
Retour sur la structuration synsystématique des alliances du Molinion caeruleae et du Juncion acutiflori
Title
Return on the synsystematic structuration of the Molinion caeruleae and Juncion acutiflori alliances
Résumé
À l’aide de tableaux synthétiques, cet article revient sur la structuration synsystématique en sous-alliances du Molinion caeruleae (quatre syntaxons) et du Juncion acutiflori (six syntaxons). Une annexe illustre l’intérêt du tableau phytosociologique comme outil heuristique de détermination.
Abstract
With the help of large tables, this article reviews the synsystematic structuring into suballiances of the Molinion caeruleae (four syntaxons) and the Juncion acutiflori (six syntaxons). An appendix illustrates the interest of the phytosociological table as a determination heuristic tool.
Dans le vaste ensemble phytosociologique des prés hygrophiles oligotrophiles, ceux du Molinion caeruleae se distinguent par leur position originale relativement aux autres prairies et pelouses, ce que ne reflète pas vraiment son nom basé sur un taxon bien plus largement répandu et absolument pas caractéristique de ce niveau hiérarchique. Cette alliance a été reconnue très tôt, puiqu’elle a été mise en évidence par Koch (1926, Jahrb. St. Gall. Naturw. Ges. 61 : 97), lectotypifiée par le Molinietum caeruleae W. Koch 1926 (Jahrb. St. Gall. Naturw. Ges. 61 : 98), source de nombreux homonymes illégitimes ultérieurs.
Dans ma thèse de 1984, j’ai reconnu que cette végétation peut apparaître dans l’oligotrophisation de prairies mésotrophiles hygrophiles (dynamique à humidité édaphique invariante, relation systémique * selon Gillet et al., 1991) ou dans l’humidification de prés et pelouses oligotrophiles mésophiles (dynamique à oligotrophie édaphique invariante, relation systémique b selon Gillet et al., 1991). D’ailleurs, ces deux origines se traduisent dans la composition floristique des moliniaies dérivées, riches en taxons hygrophiles issus de prairies hygrophiles méso- à eutrophiles et de mégaphorbiaies dans le premier cas, riches en taxons issus de pelouses et ourlets mésophiles oligotrophiles et de landes à Ericaceae dans le second.
Dans le présent article, on va compléter cette synthèse en y ajoutant le Juncion acutiflori ; on reprendra la contribution de 1984 et on y ajoutera les syntaxons décrits postérieurement à cette date. Dans les listes synsystématiques, pour ne pas trop encombrer la bibliographie, sauf indications contraires, les sources des données ne sont pas reprises ici, renvoyant au mémoire de 1984. La nomenclature taxonomique suit Flora Gallica (Tison & de Foucault, 2014) pour les taxons présents en France, Euro+Med PlantBase et Catalogue of life (www.catalogueoflife.org) pour les autres taxons selon les cas.
1. Le Molinion caeruleae
1.1. Vue d’ensemble de l’alliance
Dans la synthèse de de Foucault & Géhu (1980) affinée dans la thèse de 1984, le Molinion a été subdivisé en six sous-alliances, certaines validées ultérieurement (de Foucault, 2008) : Agrostio caninae-Molinienion caeruleae nom. ined., Serratulo seoanei-Molinienion caeruleae B. Foucault 2008, Allio angulosi-Molinienion caeruleae B. Foucault & Géhu 1980, Carici davallianae-Molinienion caeruleae B. Foucault & Géhu 1980, Deschampsio mediae-Molinienion caeruleae B. Foucault 2008, plus une sous-alliance non nommée. Le propos du présent article est de compléter et de revenir sur certaines de ces sous-alliances en s’appuyant initialement sur le tableau 1 qui synthétise la plus grande partie des colonnes N à S du tableau 219 de 1984, ces lettres n’étant plus ordonnées dans l’ordre alphabétique pour des raisons synsystématiques. Afin de rester homogène relativement à plusieurs synthèses précédentes, on a utilisé le symbole ‘r’ à la place du symbole ‘o’ pour les présences inférieures à 1 % qui s’introduisent dans de larges synthèses synsystématiques. Pour la colonne P, on n’a pas repris le syntaxon 560, marginal et, avec Selaginella selaginoides, Pinguicula vulgaris, Erica tetralix, plus rarement Narthecium ossifragum, sans doute plutôt à placer dans le Narthecio ossifragi-Caricion hostianae B. Foucault 2022 (de Foucault, 2022). Dans la suite on va reprendre plus en détail ces diverses unités.
1.2. Le Serratulo tinctoriae-Molinienion caeruleae
La colonne O du tableau 1 correspond donc au Serratulo seoanei-Molinienion caeruleae. Selon Flora Gallica, la subsp. seoanei de Serratula tinctoria ne possède pas de réelle valeur taxonomique, de sorte que l’on peut proposer la mutation du nom de la sous-alliance en Serratulo tinctoriae-Molinienion caeruleae B. Foucault 2008 nom. mut. hoc loco.
Le tableau 2 synthétise et actualise les données connues sur cette sous-alliance en tenant compte des tableaux 234 et 235 de la thèse de 1984 et en ajoutant quatre syntaxons décrits ultérieurement :
551. Serratulo seoanei-Molinietum caeruleae (Bellot 1949) Tüxen 1958 ; Espagne.
552. Centaureo radiatae-Molinietum caeruleae Tüxen & Oberd. 1958 ; Espagne ; selon Flora Iberica (http://www.floraiberica.es/floraiberica/texto/pdfs/16_159_029_Centaurea.pdf [31 octobre 2022]), le nom actuel de la centaurée éponyme serait Centaurea nigra subsp. endressii, mais Euro+Med PlantBase retient plutôt C. coronata, d’où la mutation proposée Centaureo coronatae-Molinietum caeruleae nom. mut. hoc loco.
553. Serratulo seoanei-Nardetum strictae Tüxen & Oberd. 1958 ; Espagne.
554. Cirsio filipenduli-Molinietum caeruleae P. Allorge ex B. Foucault 2008 ; Pays basque (de Foucault, 1984) ; photos 1 et 2.


555. Erico scopariae-Molinietum caeruleae Weevers ex B. Foucault 2008 ; Landes de Gascogne (de Foucault, 1984) ; atteint la Double périgourdine (de Foucault, 1993b) et, appauvri, le Lauragais (de Foucault, 2020b : 11) ; photo 3.

556. Moliniaie à Allium ericetorum P.A. Duvign. 1966 (initialement sub moliniaie à Allium ochroleucum), validée ici sous le nom de Filipendulo vulgaris-Caricetum lepidocarpae P.A. Duvign. ex ass. nov. hoc loco, typus nominis hoc loco: relevé 2 du tableau 5 in Duvigneaud (1966, Bull. Soc. Roy. Bot. Belgique 99 (2) : 300) ; massifs serpentiniques du Limousin ; association évoquée aussi par de Foucault (1986a : 306).
557. Peucedano gallici-Molinietum caeruleae Delelis & Géhu ex B. Foucault ; centre de la France (Delelis-Dusollier & Géhu, 1975 ; de Foucault, 1984) ; cf. infra; photo 4.

559. Groupement à Erica vagans-Molinia caerulea.
S-Mc. Simethido mattiazzii-Molinietum caeruleae ass. nov. hoc loco (de Foucault, 1993b, tableau 2 : relevés 7 à 12), typus nominis hoc loco : relevé 7 du tableau 2 in de Foucault (1993b, Bull. Soc. Bot. Centre-Ouest, n. s., 24 : 165) ; camp militaire de Montmorillon (Vienne) ; pourrait peut-être aussi correspondre au pré oligotrophile mésohygrophile dérivé du Festuco filiformis-Simethidetum mattiazzii de l’Orléanais (de Foucault, 2023).
H-Sh. Helictochloo marginatae-Scorzoneretum humilis B. Foucault 1993 nom. mut. hoc loco (de Foucault, 1993b, tableau 5, sub Avenulo sulcatae-Scorzoneretum humilis) ; Touraine ; photo 5.

C-Sh. Cirsio filipenduli-Scorzoneretum humilis B. Foucault 1993 ; a – de Bretagne péninsulaire (de Foucault, 1993b, tableau 9) ; b – de la presqu’île de Guérande (ibid., tableau 7).

S-Sp. Serratulo tinctoriae-Succisetum pratensis B. Foucault 1995 (de Foucault, 1995a, tableau 14) ; Cotentin, Argonne.
558. Moliniaie de New Forest, se confondant pratiquement avec l’association précédente dont l’aire s’étendrait désormais au sud-est du Royaume-Uni.
En définitive, on peut proposer d’inclure dans cette sous-alliance le syntaxon non nommé décrit par la colonne P du tableau 1 qui inclut notamment le Peucedano gallici-Molinietum caeruleae décrit à titre provisoire par Delelis-Dusollier & Géhu (1974 : 145 et tableau 2) et confirmé par de Foucault (1984 : 356 et tableau 132) ; il est temps de valider définitivement ce nom : Peucedano gallici-Molinietum caeruleae Delelis & Géhu ex ass. nov. hoc loco, typus nominis hoc loco : relevé 7 du tableau 132 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) (on préfère choisir un des relevés de ce tableau à l’un de ceux de Delelis & Géhu plutôt paucispécifiques) publié ci-dessous :
Courcy-aux-Loges (45), forêt domaniale d’Orléans, carrefour D 921 × route de la Vallée des Joncs, ~ N 48° 02’ 46,2’’, E 2° 12’ 46,7’’, 132 m, 12/07/1982, 85 %, 30 m², 31 taxons
- combinaison caractéristique : Molinia caerulea 32, Peucedanum gallicum 22, Succisa pratensis 22, Potentilla erecta 22, Danthonia decumbens *d. 22, Carex flacca *f. 22, Scorzonera humilis 12, Agrostis canina *c. (12), Serratula tinctoria 11, Juncus conglomeratus 12, Erica cinerea 12, Calluna vulgaris 12, Betonica officinalis *o. 12, Genista tinctoria 12, Carex panicea 12, tomentosa +, Silaum silaus +,
- autres taxons : Agrostis capillaris 23, Festuca filiformis 23, Avenella flexuosa *f. 12, Melampyrum pratense 12, Holcus mollis *m. +, Carex pilulifera *p. +, Hypericum pulchrum +, Teucrium scorodonia +, Lathyrus linifolius +, Solidago virgaurea *v. +, Agrimonia eupatoria *e. +, Polygala vulgaris *v. +, Betula pubescens j +°, Achillea ptarmica *p. r.
Il existe sur le versant sud, audois, de la Montagne Noire un pré hygro-oligotrophile original systémiquement relié à une pelouse à Serapias lingua et Danthonia decumbens qui semble devoir se rapporter aussi à cette sous-alliance et dont le tableau 3 rapporte deux relevés [rel. 1 : Villardonnel (11), entre la D 118 et le chemin de la Canal, 26/05/2020 ; rel. 2 : Montolieu (11), domaine de Peyremale, relevé repris in de Foucault & Klesczewski (2021, haut de p. 15)]. Les connaissances acquises sur le fonctionnement systémique de ces végétations herbacées vivaces incitent à considérer qu’il s’agit d’une association originale nouvelle, le Serapiado linguae-Scorzoneretum humilis ass. nov. hoc loco (photo 7), typus nominis hoc loco : relevé 1 du tableau 3 hoc loco retranscrit ci-après :
cf. localisation supra, N 43° 20’ 10’’, E 2° 17’ 58,2’’, 361 m, 26/05/2020, 100 %, 90 m², 30 taxons
- combinaison caractéristique : Scorzonera humilis 3, Carex panicea 3, Dactylorhiza maculata 2, Agrostis canina *c. 2, Carex flacca *f. 2, Juncus conglomeratus 2, acutiflorus 1, Luzula multiflora *m. 1, Filipendula vulgaris 1, Serratula tinctoria +, Kickxia commutata +,
- autres taxons : Anthoxanthum odoratum 3, Danthonia decumbens *d. 3, Serapias lingua 2, Ajuga reptans 2, Holcus lanatus *l. 2, Ranunculus acris s. l. 2, Centaurea jacea 2, Rumex acetosa *a. 2, Prunella vulgaris 1, Lathyrus pratensis 1, Briza media *m. 1, Schedonorus arundinaceus *a. 1, Cynosurus cristatus 1, Fraxinus angustifolia *a. j 1, Lychnis flos-cuculi *f. 1, Plantago lanceolata +, Dactylis glomerata *g. +, Cerastium fontanum *vulgare +, Leucanthemum ircutianum + ;

le relevé 2 du tableau 3 correspond à une variation moins acidiphile différenciée par Schoenus nigricans, Juncus subnodulosus et Lotus maritimus, de transition vers le Cirsio tuberosi-Schoenetum nigricantis (cf. infra, § 1.6).
L’appauvrissement floristique de cette sous-alliance relativement aux autres du Molinion caeruleae peut justifier son rattachement plutôt au Juncion acutiflori comme le proposent Julve (1993) puis Bardat et al. (2004).
1.3. L’Allio angulosi-Molinienion caeruleae
La colonne Q du tableau 1 (Allio angulosi-Molinienion caeruleae) ne pose pas de problème particulier ; elle synthétise les syntaxons suivants repris plus en détail dans le tableau 4 et répartis en deux groupes, le syntaxon D-Mc ne se rattachant bien à aucun des deux.
* Groupe de syntaxons à Allium angulosum (photo 8), A. suaveolens, Lotus maritimus, Symphytum officinale, Schoenus nigricans…
561. Oenantho lachenalii-Molinietum caeruleae G. Phil. 1960 ; Allemagne.
562. Cirsio tuberosi-Molinietum caeruleae (Scherrer 1925) Oberd. et al. 1967 ; Allemagne et France orientale, en Alsace (et peut-être jusqu’en Champagne ; de Foucault & Julve, 2002 : 86 et photo 9 ; non cité toutefois par Royer et al., 2006) ; le relevé suivant semble aussi y appartenir :
Soleymieu (38), sous la ferme de Billonay, ~ N 45° 43’ 30,8’’, E 05° 20’ 32’’, 327 m, 32 taxons


