
Nouvelles interprétations phytosociologiques de la végétation de la Tunisie centrale
Title
New phytosociological interpretations of the vegetation of central Tunisia
Résumé
La reprise d’une publication de 1954 sur la végétation de la Tunisie centrale selon les paradigme et nomenclature phytosociologiques actuels a permis de définir vingt-trois associations, sept alliances et un ordre au sein de classes steppiques et halophiles nord-africaines (Lygeo-Macrochloetea tenacissimae, Pergulario-Pulicarietea crispae, Notoceratetea bicornis, Salicornietea fruticosae, Pegano-Salsoletea vermiculatae, Saginetea maritimae).
Abstract
The revision of a 1954 publication on the vegetation of central Tunisia according to the current phytosociological paradigm and nomenclature has made it possible to define twenty-three associations, seven alliances and one order within the North African steppe and halophytic classes (Lygeo-Macrochloetea tenacissimae, Pergulario-Pulicarietea crispae, Notoceratetea bicornis, Salicornietea fruticosae, Pegano-Salsoletea vermiculatae, Saginetea maritimae).
Il y a soixante-dix ans, Gilbert Long publiait son travail de thèse sur la végétation de la Tunisie centrale, territoire centré sur la ville de Kasserine (Long, 1954 ; carte 1). Sur le plan géomorphologique, l’auteur distinguait les hauts plateaux (au-dessus de 1 000 m d’altitude : Ben Driès, Bled Zelfane), les hautes plaines sablonneuses (500 à 900 m), les basses plaines de la région gamoudienne (de Gamouda, au nord-ouest de Sidi Bou Zid) et les plaines salées ou sebkhas (dépressions à fond plat, généralement inondables, où les sols salés limitent la végétation, pouvant être lacustres ou en communication avec la mer). En dehors du tableau VI de cet auteur, les altitudes des relevés ne sont malheureusement pas indiquées. En outre, plusieurs des associations décrites sont invalides (noms de syntaxons basés sur trois taxons, alors que le maximum doit être de deux ; exemple « Peganeto-Sisymbrieto-Mesembrianthemetum crystallini » en p. 163 du mémoire) ou illégitimes (utilisation du terme sebkharum pour dénommer des syntaxons halophiles). D’autre part, beaucoup de tableaux de steppes et sansouires sont biologiquement hétérogènes, mêlant végétation vivace et végétation thérophytique. On propose de reprendre ici ces données et de les réinterpréter selon le paradigme et le code de nomenclature phytosociologiques actuels. Ce travail s’inscrit dans la poursuite de synthèses antérieures sur la végétation nord-africaine, par exemple de Foucault & Lazare (2020), de Foucault & Noble (2023), de Foucault (2024), d’autres publications étant citées dans le cours du texte.
Seront successivement abordées la végétation ligneuse, la végétation herbacée non halophile (= glycophile) des steppes (vivace puis thérophytique) et la végétation halophile (vivace puis thérophytique). La nomenclature des taxons suit Plants of the World Online (POWO) ; les signes j et ! désignent respectivement une forme juvénile d’un taxon potentiellement arbustif ou arborescent et une forme plus développée que la moyenne des individus du taxon ; dans les tableaux et les listes synsystématiques du texte, le signe * remplace ‘subsp.’ ou ‘var.’. Pour simplifier la présentation des résultats obtenus, on utilisera parfois la notion de groupe sociologique, défini comme un ensemble de taxons à affinités sociologiques, notamment mis en évidence lors de la mise en ordre des tableaux phytosociologiques.
1. La végétation ligneuse
La végétation ligneuse est d’abord représentée par un boisement décrit sur seulement un petit nombre de relevés par Long en son tableau I sous le nom de Pinetum halepensis juniperetosum phoeniceae selon la liste synsystématique suivante (sur quatre relevés) : Juniperus phoenicea *ph. ! 4, Pinus halepensis 3, Juniperus oxycedrus *o. ! 2, Quercus ilex 2, Phillyrea media ! 1, Pistacia atlantica 1. Comme les montrent les photos 3 (planche II) et 6 (planche III) de l’auteur, il apparaît effectivement que J. phoenicea est bien représenté par des formes arborescentes munies d’un tronc (il en est de même de J. oxycedrus, comme d’ailleurs en Algérie ; cf. de Foucault, 2020 : 48 et photo A58) et donc que la végétation décrite est plutôt originale : Pino halepensis-Juniperetum phoeniceae (Long 1954) ass. nov. et stat. nov. hoc loco [basionyme : Pinetum halepensis juniperetosum phoeniceae Long 1954 (Ann. Serv. Bot. Agron. Tunisie 27 : 57)], typus nominis hoc loco : composante arborescente du relevé 2 du tableau I in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte).
