
Une excursion botanique au mont Roraima, tepuy du Venezuela
Title
A botanical excursion on Mount Roraima, Venezuela’s tepuy
Résumé
Cet article relate une excursion botanique sur le mont Roraima, tepuy du Venezuela, effectuée durant la saison des pluies de l’été 1999, avec citation des principaux végétaux vasculaires rencontrés et photographiés.
Abstract
This article describes a botanical excursion to Mount Roraima, tepuy in Venezuela, during the rainy season in the summer of 1999, with a list of the main vascular plants encountered and photographed.
Au cours de l’été 1999, un voyage au Venezuela nous a offert l’occasion de pénétrer au cœur de la Gran Sabana et de la région des tepuys, au point de convergence de ce pays avec le Brésil et le Guyana (carte 1), malheureusement en pleine saison des pluies (ce qui explique la qualité un peu faible de certaines photos, correspondant en outre à des scans de diapositives). Tepuys (ou tepuis) est le nom donné par l’ethnie amérindienne des Pemón à ces inselbergs, qu’ils considèrent comme le domaine de leurs dieux, surplombant les savanes d’un millier de mètres (photo 1).
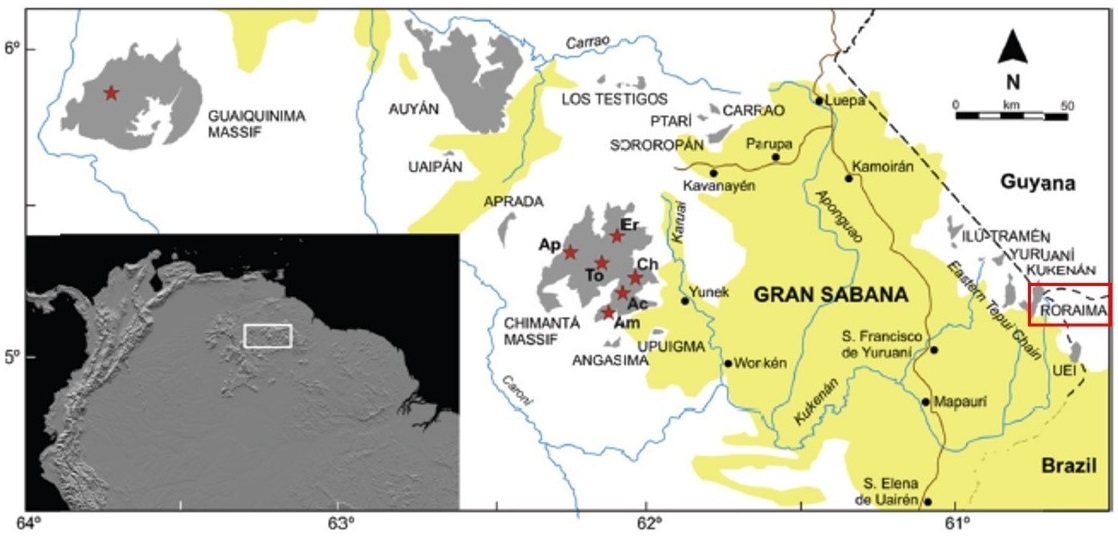

Au plan géologique, ces inselbergs sont formés de grès dérivant de sédiments sableux triasiques et crétacés qui se sont déposés au-dessus du bouclier guyanais précambrien issu du démantèlement des granites et gneiss du Gondwana, puis le massif gréseux a été découpé ultérieurement par l’érosion. Plusieurs massifs se séparent, entourant le cœur de la Gran Sabana : le massif du Guaiquinima au nord-ouest, le massif de l’Auyán au nord, le massif du Chimantá au centre, la chaîne orientale à l’est où se place le Roraima, culminant à 2 810 m, qui nous occupera ici, et le Kukenán (photos 1 et 2), enfin l’ensemble Pico Tamacuari – Sierra de la Neblina au sud-ouest (caché par le médaillon de l’Amérique du Sud sur la carte 1). L’Auyán tepuy est célèbre pour le fameux Salto Angel (le saut Angel, du nom de l’aviateur américain échoué en 1937 au sommet du tepuy qui l’aurait découvert, et non le « saut de l’Ange »), qui serait la plus haute cascade au monde (979 m, dont 807 de chute ininterrompue). D’une manière générale, ces cascades donnent naissance à des cours d’eau qui drainent la Gran Sabana et rejoignent l’Orénoque ; on peut parfois y observer des Apocynaceae Asclepiadoideae submergées comme Ditassa taxifolia. Les masses rocheuses retiennent les nuages et le climat général y est humide et très brumeux, avec des variations de température qui peuvent atteindre 40 °C en quelques heures. La dégradation du grès fournit du sable, l’acidité de ces matériaux et les conditions climatiques, froides et brumeuses, ont déterminé des dépôts de tourbe ; ces reliefs constituent de véritables monuments naturels et artistiques, d’ailleurs inscrits au Patrimoine mondial de l’humanité par l’UNESCO (Parc national de Canaima).

