
Premier bilan de la bryoflore des gorges du Rimouren (Ardèche méridionale)
Title
First assessment of the bryophyte flora of the gorges du Rimouren (Southern Ardèche)
Résumé
En 2022, dans le cadre de l’actualisation des données ZNIEFF de la région biogéographique méditerranéenne, le Conservatoire botanique national du Massif central a inventorié la bryoflore de cinq ZNIEFF ardéchoises de type 1, parmi lesquelles les gorges de Rimouren, parcourues le 14 avril 2022. Parmi les espèces recensées, plusieurs taxons remarquables pour leur statut de menace selon les critères IUCN et leur rareté en région Auvergne-Rhône-Alpes (AuRA) ou en France métropolitaine méritent à notre avis d’être valorisés. Certains parmi ces taxons appartiennent à une communauté bryologique originale et peu connue, qui est présenté dans les détails.
Abstract
In 2022, as part of the updating of ZNIEFF data for the Mediterranean biogeographical region, the Conservatoire botanique national du Massif central inventoried the bryoflora of five type 1 ZNIEFFs in the Ardèche, including the Rimouren gorge, identified with the code 820030234. Among the species inventoried, several taxa remarkable for their threatened status according to IUCN criteria and their rarity at regional (Auvergne-Rhône-Alpes, AuRA) or national level, deserve in our opinion to be valued. Some of these taxa constitute an original and little known bryological community, which is presented in detail.
1. Le Rimouren : un cours d’eau intermittent en marge du domaine méditerranéen
Site ardéchois peu connu et très sauvage, les gorges du Rimouren (planche 1) s’étendent sur près de six kilomètres depuis le hameau éponyme (sur la commune de Gras) jusqu’à la confluence du ruisseau avec celui d’Imbours (Saint-Montan), les deux ruisseaux formant alors celui de la Conche. Suivant une orientation sud-ouest/nord-est, elles se trouvent à la limite septentrionale de la région méditerranéenne. Le bioclimat méditerranéen humide de variante fraîche selon la classification d’Emberger (Daget, 1977 ; Daget & David, 1982 ; Quézel & Médail, 2003) se caractérise par un cumul moyen annuel des précipitations de 900-1 000 mm, une température moyenne annuelle de 13.6-14.6 °C, une maxima du mois le plus chaud (juillet) autour de 31 °C et une mimima du mois le plus froid (janvier) entre 0,5 et 2,4 °C[1]. Le paysage géologique est constitué de roches calcaires du Crétacé telles que les calcaires à rudistes, calcarénites et biocalcarénites, avec des insertions de vires marneuses horizontales vers la partie terminale des gorges.

Le Rimouren est un cours d’eau intermittent soumis à un régime hydrologique complexe déterminé par deux facteurs principaux :
- la structure karstique des gorges, à l’origine d’un important réseau de tunnels, grottes, cavités souterraines, mais aussi de nombreuses pertes, résurgences et exsurgences le long de son parcours ;
- les précipitations irrégulières, un maximum pluviométrique automnal et un minimum estival (saison la plus chaude), constituant ainsi une période de sécheresse (sensu Bagnouls & Gaussen, 1957) et entraînant un débit faible, voire nul. Bien que l’automne correspondrait théoriquement à la saison de crue, la remise en eau du ruisseau, au moins localement, est en réalité aléatoire à cause des phénomènes karstiques.
Il en résulte un écoulement temporaire fort dépendant des facteurs géoclimatiques, se présentant sous la forme d’une alternance de tronçons en eau courante (phases euréique et oligoréique), en assec (phases hyporéique et édaphique) et de profondes vasques en eau isolées (phase aréique) (planche 2). Déconnectées du réseau d’eau superficiel, ces dernières sont alimentées par les écoulements et résurgences souterrains. Ce type de système hydrologique et son fonctionnement sont décrits de façon très précise par Gallart et al. (2012) et Bonada et al. (2020)[2]. Juste en aval du hameau de Rimouren, à l’approche des gorges proprement dites, le milieu est ouvert. Les berges, terrassées ou en pente adoucie, sont formées de barres rocheuses basses et érodées, de grandes dalles et de murettes en pierre qui, en fonction du niveau topographique, alternent avec des fourrés rivulaires à saules (Salix spp.), des ourlets à Brachypodium retusum et des matorrals à genévrier (Juniperus spp.). L’eau court sans discontinuer, avec un débit faible à modéré et un lit généralement peu profond, ce qui permet la connexion de plusieurs grandes vasques circulaires (celles-ci très profondes) présentes en occasion des sauts de roche, où des cascades se forment et sont accompagnées de tufs détrempés et actifs. Ceux-ci se forment également sur les dalles horizontales et les petits gradins rocheux humides.