– Molinion caeruleae et unités supérieures : Molinia caerulea 5, Carex tomentosa 2, C. panicea 2, Succisa pratensis 2, Silaum silaus 1, Inula salicina 1, Cirsium tuberosum 1, Lotus maritimus 1, Deschampsia media *m. 1, Genista tinctoria +, Jacobaea erucifolia *e. +,
– autres taxons : Carex flacca *f. 2, Agrostis stolonifera 1, Gymnadenia conopsea +, Prunella vulgaris +, Danthonia decumbens *d. +, Ononis spinosa *procurrens +, Galium verum *v. +, Mentha aquatica +, Euphorbia flavicoma *verrucosa +, Galium album +, Brachypodium rupestre *r. +, Cervaria rivini +, Briza media *m. +, Potentilla reptans 1, Juncus inflexus +, J. articulatus *a. +, Cornus sanguinea *s. j +, Ligustrum vulgare j +, Juniperus communis *c. j +, Crataegus monogyna j +, Rhamnus cathartica j +.
Provenant du Bugey, la liste suivante paraît s’y rattacher aussi : Lhuis (01), bord de la D 41, vers 400 m, 14/07/1989
– Molinion caeruleae et unités supérieures : Molinia caerulea, Carex tomentosa, C. panicea, Cirsium tuberosum, Inula salicina, Lotus maritimus, Silaum silaus, Betonica officinalis *o., Succisa pratensis, Genista tinctoria, Dactylorhiza maculata, Jacobaea erucifolia *e., Colchicum autumnale,
– autres taxons : Galium verum *v., Ononis spinosa *procurrens, Carex flacca *f., Briza media *m., Gymnadenia conopsea, Poterium sanguisorba *s., Cirsium acaulon, Viola hirta, Arrhenatherum elatius *e., Centaurea jacea, Euphorbia flavicoma *verrucosa, Danthonia decumbens *d.
563. Iridetum sibiricae G. Phil. 1960 ; Allemagne.
564. Kadenio dubiae-Juncetum subnodulosi Korneck 1962 ; Allemagne.
565. Sanguisorbo officinalis-Silaetum silai Knapp 1946 ; Europe centrale.
566. Centaureo pannonicae-Molinietum caeruleae (Jarai 1958) B. Foucault & Géhu 1980 ; Hongrie ; le type désigné en 1980 a été qualifié de lectotype, il s’agit en fait d’un holotype.
567. Succiso pratensis-Molinietum caeruleae Soó 1968 apud Kuyper et al. 1978 ; Hongrie.
568. Molinietum caeruleae sensu Špánicková 1978 ; ex-Tchécoslovaquie.
V-Is. Violo elatioris-Inuletum salicinae Didier & J.-M. Royer 1989 (Didier & Royer, 1989, tableau II) ; Champagne crayeuse.
* Groupe de syntaxons à Danthonia decumbens subsp. d., Dactylorhiza maculata
569. Molinietum caeruleae sensu Grynia 1962 p. p. (pour l’autre partie du syntaxon initial voir le n° 549 infra) ; Pologne.
570. Peucedano rocheliani-Molinietum caeruleae Boscaiu 1965 ; Roumanie.
571. Molinietum caeruleae à Koeleria delavignei Shelyag-Sosonko et al. 1981 ; Ukraine.
572. Gentiano pneumonanthes-Molinietum caeruleae Ilijanić 1968 nom. prov. ; Croatie.
573. Schedonoro arundinacei-Molinietum caeruleae Mayot in Ferrez et al. 2011 (Mayot, 1977, tableau IX, sub Festuco arundinaceae-Molinietum caeruleae); Jura inférieur. Ce nom publié en 2011 est pourtant invalide car le premier taxon éponyme manque dans le type choisi par les validateurs (art. 3f) ; on le valide correctement ici : Schedonoro arundinacei-Molinietum caeruleae Mayot & B. Foucault ass. nov. hoc loco, typus nominis hoc loco: relevé 9 du tableau IX in Mayot (Essai d’interprétation de la végétation… hors texte), publié ci-après
région d’Orgelet (39, aucune localisation plus précise donnée par l’auteur, à la différence des autres tableaux de sa thèse), vers 500 m, 32 taxons
– Molinion caeruleae et unités supérieures : Succisa pratensis 2, Carex pulicaris 2, Molinia caerulea 1, Scorzonera humilis 1, Genista tinctoria 1, Carex panicea 1, C. hostiana +, Serratula tinctoria +, Juncus conglomeratus +, Sanguisorba officinalis +, Silaum silaus +, Potentilla erecta +, Galium boreale +,
– autres taxons : Centaurea jacea 1, Briza media *m. 1, Agrostis stolonifera 1, Anthoxanthum odoratum 1, Festuca filiformis 1, F. rubra *r. +, Schedonorus arundinaceus *a. +, Lotus corniculatus *c. +, Danthonia decumbens *d. +, Ranunculus acris s. l. +, Holcus lanatus *l. +, Leontodon hispidus *h. +, Cerastium fontanum *vulgare +, Gymnadenia conopsea +, Trifolium montanum *m. +, Nardus stricta +, Pedicularis sylvatica *s. +, Cirsium palustre +.
574. Selino carvifoliae-Juncetum subnodulosi (P. Allorge 1922) B. Foucault 2008 ; plaines de la France nord-orientale ; voir aussi Bournérias (1949 : 35 et tableau VI) et de Foucault et al. (1992b) ; le tableau 5 [rel. 1 – Daours (80), le marais, 15/04/1984 ; rel. 2 – Monceaux (60), le bois de Monceaux, 12/06/1986] en rapporte deux relevés inédits de Picardie orientale.
D-Mc. Dactylorhizo praetermissae-Molinietum caeruleae J.-M. Royer, Thévenin & Didier in J.-M. Royer et al. 2006 (Royer et al., 2006, tableau 7), qui se rattache mal à l’un des deux groupes précédents ; France orientale (Marne et Aube).
Cette sous-alliance est donc représentée en France au moins par le Cirsio tuberosi-Molinietum caeruleae (Scherrer 1925) Oberd. et al. 1967, l’Oenantho lachenalii-Molinietum caeruleae G. Phil. 1960, le Dactylorhizo praetermissae-Molinietum caeruleae J.-M. Royer, Thévenin & Didier in J.-M. Royer et al. 2006 et, en irradiation occidentale, le Schedonoro arundinacei-Molinietum caeruleae Mayot & B. Foucault, le Selino carvifoliae-Juncetum subnodulosi (P. Allorge 1922) B. Foucault 2008.
1.4. Le Carici davallianae-Molinienion caeruleae
Dans la synthèse de 1984, le Molinietum caeruleae W. Koch 1926 n’avait pas été considéré, sans doute en raison de son nom assez peu expressif. En injectant la synthèse des vingt relevés du tableau X de cet auteur dans notre tableau 6 (colonne Mc), on s’aperçoit que cette association type du Molinion caeruleae se place plutôt bien dans la colonne R (Carici davallianae-Molinienion caeruleae) du tableau 1, ce qui a pour conséquence immédiate de réduire ce Carici-Molinienion caeruleae à l’autonyme Eu-Molinienion caeruleae. Le tableau 6 actualise les syntaxons qui s’y rarttachent.
575. Ranunculo polyanthemoidis-Molinietum caeruleae Rameau & J.-M. Royer 1978 ; plateau de Langres (Rameau & Royer, 1978 ; Royer & Didier, 1996 ; de Foucault, 2006 : 488).
576. Trollio europaei-Molinietum caeruleae Guin. in Guin. & G. Lemée 1950 ; haut Jura ; photo 10.

577. Gentiano asclepiadeae-Molinietum caeruleae Oberd. 1957 ; massifs du sud-ouest de l’Allemagne et du nord-ouest de la Suisse ; cité de Haute-Savoie vers 900 m d’altitude par de Foucault & Philippe (1990 : 395) ; photo 11.

578. Juncetum subnodulosi W. Koch 1926 ; nord-est de la Suisse.
579. Juncetum subnodulosi schoenetosum ferruginei G. Lang 1973 ; environs du lac de Constance (sud Allemagne et nord Suisse).
580. Juncetum subnodulosi sensu Oberd. 1957 ; massifs du sud-ouest de l’Allemagne.
581. Euphrasio marchesettii-Molinietum caeruleae Poldini 1973 ; massif du Frioul (nord-est de l’Italie).
582. Selino carvifoliae-Molinietum caeruleae Bal.-Tul. 1972 ; ex-Tchécoslosvaquie.
583. Seslerietum uliginosae Zlatnik 1928 ; ex-Tchécoslosvaquie.
584. Groupement à Cirsium canum-Molinia caerulea ; ex-Tchécoslosvaquie.
585. Carici davallianae-Molinietum caeruleae Špánicková 1978 ; ex-Tchécoslosvaquie.
586. Groupement à Sesleria caerulea-Primula farinosa ; Finlande.
587. Groupement à Tomenthypnum nitens-Juncus subnodulosus ; Suède.
Mc. Molinietum caeruleae W. Koch 1926 (Koch, 1926, tableau X) ; nord Suisse.
1.5. L’Agrostio caninae-Molinienion caeruleae
La quatrième sous-alliance du Molinion caeruleae rassemblant des prés hygrophiles oligotrophiles acidiclinophiles avait été définie dans ma thèse (de Foucault, 1984) sous le nom d’Agrostio caninae-Molinienion caeruleae nom. ined. (tableau 1 : colonne N), elle a été publiée, validée sous le nom de Junco acutiflori-Molinienion caeruleae B. Foucault & Géhu ex B. Foucault 2019 (Evaxiana 6 : 57) et typifiée par le Succiso pratensis-Silaetum silai J. Duvign. ex B. Foucault 2019. Le tableau 7 actualise le tableau 233 du document de thèse, les numéros 535 à 550 étant les codes des syntaxons qui s’y rattachent (le syntaxon 542 n’est pas repris ici, maintenant placé dans les Nardetea strictae et le Nardo strictae-Juncion squarrosi ; dans ce tableau 7, certains syntaxons ont été déplacés pour une meilleure esthétique) et en ajoutant deux associations décrites ultérieurement (colonnes C-Sh et S-Ss).
* Groupe d’associations subatlantiques à Silaum silaus et Juncus acutiflorus
535. Succiso pratensis-Silaetum silai J. Duvign. ex B. Foucault 2019 race subatlantique-subcontinentale ; nord et nord-est de la France, Fagne et Famenne belges (photo 12) ; peut-être jusqu’au plateau de Chambaran (de Foucault, 1997b : 243) ; le relevé suivant en offre un nouvel exemple inédit :
Montagnat (01), parc de Noirefontaine, route de Pont d’Ain, ~ N 46° 10’ 04,9’’, E 05° 16’ 07,2’’, 237 m, 03/07/1988, 90 %, 25 m², 33 taxons
- combinaison caractéristique : Carex panicea 3, Succisa pratensis 2, Juncus conglomeratus 2, acutiflorus 1, Silaum silaus 1, Lotus pedunculatus 1, Betonica officinalis *o. +, Carex pallescens +, Genista tinctoria +, Potentilla erecta 1,
- autres taxons : Anthoxanthum odoratum 2, Danthonia decumbens *d. 2, Carex flacca *f. 2, Achillea ptarmica *p. 2, Angelica sylvestris *s. 2, Festuca rubra s. l. 2, Cirsium palustre 1, Centaurea jacea 1, Myosotis scorpioides 1, Agrostis capillaris 1, Poa trivialis *t. 1, Lotus corniculatus *c. 1, Trifolium medium *m. 1, Achillea millefolium +, Carex hirta +, Lysimachia nummularia +, Ajuga reptans +, Leucanthemum ircutianum +, Veronica chamaedrys *ch. +, Lychnis flos-cuculi *f. +, Cytisus scoparius *s. j +, Salix caprea j +, Alnus glutinosa j +.

536. Succiso pratensis-Silaetum silai J. Duvign. ex B. Foucault 2019 race thermo-atlantique ; ce syntaxon est en définitive suffisamment distinct de l’association initiale de Duvigneaud (1955 ; ici syntaxon 535) pour justifier une nouvelle association vicariante à répartition plus occidentale : Cirsio dissecti-Silaetum silai ass. nov. hoc loco, typus nominis hoc loco: relevé 3 du tableau 56 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) publié ci-après
Sainte-Anne-sur-Brivet (44), La Turcandais, N 47° 27’ 42,9’’, O 1° 59’ 16,5’’, 7 m, 21/05/1981, 100 %, 30 m², 45 taxons
- combinaison caractéristique : Silaum silaus 32, Succisa pratensis 22, Scorzonera humilis 22, Carex panicea 22, Colchicum autumnale 22, Juncus acutiflorus 12, Ophioglossum vulgatum 12, Cirsium dissectum +2, Danthonia decumbens *d. 12, Luzula multiflora *m. +, Carex leporina *l. +, Ranunculus flammula *f. +, Betonica officinalis *o. +2, Coeloglossum viride +,
- autres taxons : Centaurea decipiens 22, Festuca rubra *r. 23, Anthoxanthum odoratum 22, Ajuga reptans 11, Taraxacum 12, Lysimachia nummularia 11, Lychnis flos-cuculi *f. 11, Leucanthemum ircutianum 11, Pulicaria dysenterica 12, Ranunculus acris s. l. 12, Schedonorus arundinaceus *a. 11, Rumex acetosa *a. 12, Plantago lanceolata 12, Lathyrus pratensis 12, Cardamine pratensis 11, Carex pilulifera *p. 12, Briza media *m. 11, Hypochaeris radicata 12, Anacamptis laxiflora 11, A. morio *m. +, Daucus carota *c. +, Gaudinia fragilis +, Cynosurus cristatus +, Oenanthe silaifolia +, Cerastium fontanum *vulgare +, Holcus lanatus *l. +, Trifolium pratense +, Pilosella officinarum +, Vicia segetalis +, Medicago lupulina +, Carex disticha 11.
Ch-Sh. Carici hostianae-Scorzoneretum humilis B. Foucault 2019 (de Foucault, 2019a, tableau 12 et photo 10) ; pays de Sault (Pyrénées audoises).
537. Filipendulo ulmariae-Scorzoneretum humilis Géhu 1961, association un peu complexe dont une partie est maintenant intégrée au Junco conglomerati-Scorzoneretum humilis Trivaudey 1997 (Trivaudey, 1997) ; nord à est de la France.
538. Groupement à Alchemilla gr. vulgaris-Scorzonera humilis Malcuit 1928, peut-être une forme du Junco conglomerati-Scorzoneretum humilis Trivaudey 1997.
539. Selino carvifoliae-Juncetum acutiflori G. Phil. 1960 (Philippi, 1960), association connue aussi du nord de la France (de Foucault, 1993a ; de Foucault & Matysiak, 1995 ; de Foucault et al., 2000) ; photo 13.