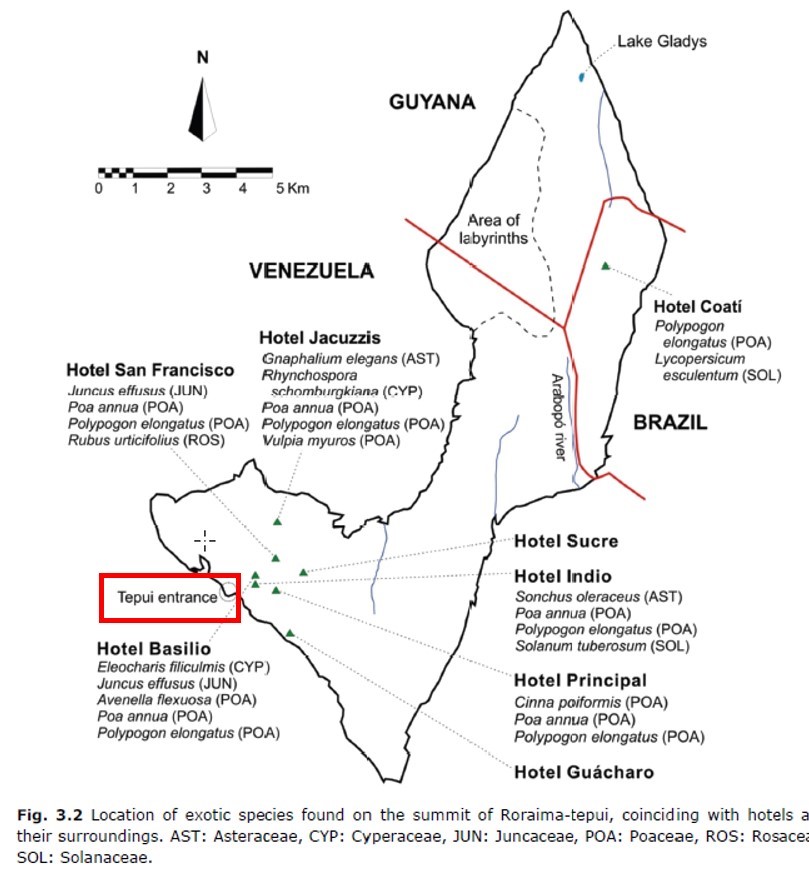
La plupart de ces tepuys sont accessibles seulement par des hélicoptères qui déposent les chercheurs et visiteurs au sommet, seul le mont Roraima (ou de manière abrégée le Roraima) est accessible à pied. Le plus simple est de partir de la petite ville de Santa Elena de Uairén (aisément accessible par bus publics depuis Caracas via El Tigre), de louer les services d’un guide Pemón qui mène ses clients en deux jours au pied du tepuy, que l’on escalade le lendemain en traversant une forêt hygrophile pour atteindre une grande faille oblique menant de la base au sommet de l’inselberg en passant sous quelques cascades (figures 1 et 2). Les « hébergements » consistent en des abris sous roche sommaires parfois pompeusement qualifiés d’« hôtels » (carte 2). En général, les visiteurs restent une ou deux nuits au sommet et redescendent par la même voie jusqu’à la Gran Sabana puis la petite ville de départ. Ils sont instamment priés de revenir avec leurs déchets et de ne pas ramener des morceaux de grès ou de sable.