Bordé par des falaises, des affleurements rocheux et éboulis ou des versants boisés en pente raide, le cœur des gorges du Rimouren offre deux types de géomorphologie :
- l’un très encaissé, d’ambiance canyon, à lit étroit, où l’eau a façonné la roche en lui conférant des formes douces et sinueuses et une surface souvent polie. Les tronçons à lit large de quelques mètres alternent avec des passages larges de moins d’un mètre prenant l’aspect de défilé, des sauts de roche qui donnent lieu à des cascades durant la période en eau, des vasques circulaires ou allongées. Sur les roches, les dépôts de tuf d’épaisseur variable sont récurrents et souvent humides (actifs), y compris dans les parties du lit en assec, ce qui confirme la présence de suintements tufeux (quasi) permanents ;
- l’autre à lit plus large, plus faiblement creusé et constitué de roche affleurante, galets et gros blocs éboulés, à topographie plutôt plane, souvent en correspondance avec des pentes de versant plus faibles avec absence de parois directement en bord du ruisseau. Ce contexte permet l’expression d’une ripisylve issue des chênaies et matorrals méditerranéens situés en mi- et haut-versant. En effet, Prunus maheleb, Phillyrea latifolia, Rhamnus alaternus et Piptatherum paradoxum ont déjà commencé à coloniser le lit, illustrant les longues périodes d’assec que connaît localement le Rimouren. Les tufs en bord du lit sont moins fréquents, de grande taille mais tous secs (inactifs), témoignant de la courte temporalité des suintements.
2. Une bryoflore remarquable d’affinité méditerranéo-océanique
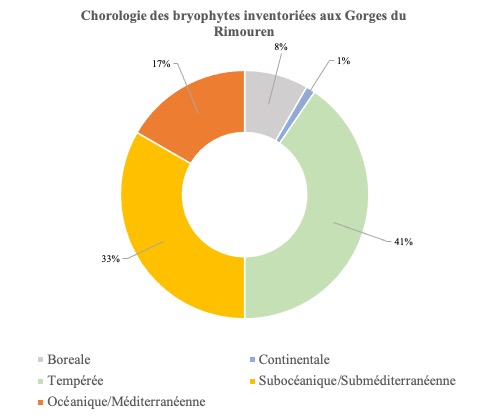
La requête de l’outil LOBELIA[3] en date du 7 janvier 2022 (précédant de quelques mois notre passage sur le site) indique que l’état des connaissances de la bryoflore des gorges du Rimouren est nul. Nous avons parcouru les gorges en suivant le talweg depuis le hameau de Rimouren jusqu’aux alentours de la grotte de Chironlong. Deux détours sur le versant boisé d’ubac nous ont permis de prospecter succinctement les matorrals à chêne vert, les clairières à pelouses rases et écorchées du bois communal de Saint-Montant, les barres rocheuses de mi-versant et un éboulis calcaire intraforestier, l’entrée de la grotte de Pascaloune. Dans l’objectif de maximiser les probabilités d’observer des espèces déterminantes ZNIEFF, nous avons choisi de concentrer les efforts de recherche dans les microhabitats favorables le long du talweg, à savoir suintements et sources pétrifiantes, affleurements rivulaires et fissures rocheuses, balmes et microbalmes, entrées de grottes et cavités des barres rocheuses. Les prospections ont permis d’y inventorier 86 taxons (autant dire que l’inventaire est loin d’être exhaustif ; la nomenclature suit TaxRef v. 15 ; Gargominy et al., 2021), parmi lesquels dix sont des hépatiques. Une rapide analyse de la représentativité des types chorologiques (sensu Düll, 1983, 1984, 1985, 1992) révèle une bryoflore principalement partagée entre taxons tempérés (40,5 %) et taxons subocéaniques et/ou subméditerranéens (33,3 %). Aussi, tandis que la part des éléments océaniques-méditerranéens atteint près de 17 % (Bryum gemmiparum, Fissidens crispus, Habrodon perpusillus, Ptychostomum donianum, Scorpiurium deflexifolium et Timmiella anomala), les éléments d’affinités continentale et boréale au sens large représentent respectivement 1,2 et 8,3 % de la bryoflore inventoriée (figure 1).
