une nouvelle alliance, le Styraco officinalis-Arbution andrachnes all. nov. hoc loco, typus nominis hoc loco : le Rubio laurae-Arbutetum andrachnes nov.
Les colonnes 17 à 20 s’isolent par la présence remarquable de Berberis cretica qui participe à des fourrés développés au-dessus de 1 250 m d’altitude, donc plutôt dans le massif du Troodos (Chypre du Sud), en lien avec les associations arborescentes du Pinion pallasianae. La colonne 20 a été antérieurement décrite comme Aceri sempervirentis-Berberidetum creticae Quézel & Barbero 1980 rattaché à l’Asparago acutifolii-Berberidion creticae des montagnes de Crète (de Foucault, 2021b). La colonne 21 est l’Aceri sempervirentis-Zelkovetum abeliceae Zaffran ex ass. nov. hoc loco [syn. : Zelcovo-Aceretum sempervirentis Zaffran 1990 et Vincetoxico-Zelcovetum abeliceae Zaffran 1990 nom. inval.)] toujours de Crète, typus nominis hoc loco : relevé 62 du tableau 19 in Zaffran (1990, Contributions à la flore et à la végétation de la Crète, hors texte). Les colonnes 17 à 19, des montagnes de Chypre, en sont assez distinctes. Les deux premières peuvent être décrites comme Roso chionistrae-Berberidetum creticae ass. nov. hoc loco (1 250-1 750 m), typus nominis hoc loco : composante arbustive du relevé 5 du tableau 10 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV, hors texte), et Cotoneastro racemiflori-Berberidetum creticae ass. nov. hoc loco (1 700-1 950 m), typus nominis hoc loco : composante arbustive du relevé 3 du tableau 11 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 30). Ces deux associations peuvent être rapprochées dans le Roso chionistrae-Berberidion creticae all. nov. hoc loco, caractérisée ou différenciée de l’alliance crétoise par Rosa chionistra (endémique chypriote), Crataegus monogyna, Cistus creticus et Rubia tenuifolia, typus nominis hoc loco : le Roso chionistrae-Berberidetum creticae nov.
5. La végétation vivace intraforestière
C’est maintenant le tableau synthétique 4 qui servira de base à l’analyse de cette végétation. Les douze colonnes se répartissent en trois ensembles.
Dans l’ensemble formé par les colonnes 1 à 5, correspondant à l’étage thermoméditerranéen, on peut au moins mettre en exergue :
- la colonne 2 comme Teucrio canescentis-Acanthoprasietum integrifolii ass. nov. hoc loco, typus nominis hoc loco: composante basse du relevé 4 du tableau 2 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 17, en remplaçant Teucrium divaricatum subsp. canescens par T. canescens et Ballota integrifolia par Acanthoprasium integrifolium) ;
- les colonnes 4 et 5 comme deux variations de l’Helichryso stoechadis-Thymetum integri nov. hoc loco, typus nominis hoc loco: composante basse du relevé 6 du tableau 5 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV : 20).
Tout cet ensemble paraît avoir la valeur d’une alliance caractérisée ou différenciée par Phagnalon graecum, Teucrium divaricatum, Piptatherum coerulescens, Thymelaea hirsuta, Convolvulus lineatus, Sarcopoterium spinosum (photo 29), Zygophyllum creticum, Thymus integer, Achnatherum bromoides, Melica minuta, Micromeria nervosa (photo 30), Helichrysum stoechas, Thliphthisa suberosa (= Galium suberosum), Carex halleriana, le Phagnalo graeci-Micromerion nervosae all. nov. hoc loco, typus nominis hoc loco : le Teucrio canescentis-Acanthoprasietum integrifolii nov. Avec Thymbra capitata et Sarcopoterium spinosum, ainsi que sa chorologie est-méditerranéenne, cette alliance devrait bien trouver sa place dans les Cisto cretici-Micromerietea julianae Oberd. 1954 et les Sarcopoterietalia spinosi Eig 1939, à côté du Sarcopoterio spinosi-Genistion fasselatae M.J. Costa et al. 1984 (de Foucault, 2020b).