543. Molinietum caeruleae sensu Tüxen 1937 nom. illeg. (art. 31, non W. Koch 1926) ; Allemagne du nord-est ; à renommer, mais les données d’origine sont synthétiques de sorte qu’on ne peut en extraire un type.
540. Kadenio dubiae-Juncetum acutiflori Oberd. 1957 nom. mut. hoc loco (initialement sub Cnidio venosi-Juncetum acutiflori) ; sud-ouest de l’Allemagne ; passage entre les deux groupes, ainsi que l’association suivante.
S-Ss. Selino carvifoliae-Silaetum silai Misset in J.-M. Royer, Felzines, Misset & Thévenin 2006 (Royer et al., 2006, tableau 6) ; Ardennes françaises.
* Groupe d’associations plus continentales à Sanguisorba officinalis, Galium boreale…
541. Scorzonero humilis-Molinietum caeruleae Oberd. & Krause 1955 ; sud-ouest de l’Allemagne.
542. Molinietum balticum F. Fukarek 1961 nom. illeg. ; nord de l’Allemagne.
545. Gentiano pneumonanthes-Molinietum arundinaceae sensu Kuyper et al. 1978 (tableau VIII, sub Gentiano pneumonanthes-Molinietum arundinaceae Ilijanić 1968) ; la comparaison avec le tableau princeps de Ilijanić (1968, tableau IV) montre que ce rattachement n’est pas correct et qu’il faut définir un autre nom : Cirsio rivularis-Selinetum carvifoliae (Kuyper, Leeuwenberg & Hübl 1978) nom. nov. hoc loco, typus nominis hoc loco: relevé 116 du tableau VIII in Kuyper et al. (1978, Linzer Biol. Beiträge 10 (2) hors texte) ; Autriche.
546. Willemetio stipitatae-Caricetum paniceae Moravec 1965 ; ex-Tchécoslosvaquie.
547. Groupement à Cirsium rivulare-Molinia caerulea Kopecký 1960.
548. Molinietum caeruleae sensu Resmeriţà 1977 ; encore un nom illégitime (art. 31, non W. Koch 1926) à renommer : Gladiolo imbricati-Molinietum caeruleae (Resmeriţà 1977) nom. nov. hoc loco, typus nominis hoc loco: relevé du tableau 3 in Resmeriţà (1977, Doc. Phytosoc., n. s., I : 249-250) ; Carpates roumaines.
549. Molinietum caeruleae sensu Grynia 1962 p. p. ; Pologne.
550. Junco–Molinietum caeruleae auct. medioeur.
Dans le premier groupe de syntaxons de cette sous-alliance, on peut placer aussi la moliniaie acidiclinophile à Gladiolus palustris de Haute-Savoie (forêt de Planbois), malheureusement connue d’un unique relevé : Gladiolo palustris-Juncetum acutiflori ass. nov. hoc loco, typus nominis hoc loco : relevé in de Foucault & Philippe (1990, Bull. Soc. Bot. Centre-Ouest, n. s., 21 : haut de p. 393) ; le nom de « Gladiolo palustris-Molinietum caeruleae » ne peut être utilisé ici en dépit de la dominance de Molinia caerulea car ce nom est déjà utilisé pour un syntaxon slovène assez différent (Dakskobler et al., 2021). Sous le nom de Trollio europaei-Molinietum caeruleae juncetosum acutiflori, basé sur un unique relevé, Bailly (in Ferrez et al., 2011 : 210) évoque un pré oligotrophile mésohygrophile acidiclinophile montagnard à ratttacher au Junco acutiflori-Molinienion caeruleae plutôt qu’à l’Eu–Molinienion caeruleae auquel se rattache le Trollio-Molinietum typicum. Le relevé suivant paraît aussi se rattacher à cette sous-alliance :
Saint-Évroult-Notre-Dame-du-Bois (61), forêt de Saint-Évroult, berme de la D 50, non loin de Saint-Hubert, ~ N 48° 45’ 55,9’’, E 0° 25’ 39,5’’, 292 m, 27/04/1994, 44 taxons
- combinaison caractéristique : Selinum carvifolia 2, Carex flacca *f. 2, Succisa pratensis 2, Betonica officinalis *o. 2, Molinia caerulea 1, Scorzonera humilis 1, Dactylorhiza maculata 1, Juncus effusus 1, Silaum silaus +, Lotus pedunculatus +, Carex panicea +, Potentilla erecta +, Ophioglossum vulgatum +, Jacobaea erucifolia +, Trocdaris verticillatum +, Colchicum autumnale +,
- autres taxons : Primula veris *v. 2, Centaurea decipiens 2, Festuca rubra *r. 2, Dactylis glomerata *g. 1, Lathyrus pratensis 1, Viola hirta 1, riviniana 1, Angelica sylvestris *s. 1, Carex caryophyllea 1, Achillea millefolium 1, Neottia ovata 1, Vicia sepium 1, Potentilla sterilis 1, Ajuga reptans 1, Clinopodium vulgare 1, Leucanthemum ircutianum 1, Hieracium lachenalii 1, H. virgultorum +, Hypericum gr. maculatum +, Fragaria vesca +, Brachypodium sylvaticum +, Ranunculus acris s. l. +, Pimpinella major +, Rumex acetosa *a. +, Pilosella officinarum +, Teucrium scorodonia +, Poterium sanguisorba *s. +, Salix atrocinerea j +,
prairie forestière riche en taxons des ourlets mésophiles acidiphiles du Conopodio majoris-Teucrion scorodoniae et rappelant le Selino carvifoliae-Juncetum acutiflori, mais ici avec une petite tonalité atlantique indiquée par Trocdaris verticillatum.
1.6. Le Deschampsio mediae-Molinienion caeruleae
Lors d’excursions et randonnées dans la nature audoise, on observe de temps à autre des taxons caractéristiques de l’alliance du Molinion caeruleae et unités supérieures, dont Cirsium tuberosum, Inula salicina, Lotus maritimus, Schoenus nigricans, et pourtant aucune association de cette alliance ne semble décrite dans ce département de l’Aude. Le tableau 8 rapproche onze relevés d’un tel syntaxon, qui paraît bien ne correspondre à aucune association connue au sein du Molinion caeruleae, plus précisément du Deschampsio mediae-Molinienion caeruleae. On propose donc de définir ici le Cirsio tuberosi-Schoenetum nigricantis ass. nov. hoc loco (photo 14 ; le nom de Cirsio tuberosi-Molinietum caeruleae est déjà occupé pour une association médio-européenne relevant de l’Allio angulosi-Molinienion caeruleae, cf. supra, syntaxon 562), typus nominis hoc loco : relevé 1 du tableau 8 hoc loco, retranscrit ici
Saint-Martin-le-Vieil (11), près de l’abbaye de Villelongue, La Jambe de l’Homme, N 43° 18’ 17,6’’, E 2° 09’ 49,2’’, 180 m, 40 %, 50 m², 26/07/2020, 21 taxons
- combinaison caractéristique : Schoenus nigricans 3, Molinia caerulea 3, Succisa pratensis 3, Carex flacca *f. 3, Cirsium tuberosum 2, Lotus dorycnium 2, Centaurea jacea (*timbalii ?) 2, Prunella hyssopifolia 1, Filipendula vulgaris 1, Scorzonera humilis +, Pulicaria dysenterica +,
- autres taxons : Centaurium erythraea *e. 2, Leucanthemum pallens 1, Cervaria rivini +, Danthonia decumbens *d. +, Genista pilosa *p. +, Fraxinus angustifolia *a. j 1, Erica scoparia *s. j 1, Ligustrum vulgare j 1, Arbutus unedo j +, Viburnum tinus j +.

Les relevés 10 et 11 correspondent à une variation d’exposition nord différenciée par Sesleria caerulea et Potentilla erecta. Au voisinage du relevé 4, on a aussi noté la présence du Blackstonio perfoliatae-Isolepidetum cernuae B. Foucault 2019 : Isolepis cernua 2, Linum catharticum 2, Blackstonia perfoliata 1.
Cette association développée à une altitude moyenne de 240 m accueille en moyenne environ 18,5 taxons par relevé. On aurait pu s’attendre à rencontrer cette association dans la Piège audoise, où apparaissent des pelouses parfois massivement dominées par la molinie. Il n’est pas exclu d’en rencontrer à l’avenir, mais pour l’instant on n’a observé que des pelouses des Brometalia erecti présentant de tels faciès.
Parmi les taxons compagnes, les formes juvéniles de plusieurs arbustes témoignent d’une dynamique progressive possible vers un fourré à Erica scoparia, dont le tableau 9 rapporte six relevés. La présence de cette bruyère acidiphile et d’Arbutus unedo incite à placer ce fourré dans les Cytiso villosi-Genistetalia monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 et l’Ericion arboreae (Rivas Mart. et al. 1986) Rivas Mart. 1987 (classe des Pistacio lentisci-Rhamnetea alaterni), bien qu’Erica arborea manque ici, alors que des taxons plutôt caractéristiques des Rhamno catharticae-Prunetea spinosae (Ligustrum vulgare, Juniperus communis subsp. c., Viburnum lantana surtout) le différencient des autres associations connues de cet ordre (de Foucault, 2021b) ; on peut définir ici le Junipero communis-Phillyreetum angustifoliae ass. nov. hoc loco (photo 15), accueillant en moyenne 14,5 taxons par relevé, typus nominis hoc loco : relevé 4 du tableau 9 hoc loco, retranscrit ci-après :
Saint-Martin-le-Vieil (11), « Ficelle », au bord de la D 64, N 43° 18’ 39,9’’, E 2° 09’ 24’’, 218 m, 31 mai 2023, 100 %, 40 m², 16 taxons
- Pistacio-Rhamnetea alaterni, Cytiso-Genistetalia monspessulanae: Erica scoparia *s. 3, Phillyrea angustifolia 2, Arbutus unedo 2, Quercus coccifera 2, Rhamnus alaternus *a. 1, Smilax aspera +, Asparagus acutifolius +, Viburnum tinus +, Lonicera implexa +, Pistacia terebinthus +,
- Rhamno c.-Prunetea spinosae: Ligustrum vulgare 2, Juniperus communis *c. 1, Viburnum lantana 1,
- autres taxons : Fraxinus angustifolia *a. j 3, Quercus ilex j 2, pubescens j .
Sur Saint-Martin-le-Vieil, au voisinage de cette association, on peut encore remarquer le développement d’un bel ourlet inédit à Iris graminea, Filipendula vulgaris, Cervaria rivini, Serratula tinctoria…

Annexe du tableau 8 (tous ces relevés proviennent de l’Aude) – rel. 1 : Saint-Martin-le-Vieil, près de l’abbaye de Villelongue, ‘La Jambe de l’Homme’, 26/07/2020 ; rel. 2 : Saint-Martin-le-Vieil, sur la D 64, 07/10/2014 ; rel. 3 : Alzonne, vallée de la Vernassonne, vers la Migance, 29/07/2022 ; rel. 4 : Axat, nord-est, sur la D 117, vers la Crémade, 05/07/2020 ; rel. 5 et 6 : Alzonne, la Migance, 18/05/2023 ; rel. 7 : Alzonne, les Sesquières, 18/05/2023 ; rel. 8 et 9 : Saint-Martin-le-Vieil, ‘Ficelle’, bord de la D 64, 31/05/2023 ; rel. 10 : Saint-Martin-le-Vieil, ‘Borde Neuve’, D 64, 18/05/2023 ; rel. 11 : Saint-Martin-le-Vieil, D 64, Villelongue, derrière la Tuilerie,18/05/2023.
Annexe du tableau 9 (tous ces relevés proviennent de l’Aude) – rel. 1 : Alzonne, la Migance, 18/05/2023 ; rel. 2 : Alzonne, les Sesquières, 18/05/2023 ; rel. 3 : Saint-Martin-le-Vieil, ‘Borde Neuve’, D 64, 18/05/2023 ; rel. 4 à 6 : Saint-Martin-le-Vieil, ‘Ficelle’, bord de la D 64, 31/05/2023.
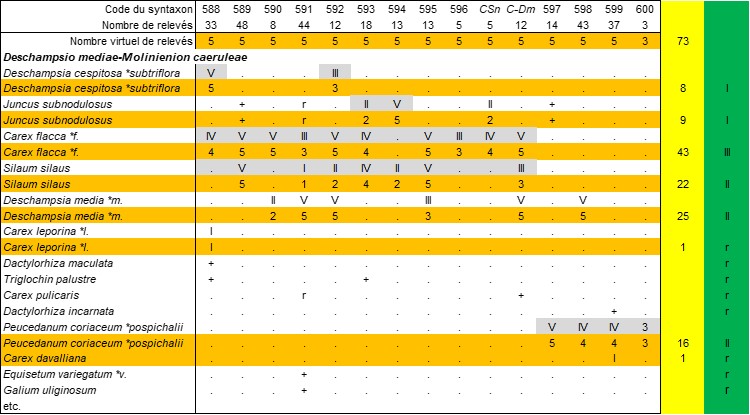
Cette nouvelle association offre l’occasion d’actualiser le Deschampsio mediae-Molinienion caeruleae à partir du tableau 238 de notre thèse de 1984 (tableau 10) ; les numéros de syntaxons 588 à 600 sont ceux utilisés dans ce document de 1984 ; deux syntaxons décrits ultérieurement sont aussi inclus.
Deschampsio mediae-Molinienion caeruleae B. Foucault 2008
Typus suballianciae : Blackstonio perfoliatae-Silaetum silai B. Foucault 2008 (J. Bot. Soc. Bot. France 43 : 44), syntaxon n° 589.
* Groupe de syntaxons à Silaum silaus, Cirsium tuberosum, Lotus maritimus, Genista tinctoria, Plantago maritima subsp. serpentina, Pulicaria dysenterica, Inula salicina…
588. Deschampsio subtriflorae-Molinietum caeruleae Rivas Goday & Borja 1961 nom. mut. hoc loco. (initialement sub Deschampsio refractae-Molinietum caeruleae) ; Espagne centrale.
589. Blackstonio perfoliatae-Silaetum silai B. Foucault 2008 ; centre-ouest de la France jusqu’en Basse-Normandie méridionale (Lemée, 1933 ; de Foucault, 1984) ; photo 16.