Il nous semble également intéressant de remarquer la présence d’une espèce habituellement silicicole, Imbribryum alpinum, accompagnées d’autres Bryacées telles que Bryum gemmiparum (photo 1) et Ptychostomum pseudotriquetrum sur les dalles calcaires humides tufeuses en bord de ruisseau et les tufs des suintements. Ces espèces se présentent alors souvent sous des formes réduites, voire méconnaissables, comme l’a remarqué déjà Hugonnot (2010) dans le bois de Païolive.

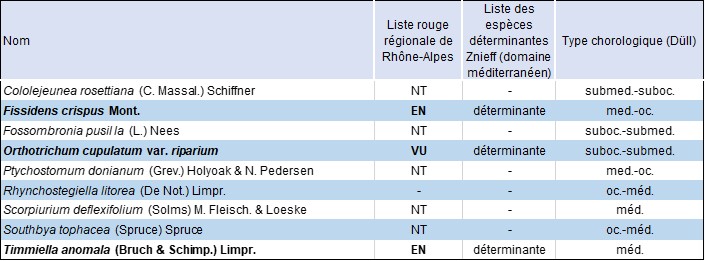
Parmi les 86 espèces recensées, bien qu’aucune ne figure sur la Liste rouge d’Europe (Hoddgets et al., 2019), huit apparaissent sur la Liste rouge de l’ex-région Rhône-Alpes (CBNA, 2022) :
- trois aux rangs en danger (EN) et vulnérable (VU), du fait d’un faible nombre de populations : Fissidens crispus, Orthotrichum cupulatum riparium et Timmiella anomala (photo 2) ;
- cinq au rang quasi menacé (NT), également en raison du faible nombre de populations répertoriées à l’échelon régional : Cololejeunea rossettiana, Fossombronia pusilla, Ptychostomum donianum (figure 2, planche 3), Scorpiurium deflexifolium et Southbya tophacea.


Quoique ne figurant pas dans la Liste rouge régionale, soulignons également la présence de Rhynchostegiella litorea. D’affinité océanique-méditerranéenne, cette mousse pleurocarpe à port tapissant se distingue de Rh. tenella (dont elle était auparavant considérée comme une variété) par ses feuilles et sa