Dans l’ensemble formé par les colonnes 7 à 9, on peut distinguer
- le Teucrio kotschyani-Erophacetum baeticae ass. nov. hoc loco (colonne 7), typus nominis hoc loco: composante basse du relevé 5 du tableau 7 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 23, en remplaçant Astragalus lusitanicus par Erophaca baetica) ;
- le Crepido fraasii-Lecokietum creticae ass. nov. hoc loco (colonne 8), typus nominis hoc loco: composante basse du relevé 8 du tableau 8 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 25, en remplaçant Crepis frasii par C. fraasii et Lecoquia cretica par Lecokia c.) ;
- le Cyclamini cyprii-Lecokietum creticae ass. nov. hoc loco (colonne 9), typus nominis hoc loco: composante basse du relevé 11 du tableau 8 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 25, en remplaçant Lecoquia cretica par Lecokia c.).
Là encore, cet ensemble doit posséder la valeur d’une alliance, caractérisée par Lecokia cretica, Teucrium kotschyanum, Erophaca baetica, Sedum cyprium, le Teucrio kotschyani-Lecokion creticae all. nov. hoc loco, typus nominis hoc loco : le Crepido fraasii-Lecokietum creticae nov.
Enfin, l’ensemble rassemblant les colonnes 10 à 12 est fortement caractérisé :
- en colonne 10, le Corydalido rutifoliae-Odontarrhenetum troodi ass. nov. hoc loco, typus nominis hoc loco: composante basse du relevé 3 du tableau 11 in Barbero & Quézel (1979, Phytosoc., n. s., IV : 30, en remplaçant Alyssum troodi par Odontarrhena t. et Corydalis rutaefolia par C. rutifolia) ;
- en 11, le Salvio willeanae-Paeonietum masculae nov. hoc loco, typus nominis hoc loco: composante basse du relevé 2 du tableau 10 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV hors texte, en remplaçant Salvia grandiflora subsp. wileana par S. willeana) ;
- en 12, le Noccaeo cypriae-Euphorbietum rigoi nov. hoc loco, typus nominis hoc loco: composante basse du relevé 13 du tableau 10 in Barbero & Quézel (1979, Doc. Phytosoc., n. s., IV hors texte, en remplaçant Thlaspi cyprium par Noccaea cypria).
Ce Noccaeo cypriae-Euphorbietum rigoi peut être le typus nominis hoc loco d’une alliance fortement caractérisée par Euphorbia crassa subsp. rigoi, Anthemis tricolor, Scorzonera cana, Teucrium cyprium, par ailleurs assez riche en Orchidaceae (Neotinea maculata, Limodorum abortivum, Platanthera holmboei), le Teucrio cyprii-Euphorbion rigoi all. nov. hoc loco.
De moyenne à haute montagne chypriote, les Teucrio kotschyani-Lecokion creticae et Teucrio cyprii-Euphorbion rigoi n’ont guère de liens synfloristiques avec les Cisto-Micromerietea julianae cités à propos du Phagnalo graeci-Micromerion nervosae ; on peut les rapprocher au moins dans un ordre original, les Teucrio kotschyani-Crepidetalia fraasii ord. nov. hoc loco, caractérisé ou différencié des Sarcopterietalia spinosi par Crepis fraasii, Cicerbita cyprica, Scutellaria cypria, Jurinea cypria, Clinopodium vulgare, Pteridium aquilinum. Ces unités sont donc assez différentes du Phlomido creticae-Crepidion fraasii décrit des montagnes de Grèce (de Foucault, 2022).
La colonne 6 du tableau 4 semble se rattacher aussi à cet ordre, mais est statistiquement insuffisamment définie.