590. Cirsio tuberosi-Lotetum maritimi Vanden Berghen 1963 nom. mut. hoc loco (initialement sub Cirsio-Tetragonolobetum) ; Causses.
591 et 592. Potentillo reptantis-Deschampsietum mediae Oberd. 1957 ; 591 – race subcontinentale ; 592 – race occidentale. Après comparaison entre ces deux syntaxons, il convient de séparer plus nettement le second du premier : Galio borealis-Deschampsietum mediae ass. nov. hoc loco, typus nominis hoc loco : relevé 3 du tableau 44 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) publié ci-après sud de Benon (17), ‘Plaisance’, ~ N 46° 11’ 0,0’’, O 0° 48’ 06,5’’, 20 m, 25/06/1981, 90 %, 1 m², 17 taxons
- combinaison caractéristique : Deschampsia media *m. 33, Galium boreale 22, Carex flacca *f. 22, Serratula tinctoria 22, Genista tinctoria 22, Centaurea decipiens 11, Prunella hyssopifolia 12, Cervaria rivini 12, Carex panicea +, Succisa pratensis +2, Lathyrus pannonicus +, Iris reichenbachiana +,
- autres taxons : Brachypodium rupestre *r. 12, Galatella linosyris +, Cytisus lotoides +, Galium verum *v. +, Briza media *m. +.
Le tassement du sol du Cirsio tuberosi-Molinietum caeruleae d’Isère (cf. supra, syntaxon 562) sous l’effet de pressions biotiques le déstructure, le faisant dériver vers le Potentillo reptantis-Deschampsietum mediae :
Soleymieu (38), sous la ferme de Billonay, ~ N 45° 43’ 30,8’’, E 05° 20’ 32’’, 327 m, 14 taxons
– Molinion caeruleae et unités supérieures : Molinia caerulea 3°, Deschampsia media *m. 2, Succisa pratensis 2, Cirsium tuberosum +°, Genista tinctoria +°, Inula salicina +°,
– autres taxons : Carex flacca *f. 2, Prunella vulgaris 2, Agrostis stolonifera 2, Cervaria rivini +°, Lotus corniculatus *c. +, Potentilla reptans +, Trifolium fragiferum +, Briza media *m. +.
Sous ce nom, Rameau et Royer (1978, tableau II) ont décrit un groupement qui s’identifie plutôt au Carici lepidocarpae-Deschampsietum mediae (cf. infra, syntaxon C-Dm).
593. Loto jordanii-Molinietum caeruleae Braun-Blanq. ex B. Foucault & Géhu 1980 ; France méditerranéenne.
594. Junco ancipitis-Galietum debilis Braun-Blanq. 1952 nom. mut. hoc loco (initialement sub Junco-Galietum constricti); France méditerranéenne.
595. Molinietum mediterraneum sensu Donker & Stevelink 1962, nom. illeg. (art. 34a) validé ici sous le nom de Narcisso poetici-Caricetum tomentosae (Donker & Stevelink 1962) nom. nov. hoc loco, typus nominis hoc loco: relevé 2 du tableau II in Donker & Stevelink (1962, Meded. Landbouwh. Wageningen 61 (15) hors texte) ; il est probable que ce soit aussi cette association de France méditerranéenne qui a été observée par de Foucault & Royer (2019 : 90).
596. Groupement à Plantago maritima subsp. serpentina-Euphrasia hirtella ; mérite un rang d’association, mais les données synthétiques d’origine empêchent toute validation ; Dévoluy (1 000 à 1 300 m).
C-Sn. Cirsio tuberosi-Schoenetum nigricantis nov. (tableau 8 hoc loco).
C-Dm. Carici lepidocarpae-Deschampsietum mediae J.-M. Royer & Didier 1996 (Royer & Didier, 1996, tableau XXIV) ; plateau de Langres.
* Groupe de syntaxons à Peucedanum coriaceum subsp. pospichalii, Scilla litardierei, Carex hostiana, Sesleria caerulea, surtout de Croatie, donc sans représentant en France
597. Peucedano pospichalii-Molinietum caeruleae Horvatić 1934.
598. Deschampsietum mediae illyricum (Zeidler 1944) Horvatić 1963, nom. illeg. (art. 34a) que l’on peut renommer Scillo litardierei-Deschampsietum mediae (Horvatić 1963) nom. nov. hoc loco, typus nominis hoc loco: relevé 2 du tableau I in Gaži-Baskova & Trinajstić (1970, Acta Bot. Croatica 29 : entre p. 150 et p. 151) en remplaçant Scilla pratensis par S. litardierei.
599. Lathyro pannonici-Molinietum caeruleae Gaži-Baskova1963.
600. Groupement à Peucedanum coriaceum subsp. pospichalii-Molinia caerulea.
Floristiquement, l‘Allio schoenoprasi-Deschampsietum mediae Laclos & J.-M. Royer 2002 (de Laclos & Royer, 2001, tableau I) ne se place pas dans la présente sous-alliance malgré la présence de Deschampsia media ; il s’agit plutôt d’une pelouse voisine ou dérivée du Poetum badensis J.-M. Royer 1978 (Sedo-Scleranthetea biennis ; Royer & Ferrez, 2018) soumise à une légère humidification édaphique qui favorise l’arrivée des deux taxons mésohygrophiles éponymes, humidification incomplète pour la faire basculer vers cette sous-alliance.
1.7. Le Genisto tinctoriae-Silaenion silai
Il existe encore des associations du Molinion caeruleae qui n’entrent guère dans les sous-alliances précédentes : à la suite d’une première étude sur des pelouses du Boulonnais (de Foucault, 1986b) dans laquelle fut décrit le Dactylorhizo meyeri-Silaetum silai, Catteau (2021) a repris et étendu les investigations de terrain et a finalement retenu deux associations synthétisées dans le tableau 11 :
E-Ss. Epipactido palustris-Silaetum silai Catteau 2021 (Catteau, 2021, tableau 2).
D-Ss. Dactylorhizo fuchsii-Silaetum silai B. Foucault 1986 nom. mut. hoc loco (Catteau, 2021, tableau 7, sub Dactylorhizo meyeri-Silaetum silai) ; photo 17.
Ces deux associations marnicoles nord-atlantiques partagent Silaum silaus, Genista tinctoria, Ophioglossum vulgatum, Dactylorhiza fuchsii, Succisa pratensis, Pulicaria dysenterica, Centaurea decipiens, Neottia ovata, Carex flacca subsp. f., ensemble caractéristique d’une sous-alliance originale, le Genisto tinctoriae-Silaenion silai suball. nov. hoc loco, typus nominis hoc loco : Epipactido palustris-Silaetum silai Catteau 2021 (Botanique 7 : 41). On peut émettre l’hypothèse que cette sous-alliance atteint aussi le sud-est du Royaume-Uni (comté du Kent).

2. Le Juncion acutiflori
2.1. Vue d’ensemble de l’alliance
Deux « Juncion acutiflori » ont successivement été décrits, le Juncion acutiflori Braun-Blanq. ex Braun-Blanq. & Tüxen 1952 (Veröff. Geobot. Inst. E.T.H. Stiftung Rübel Zürich 25 : 292) [syn. : Juncion acutiflori Braun-Blanq. 1947 (Les groupements végétaux supérieurs de la France : 26) nom. inval.] et l’Anagallido tenellae-Juncion acutiflori Braun-Blanq. 1967 (Vegetatio XVI : 39). Il paraît acceptable de considérer le nom le plus ancien et de ramener l’Anagallido–Juncion acutiflori au rang de sous-alliance, suivant en cela Julve (1993) puis Bardat et al. (2004).
Dans la synthèse de de Foucault & Géhu (1980) affinée dans la thèse de 1984 déjà citée, puis les validations de 2008a et 2019b, on peut y distinguer trois sous-alliances : Anagallido tenellae-Juncenion acutiflori B. Foucault 2008, Caro verticillati-Juncenion acutiflori B. Foucault & Géhu 1980 et Polygono bistortae-Juncenion acutiflori B. Foucault & Géhu ex B. Foucault 2008. Le type de l’alliance retenue étant obligatoirement le Jacobaeo aquaticae-Juncetum acutiflori Braun-Blanq. & Tüxen 1952 (Veröff. Geobot. Inst. E.T.H. Stiftung Rübel Zürich 25 : 293, sub Senecioneto-Juncetum acutiflori) d’Irlande, traditionnellement inclus dans le Caro verticillati-Juncenion acutiflori, cette sous-alliance doit dès lors être dénommé Eu-Juncenion acutiflori (cf. infra).On va reprendre ces unités pour les actualiser et y ajouter quelques syntaxons récemment décrits sur la base des tableaux 220 à 223 de la thèse de 1984.
2.2. Le Lysimachio tenellae-Juncenion acutiflori (Braun-Blanq. 1967) B. Foucault 2008 (J. Bot. Soc. Bot. France 43 : 58) nom. mut. hoc loco
[corresp. syntax. : Anagallido tenellae-Juncion acutiflori Braun-Blanq. 1967 (Vegetatio XIV (1-4) : 39)]
Typus allianciae : Hyperico undulati-Juncetum acutiflori Teles 1970 (Agron. Lusit. 31 (1-2) : 82) désigné pour l’alliance par de Foucault & Géhu (1980, Colloq. Phytosoc. VII : 139).
Les syntaxons connus de cette unité sont synthétisés dans le tableau 12.
344. Erico tetralicis-Narthecietum ossifragi Braun-Blanq. 1967 ; Pays basque espagnol.
345. Groupement à Galium uliginosum-Juncus acutiflorus; Pays basque espagnol.
346. Arnicetum atlanticae Bellot & Casaseca 1956 ; Galice.
347. Loto pedunculati-Juncetum acutiflori (Tüxen & Oberd. 1958) B. Foucault & Géhu 1980 nom. mut. hoc loco (initialement sub Loto villosi-Juncetum acutiflori; syn. : Senecioni aquatici-Juncetum acutiflori sensu Tüxen & Oberd. 1958, nom. illeg. non Braun-Blanq. & Tüxen 1952) ; nord-ouest de la péninsule Ibérique.
348. Hyperico undulati-Juncetum acutiflori Teles 1970 ; Portugal septentrional.
349. Thysselino lancifolii-Juncetum acutiflori Teles 1970 nom. mut. hoc loco (initialement sub Peucedano lancifolii-Juncetum acutiflori) ; Portugal septentrional.
350. Lysimachio tenellae-Pinguiculetum lusitanicae Rivas Goday ex B. Foucault 2008 nom. mut. hoc loco (initialement sub Anagallido tenellae-Pinguiculetum lusitanicae) ; association ibéro-franco-atlantique pénétrant en France occidentale dans l’aire de l’Eu–Juncenion acutiflori; photo 18.