costa plus courtes. Rare à l’échélle nationale, la présence de cette espèce dans la métropole se limite aujourd’hui à la région méditerranéenne et à la façade atlantique (Hugonnot, 2007). En AuRA, l’espèce est rarissime, seulement signalée d’une localité en Sud-Ardèche. Dans les gorges du Rimouren, elle a été observée en haut de versant, dans une tonsure de matorral arborescent à chêne vert. Ce cortège de taxons patrimoniaux, neuf au total, est formé d’éléments (sub)océaniques et/ou (sub)méditerranéens (tableau 1).
3. Le groupement à Timmiella anomala et Gymnostomum calcareum : une communauté originale
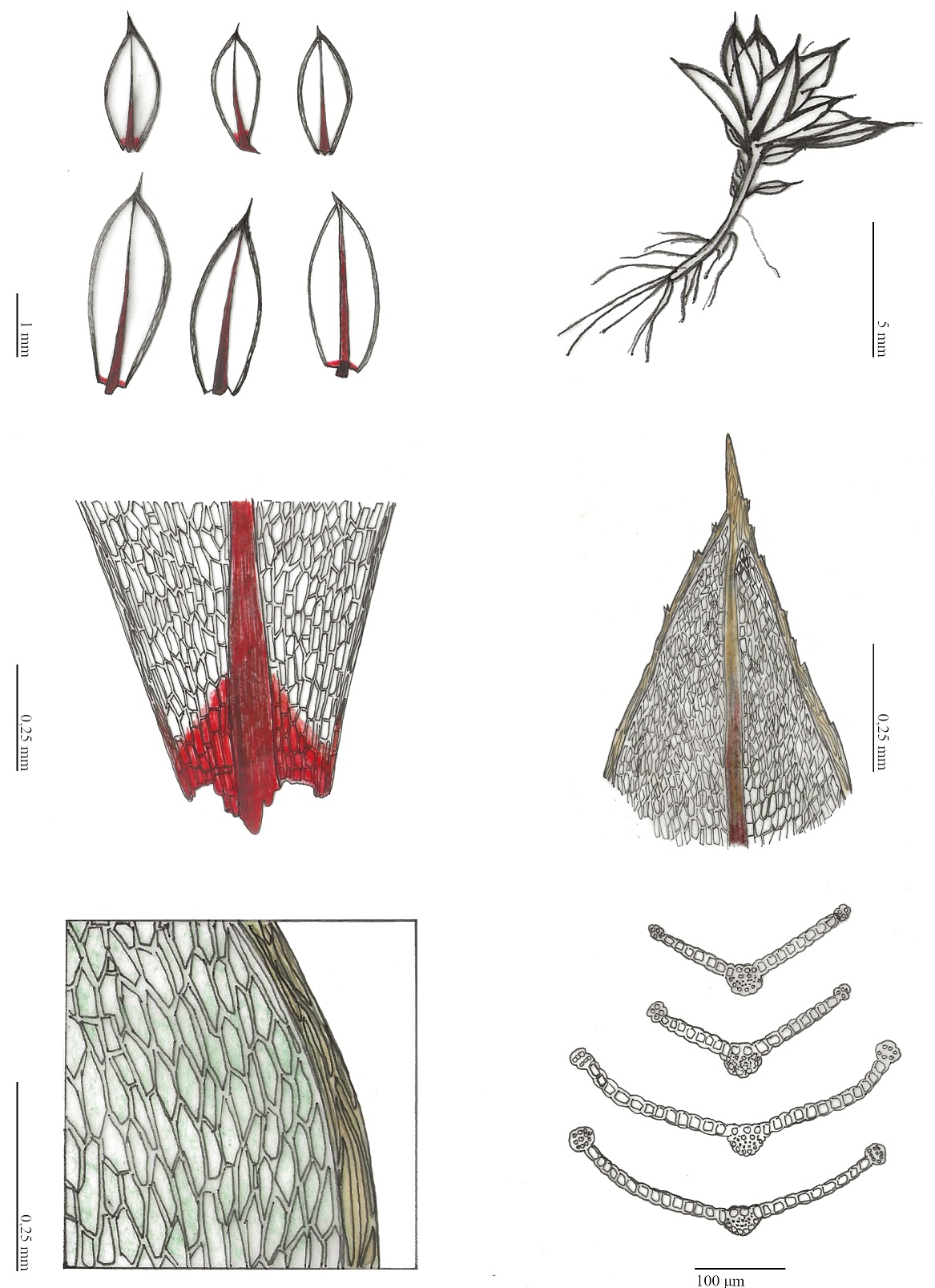
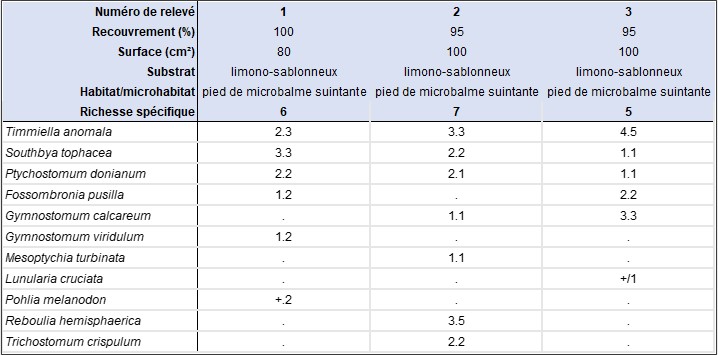
L’observation d’un microgazon dense, ras et pionnier composé de Timmiella anomala, Ptychostomum donianum et Southbya tophacea, sur un sol sablo-limoneux en pied de paroi calcaire suintante, en situation de microbalme, en rive droite du Rimouren, nous a amenés à produire trois relevés bryosociologiques[4] (tableau 2), afin de contribuer à la documentation de cette communauté bryophytique, constituée majoritairement d’acrocarpes, qui nous est apparue comme originale (planche 4, photo 3). Trois espèces constantes structurent cette communauté limicole pionnière : Timmiella anomala, Ptychostomum donianum et Southbya tophacea ; deux autres espèces, Fossombronia pusilla et Gymnostomum calcareum, sont également présents dans deux des trois relevés réalisés.