Par ailleurs, en considérant en détail le tableau VI de l’auteur décrivant le Ferulo tunetanae-Ephedretum nebrodensis Long 1954, sur lequel on reviendra plus loin, on note l’existence d’un fourré rupicole (sur 11 relevés) à Searsia tripartita V, Asparagus albus V, Juniperus phoenicea *ph. IV, Periploca laevigata IV, Prasium majus III, Jasminum fruticans III, Rhamnus lycioides *oleoides III, Rh. myrtifolia II, Phillyrea media +, Olea europaea j V, Pistacia atlantica j V, un fourré original un peu intermédiaire entre Pistacio lentisci-Rhamnetea alaterni Julve ex B. Foucault & Julve in B. Foucault 2020 méditerranéens et Zizipho loti-Searsietea tripartitae B. Foucault ex B. Foucault 2020 steppiques. On renonce pourtant à le nommer au rang association, car les relevés de Long dans ce tableau VI sont en présence-absence et non quantifiés, de sorte qu’on ne peut en retirer un holotype valide.
2. La végétation glycophile
2.1. La végétation glycophile vivace
Le tableau 1 hoc loco synthétise en vingt-et-une colonnes cette végétation et fait apparaître une césure majeure entre les colonnes 11 et 12.
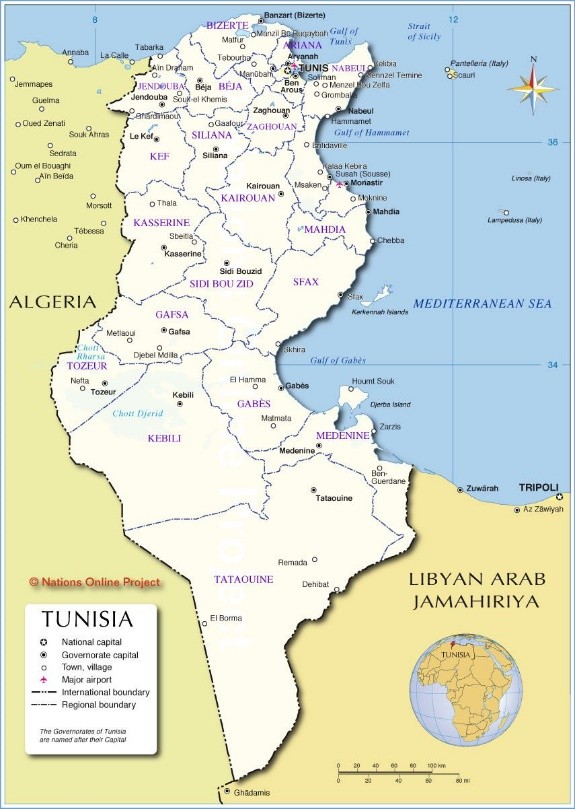
L’ensemble formé par les colonnes 1 à 11 est différencié par un imposant groupe sociologique à Macrochloa tenacissima, taxon qui renvoie aux Lygeo sparti-Macrochloetea tenacissimae Rivas Mart. 1978. On va pouvoir injecter cet ensemble dans le tableau 1 d’une synthèse précédente (de Foucault & Noble, 2023). Une fois cette opération réalisée (tableau 2 : colonne DTa à l’extrême droite ; nomenclature partiellement actualisée), on se rend compte qu’il s’intègre bien aux Stipellulo parviflorae-Lygeetalia sparti B. Foucault & Noble 2023, non loin du Launaeo angustifoliae-Lygeion sparti Le Houérou 1969, ce qui est assez logique pour deux ensembles tunisiens, tout en étant original en lui-même. Une césure secondaire sépare les colonnes 1 à 6 et 7 à 11. Cet ensemble rapproche les syntaxons suivants :
- colonne 1 : communauté associée au Pino halepensis-Juniperetum phoeniceae, cependant réduite à quatre relevés ;
- colonne 2 : « Junipereto-Rosmarineto-Artemisietum campestris Long 1954 » nom. inval., redéfini ici comme Fumano thymifoliae-Thymetum algeriensis G. Long ex ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H) du relevé 6 du tableau II in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; végétation des basses plaines ;
- colonne 3 : Plantagini albicantis-Artemisietum campestris Long 1954 p. p., bien plus original que le syntaxon de la colonne 7 (cf. infra) ; en l’absence de typification, on propose d’affecter ce nom valide à cette colonne 3, lectotypus nominis hoc loco: composante vivace (Ch + H) du relevé 8 du tableau IV in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; végétation surtout des hauts plateaux et des hautes plaines ;
- colonnes 4 à 6 : Rosmarino officinalis-Stipetum tenacissimae Long 1954, muté ici en Salvio rosmarini-Macrochloetum tenacissimae nom. mut. hoc loco; comme Salvia rosmarinus est retenu en tant que premier taxon éponyme, la colonne 4 correspondant au « rosmarineto-cistetosum libanotidis G. Long 1954 » nom. inval. doit être la variation typicum ; lectotypus nominis hoc loco de l’association et du typicum : composante vivace (Ch + H) du relevé 2 du tableau III in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en changeant Stipa tenacissima en Macrochloa t.) ; les autres variations correspondant aux colonnes 5 et 6 restent à préciser et à nommer ; comme le précédent, végétation surtout des hauts plateaux et des hautes plaines ;