351. Sibthorpio europaeae-Pinguiculetum lusitanicae Ladero & Velasco in Velasco 1980 ; monts de Tolède (Espagne) ; avec Sibthorpia europaea, Myosotis stolonifera, Wahlenbergia hederacea, cette association converge vers la végétation crénophile du Wahlenbergio hederaceae-Sibthorpion europaeae B. Foucault 2018.
C-Cp. Carici camposi-Calthetum palustris Braun-Blanq. 1967 prov. (Braun-Blanquet, 1967, tableau 16) ; Pays basque espagnol.
T-Mw. Groupement à Trocdaris verticillatum-Myosotis welwitschii (Braun-Blanquet, 1967, tableau 18) ; Pays basque espagnol.
352. Groupement à Baldellia ranunculoides-Lysimachia tenella Rivas Goday 1964 ; Espagne sud-occidentale (province de Badajoz).
353. Groupement à Juncus bulbosus-Wahlenbergia hederacea (composante vivace du groupement à Isolepis setacea-Wahlenbergia hederacea Braun-Blanq. 1967) ; Pays basque espagnol.
354. Lobelio urentis-Lotetum pedunculati Rivas Goday 1964 ; Espagne sud-occidentale (province de Badajoz).
355. Hyperico undulati-Scirpoidetum holoschoeni Rivas Goday 1964 ; Espagne sud-occidentale (province de Badajoz).
356. Succiso pratensis-Centaureetum rivularis Rivas Goday, Mayor, Lardo & Izco 1965 ; Espagne occidentale.
On ne retient pas ici les Hyperico undulati-Juncetum conglomerati J.C. Costa et al. 2004 et Carici cedercreutzii-Juncetum effusi J.C. Costa et al. 2004 de Madère très faiblement reliés à cette sous-alliance (Costa et al., 2004) ; ils semblent plutôt relever des Agrostietea stoloniferae. Malgré sa position en Catalogne, le Carici punctatae-Juncetum acutiflori O. Bolòs 1959 s’intègre mal à cette sous-alliance, et d’ailleurs guère mieux dans la suivante (de Bolòs, 1959, 1979).
2.3. L’Eu-Juncenion acutiflori
[syn. : Caro verticillati-Juncenion acutiflori B. Foucault & Géhu 1980 (Colloq. Phytosoc. VII : 141)]
Les syntaxons connus de cette unité sont synthétisés dans le tableau 13.
357 à 360. Caro verticillati-Juncetum acutiflori auct. ; 357 – race franco-basque ; 358 – race armoricaine à Thysselinum lancifolium ; 359 – race eu-atlantique ; 360 – race atlantique montagnarde ; le nom peut être muté en Trocdario verticillati-Juncetum acutiflori nom. mut. hoc loco.
361. Jacobaeo aquaticae-Juncetum acutiflori Braun-Blanq. & Tüxen 1952 nom. mut. hoc loco (Braun-Blanquet & Tüxen, 1952, tableau 31 : Senecieto-Juncetum acutiflori subass. à Carum verticillatum et subass. à Ranunculus repens) ; Irlande.
T-No. Trocdario verticillati-Narthecietum ossifragi B. Foucault 2018 (de Foucault, 2018, tableau 3) ; Montagne Noire audoise.
M-Tv. Mentho arvensis-Trocdarietum verticillati B. Foucault & Philippe 1989 nom. mut. hoc loco (de Foucault & Philippe, 1989, tableau 4, sub Mentho arvensis-Caretum verticillati) ; Morvan.
362. Caro verticillati-Molinietum caeruleae (G. Lemée 1937) J.-M. Royer, Felzines, Misset & Thévenin 2006 ; le relevé suivant est inédit :
Combreux (45), route forestière au sud-est du bourg, 27/05/1995, 15 taxons
- combinaison caractéristique : Molinia caerulea 5, Potentilla erecta 2, Agrostis canina *c. 2, Calluna vulgaris 2, Danthonia decumbens *d. 1, Erica ciliaris 1, Ulex minor 1, Trocdaris verticillatum +, Dactylorhiza maculata +, Juncus conglomeratus +, Scorzonera humilis +, Pedicularis sylvatica *s. +,
- autres taxons : Pteridium aquilinum 2, Frangula alnus *a. j 1, Ulex europaeus *e. j +,
moliniaie souvent dense, de contact avec des landes hygrophiles dont quelques chaméphytes pénètrent cette végétation. Le nom peut être muté en Trocdario verticillati-Molinietum caeruleae nom. mut. hoc loco.
363. Aristaveno setaceae-Agrostietum caninae B. Foucault 1980 nom. mut. hoc loco (initialement sub Deschampsio setaceae-Agrostietum caninae) ; France occidentale, souvent au contact de l’Hyperico elodis-Potametum polygonifolii Braun-Blanq. & Tüxen 1952 et de l’Eleocharitetum multicaulis P. Allorge ex Tüxen 1937 ; le tableau 14 en rapporte deux relevés inédits du Morbihan [rel. 1 – Carnac (56), ‘Crucuny’, 09/08/1986 ; rel. 2 – Carnac (56), ‘Le Hahon’, 09/08/1986].
364. Oenantho fistulosae-Agrostietum caninae B. Foucault 2008 ; France occidentale.
365 à 367. Cirsio dissecti-Scorzoneretum humilis B. Foucault 1980 ; 365 – race à Thysselinum lancifolium ; 366 – race eu-atlantique à Centaurea decipiens/nigra (photo 19) ; 367 – race subcontinentale à Centaurea jacea. Le relevé inédit qui suit appartient à la deuxième race :
Rosnay (36), queue de l’étang Pifaudière, ~ N 46° 43’ 28’’, E 1° 13’ 28,9’’, 105 m, 16/05/2000, 29 taxons
- combinaison caractéristique : Juncus acutiflorus 4, Cirsium dissectum 3, Trocdaris verticillatum 3, Scorzonera humilis 2, Achillea ptarmica *p. 2, Agrostis canina 2, Ranunculus flammula *f. 1, Carex panicea 1, leporina *l. +, Hydrocotyle vulgaris +, Lotus pedunculatus +, Scutellaria minor +, Molinia caerulea +, Juncus conglomeratus +, Veronica scutellata +,
- autres taxons : Mentha aquatica 1, Lycopus europaeus 1, Eleocharis palustris s. l. 1, uniglumis +, Cardamine pratensis +, Carex flacca *f. +, Holcus lanatus *l. +, Anthoxanthum odoratum +, Oenanthe peucedanifolia +, Galium palustre +, Potentilla reptans +, Alisma plantago-aquatica +, Lythrum salicaria +, Frangula alnus *a. j + ;
cette prairie hygrophile oligotrophile sur substrat paratourbeux est souvent en lien systémique avec des prairies hygrophiles plus eutrophiles, notamment l’Oenantho peucedanifoliae-Brometum racemosi B. Foucault 1981, dont le premier taxon éponyme est présent ici, prairie par ailleurs attestée dans cette région de Brenne (de Foucault et al., 1992a).

368. Groupement à Lychnis flos-cuculi-Trocdaris verticillatum B. Foucault 1984 nom. ined. ; la jonçaie oligotrophile franco-basque est assez différente du Cirsio dissecti-Scorzoneretum humilis, son homologue eu-atlantique, et mérite d’être distinguée sous le nom de Lychnido floris-cuculi-Trocdarietum verticillati ass. nov. hoc loco, typus nominis hoc loco : relevé 4 du tableau 71 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) publié ci-après :
Louhossoa (64), Zilantzenea, ~ N 43° 18’ 50,1’’, O 1° 20’ 21,2’’, 113 m, 07/05/1982, 90 %, 50 m², 27 taxons
- combinaison caractéristique : Juncus acutiflorus 33, Carex leporina *l. 32, Trocdaris verticillatum 12, Juncus conglomeratus 12, Luzula multiflora *m. 12, Lotus pedunculatus 12, Carex laevigata 12, panicea +, C. echinata *e. +, Potentilla erecta +, Succisa pratensis r,
- autres taxons : Trifolium repens 32, Anthoxanthum odoratum 32, Holcus lanatus *l. 22, Agrostis capillaris 22, Festuca rubra *r. 22, Taraxacum 22, Carex pilulifera *p. 11, Poa trivialis *t. 12, Gaudinia fragilis 12, Ajuga reptans 12, Plantago lanceolata 12, Trifolium pratense *p. 12, Lolium perenne 11, Lychnis flos-cuculi *f. +, Cirsium palustre +, Hypochaeris radicata +.
369. Lobelio urentis-Agrostietum caninae B. Foucault 2008. Les trois relevés ci-après sont inédits :
a) Montrevault-sur-Èvre (49), forêt de Leppo, ~ N 47° 14’ 34,6’’, O 1° 05’ 37,6’’, 106 m, 18/07/1990, 10 m², 16 taxons
- combinaison caractéristique : Lobelia urens 2, Danthonia decumbens *d. 2, Molinia caerulea 2, Potentilla erecta 2, Succisa pratensis 2, Agrostis canina *c. 1, Pedicularis sylvatica *s. 1, Carex demissa 1, laevigata +,
- autres taxons : Agrostis capillaris 3, Calluna vulgaris 1, Festuca filiformis 1, Polygala serpyllifolia 1, Prunella vulgaris +, Cytisus scoparius *s. j +, Juncus tenuis + ;
b) Combreux (45), route forestière au sud-est, 27/05/1995, 16 taxons
- combinaison caractéristique : Molinia caerulea 3, Juncus conglomeratus 3, acutiflorus 2, Agrostis canina *c. 2, Carex demissa 2, C. panicea 1, Scutellaria minor 1, Potentilla erecta 1, Luzula multiflora *m. 1, Lobelia urens +, Juncus bulbosus +,
- autres taxons : Calluna vulgaris 1, Genista anglica +, Myosotis scorpioides +, Carex pilulifera *p. +, Hypericum pulchrum + ;
c) Saint-Martin d’Abbat (45), forêt domaniale d’Orléans, 27/05/1995, 17 taxons
- combinaison caractéristique : Molinia caerulea 3, Agrostis canina *c. 2, Potentilla erecta 2, Carex demissa 2, Lobelia urens 1, Juncus conglomeratus 1, acutiflorus 1, Luzula multiflora *m. 1, Lotus pedunculatus +, Carex pallescens +,
- autres taxons : Calluna vulgaris 2, Carex pilulifera *p. +, Agrostis capillaris +, Danthonia decumbens *d. +, Polygala serpyllifolia +, Rubus +, Hypericum pulchrum + ;
cette végétation ouverte thermo-atlantique sur sol plutôt minéral oligotrophe inondable est souvent en mosaïque avec une végétation thérophytique (par exemple en a : Radiola linoides 1, Hypericum humifusum 1, Cicendia filiformis +, Lysimachia minima +, soit le Cicendietum filiformis P. Allorge 1922).
370. Carici punctatae-Agrostietum caninae B. Foucault 2017 (de Foucault, 2017 : 343) ; sud-ouest de la France ; photo 20.
371. Carici binervis-Agrostietum caninae B. Foucault 2008 ; association eu-atlantique atteignant la Montagne Noire (de Foucault, 2022).

Cv-Ac. Caropsio verticillato-inundatae-Agrostietum caninae B. Foucault 2008 (de Foucault, 1984, tableau 133) ; Landes de Gascogne.
L’association 357-360 offre des difficultés de validation, d’où l’indication provisoire ‘auct.’, on va essayer de les résoudre ici. Elle a été définie initialement par Lemée (1937 : 128) sous le nom de Juncetum silvatici, un nom illégitime (art. 31, non Juncetum sylvatici Braun-Blanq. 1915) ; Clément (1978 : 105, inédit) utilise à tort le nom de Peucedano lancifolii-Juncetum acutiflori Teles 1970 qui relève de la sous-alliance précédente (cf. supra, syntaxon n° 349). Oberdorfer (1979, Pflanzensoziologische Exkursionsflora : 37) utilise le nom de Caro verticillati-Juncetum acutiflori, mais c’est un nomen nudum invalide, pourtant retenu par Delassus et al. (2014) ; ce nom a été repris par moi-même (thèse, 1984 : 81) mais sous forme inédite. Ce nom est parfois attribué à Korneck (1962), mais cet auteur n’use que du nom inadéquat et illégitime de ‘Molinietum atlanticum Lemée 1937’. Oberdorfer (1983, Süddeutsche Pflanzengesellschaften III : 384) utilise aussi Caro verticillati-Juncetum acutiflori sans plus de validation, nom repris par Robbe (1993). Malgré un usage courant, il semble donc bien que cette association soit toujours invalide ; on la valide définitivement ici : Trocdario verticillati-Juncetum acutiflori (G. Lemée 1937) Oberd. ex ass. nov. hoc loco,
typus nominis hoc loco de l’association et du typicum : relevé 23 du tableau 13 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) publié ici :
forêt de Saint-Sever (commune de Noues-de-Sienne, 14), ‘La Forge’, ~ N 48° 48’ 06,6’’, O 1° 02’ 48,9’’, 257 m, 03/09/1982, 90 %, 20 m², 22 taxons
- combinaison caractéristique : Juncus acutiflorus 32, Trocdaris verticillatum 22, Hydrocotyle vulgaris 22, Carex echinata *e. +2, Scutellaria minor +, Wahlenbergia hederacea +2, Epilobium palustre (+), Viola palustris 11, Agrostis canina *c. 22, Ranunculus flammula *f. +, Potentilla erecta 12, Cirsium dissectum +2, Molinia caerulea 22, Carex panicea 12, nigra 12, Succisa pratensis 11, Lotus pedunculatus +,
- autres taxons : Juncus effusus 22, Anthoxanthum odoratum +, Juncus bulbosus +, squarrosus +, Nardus stricta + ;
typus nominis hoc loco du juncetosum subnodulosi subass. nov. hoc loco, sous-association neutrophile différenciée par le taxon éponyme, Dactylorhiza incarnata, Carex hostiana… : relevé 14 du tableau 64 in de Foucault (1984, Systémique, structuralisme et synsystématique… hors texte) publié ici :
Carentan-les-Marais (50), réserve naturelle du Marais de la Taute, ~ N 49° 15’ 55,2’’, O 01° 13’ 15,1’’, 1 m, 17/06/1982, 100 %, 25 m², 27 taxons
- combinaison caractéristique : Molinia caerulea 23, Agrostis canina *c. 22, Hydrocotyle vulgaris 22, Carex panicea 23, Cirsium dissectum 12, Juncus acutiflorus 12, Trocdaris verticillatum 12, Carex pulicaris 13, echinata *e. 13, Eriophorum angustifolium *a. 11, Dactylorhiza maculata 12, Ranunculus flammula *f. +, Potentilla erecta 12, Scorzonera humilis +, Succisa pratensis +, Lotus pedunculatus +,
- taxons différentiels du juncetosum subnodulosi: Juncus subnodulosus 32, Carex hostiana 22, Dactylorhiza incarnata +,
- autres taxons : Anthoxanthum odoratum 22, Holcus lanatus *l. 12, Festuca rubra *r. 13, Cardamine pratensis +, Briza media *m. 12, Cirsium palustre +, Schedonorus arundinaceus *a. +, Quercus robur j +.
2.4. Le Bistorto officinalis-Juncenion acutiflori B. Foucault & Géhu ex B. Foucault 2008 nom. mut. hoc loco (J. Bot. Soc. Bot. France 43 : 59, sub Polygono bistortae -Juncenion acutiflori)
[syn. : Polygono bistortae-Juncenion acutiflori B. Foucault & Géhu 1980 (Colloq. Phytosoc. VII : 145) nom. inval. (art. 3b)]
Typus suballianciae : Pediculari mixtae-Molinietum caeruleae Gruber 1978 (La végétation des Pyrénées ariégeoises et catalanes occidentales : 78) désigné par le validateur (2008 : 59) ; ici, on confirme ce choix en suivant Corriol et al. (2023) pour estimer que cette thèse est une publication effective.
Les syntaxons connus de cette unité sont synthétisés dans le tableau 15.
373. Pediculari mixtae-Molinietum caeruleae Gruber 1978 ; Pyrénées centro-occidentales.
374 et 375. Ligulario sibiricae-Molinietum caeruleae B. Foucault & Géhu ex B. Foucault 2019 [syn. : Ligulario sibiricae-Molinietum caeruleae B. Foucault & Géhu 1980 (Colloq. Phytosoc. VII : 145) nom. inval., nom finalement validé in de Foucault (2019b, J. Bot. Soc. Bot. France 86 : 97)], caractérisé par Ligularia sibirica (photo 21) ; 374 – race du Capcir, avec Juncus balticus subsp. pyrenaeus (photo 22) ; 375 – race des monts Dore.