Timmiella anomala
Les Timmiella se reconnaissent in situ et à l’état hydraté à leur couleur vert clair presque luminescent (Hugonnot, 2010), leurs feuilles à marge pâle et apex denticulé. T. anomala se différencie de T. barbuloides (également présente dans le Bas-Vivarais) par leur sexualité ainsi que par l’anneau de la capsule. Ce taxon d’affinité méditerranéenne est très rare en France, d’autant plus rare qu’on s’éloigne de la frange la plus chaude du secteur méditerranéen (Hugonnot, 2010). En région Auvergne-Rhône-Alpes, les populations se situent toutes dans le Bas-Vivarais (gorges de l’Ardèche et de la Ligne).
Ptychostomum donianum
D’identification relativement aisée, cette Bryacée méditerranéo-océanique vivant en colonie lâche et plane se reconnaît à son limbidium épais (deux-quatre strates de cellules) et à son arête terminale très courte ornée de dents géminées, parfois peu marquées. Rare en France métropolitaine et peu fréquente à l’intérieur des terres, cette espèce thermophile trouve son optimum sur les pourtours méditerranéen et atlantique (Hugonnot, 2010). En AuRA, elle est principalement connue dans le Bas-Vivarais et dans la Drôme.
Southbya tophacea
D’affinité océanique-méditerranéenne, cette hépatique au port prostré se reconnaît à ses feuilles succubes et opposées, vert clair et opaque. Basiphile, elle s’observe majoritairement sur suintements tufeux. Relativement rare en France métropolitaine (Hugonnot & Chavoutier, 2021), elle occupe la façade atlantique et la région méditerranéenne. En AuRA, elle est repartie principalement dans les secteurs méridionaux de l’Ardèche et de la Drôme.
Fossombronia pusilla
D’affinité subméditerranéenne-subatlantique, Fossombronia pusilla est disséminée dans une grande partie de la France. Cette terricole pionnière investit les terres nues dans des contextes variés (berges des ruisseaux, ornières de chemins forestiers, cultures, etc.).
La présence abondante des trois premiers taxons, associée à celles d’autres espèces occupant un espace plus réduit, comme Mesoptychia turbinata et Lunularia cruciata, amènent à positionner cette communauté au sein du Cephaloziello baumgartneri-Southbyon nigrellae Guerra & Gil 1982, alliance méditerranéo-atlantique plutôt photophile, épilithique, principalement de rochers et protosols calcaires (Guerra & Gil, 1982 ; Bardat & Hauguel, 2002). Cette communauté, qui ne saurait être assimilée à l’heure actuelle en l’état à aucune association décrite en Europe, est à rattacher au groupement à Timmiella anomala et Gymnostomum calcareum (Celle & Hugonnot, 2012).
Ce groupement semble en France n’avoir été signalé que des gorges de l’Ardèche, du Chassezac et de la Baume. Dans les premières, il apparaît de manière ponctuelle sur des abrupts rocheux recouverts de placages minces de sables limoneux soumis à des suintements temporaires carbonatés : ici, la communauté est formée également par l’association de T. anomala, L. cruciata, G. calcareum et P. donianum (Celle & Hugonnot, 2012). À quelques dizaines de kilomètres des gorges du Rimouren, dans le bois de Païolive, cette communauté est strictement inféodée aux placages d’alluvions déposés sous les tables calcaires à proximité immédiate du cours du Chassezac et plus rarement de la Baume (Hugonnot, 2010). De fait, la découverte du groupement dans les gorges du Rimouren élargit son spectre écologique, le Rimouren étant un cours d’eau de plus petit gabarit.
Comme l’expliquent précisément Celle et Hugonnot (2012), le décapage régulier des substrats provoqué par les crues permet dans un premier temps la recolonisation des rochers par des communautés saxicoles du Cinclidotion fontinaloidis G. Phil. 1956. Puis, les dépôts limono-sableux, qui vont progressivement se constituer dans les zones à l’abri des courants les plus forts, peuvent être investis par le groupement à Didymodon nicholsonii (Cinclidotion fontinaloidis). Enfin, tandis que le Leptodictyo riparii-Fissidentetum crassipedis G. Phil. 1956 s’installe sur les limons qui s’enrichissent en matières organiques, le groupement à Timmiella anomala et Gymnostomum calcareum se développe sur des placages limono-sableux soumis à des suintements temporaires. En l’absence d’une régénération cyclique induite par la dynamique alluviale, ces communautés pionnières laisseraient progressivement place à des ourlets.