1. Un peu d’histoire des découvertes botaniques
Les frères R.H. et R. Schomburgk semblent avoir été les premiers botanistes à parvenir au pied du Roraima en 1838 puis en 1842. Cet inselberg fut escaladé et exploré en 1884 par Everard F. im Thurn (1852-1932) qui a relaté son expédition botanique (im Thurn, 1887). À ce botaniste fut dédié le genre Thurnia Hook. f., renfermant trois espèces au sein de la famille des Thurniaceae (photo 3), et plusieurs espèces d’autres genres, dont certaines citées ci-après ; son prénom a en outre été utilisé pour nommer le genre Everardia au sein de la famille des Cyperaceae, aujourd’hui inclus dans Cephalocarpus Nees qui renferme vingt espèces. Les taxons reconnus par im Thurn lors de son expédition furent ultérieurement décrits et publiés par D. Oliver (1830-1916). Au xxe siècle, plusieurs missions botaniques ont été menées par des équipes américaines du New York Botanical Garden et du Missouri Botanical Garden dirigées notamment par B. Maguire (1904-1991) et J.A. Steyermark (1909-1988). Signalons encore l’expédition pluridisciplinaire menée par une équipe française en 1998 et qui a donné lieu à un beau livre (Beau-Douëzy & Cambornac, 1999) sur la Sierra de la Neblina (3 014 m) au sud-ouest de la Gran Sabana.

La connaissance des habitats présents sur le plateau terminal n’a en revanche pas été menée. Ule (1915) a décrit quelques aspects de cette végétation ; plus récemment Brewer-Carias (1987) a réalisé une synthèse qui n’a pu être consultée et Huber (1988) a fait un peu le même travail pour le Pantepui (situé dans le massif de Chimantá, 3 045 m). On peut encore signaler un compte rendu (de Foucault, 2001), modeste et dépourvu de photos, qui servira cependant de base au présent article.
D’un point de vue phytogéographique, considérée comme très ancienne (Crétacé, voire même Jurassique), cette flore se rattache à la région des hauts plateaux guyanais (Region of the Guyana Highlands), à la province du Guyana (Guyana Province) et à l’empire néotropical (Takhtajan, 1986) ; un endémisme très marqué s’y manifeste suite à l’isolement des tepuys dans leur ensemble par rapport à la Gran Sabana et des tepuys entre eux. Toutefois les trois familles citées à l’époque par le phytogéographe russe comme caractéristiques de cette région (Hymenophyllopsidaceae, Tepuianthaeae et Saccifoliaceae) sont aujourd’hui incluses dans des familles bien plus larges (respectivement Cyatheaceae, Thymelaeaceae et Gentianaceae).
2. Quelques aspects de la végétation du Roraima
Depuis la Gran Sabana, l’ascension le long de la faille oblique traverse des boisements bas néphéliphiles favorisés par les nuages et les embruns issus des cascades, dénommés bocacillos accueillant des Weinmannia (Cunoniaceae), Besleria (Gesneriaceae), Clusia (Clusiaceae), Hedyosmum tepuiense (Chloranthaceae), Hyptidendron arboreum (Lamiaceae arbustive), Euterpe catinga var. roraimae (Arecaceae), Podocarpus roraimae (Podocarpaceae), Pouteria rigida (Sapotaceae), Moronobea intermedia (Clusiaceae), Heptapleurum rugosum (Araliaceae), Cattleya violacea (Orchidaceae).
Sur le plateau sommital, les parties de roche nue sont souvent colonisées par des Cyanobactéries pionnières (notamment du genre Stigonema, Stigonemataceae) qui les rendent extrêmement glissantes en saison humide. On y parcourt aussi une savane à Stegolepis guianensis (photo 4), Connellia quelchii (photo 5), des Poaceae, des Cyperaceae du genre Cephalocarpus (incl. Everardia), des Orchidaceae (Epidendrum, Habenaria…).


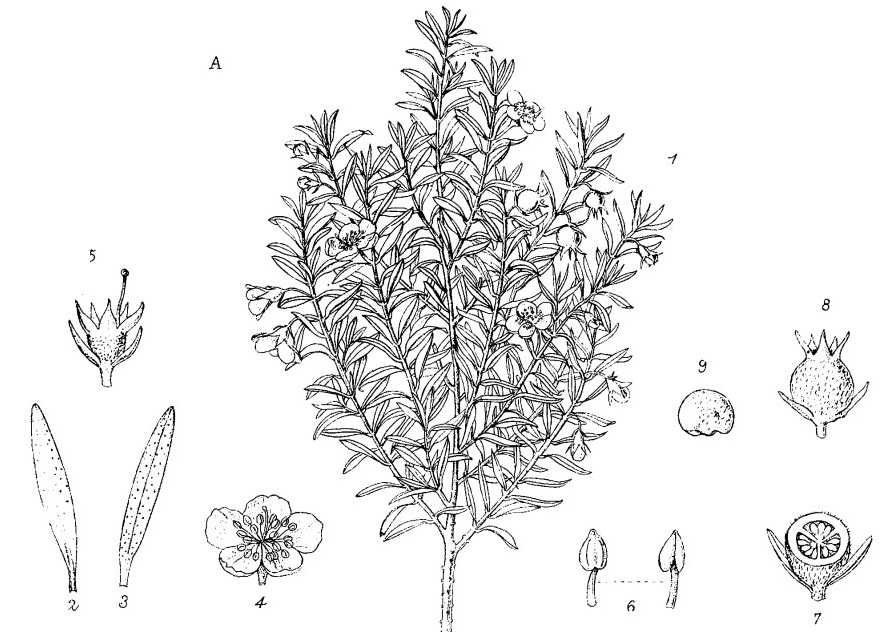
Ce qui paraît être une lande mésophile est caractérisé par des Ericaceae comme Vaccinium roraimense (photo 6), Psammisia coarctata (photo 7), Gaultheria setulosa (photo 8), Ledothamnus guyanensis (photo 9), Bejaria imthurnii (photo 10), ainsi que Ugni myricoides (photo 11 et figure 3) et Sauvagesia imthurniana (photo 12).