376 et 377. Epikeroto pyrenaei-Juncetum acutiflori (Luquet 1926) B. Foucault & Julve in B. Foucault, Beaufils, Gilbert & Lépagnole 2021 ; 376 – race des monts Dore ; 377 – race du Vivarais ; atteint les Pyrénées audoises (de Foucault et al., 2021).
E-Sh. Epikeroto pyrenaei-Scorzoneretum humilis B. Foucault 1986 nom. mut. hoc loco ; Aubrac (de Foucault, 1986c, sub Selino pyrenaei-Scorzoneretum humilis) et monts Dore (Julve, 1983).
P-Sh. Prunello hastifoliae-Scorzoneretum humilis B. Foucault 1986 ; Aubrac.
2.5. Le Junco conglomerati-Molinienion caeruleae (V. Westh. 1969) suball. nov. et stat. nov. hoc loco
[syn. : Junco-Molinienion caeruleae (V. Westh. 1969) B. Foucault & Géhu 1980 (Colloq. Phytosoc. VII : 143) nom. inval. (art. 3o, 5a, 8), Juncenion acutiflori Delpech in Bardat et al. (2004, Patrimoines naturels 61 : 65) nom. prov. ; corresp. syntax. : Junco-Molinion caeruleae V. Westh. in V. Westh. & Den Held 1969 (Plantengemeenschappen in Nederland)]
Typus nominis hoc loco : Cirsio dissecti-Molinietum caeruleae G. Sissingh & De Vries (1942) 1946.
Sous-alliance d’appauvrissement chorologique vers les régions plus continentales : raréfaction des taxons ibéro- à eu-atlantiques (Trocdaris verticillatum, Lysimachia tenella, Wahlenbergia hederacea…), ainsi que des taxons d’optimum montagnard (Epikeros pyrenaeus, Veratrum album), pouvant être différenciée par Sanguisorba officinalis. Les syntaxons connus de cette unité sont synthétisés dans le tableau 16, à l’exception du n° 386, placé dans le Nardo strictae-Juncion squarrosi donc non repris ici, et du n° 378.
Ce dernier correspond au groupement à Juncus acutiflorus-Eriophorum gracile, un rarissime bas-marais flottant localisé dans l’aire générale de l’Eu–Juncenion acutiflori (une tourbière de Mayenne) qui trouve mieux sa place dans les Scheuchzerietalia palustris Nordh. 1936 et le Caricion lasiocarpae Vanden Berghen in Lebrun et al. 1949 sous le nom de Junco acutiflori-Eriophoretum gracilis ass. nov. hoc loco, typus nominis hoc loco : relevé in Corillion & Planchais (1963, Pollen & Spores V (2) : 379). Le Caricion lasiocarpae peut se subdiviser en trois-sous-alliances selon la réaction ionique du substrat et l’étage climatique ; en l’absence de synthèse récente sur cette alliance, on pose l’hypothèse que la sous-alliance autonyme en est le volet acidiphile (Eu-Caricenion lasiocarpae ; syn. : Junco acutiflori-Caricenion lasiocarpae J.-M. Royer in Bardat et al. 2004 nom. prov. inval.), à laquelle appartient alors l’association de Mayenne. Les deux autres sous-alliances sont plutôt neutro-basiphiles, le Trichophorenion alpini Julve ex J.-M. Royer in Bardat et al. 2004 nom. mut. hoc loco (initialement sub Eriophorenion alpini) boréo-montagnard et le Junco subnodulosi-Caricenion lasiocarpae (Julve 1993) J.-M. Royer in Bardat et al. 2004 nom. prov. inval. planitiaire. On peut valider ici cette dernière sous-alliance : Junco subnodulosi-Caricenion lasiocarpae Julve ex suball. nov. hoc loco, typus nominis hoc loco : Junco subnodulosi-Caricetum lasiocarpae B. Foucault 2008 (J. Bot. Soc. Bot. France 43 : 48).
379. Comaro palustris-Juncetum acutiflori (Braun-Blanq. 1915) H. Passarge 1964 ; nord-est de l’Allemagne.
380. Crepido paludosae-Juncetum acutiflori Oberd. 1957 ; sud-ouest de l’Allemagne.
381-382. Cirsio dissecti-Molinietum caeruleae G. Sissingh & De Vries (1942) 1946 ; 381 – des plaines subatlantiques ; 382 – d’Allemagne orientale.
383. Groupement à Ophioglossum vulgatum-Agrostis canina ; original, mais non typifiable ; Pays-Bas du Nord.
384. Groupement à Epikeros pyrenaeum-Viola palustris ; semble remplacer l’Epikeroto pyrenaei-Scorzoneretum humilis (syntaxon E-Sh dans la sous-alliance précédente) dans des régions du Vivarais oriental (Delpech, 1980) ; on propose de l’élever au rang d’association : Epikeroto pyrenaei-Caricetum echinatae B. Foucault & Delpech ass. nov. hoc loco, typus nominis hoc loco: relevé 29 du tableau in Delpech (1980, Colloq. Phytosoc. VII hors texte).
385. Caricetum echinato-paniceae (Birse & Robertson 1976) Birse 1980 (Birse, 1980, tableau p. 40-42) ; Écosse.
Tx. Juncetum acutiflori sensu Tüxen 1937 (Tüxen, 1937 : 64) ; nord-est de l’Allemagne.
P-Ja. Potentillo erectae-Juncetum acutiflori Birse & Robertson 1976 (Birse, 1980, tableau p. 81-84) ; Écosse.
387 et 388. Junco acutiflori-Molinietum caeruleae Tüxen & Preising 1951 ; 387 – race euro-subatlantique ; 388 – race irlandaise.
389. Eleocharito multicaulis-Agrostietum caninae ass. nov. hoc loco, typus nominis hoc loco: relevé 442 du tableau 5 in Bournérias & Maucorps (Doc. Phytosoc. 9-14 : 35) ; Hauts-de-France.
390. Aristaveno setaceae-Caricetum serotinae Pietsch 1964 ; Allemagne.
391. Ranunculo repentis-Juncetum acutiflori B. Foucault & Wattez ass. nov. hoc loco, typus nominis hoc loco : relevé 26 du tableau 3 in Wattez (1978, Colloq. Phytosoc. V hors texte) ; Hauts-de-France.
J-Cn. Junco articulati-Caricetum nigrae Braun-Blanq. & Tüxen 1952 (de Foucault, 1984, tableau 116, sub Potentillo anserinae-Caricetum fuscae) ; nommé « groupement à Carex fusca-Potentilla anserina » dans le tableau 35 de Braun-Blanquet & Tüxen (1952), mais bien en tant qu’association dans le texte des mêmes auteurs (en p. 306) ; association à caractère plutôt subatlantique isolée dans un contexte général eu-atlantique (littoral du Cotentin, de l’Irlande, de l’Écosse, de la Suède).
392. Caricetum trinervi-nigrae V. Westh. 1947 nom. mut. hoc loco (initialement sub Caricetum trinervi-fuscae) ; du littoral du Pas-de-Calais aux Pays-Bas ; combinaison de Carex trinervis (photo 23) et C. nigra.

393. Ophioglosso azorici-Agrostietum caninae Géhu ex B. Foucault 1988 (de Foucault, 1988, tableau 13) ; littoral du Pas-de-Calais et platières de Fontainebleau ; photo 24.

394. Carici demissae-Agrostietum caninae B. Foucault 2022 ; association nord- et subatlantique.
395. Groupement à Carex binervis-Molinia caerulea.
396. Nardo strictae-Caricetum binervis Braun-Blanq. & Tüxen 1952 ; Irlande.
H-Lt. Hydrocotylo vulgaris-Lysimachietum tenellae B. Foucault, Wattez & Santune ex B. Foucault 2008 nom. mut. hoc loco (de Foucault et al., 1999, tableau 7 : relevés 73, 75 et E3), rarissime association vicariante nord-atlantique du Lysimachio tenellae-Pinguiculetum lusitanicae (syntaxon 350 dans le Lysimachio tenellae-Juncenion acutiflori) ; Hauts-de-France, puis reconnue aussi en Haute-Normandie (de Foucault, 2008b : 45) ; photo 25.

C-Ac. Le Carici pallescentis-Agrostietum caninae ass. nov. hoc loco est une prairie acidiphile oligotrophile développée sur pseudogley, notamment après des défrichements forestiers suivis d’une humidification édaphique, dont le tableau 9 in de Foucault (1995b) rapporte treize relevés du département du Nord, typus nominis hoc loco : relevé 11 du tableau 9 in de Foucault (1995b, Bull. Soc. Bot. N. France 48 (1) : 18) (photos 26 et 27). Cette association est signalée aussi de ce département par de Foucault (1993a, liste p. 48) et existe dans le département voisin de l’Aisne (de Foucault, 1994, tableau IX : relevés 2 à 5), en Argonne (un relevé inédit), jusque dans les Pyrénées audoises (de Foucault, 2019 : 49) ; Gelez et al. (2015 : 43) en rapportent un de plus du nord de la France. Les relevés suivants, encore proches d’un ourlet oligotrophile, sont inédits :
a) Prunay-Cassereau (41), forêt de Prunay, vers la Linoterie, ~ N 47° 42’ 55,7’’, E 0° 53’ 45,2’’, 144 m, 19/07/1990, 17 taxons
- combinaison caractéristique : Agrostis canina *c. 1, Carex pallescens +, Juncus conglomeratus +, Luzula multiflora *m. +,
- autres taxons : Carex pilulifera *p. 3, Veronica officinalis 2, Teucrium scorodonia 2, Quercus petraea *p. j 2, Rubus 2, Hedera helix 2, Agrostis capillaris 1, Carpinus betulus j 1, Hypericum pulchrum 1, Calluna vulgaris 1, Melampyrum pratense +, Luzula pilosa +, Avenella flexuosa *f. + ;
b) Ingrannes (45), forêt domaniale d’Orléans, vers le carrefour des Huit Routes, ~ N 48° 01’ 20,7’’, E 2° 15’ 21,9’’, 141 m, 20/05/1997, 90 %, 150 m², 25 taxons
- combinaison caractéristique : Molinia caerulea 5, Juncus conglomeratus 1, Luzula multiflora *m. 1, congesta +, Agrostis canina *c. +, Carex pallescens +, Juncus effusus +, J. acutiflorus +, Potentilla erecta +, Lotus pedunculatus +,
- autres taxons : Calluna vulgaris 1, Carex pilulifera *p. 1, Rubus 1, Hypericum pulchrum 1, Agrostis capillaris +, Hypochaeris radicata +, Calamagrostis epigejos *e. +, Teucrium scorodonia +, Lonicera periclymenum *p. +, Quercus petraea *p. j +, Anthoxanthum odoratum +, Pteridium aquilinum +, Populus tremula j +, Veronica officinalis +, Lysimachia vulgaris +.