Outre ce groupement, soulignons la présence de deux autres espèces remarquables dépendant de la dynamique alluviale du cours d’eau : Orthotrichum cupulatum var. riparium et Scorpiurium deflexifolium.
Orthotrichum cupulatum var. riparium
Orthotric subocéanique d’affinité montagnarde reconnaissable principalement à ses capsules émergentes à coiffe glabre, caractère qui la différencie de la variété type à coiffe poilue, et à son écologie saxicole puisqu’elle est spécifiquement inféodée aux rochers calcaires submersibles de basse altitude. Il peut être confondu avec deux autres taxons à écologie primaire saxicole et calcicole, mais plus répandus en France et assez fréquemment relevés dans les gorges du Rimouren : O. cupulatum Brid. var. cupulatum et O. anomalum Hedw. Peu fréquent en Ardèche méridionale (Hugonnot, 2010), il est rare et disséminé dans la région AuRA. Dans les gorges du Rimouren, il a été observé sur rochers et dalles calcaires chauds submersibles en bord de ruisseau au niveau d’un tronçon en assec.
Scorpiurium deflexifolium
Pleurocarpe méditerranéenne de taille moyenne formant des tapis vert olive, à feuilles de taille réduite (< 1 mm), ovales-elliptiques et dont les marges sont dentées dans la moitié supérieure. Il peut être confondu avec S. circinatum (Bruch) M. Fleisch. & Loeske hors de ce milieu (Hugonnot, 2010). Amphibie, étroitement inféodée aux supports (rochers et écorces) submersibles régulièrement exposés aux crues, l’espèce est disséminée dans le bassin méditerranéen, en Corse et dans le Sud-Ouest. En AuRA, les populations relativement nombreuses se situent majoritairement dans les sud de l’Ardèche. Dans les gorges du Rimouren, cette pleurocarpe a été observée fréquemment sur les rochers rivulaires du cours d’eau.
4. La grotte de Pascaloune et ses environs proches
Insérée dans le contexte karstique des gorges du Rimouren, la grotte de Pascaloune occupe une place discrète à mi-versant d’une pente boisée en situation d’ubac. Cependant, son rôle dans le système karstique est assez important : il s’agit d’une ancienne perte du Rimouren d’une profondeur d’environ 190 m et se développant sur 1,2 km, formant une rivière souterraine en relation hydrologique avec le bassin d’alimentation des sources de Tourne (Syndicat mixte Ardèche Claire, 2016). À l’entrée de cette grotte (photo 4) et en situation très ombragée sous le porche, le sol compact surmonté d’une fine accumulation limono-sableuse accueille un tapis ras et irrégulier de mousses de très petite taille. Nous y collectons un petit Fissidens à limbidium mince qui attire notre attention. L’identification de cet échantillon a posteriori révèlera son identité : il s’agit de F. crispus. À quelques pas de cette grotte, dans une fissure de rocher calcaire en situation ombragée, nous relevons la présence de Cololejeunea rossetiana.

Fissidens crispus
Petit Fissidens méditerranéo-océanique, reconnaissable à son limbidium étroit (un-deux rangs de cellules), son aréolation foliaire dense et opaque en raison de cellules de petite taille (4-10 μm), épaisses et à parois convexes, voire mammilleuses (aspect bombé, protubérant). Espèce rare en France, présente dans le Midi et en Corse (Sotiaux at al., 2007). Signalée il y a vingt ans sur le littoral de la Manche, dans la Somme, au bois de Cise (Boudier et al., 2000), elle y est aujourd’hui considérée comme exceptionnelle (Hauguel et al., 2013). En région AuRA, elle est connue seulement de deux localités situées dans les gorges de l’Ardèche, à quelques kilomètres des gorges du Rimouren, et de deux autres en basse vallée du Rhône (Drôme), en rive gauche du fleuve entre Valence et Pierrelatte.
Cololejeunea rossettiana
Petite hépatique subméditerranéenne-subocéanique et pionnière. La taille minuscule des feuilles, l’absence d’amphigastres et l’organisation en colonies associées à l’écologie mésoxérophile et saxicole constituent un faisceau d’indices permettant in situ une première identification au rang générique (Hugonnot & Chavoutier, 2021) qui doit obligatoirement être affinée en laboratoire. Peu fréquente en France métropolitaine, en AuRA, cette espèce est signalée de façon disséminée du Cantal à l’Ain, avec un foyer plus important en Ardèche méridionale.