- colonne 7, une autre partie du Plantagini albicantis-Artemisietum campestris Long 1954 p. p., laissée à part car définie par seulement quatre relevés ;
- colonnes 8 et 9 : « Rosmarineto-Stipeto-Rumicetum papillosae Long 1954 » nom. inval., redéfini ici comme Diantho serrulati-Macrochloetum tenacissimae G. Long ex ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H) du relevé 8 du tableau V in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte), la colonne 8 correspondant alors au typicum ; végétation des basses plaines gamoudiennes ;
- colonne 10 : Launaeo mucronatae-Erodietum glaucophylli Long 1954, lectotypus nominis hoc loco: composante vivace (Ch + H) du relevé 6 du tableau VII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte), de la région gamoudienne ;
- enfin colonne 11 : Ferulo tunetanae-Ephedretum villarsii Long 1954 muté ici en Ferulo tunetanae-Ephedretum nebrodensis nom. mut. hoc loco; ce syntaxon rupicole ne peut être lectotypifié car les onze relevés sont en présence-absence ; quoi qu’il en soit, il montre un bel exemple de végétation chasmophytique vicariante des syntaxons des Asplenietea trichomanis avec Umbilicus horizontalis, Rupicapnos numidica, Malva subovata subsp. o., Capparis spinosa, Chaenorhinum flexuosum, Sedum dasyphyllum et les fougères Asplenium ceterach, Hemionitis pteridioides, Cosentinia vellea ; son placement dans les Lygeo-Stipetea provient sans doute du fait que les listes de l’auteur sont trop larges, mélangeant la végétation des fentes rocheuses et la végétation voisine ; en un mot, un précieux syntaxon à réviser et à typifier.
À l’exception de la colonne 11, complexe, tout cet ensemble de dix colonnes possède bien la valeur d’une alliance originale au sein des Lygeo-Macrochloetea tenacissimae, le Deverro scopariae-Thymion algeriensis all. nov. hoc loco, caractérisée notamment par Thymus algeriensis, Deverra scoparia, Artemisia herba-alba, Paronychia kapela subsp. k., Helianthemum kahiricum, H. virgatum subsp. africanum, Pseudopodospermum undulatum, Ebenus pinnata, Moraea sisyrinchium, Astragalus armatus, Cardopatium amethystinum, typus nominis hoc loco : le Diantho serrulati-Macrochloetum tenacissimae G. Long ex nov.
Les colonnes 12 à 21 de ce même tableau 1 forment un ensemble floristiquement assez distinct du précédent. La colonne 12 est une végétation vivace à Peganum harmala fort pauvre car issue d’un complexe surtout thérophytique (cf. infra, Sisymbrio irionis-Mesembryanthemetum crystallini prov.), qui rappelle cependant le Pegano harmalae-Bassietum muricatae décrit par Lemée (1953) du Sahara.
Avec Stipagrostis pungens et Centropodia forskaolii, la colonne 13 relève de l’Aristidion pungentis Géhu et al. 1994 muté ici en Stipagrostion pungentis nom. mut. hoc loco ; il s’agit de l’Aristido pungentis-Rumicetum laceri G. Long 1954, valide et muté ici en Stipagrostio pungentis-Rumicetum rosei nom. mut. hoc loco, lectotypus nominis hoc loco : composante vivace (G + Ch + H) du relevé 7 du tableau X in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte en changeant Aristida pungens en Stipagrostis p. et Rumex tingitanus var. lacerus en R. roseus) ; cette végétation colonise des sables grossiers mouvants à très faible capacité d’échange.