3. Conclusion
En définitive, avec ces nouvelles données, on propose de structurer les alliances du Molinion caeruleae et du Juncion acutiflori selon le schéma synsystématique suivant (tableau 17) :
Molinion caeruleae W. Koch 1926
A-Mc. Allio angulosi-Molinienion caeruleae B. Foucault & Géhu 1980 (tableau 4)
Eu-Mc. Eu-Molinienion caeruleae (tableau 6)
D-Mc. Deschampsio mediae-Molinienion caeruleae B. Foucault 2008 (tableau 10)
G-Ss. Genisto tinctoriae-Silaenion silai nov. (tableau 11)
Juncion acutiflori Braun-Blanq. ex Braun-Blanq. & Tüxen 1952
L-Ja. Lysimachio tenellae-Juncenion acutiflori (Braun-Blanq. 1967) B. Foucault 2008 (tableau 12)
Eu-Ja. Eu-Juncenion acutiflori (tableau 13)
B-Ja. Bistorto officinalis-Juncenion acutiflori B. Foucault & Géhu ex B. Foucault 2008 (tableau 15)
Ja-Mc. Junco acutiflori-Molinienion caeruleae B. Foucault 2019 (tableau 7)
Jc-Mc. Junco conglomerati-Molinienion caeruleae (V. Westh. 1969) B. Foucault nov. (tableau 16)
S-Mc. Serratulo tinctoriae-Molinienion caeruleae B. Foucault 2008 nom. mut. (tableau 2)
Annexe : Le tableau phytosociologique comme outil heuristique de détermination
Dans le domaine de la taxonomie, déterminer c’est attacher à un individu ou une catégorie d’individus se ressemblant fortement (un taxon) nouveau une catégorie et un nom connus. Pour faciliter la reconnaissance des plantes en botanique, on a introduit l’usage des flores, ouvrages permettant d’accéder à l’ensemble des idiotaxons d’un territoire donné.
Parallèlement, on peut alors concevoir l’établissement de documents permettant la reconnaissance de la végétation [ensemble des sytaxons élémentaires (Sy-E T) ou, au moins, des associations végétales] d’un territoire donné au moyen de clés dites synsystématiques. Normalement, comme on le fait en botanique, on devrait pour cela s’appuyer sur la possibilité de hiérarchiser les invariants floristiques caractérisant les unités synsystématiques, en allant des plus élevées aux plus basses :
démarche théorique, uniquement floristique
- invariants floristiques
pourtant, cette démarche est très exceptionnellement suivie : on la reconnaît par exemple dans une clé floristique des landes ibériques établie par Rivas-Martínez (1979). On peut s’appuyer par ailleurs sur le fait que chaque Sy-E T est relié à une synécologie précise en une loi phytosociologique qualitative (de Foucault, 1997a), les invariants synécologiques étant susceptibles d’être eux-mêmes hiérarchisés ; la clé utilise alors cette hiérarchie pour mener progressivement à une dichotomie ultime au niveau de laquelle on passe finalement au Sy-E en s’appuyant sur la loi phytosociologique connue :
démarche concrète, uniquement écologique
invariants écologiques ![]()
Disons toutefois que cette démarche quasi (c’est-à-dire à l’exception de son dernier stade) synécologique est elle-même rarement suivie : on la retrouve surtout dans les clés de détermination du Guide des groupements végétaux du Bassin parisien (Bournérias, 1968, 1984 ; Bournérias et al., 2001).
Le plus souvent, concrètement, la démarche utilisée est mixte, puisque les invariants synécologiques et synfloristiques s’épaulent dans les dichotomies pour déboucher finalement sur le syntaxon cherché ou, plus exactement, sur la loi phytosociologique dont un des éléments est le syntaxon cherché :
démarche concrète, mixte
invariants écologiques et floristiques ![]()
Historiquement, le premier essai paraît être celui de Molinier (1934), suivi de ceux de Meltzer & Westhoff (1942), Bournérias (1949), Braun-Blanquet (1956), Guinochet (1970), Rameau & Royer (1972), Barbe (1972), Guinochet & de Vilmorin (1973), Corriol (2008)…
En seconde partie de cette annexe, on souhaite présenter une approche plus globale de la détermination, à l’aide de tableaux phytosociologiques synthétiques préparés sous le logiciel Excel® (il faudrait voir si la démarche est applicable aussi sur les logiciels libres de type OpenOffice) en prenant le cas du tableau X de Koch (1926, Molinietum caeruleae) et du tableau synthétique 17.
Le principe est de considérer la colonne synthétique issue du syntaxon à déterminer et de l’injecter dans le tableau synthétique taxon par taxon. Dans les végétations riches en taxons et en syntaxons, retrouver les taxons peut devenir très vite fastidieux, même en utilisant la fonction Rechercher… du logiciel. Pour faciliter et accélerer cette opération, on peut introduire à l’extrême gauche du tableau synthétique ce que l’on peut dénommer une colonne pivot en commençant par les chiffres 1, puis 2 et en incrémentant automatiquement la suite jusqu’au dernier taxon (tableau 18a : colonne A marquée en couleur). Ensuite on sélectionne la totalité du tableau (ici colonnes A à H), puis on applique successivement les fonctions Trier et filtrer, Tri personnalisé…, Trier par colonne B, de A à Z (si le titre du tableau est mis en forme selon la fonction Fusionner et centrer, il faut annuler cette mise en forme en cliquant dessus) ; on obtient alors le tableau 18b dans lequel on peut aisément entrer les présences de la colonne à déterminer puisque les taxons sont rangés alphabétiquement, surtout si de plus on a ordonné les taxons de la colonne à déterminer selon le même ordre (tableau 18c, colonne colorée). Dans une dernière manipulation, on sélectionne à nouveau tout le tableau sans oublier la colonne nouvellement ajoutée (donc de A à I dans le tableau 18c) et on va utiliser la colonne pivot A pour réordonner le tableau, successivement Trier et filtrer, Tri personnalisé…, Trier par colonne A, de A à Z (tableau 18d). Le tableau synthétique reprend sa structure initiale. En comparant la colonne I à l’ensemble des colonnes C à H, on voit que l’on peut rattacher I au Molinion caeruleae (absence des taxons du Juncion acutiflori) et plutôt à la colonne D décrivant l’Eu–Molinienion caeruleae.
Évidemment, pour appliquer cette méthode, il faut disposer des tableaux synthétiques au format Excel. Un tableau de classe réduit suffit (voir par exemple les synthèses des Crypsietea aculeatae in de Foucault, 2021a ou des Thero-Suaedetea splendentis in de Foucault & Guitton, 2021). Dans le cas de classes imposantes (nombreux taxons et syntaxons), on préconise de réaliser les tableaux d’alliance puis de condenser chacun d’eux en une colonne synthétique pour élaborer le tableau de classe. Comment passer des premiers au final ? En p. 15 de ma thèse de 1984, j’ai brièvement exposé la méthode ; celle-ci n’ayant fait l’objet d’aucune présentation officielle, je l’expose ici avec plus de détails. Considérons le tableau 10 (Deschampsio mediae-Molinienion caeruleae) que l’on cherche à condenser en une colonne unique. Pour les syntaxons représentés par au moins cinq relevés, on procède comme s’il y avait réellement cinq relevés dans chacun d’eux (tableau 19 ci-dessous, lignes en bistre), donc V devient 5, IV devient 4…, on ne touche pas aux + et r ou on les ignore. On peut alors additionner toutes les présences supérieures ou égales à 1 et les transformer en chiffres romains sur la base du nombre total fictif de relevés (ici 73 ; tableau 19 : colonne en vert) ; pour les + et r, on estime leur fréquence pour choisir au final entre + et r (ici seulement des r). C’est cette colonne synthétique finale qui est reprise dans la colonne D-Mc du tableau 17. Dans toutes ces manipulations, les colonnes en couleur ne sont que des artifices intermédiaires, elles ne doivent pas être conservées dans les tableaux définitifs publiés.
Ces aspects pratiques autour du tableau phytosociologique viennent compléter l’approche théorique et pratique exposée in de Foucault (2020a). Il est évident que toute cette heuristique n’est possible que si les tableaux Excel sont mis à la disposition de tous les phytosociologues qui souhaitent les utiliser, au contraire des tableaux publiés au format texte voire, pire encore, coupés sur plusieurs pages parce que trop volumineux. À ce titre, la revue Carnets botaniques de la Société botanique d’Occitanie publiée en ligne avec les tableaux comme matériel supplémentaire est tout à fait adaptée à cette problématique. En cas de publication d’associations ou sous-associations nouvelles, il ne faut toutefois pas oublier de publier les relevés types dans le texte lui-même (et pas simplement dans le matériel supplémentaire) sous peine d’invalidité selon les art. 1 (§ 2 et 3) et 2 (Note 4, § 5) de la v. 4 de l’ICPN.
Bibliographie
Barbe J., 1972. Clé de détermination des groupements végétaux des environs de la station de Bonnevaux (Jura central). Bulletin de la Fédération des sociétés d’histoire naturelle de Franche-Comté 74 : 104-111.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Patrimoines naturels 61 : 1-171.
Birse E.L., 1980. Plant communities of Scotland, a preliminary phytocoenonia. Soil Survey of Scotland 4 : 1-235.
Bolòs O. (de), 1979. Sur quelques groupements herbacés hygrophiles du Montseny (Catalogne). Phytocoenologia 6 : 202-208.
Bournérias M., 1949. Les associations végétales de l’antique forêt de Beine. Encyclopédie biogéographique et écologique III : 1-163.
Bournérias M., 1968. Guide des groupements végétaux du Bassin parisien. SEDES, Paris, 290 p.
Bournérias M., 1984. Guide des groupements végétaux du Bassin parisien, 3e édition. SEDES et Masson, Paris, 483 p.
Bournérias M. & Maucorps J., 1975. Les landes oligotrophes des “usages” de Versigny (dép. de l’Aisne). Documents phytosociologiques 9-14 : 19-38.
Bournérias M., Arnal G. & Bock Ch., 2001. Guide des groupements végétaux du Bassin parisien. Belin, Paris, 640 p.
Braun-Blanquet J., 1947. II – Les groupements végétaux supérieurs de la France. In J. Braun-Blanquet, L. Emberger & R. Molinier, Instructions pour l’établissement de la carte des groupements végétaux de la France, Montpellier : 19-32.
Braun Blanquet J., 1956. Clé écologique pour la détermination des classes, ordres et alliances phytosociologiques du Midi méditerranéen. Communications de la SIGMA 132 (II) : 9-16.
Braun Blanquet J., 1967. Vegetationsskizzen aus dem Baskenland mit Ausblicken auf des weitere Ibero-Atlantikum. Vegetatio XIII : 117-147, XIV : 1-126.
Braun Blanquet J. & Tüxen R., 1952. Irische Pflanzengesellschaften. Veröffentlichungen des Geobotanischen Institutes der ETH, Stiftung Rübel, in Zürich 25 : 223-421.
Catteau E., 2021. Caractérisation des végétations et séries marnicoles du Boulonnais (Pas-de-Calais, France). Botanique 7 : 37-71.
Clément B., 1978. Contribution à l’étude phytoécologique des monts d’Arrée ; organisation et cartographie des biocénoses ; évolution et productivité des landes. Thèse de doctorat de 3e cycle, université de Rennes, 260 p.
Clément B., 1981. Compte-rendu de la session de l’Amicale internationale de phytosociologie en Bretagne du 22 au 29 juillet 1979. Documents phytosociologiques, n. s., V : 467-501.
Clément B., 1987. Structure et dynamique des communautés et des populations végétales des landes bretonnes. Thèse de doctorat d‘État, université de Rennes, 320 p.
Corillion R. & Planchais N., 1963. Recherches sur la végétation actuelle et passée d’une lande tourbeuse armoricaine : Malingue (Mayenne). Pollen et Spores V (2) : 373-386.
Corriol G., 2008. Essai de clé typologique des groupements végétaux de Midi-Pyrénées et des Pyrénées françaises ; I – Introduction et pelouses acidophiles (Nardetea et Caricetea curvulae). Le Monde des plantes 495 : 3-13.
Corriol G., Sanz Th. & Van Es J., 2023. Contribution au Prodrome des végétations de France : les Elyno myosuroidis – Seslerietea caeruleae Braun-Blanq. 1948. Carnets botaniques 120 : 1-87.
Costa J.C., Capelo J., Jardim R., Sequeira M., Lousã M., Espírito-Santo M.D. & Rivas-Martínez S., 2004. Notas do Herbário da Estação Florestal Nacional (LISFA), XVIII, 3 – A Vegetação da Madeira VII, a classe Molinio-Arrhenatheretea Tüxen 1937 e IsoetoNanojuncetea Br.-Bl. & Tüxen 1937 ex Westhoff, Dijk & Passchier. Silva Lusitana 11 (2) : 251-256.
Dakskobler I., Seliškar A. & Vreš B., 2021. Phytosociological analysis of Gladiolus palustris sites in northwestern, western and southwestern Slovenia. Folia Biologica et Geologica 62 (1) : 59-159.
Delassus L., Magnanon S. et 24 coauteurs, 2014. Classification physionomique et phytosociologique des végétations de Basse-Normandie, Bretagne et Pays de la Loire. Les Cahiers scientifiques et techniques (CBN de Brest) 1 : 1-262.
Delelis-Dusollier A. & Géhu J.-M., 1975. Apport à la phytosociologie de quelques forêts thermo-acidiphiles ligériennes et de leurs stades d’altération. Colloques phytosociologiques III, La végétation des forêts caducifoliées acidiphiles : 141-156.
Delpech R., 1980. Les prairies tourbeuses du haut Vivarais (Ardèche, France). Colloques phytosociologiques VII, La végétation des sols tourbeux : 265-275.
Didier B. & Royer J.-M., 1989. Étude phytosociologique des prairies de fauche inondables des vallées de l’Aube, de la Seine et de la Marne (Champagne crayeuse). Colloques phytosociologiques XVI, Phytosociologie et pastoralisme : 195-208.
Donker M. & Stevelink A., 1962. Einige Wiesenvegetationen (Gaudinieto-Arrhenatheretum; Molinietum mediterraneum; Caricetum divisae) im Vistre-Tal bei le Cailar. Mededelingen van de Landbouwhogeschool te Wageningen 61 (15) : 1-32.
Duvigneaud J., 1955. Notes sur quelques groupements végétaux de la Fagne mariembourgeoise. Bulletin de la Société royale de botanique de Belgique 87 : 145-155.
Duvigneaud P., 1966. Notes sur la biogéochimie des serpentines du sud-ouest de la France. Bulletin de la Société royale de botanique de Belgique 99 (2) : 271-330.