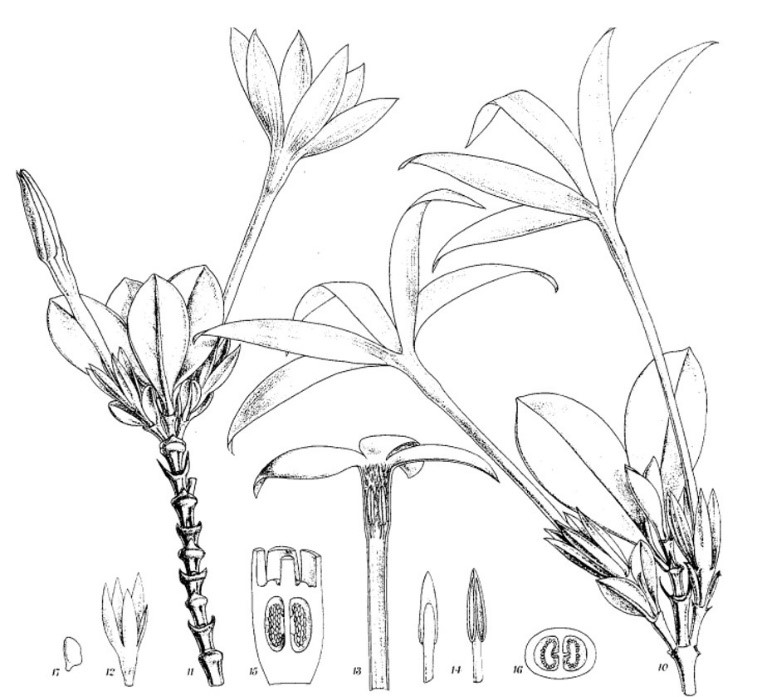
Sur ce plateau, il n’existe pas de véritable forêt, mais seulement des fourrés dominés par Bonnetia roraimae (photo 13), Heptapleurum rugosum (Araliaceae), Weinmannia laxiramea (photo 14), Stomatochaeta condensata (photo 15), Cyrilla racemiflora (photos 16 et 17), Sciodaphyllum monospermum (figure 4), Maguireothamnus speciosus (photo 18 et figure 5), Drimys roraimensis (Winteraceae), Ilex apicidens, I. retusa.