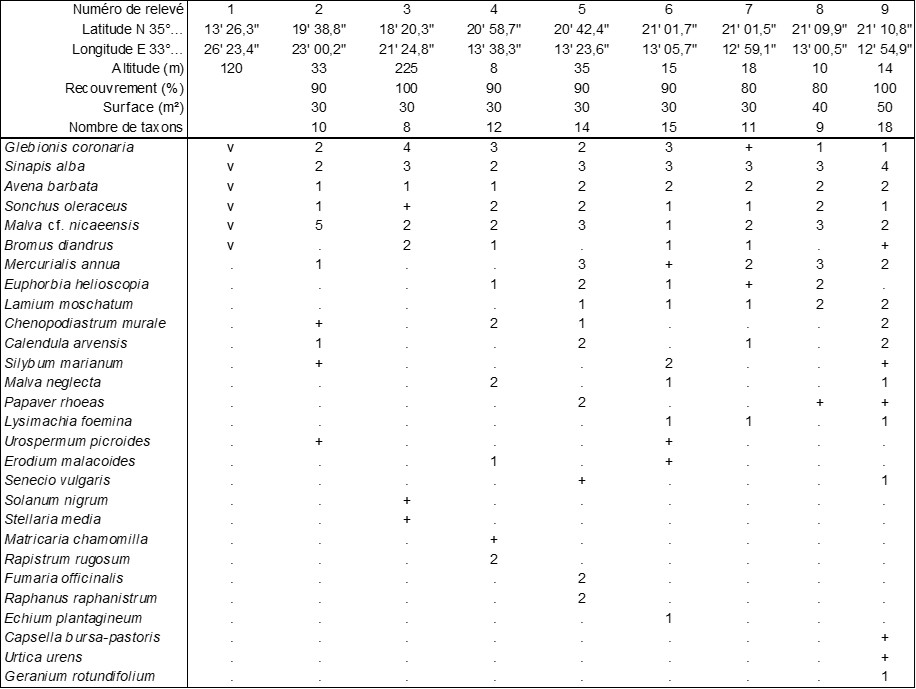
Annexe du tableau 5 – rel. 1 : Minareliköy, Panagia Pergaminiotissa Kirche, 12/03/2025 ; rel. 2 : Namık Kemal sokak, Çatalköy, près du magasin Leather & Fur, 12/03/2025 ; rel. 3 : Bellapaïs Village, 12/03/2025 ; rel. 4 : Dumlupınar sokak, Çağlayan, Alsancak, près du Fish Restaurant, 13/03/2025 ; rel. 5 : Yeşiltepe, Alsancak, vers le croisement Ecevit caddesi ´ Lazmarin sokak, 13/03/2025 ; rel. 6 : Çağlayan, Alsancak, vers Soğular sokak et Tumalar sokak, 14/03/2025 ; rel. 7 : Çağlayan, Alsancak, vers le 5 Savcilak sokak, 14/04/2025 ; rel. 8 : Çağlayan, Alsancak, vers le 25 Savcilak sokak, 14/03/2025 ; rel. 9 : Çağlayan, Alsancak, Zefiros sokak, 14/03/2025.
6. Une friche thérophytique
Le tableau 5 rapporte neuf relevés d’une friche urbaine thérophytique à hautes densité et biomasse fréquemment rencontrée dans les étages inférieurs de Chypre du Nord (photo 31). Elle est caractérisée par Glebionis coronaria (photo 32), Sinapis alba, Bromus diandrus, Lamium moschatum (un lamier est-méditerranéen annuel qui rappelle fortement le vivace L. album, absent de Chypre ; photo 33), Urospermum picroides (photo 34), Chenopodiastrum murale… Une mauve reste de détermination encore incertaine faute d’avoir pu observer les méricarpes parfaitement mûrs. Quoi qu’il en soit, on est en présence d’une association non encore définie, le Glebionido coronariae-Sinapietum albae ass. nov. hoc loco, typus nominis hoc loco : relevé 5 du tableau 5 hoc loco, accueillant en moyenne douze taxons par relevé. Il semble qu’on puisse placer cette association au sein des Sisymbrietea officinalis Korneck 1974 et du Malvion parviflorae (Rivas Mart. 1978) Brullo in Brullo & Marcenò 1985 (de Foucault, 2012).



7. Conclusion
En combinant données de terrain et exploitation moderne de la littérature phytosociologique, on a décrit ici des syntaxons nouveaux pour la végétation de l’île de Chypre. En particulier, on a défini six alliances nouvelles et un ordre nouveau. On a souligné par ailleurs le déficit de connaissances pour les régions est-méditerranéennes relativement aux régions ouest- et centre-méditerranéennes. Notamment on a identifié la classe des Charybdido pancratii-Asphodeletea ramosi récemment décrite comme étant à étudier dans cette île. Il faut espérer que des publications telles que celle-ci y stimuleront de nouvelles recherches. Il reste cependant d’autres publications antérieures sur les régions est-méditerranéennes à exploiter selon la méthodologie suivie ici.
Bibliographie
Alziar G., 1999 (publié 2000). Compte rendu du 4ème Iter Mediterraneum. Bocconea 11 : 5-83.