5. Conservation
Outre son statut de ZNIEFF de type 1, cet espace naturel constitue un site Natura 2000 de premier ordre en étant à la fois une zone de protection spéciale (ZPS) et une zone spéciale de conservation (ZSC). Il s’inscrit pleinement dans les enjeux de conservation similaires à ceux d’autres sites de l’Ardèche méridionale.
Situées à environ dix kilomètres au nord/nord-est de la Réserve naturelle nationale des Gorges de l’Ardèche, les discrètes gorges du Rimouren, situées à l’écart des sites les plus prisés (comme les gorges de l’Ardèche qui connaît une affluence annuelle d’environ 1,5 million de spectateurs entre avril et septembre) et jouissant d’une relative tranquillité, sont ainsi épargnées des impacts de la fréquentation touristique estivale du Bas-Vivarais. Il est principalement connu pour ses grottes de Pascaloune et Chironlong qui sont explorées et visitées à des fins spéléologiques.
Néanmoins, les gorges du Rimouren ne sont pas à l’abri d’un changement des flux touristiques des sites les plus emblématiques et accessibles vers des sites périphériques méconnus, en particulier par l’essor des activités de plein nature telles que le canyoning. Aujourd’hui, le talweg n’est pas parcouru par des sentiers de randonnée balisés et conserve un caractère sauvage et authentique très marqué.
Une augmentation de la fréquentation du site nécessiterait des mesures adéquates afin de limiter et de canaliser les flux. L’activité de canyoning pourrait avoir, par le piétinement, des répercussions néfastes notamment sur le groupement bryophytique pionnier à Timmiella anomala et Gymnostomum calcareum situés au pied des parois rocheuses à quelques pas de la rivière, d’autant plus que le changement climatique renforcera cet impact par les répercussions qu’il aura probablement dans un futur proche sur l’activité des tufières et de ces mêmes communautés dont le maintien dépend de l’existence de suintements en pied de parois rocheuses. Enfin, le maintien d’un équilibre de perturbations hydrologiques cycliques et fréquentes, mais également d’une stabilité dans la qualité des eaux (en évitant l’eutrophisation et les pollutions) sont des conditions sine qua non à la conservation de cette communauté pionnière. Aussi, le petit Fissidens terricole découverte à l’entrée de la grotte de Pascaloune et menacé dans l’ex-région Rhône-Alpes pourrait souffrir du piétinement et d’éventuels aménagements liés à l’activité spéléologique. En effet, le piétinement et l’eutrophisation du sol dus à sa situation de point d’attrait des randonneurs et visiteurs, et ce malgré la faible fréquentation, constituent des menaces prégnantes qui pèsent sur la population de Fissidens crispus.
[1] Valeurs des indices S (indice sécheresse estivale) et Q2 (indice pluviométrique) calculés à partir des données historiques MétéoFrance 1981-2010 des stations de Bourg-Saint-Andéol et Vallon-Pont-d’Arc (Infoclimat.fr, consulté le 08/10/2022).
[2] Phases hydrologiques originairement décrites par Gallart et al. (2012) et Bonada et al. (2020) avec les termes anglais de « eurheic and oligorheic states », « hyporheic and edaphic states », « isoleted pools phase in arheic state », traduit en français pour l’occasion.
[3] Système d’information mutualisé utilisé par le Conservatoire botanique national du Massif central.
[4] Chaque relevé bryosociologique a fait l’objet prélèvement de substrat (conservée dans l’herbier privé d’A. Labroche). L’identification des espèces et leur attribution d’un coefficient d’abondance-dominance ont été effectuées au laboratoire.
Bibliographie
Anonyme, 2016. Étude d’identification et préservation des ressources souterraines stratégiques pour l’alimentation en eau potable du bassin versant de l’Ardèche, phase 1. AERMC, Syndicat mixte Ardèche Claire, SAGE Ardèche, Région Auvergne-Rhône-Alpes, bureau d’étude G. Rabin, ACTEON, Idées Eaux, 358 p.
Bagnouls F. & Gaussen H.,1957. Les climats biologiques et leur classification. Annales de géographie 66 (355) : 193-220.