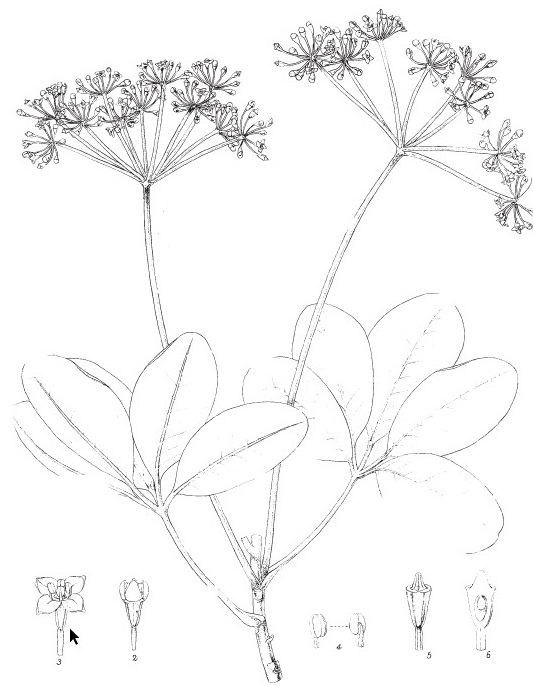

L’eau et le relief tourmenté du plateau sommital favorisent la formation de petits marécages plus ou moins tourbeux, dont l’oligotrophie détermine un bon développement de plantes insectivores. Cinq genres peuvent en effet s’y rencontrer : Utricularia (photos 19 et 20), Genlisea, Drosera (photo 21), Brocchinia (photo 22) et Heliamphora (photo 23). Beau-Douëzy et Cambornac (1999) rappellent que ces plantes possèdent des systèmes de capture et d’assimilation assez diversifiés : primitifs (les feuilles forment des tubes dans lesquels peuvent tomber des débris végétaux et d’insectes et où se produit une dégradation bactérienne libérant des éléments nutritifs pour l’hôte ; Brocchinia hechtioides, B. reducta ; voir aussi Owen & Thomson, 1991), primitifs évolués (pièges passifs avec attraction des proies par du nectar, lesquelles tombent au fond du piège et sont décomposées par des bactéries, l’hôte absorbant les résidus issus de cette décomposition ; Heliamphora), semi-passifs (avec des feuilles pièges possédant une partie en filaments creusés en spirale terminés par une « bouche » attirant des micro-organismes qui seront digérés par des glandes ; Genlisea), actifs à succion (bien connus ; Utricularia) et actifs à tentacules gluants (Drosera). Outre ces végétaux, s’y développent aussi des Xyridaceae comme Orectanthe sceptrum (photo 24), des Eriocaulaceae comme des Paepalanthus (photo 25), ainsi que Harperocallis schomburgkiana (photo 26), Nietneria corymbosa (Nartheciaceae), Pseudolycopodiella caroliniana (Lycopodiaceae). Cette conjonction d’Eriocaulaceae, de Xyridaceae et de taxons insectivores caractérise la structure systématique S(Xyridaceae-Eriocaulaceae) rassemblant des bas-marais oligotrophiles amphibies d’affinités tropicales (photo 27), à laquelle peuvent aussi participer des Isoetes (de Foucault, 1988), ce dernier genre justement représenté sur le Roraima par I. killipii.









On peut enfin citer les fentes de rochers, plus ou moins riches en ptéridophytes chasmophiles, notamment Pterozonium cyclophyllum (photo 28), Cyathea ctenitoides, C. dejecta (Cyatheaeae).

Bibliographie
Beau-Douëzy J.-Ph. & Cambornac M., 1999. Neblina, de brumes et de senteurs. La Martinière, Paris, 140 p.
Brewer-Carias C., 1987. The lost world of Venezuela and its vegetation. Cromotip, Caracas, 223 p.
Brown N.E. et al., 1901. Reports of two botanical collections made by Messrs. F.V. McConnell and J.J. Quelch at Mount Roraima in British Guiana. Transactions of the Linnean Society of London, ser. Botany, VI, part 1 : 1-107, suivi de planches.
Foucault B. (de), 1988. Les végétations herbacées basses amphibies : systémique, structuralisme, synsystématique. Dissertationes Botanicae 121 : 1-150.
Foucault B. (de), 2001. Quelques aspects de la végétation du Venezuela. Bulletin de la Société de botanique du nord de la France 54 (2) : 45-49.
Im Thurn E.F., 1887. Notes on the plants observed during the Roraima expedition of 1884. In The botany of the Roraima expedition of 1884, Transactions of the Linnean Society of London, ser. Botany, II, part 13 : 249-270.
Maguire B. et al., 1953-1965. The botany of the Guayana Highland: a report of the Kunhardt, the Phelps, and the New York Botanical Garden Venezuela expeditions. New York Botanical Garden, plusieurs volumes.
Oliver D., Ridley H.N., Baker J.G. & Mitten W., 1887. A list of the species collected, and Determinations of those are new. In The botany of the Roraima expedition of 1884, Transactions of the Linnean Society of London, ser. Botany, II, part 13 : 271-300 suivi de planches illustrées.
Owen T.P. & Thomson W.W., 1991. Structure and function of a specialized cell wall in the trichomes of the carnivorous bromeliad Brocchinia reducta. Canadian Journal of Botany 69 : 1700-1706.
Safont Crespo E., 2015. Flora and vegetation of the Guyana Highlands, past dynamics, global warming and conservation guidelines. Thèse de doctorat, Barcelona, 314 p.
Steyermark J.A., 1984. Flora of the Venezuelan Guayana, I. Annals of the Missouri Botanical Garden 71 : 297-340.
Ule E., 1915. Die Vegetation des Roraima. Beiblatt zu den Jahrbüchern 115 : 42-53.