Alziar G. & Guittonneau G.-G., 2004. Compte rendu des journées d’études de la Société botanique de France à Chypre (5-12 avril 2001 et 2-9 mai 2002). Journal de botanique de la Société botanique de France 25 : 5-25.
Barbero M. & Quézel P., 1979. Contribution à l’étude des groupements forestiers de Chypre. Documents phytosociologiques, n. s., IV : 9-34.
Brandes D., 2020. Some contributions to the wall flora of North Cyprus. Braunschweiger Geobotanische Arbeiten 14 : 11-33.
Brullo S. & Guarino R., 1998. Syntaxonomy of the Parietarietea judaicae class in Europe. Annali di Botanica (Roma) LVI (1) : 109-146.
Costa M., Géhu J.-M., Peris J.B., Biondi E. & Arnold N., 1984. Sobre la vegetación termomediterránea litoral de la isla Chipre. Documents phytosociologiques, n. s., VIII : 365-376.
Coulot P., 2000. Approche de la flore de l’île de Chypre. Le Monde des plantes 470 : 16-20.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Sisymbrietea officinalis Korneck 1974. Journal de botanique de la Société botanique de France 58 : 55-116.
Foucault B. (de), 2014. Contribution au prodrome des végétations de France : les Parietarietea judaicae Rivas-Mart. in Rivas Goday 1964. Acta Bot. Gallica: Botany Letters 161 (4) : 403-427.
Foucault B. (de), 2020a. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2020b. Sur la synsystématique des Rosmarinetea officinalis ouest-méditerranéens. Carnets botaniques 4 : 1-27 et 26 tableaux, https://doi.org/10.34971/s9d8-qj43.
Foucault B. (de), 2021a. Apports de la 155e session extraordinaire de la Société botanique de France à la connaissance de la végétation provençale. Journal de botanique de la Société botanique de France 93 : 62-86.
Foucault B. (de), 2021b. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de), 2021c. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de), 2022. Nouvelles considérations phytosociologiques sur la végétation de la Grèce. Carnets botaniques 109 : 1-27, https://doi.org/10.34971/Q6KD-W822.
Foucault B. (de), 2023. Sur la végétation aérohalophile à Crithmum maritimum, essai de synthèse. Carnets botaniques 147 : 1-23, https://doi.org/10.34971/4XBV-3673.
Foucault B. (de), 2024. La classe des Charybdido pancratii-Asphodeletea ramosi en Méditerranée occidentale. Carnets botaniques 226 : 1-8, https://doi.org/10.34971/WW5N-7K72.
Foucault B. (de) & Bioret F., 2010. Contribution au prodrome des végétations de France : les Saginetea maritimae Westhoff et al. 1962. Journal de botanique de la Société botanique de France 50 : 59-83.
Foucault B. (de) & Jasprica N., 2019. Sur une classe de végétation méconnue en France : les Dittrichietea viscosae Trinajstić, B. Foucault & Jasprica cl. nov. Botanique 5 : 155-170.
Géhu J.-M., Costa M., Biondi E., Franck J. & Arnold N., 1984. Données sur la végétation maritime des côtes méridionales de l’île de Chypre (plages, dunes, lacs salés et falaises). Documents phytosociologiques, n. s., VIII : 343-364.
Géhu J.-M., Costa M. & Uslu T., 1990. Analyse phytosociologique de la végétation littorale des côtes de la partie turque de l’île de Chypre dans un souci conservatoire. Documents phytosociologiques, n. s., XII : 203-234.
Géhu J.-M., Aspostolidès N., Géhu-Franck J. & Arnold K., 1992. Premières données sur la végétation littorale des îles de Rhodos et de Karpathos (Grèce). Colloques phytosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 545-582.
Hand R., Hadjikyriakou G.N. & Christodoulou C.S. (ed.), 2011 (continuously updated). Flora of Cyprus – a dynamic checklist, http://www.flora-of-cyprus.eu/.
Meikle R.D., 1977 et 1985. Flora of Cyprus, I et II. The Bentham-Moxon Trust, Kew, 1 969 p.
Zaffran J., 1990. Contributions à la flore et à la végétation de la Crète. Thèse de doctorat d’État, Presses de l’université de Provence, Aix-en-Provence, 615 p. et tableaux.
Remerciements
Je remercie tout particulièrement Pierre Coulot qui a mis à ma disposition sa publication de 2000 sur ses observations floristiques, ainsi que la flore de Chypre de Meikle.