Bardat J. & Hauguel J.-Ch., 2002. Synopsis bryosociologique pour la France. Cryptogamie, Bryologie 23 (4) : 279-343
Bonada N., Cañedo-Argüelles M., Gallart F. et al., 2020. Conservation and Management of Isolated Pools in Temporary Rivers. Water 12, 2870, doi : 10.3390/w12102870.
Boudier P., Hauguel J.-Ch. & Wattez J.-R., 2000. Contribution à la bryoflore du nord de la France. Compte rendu des cinquièmes rencontres bryologiques de la SBCO. Bulletin de la Societé botanique du Centre Ouest, NS, 31 : 507-542.
Celle J. & Hugonnot V., 2012. Les communautés bryophytiques du lit mineur des gorges de l’Ardèche comme témoins de sa richesse écosystémique. Ecologia Mediterranea 38 (2) : 29-41.
Daget Ph. 1977. Le bioclimat méditerranéen : caractères généraux, modes de caractérisation. Vegetatio 34 : 1-20.
Daget Ph. & David P., 1982. Essai de comparaison de diverses approches climatiques de la méditerranéité. Ecologia Mediterranea 8 (1-2) : 33-48.
Düll R., 1983. Distribution of the European and Macaronesian liverworts (Hepaticophytina). Bryologische Beiträge 2 : 1-115.
Düll R., 1984. Distribution of the European and Macaronesian mosses (Bryophytina). Bryologische Beiträge 4 : 1-114.
Düll R., 1985. Distribution of the European and Macaronesian mosses (Bryophytina), II. Bryologische Beiträge 5 : 110-232.
Düll R., 1992. Distribution of the European and Macaronesian Mosses (Bryophytina), annotations and progress. Bryologische Beiträge 8-9 : 1-223.
Gallart F., Prat N. & Garcìa-Roger E.-M., 2012. A novel approach to analysing the regimes of temporary streams in relation to their controls on the composition and structure of aquatic biota. Hydrology and Earth Systeme Sciences 16 : 3165-3182.
Gargominy O., Tercerie S., Régnier C. et al., 2021. Taxref v. 15.0, référentiel taxonomique pour la France. UMS PatriNat, Muséum national d’histoire naturelle, Paris, archive de téléchargement contenant 8 fichiers. https://inpn.mnhn.fr/telechargement/referentielEspece/taxref/15.0/menu
Guerra J. & Gil J.A., 1982. Comunidades briofíticas mediterráneas de protosuelos calcáreos húmidos. Folia Botanica Miscellanea 3 : 87-94.
Hauguel J.-Ch. (coord.), Wattez J.-R., Prey T., Messean A., Larere P. & Toussaint B., 2013. Inventaire des bryophytes de la Picardie : raretés, protections, menaces et statuts, version 3a. Centre régional de phytosociologie agréé Conservatoire botanique national Bailleul, 66 p.
Hoddgets N., Cálix M., Englefield E. et al., 2019. A miniature world in decline: European Red List of Mosses, Liverworts and Hornworts. IUCN, Brussels, Belgium, 88 p.
Holyoak D.T., 2021. European Bryaceae. British Bryological Society, Pisces Publications, Newbury, UK, 344 p.
Hugonnot V., 2007. Les bryophytes de l’ile de Porquerolles (Hyères, Var, France) : inventaire, atlas, groupements bryophytiques et intérêt patrimonial. Scientific Reports of Port-Cros national Park 22 : 67-194.
Hugonnot V., 2010. Mousses et hépatiques de Païolive (Ardèche et Gard, France). Société btanique du Centre-Ouest. Jarnac, 293 p.
Hugonnot V. & Chavoutier J.-L., 2021. Les bryophytes de France, 1 – Anthocérotes et Hépatiques. Éd. Biotope, Mèze, 652 p.
Quézel P. & Médail F., 2003. Que faut-il entendre par « forêts méditerranéennes » ? Forêt Méditerranéenne XXIV (1) : 11-31.
Sotiaux A., Pioli A., Royaud A., Schumacker R. & Vandenpoorten A., 2007. A checklist of the bryophytes of Corsica (France): new records and a review of the literature. Journal of Bryology 29 : 41-53, doi : 10.1179/174328207X171872.
Sites internet
https://www.grandsitedefrance.com/component/content/article/71-gorges-de-lardeche.html, consulté le 19 janvier 2023.
Remerciements
Les auteurs souhaitent remercier Nicolas Bianchin (CBNMC) pour la relecture du texte et les conseils avisés.