Ferrez Y., Bailly G., Beaufils Th., Collaud R., Caillet M., Fernez Th., Gillet F., Guyonneau J., Hennequin Ch., Royer J.-M., Schmitt A., Vergon-Trivaudey M.-J., Vadam J.-C. & Vuillemenot M., 2011. Synopsis des groupements végétaux de Franche-Comté. Les Nouvelles Archives de la flore jurassienne et du nord-est de la France, n° sp. 1 : 1-281.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’Etat, université de Rouen, 675 p.
Foucault B. (de), 1986a. Données phytosociologiques sur la végétation observée lors de la douzième session de la SBCO en Limousin et Marche. Bulletin de la Société botanique du Centre-Ouest, n. s., 17 : 291-308.
Foucault B. (de), 1986b. Quelques données phytosociologiques peu connues sur la végétation du Boulonnais et de la Côte d’Opale (Pas-de-Calais, France). Documents phytosociologiques, n. s., X (2) : 93-116.
Foucault B. (de), 1986c. Contribution à une étude systémique des prairies de l’Aubrac (Massif central français). Documents phytosociologiques, n. s., X (1) : 255-305.
Foucault B. (de), 1988. Les végétations herbacées basses amphibies : systémique, structuralisme, synsystématique. Dissertationes Botanicae 121 : 1-150.
Foucault B. (de), 1993a. Observations floristiques et phytosociologiques sur la commune de Wallers-Trélon et ses environs. Natura Mosana 46 (2) : 37-52.
Foucault B. (de), 1993b. Nouvelles recherches sur les pelouses de l’Agrostion curtisii et leur syndynamique dans l’ouest et le centre de la France. Bulletin de la Société botanique du Centre-Ouest, n. s., 24 : 151-178.
Foucault B. (de), 1994. Compte rendu de l’excursion de la SBNF en forêt d’Andigny (Aisne). Contribution à la connaissance phytosociologique des forêts du Nord. Bulletin de la Société de botanique du nord de la France 47 (2) : 5-20.
Foucault B. (de), 1995a. Contribution à une monographie phytosociologique de la Hague (Manche, France). Bulletin de la Société de botanique du nord de la France 48 (4) : 45-90.
Foucault B. (de), 1995b. Contribution à la connaissance phytosociologique des forêts sur sable tertiaire du département du Nord. Bulletin de la Société de botanique du nord de la France 48 (1) : 13-26.
Foucault B. (de), 1997a. Nouvelles réflexions sur les lois qualitatives du monde végétal et leur valeur heuristique. Acta Botanica Gallica 144 (1) : 129-144.
Foucault B. (de), 1997b. Contribution à la connaissance phytosociologique des forêts et lisières mésophiles du plateau de Chambaran (Isère et Drôme, France). Bulletin mensuel de la Société linnéenne de Lyon 66 (9) : 233-250.
Foucault B. (de), 2006. La montagne châtillonnaise (journée du 14 juillet 2005). Bulletin de la Société botanique du Centre-Ouest, n. s., 37 : 483-490.
Foucault B. (de), 2008a. Validation nomenclaturale de syntaxons inédits ou invalides. Journal de botanique de la Société botanique de France 43 : 43-61.
Foucault B. (de), 2008b. Note phytosociologique sur les végétations observées en Haute-Normandie. Bulletin de la Société de botanique du nord de la France 61 (1-4) : 43-46.
Foucault B. (de), 2017. Données phytosociologiques nouvelles sur la végétation des Cévennes occidentales (département de la Lozère) et du Haut-Languedoc (département de l’Hérault). Bulletin de la Société botanique du Centre-Ouest, n. s., 48 : 334-359.
Foucault B. (de), 2018. Caractérisation des habitats de la Bézalado et de la Sagne Grande, commune des Martys (Aude, France). Evaxiana 4 : 143-171.
Foucault B. (de), 2019a. Contribution à la connaissance phytosociologique de la végétation du pays de Sault (Aude, France). Evaxiana 6 : 29-129.
Foucault B. (de), 2019b. Errata et compléments à propos du Prodrome des végétations de France, sixième note. Journal de botanique de la Société botanique de France 86 : 93-102
Foucault B. (de), 2020a. Approche théorique et pratique du tableau phytosociologique. Botanique 6 : 37-51.
Foucault B. (de), 2020b. Données phytosociologiques sur les forêts de la Malepère (Aude, France) et leurs lisières. Carnets botaniques 9 : 1-15 et 15 tableaux, https://doi.org/10.34971/5nt2-zy92.
Foucault B. (de), 2021a. Contribution à la connaissance de la végétation des étangs asséchés et des canaux du Narbonnais. Carnets botaniques 38 : 1-15 + 3 tableaux ; https://doi.org/10.34971/387K-6576.
Foucault B. (de), 2021b. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49
Foucault B. (de), 2022. Miscellanées phytosociologiques III : syntaxons nouveaux, validés ou complétés. Carnets botaniques 112 : 1-22, https://doi.org/10.34971/6AX1-0K40.
Foucault B. (de), 2023. Miscellanées phytosociologiques IV : syntaxons nouveaux, validés ou complétés. Carnets botaniques 127 : 1-18 + 14 tableaux, https://doi.org/10.34971/1d8e-ky97.
Foucault B. (de) & Géhu J.-M., 1980. Essai synsystématique et chorologique sur les prairies à Molinia coerulea et Juncus acutiflorus de l’Europe occidentale. Colloques phytosociologiques VII, La végétation des sols tourbeux : 135-164.
Foucault B. (de) & Philippe Th., 1989. Systémique des prairies du Morvan (Massif central, France). Colloques phytosociologiques XVI, Phytosociologie et pastoralisme : 101-141
Foucault B. (de) & Philippe Th., 1990. Données phytosociologiques sur la végétation observée lors de la seizième session de la SBCO en Haute-Savoie. Bulletin de la Société botanique du Centre-Ouest, n. s., 21 : 391-412.
Foucault B. (de), Frileux P.-N. & Delpech R., 1992a. Contribution à l’étude phytosociologique des systèmes prairiaux de la Brenne (Indre, France). Documents phytosociologiques, n. s., XIV : 273-303.
Foucault B. (de), Bournérias M. & Wattez J.-R., 1992b. Données floristiques et phytosociologiques récentes sur les milieux tourbeux basiclines du marais de Sacy-le-Grand (Oise). Bulletin de la Société botanique de France 139, Lettres botaniques (1) : 75-91.
Foucault B. (de) & Matysiak J.-P., 1995. Étude phytosociologique du complexe humide de Raimbeaucourt (département du Nord, France), ZNIEFF 11, type I. Application de la théorie des stratégies adaptatives. Natura Mosana 48 (3) : 57-76.
Foucault B. (de), Wattez J.-R. & Santune V., 1999. La végétation de l’ex-pré communal de Saint-Josse (Pas-de-Calais) ; son évolution sur une période d’une trentaine d’années. Bulletin de la Société de botanique du nord de la France 52 (2-3) : 23-37.
Foucault B. (de), Grzemski B., Toussaint B., Leduc A. & Grzemski M.-N., 2000. Compte-rendu de la sortie aux environs d’Ohain et Anor (mai 2000). Bulletin de la Société de botanique du nord de la France 53 (2-3) : 17-20.
Foucault B. (de) & Julve Ph., 2002. Observations phytosociologiques sur la végétation de la région de Reims (Marne). Bulletin de la Société de botanique du nord de la France 55 (3-4) : 73-87.
Foucault B. (de) & Royer J.-M., 2019. Quelques données phytosociologiques sur le bas Languedoc à propos de la session de la SBF en avril 2018. Journal de botanique de la Société botanique de France 87 : 83-96.
Foucault B. (de) & Guitton H., 2021. Une déclinaison moderne des Thero-Suaedetea splendentis. Carnets botaniques 76 : 1-9 + 2 tableaux, https://doi.org/10.34971/3XK3-N640.
Foucault B. (de), Beaufils L., Gilbert B. & Lépagnole C., 2021. Quelques données sur la végétation des Pyrénées audoises à travers celle du plateau de Counozouls (Aude). Carnets botaniques 74 : 1-12 + 9 tableaux, https://doi.org/10.34971/x1yh-sv34.
Foucault B. (de) & Klesczewski M., 2021. Contribution à une monographie phytosociologique du domaine de Peyremale (commune de Montolieu, Aude). Carnets botaniques 47 : 1-18 + 16 tableaux, https://doi.org/10.34971/B79J-2H81.
Gaži-Baskova V. & Trinajstić I., 1970. Floristische Gliederung, syndinamisch-syngenetische Verhältnisse und geographische Verbreitung des Deschampsietum mediae illyricum (Zeidler) H-ić. Acta Botanica Croatica 29 : 149-156.
Gelez W., Delplanque S. & Catteau E., 2015. Contribution à la connaissance des végétations du Parc naturel régional Scarpe- Escaut. Bulletin de la Société de botaniqe du nord de la France 68 (1-2) : 37-52.
Ghestem A. & Vilks A., 1978. Premières données phytosociologiques sur les formations prairiales hygrophiles du Limousin et de la Marche (nord-ouest du Massif central, France). Colloques phytosociologiques V, La végétation des prairies inondables : 153-163.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Gruber M., 1978. La végétation des Pyrénées ariégeoises et catalanes occidentales. Thèse de doctorat d’État, université de Marseille, 305 p. et tableaux.
Guinochet M., 1970. Clé des classes, ordres et alliances phytosociologiques de la France. Naturalia Monspeliensia, botanique, 21 : 79-119.
Guinochet M. & de Vilmorin R., 1973. Flore de France, I. CNRS, Paris, 366 p.
Ilijanić, L. 1968. Die Ordnung Molinietalia in der Vegetation Nordostkroatiens. Acta Botanica Croatica 26-27 : 161-180.
Julve Ph., 1983. Les groupements de prairies humides et de bas-marais ; étude régionale et essai de synthèse à l’échelle de l’Europe occidentale. Thèse de doctorat de 3e cycle, université de Paris-Sud Orsay, 224 p.
Julve Ph., 1993. Synopsis phytosociologique de la France (communautés de plantes vasculaires). Lejeunia, n. s., 140 : 1-160.
Koch W., 1926. Die Vegetationseinheiten der linthebene unter Berücksichtigung der Verhältnisse in der Nordostschweiz. Jahrbuch der St. Gallischen Naturwissenschaftlichen Gesellschaft 61 : 1-144.
Korneck D., 1962. Die Pfeifengraswiesen und ihre wichstigten Kontaktgesellschaften in der nördlichen Oberrheinebene und im Schweinfurter Trockengebiet ; II – Die Molinieten feuchter Standorte. Beiträge zur naturkundlichen Forschung in Südwestdeutschland XXI (2) : 165-190.
Kuyper T.W., Leeuwenberg H.F.M. & Hübl E., 1978. Vegetationskundliche Studie an Feucht-, Moor- und Streuwiesen im Burgenland und östlichen Niederösterreich. Linzer biologische Beiträge 10 (2) : 231-321.
Laclos E. (de) & Royer J.-M., 2001 (publié 2002). Étude phytoécologique des dépressions des dalles calcaires à Deschampsia media et divers Juncus de Bourgogne et du sud du Jura. Bulletin de la Société botanique du Centre-Ouest, n. s., 32 : 53-72.
Lemée G., 1933. Études phytogéographiques sur les plaines jurassiques normandes ; II – Les buttes calloviennes des environs d’Alençon. Bulletin de la Société botanique de France 80 : 814-823.
Lemée G., 1937. Recherches écologiques sur la végétation du Perche. Thèse de doctorat d’État, université de Paris, 388 p.
Mayot J., 1977. Essai d’interprétation de la végétation de la partie inférieure du Jura central (feuille au 1/50 000e d’Orgelet). Thèse de doctorat d’État, université de Besançon, 248 p.
Meltzer J. & Westhoff V., 1942. Inleiding tot de plantensociologie. G.W. Breughel, S’Graveland, 326 p.
Molinier R., 1934. Études phytosociologiques et écologiques en Provence occidentale. Annales du Muséum d’histoire naturelle de Marseille 27 : 1-274.
Oberdorfer E., 1979. Pflanzensoziologische Exkursionsflora. E. Ulmer, Stuttgart, 997 p.
Oberdorfer E., 1983. Süddeutsche Pflanzengesellschaften, III. G. Fischer, Iena, 455 p.
Philippi G., 1960. Zur Gliederung der Pfeifengraswiesen im südlichen und mittelren Oberrheingebiet. Beiträge zur naturkundlichen Forschung in Südwestdeutschland 19 : 138-187.
Rameau J.-C. & Royer J.-M., 1972. Clé de détermination des groupements végétaux présents sur les terrains calcaires jurassiques du SE du Bassin parisien et de la Bourgogne. Revue de la Fédération française des sociétés de sciences naturelles 11 (48) : 31-64.
Rameau J-C. & Royer J.-M., 1978. Les moliniaies du plateau de Langres. Colloques phytosociologiques V, La végétation des prairies inondables : 269-286.
Resmeriţà I., 1977.La classe des Molinio-Arrhenatheretea Tx. 1937 dans les Carpathes roumaines. Documents phytosociologiques, n. s., I : 241-267.
Rivas-Martínez, S., 1979. Brezales y jarales de Europa occidental (Revision fitosociologica de las clases Calluno-Ulicetea y Cisto-Lavanduletea). Lazaroa 1 : 5-127.
Robbe G., 1993. Les groupements végétaux du Morvan. Société d’histoire naturelle d’Autun, 160 p.
Royer J.-M. & Didier B., 1996. Flore et végétation des marais tufeux du plateau de Langres. Mémoires de la Société de sciences naturelles et d’archéologie de la Haute-Marne 2 : 1-112.
Royer J.-M., Felzines J.-C., Misset C. & Thévenin S., 2006. Synopsis commenté des groupements végétaux de la Bourgogne et de la Champagne-Ardenne. Bulletin de la Société botanique du Centre-Ouest, n. s., n° sp. 25 : 1-394.
Royer J.-M. & Ferrez Y., 2018. Contribution au prodrome des végétations de France : les Sedo albi – Scleranthetea biennis Braun-Blanq. 1955. Documents phytosociologiques, 3e série, 7 : 179-281.
Teles A.N., 1970. Os lameiros de montanha do norte de Portugal. Subdivisios para a sua caracterizaçao fitosociologia e quimica. Agronomia Lusitana 31 (1-2) : 5-132.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.
Trivaudey M.-J., 1997. Contribution à l’étude phytosociologique des prairies alluviales de l’est de la France (vallées de la Saône, de la Seille, de l’Ognon, de la Lanterne et du Breuchin), approche systémique. Dissertationes botanicae 284 : 1-216 et tableaux.
Tüxen R., 1937. Die Pflanzengesellschaften Nordwestdeutschlands. Mitteilungen der Floristisch-soziologische Arbeitsgemeinschaft Niedersachsen 3 : 1-170.
Wattez J.-R., 1978. Les jonçaies acidoclines à Juncus acutiflorus Ehrh. du nord de la France. Colloques phytosociologiques V, La végétation des prairies inondables : 319-338.
Westhoff V. & Den Held A.J., 1969. Plantengemeenschappen in Nederland. Thieme, Zutphen, 324 p.
Remerciements
Ils s’adressent à Vincent Gaudillat (MNHN, Service du patrimoine naturel) et Jean-Marie Royer pour leur aide à la recherche de documents bibliographiques.