La colonne 14 correspond au « Plantagineto-Silybetum eburnei G. Long 1954 », qui comprend aussi une importante composante thérophytique, mais dont on peut considérer la composante vivace comme un Marrubio alyssi-Peganetum harmalae ass. nov. hoc loco, typus nominis hoc loco : composante vivace (G + Ch + H) du relevé 10 du tableau XII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; cette association rudérale se développe plutôt sur des sols alluvionnaires compacts fins et légèrement salés (sols dits « de garâa »), d’où la présence de quelques halophytes.
Laissant pour l’instant de côté la colonne 14 dont on testera la place dans un autre paragraphe (§ 3.1), les colonnes 15 à 21 peuvent provisoirement être rattachées aux Gymnocarpo decandri-Antirrhinetalia fruticosi Guin. 1952 et aux Pergulario tomentosae-Pulicarietea crispae Quézel 1965 (de Foucault & Lazare, 2020). Ce sont
- colonne 15 : le « Chrysanthemeto-Peganetum harmalae Long 1954 », association biologiquement complexe dont on peut isoler la composante vivace en tant que Centaureo dimorphae-Peganetum harmalae ass. nov. hoc loco, typus nominis hoc loco: composante vivace (G + Ch + H) du relevé 2 du tableau XI in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; cette association se développe à la place de l’Eragrostio papposae-Artemisietum campestris (cf. syntaxon suivant) sur des terres cultivées sablonneuses, ce qui explique la place prise par une communauté thérophytique associée (cf. infra, Glebionido coronariae-Anacycletum cyrtolepidioidis) ;
- colonnes 16 à 18 : « Eragrostieto-Zizipheto-Artemisietum campestris Long 1954 p. p. », syntaxon invalide que l’on redéfinit comme Eragrostio papposae-Artemisietum campestris G. Long ex ass. nov. hoc loco, typus nominis hoc loco: composante vivace (G + Ch + H) du relevé 5 du tableau VIII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; végétation des sables fixés d’origine fluviatile et éolienne des basses plaines ;
- colonnes 19 et 20 : une autre partie de l’« Eragrostieto-Zizipheto-Artemisietum campestris Long 1954 », redéfinie comme Zygophyllo scabri-Plantaginetum albicantis ass. nov. hoc loco, typus nominis hoc loco: composante vivace (G + Ch + H) du relevé 25 du tableau VIII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Fagonia kahirina par Zygophyllum scabrum) ;
- colonne 21 : le reste de l’« Eragrostieto-Zizipheto-Artemisietum campestris Long 1954 », redéfinie comme Rumici rosei-Plantaginetum albicantis ass. nov. hoc loco, typus nominis hoc loco: composante vivace (G + Ch + H) du relevé 44 du tableau VIII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Rumex tingitanus var. lacerus par R. roseus).
L’ensemble formé par ces colonnes 15 à 21 possède aussi la valeur d’une alliance caractérisée par Atractylis carduus subsp. c., Echiochilon fruticosum, Peganum harmala, Marrubium alysson, Centaurea dimorpha, Cleome arabica, Stipagrostis ciliata, S. obtusa, Thapsia garganica, le Centaureo dimorphae-Atractylidion cardui all. nov. hoc loco, typus nominis hoc loco : l’Eragrostio papposae-Artemisietum campestris G. Long ex nov.
2.2. La végétation glycophile thérophytique
Le tableau 3 hoc loco synthétise en dix-neuf colonnes cette végétation et fait apparaître une césure majeure entre les colonnes 10 et 11.
Le premier ensemble est caractérisé par le groupe sociologique à Brachypodium distachyon et réunit les syntaxons suivants :
- colonne 1 : le « Junipereto-Rosmarineto-Artemisietum campestris Long 1954 » nom. inval., sans doute à rattacher au syntaxon suivant en tant que variation ;
- colonnes 2 à 4 : le « Rosmarineto-Stipeto-Artemisietum campestris Long 1954 » nom. inval., dont on peut considérer la composante thérophytique comme Anacyclo cyrtolepidioidis-Stipelluletum capensis ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 8 du tableau III in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Stipa retorta par Stipellula capensis) ; végétation surtout des hauts plateaux et des hautes plaines ;
- colonnes 5 et 6 : le Plantagineto-Artemisietum campestris Long 1954, biologiquement complexe, dont on peut isoler la composante thérophytique comme Astragalo crenati-Cutandietum divaricatae ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 4 du tableau IV in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; comme le précédent, végétation surtout des hauts plateaux et des hautes plaines ;
- colonnes 7 et 8 : le « Rosmarineto-Stipeto-Resedetum papillosae Long 1954 » nom. inval., dont on peut extraire la composante thérophytique comme Sileno deserticolae-Resedetum duriaeanae ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 6 du tableau V in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Reseda papillosa par R. duriaeana) ; végétation des basses plaines gamoudiennes ;
- colonne 9 : le Launaeo mucronatae-Erodietum glaucophylli Long 1954, association biologiquement complexe dont on peut mettre en exergue la composante thérophytique comme Pterantho dichotomi-Sulletum spinosissimae ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 6 du tableau VII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Hedysarum spinosissimum par Sulla spinosissima), de la région gamoudienne ;
- colonne 10 : le Ferulo tunetanae-Ephedretum villarsii Long 1954 prov., complexe rupicole dont la composante thérophytique semble posséder la valeur d’un ourlet vernal à Parietaria mauritanica qui pourrait appartenir au Parietarion lusitanico-mauritanicae Rivas Mart. et al. 2001, surtout ibérique, au sein des Cardaminetea hirsutae Géhu 1999 (Rivas-Martínez et al., 2001 ; de Foucault, 2009) ; il s’agit sans doute d’une association originale, mais qui ne peut être validée car les onze relevés sont publiés en présence-absence.
À l’exception de la colonne 10, placée dans les Cardaminetea hirsutae, tout cet ensemble semble devoir s’intégrer aux Notoceratetea bicornis Nègre 1959. En considérant la synthèse de de Foucault & Lazare (2020, tableau XIII), on s’aperçoit que la végétation décrite ici (colonne SHc du tableau 4 ; la colonne SPl voisine sera présentée plus loin, cf. § 3.2) entre dans le premier ordre, les Koelerio pubescentis-Schismetalia barbati Aidoud et al. ex B. Foucault & Lazare 2020, muté ici en Rostrario litoreae-Schismetalia barbati nom. mut. hoc loco, non loin des trois alliances marquées par le groupe sociologique à Muricaria prostrata (AMp, AAl et Lc), sans toutefois s’identifier à l’une d’elles. On peut donc définir au sein de cet ordre le Sileno oliverianae-Hippocrepidion ciliatae all. nov. hoc loco, caractérisé par Hippocrepis ciliata, Silene oliveriana, S. muscipula subsp. deserticola, Reichardia tingitana, Reseda duriaeana, R. decursiva, Ononis reclinata subsp. mollis, Neatostema apulum, Medicago truncatula, Eruca pinnatifida, Carrichtera annua, Anthemis stiparum, Picris asplenioides subsp. a., Didesmus bipinnatus, Helianthemum ledifolium…, typus nominis hoc loco : le Sileno deserticolae-Resedetum duriaeanae nov.
L’ensemble formé par les colonnes 11 à 19 se différencie par le groupe sociologique à Ifloga spicata. Il rassemble :
- colonne 11 : le « Peganeto-Sisymbrieto-Mesembrianthemetum crystallini Long 1954 » nom. inval., dont on pourrait isoler la composante thérophytique en tant que Sisymbrio irionis-Mesembryanthemetum crystallini mais le tableau IX de l’auteur ne fournit qu’une liste synthétique, de sorte qu’on ne peut valablement la valider ; elle pourrait se ranger dans le Mesembryanthemion crystallini Rivas Mart. et al. 1993 au sein des Sisymbrietea officinalis Korneck 1974 (de Foucault, 2012) ; Long précise que ce groupement halonitrophile est lié à l’azote édaphique apporté par la concentration d’animaux sauvages (insectes, petits vertébrés) sur une association initialement halophile ;
- colonne 12 : l’Aristido pungentis-Rumicetum laceri Long 1954, groupement biologiquement complexe dont la composante thérophytique peut être isolée comme Delphinio pubescentis-Silenetum arenarioidis ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 5 du tableau X in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; végétation des sables grossiers mouvants à très faible capacité d’échange associée au Stipagrostio pungentis-Rumicetum rosei.
- colonne 13 : le Chrysanthemo coronarii-Peganetum harmalae Long 1954, autre groupement biologiquement complexe dont la composante thérophytique peut être isolée comme Glebionido coronariae-Anacycletum cyrtolepidioidis ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 4 du tableau XI in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Chrysanthemum coronarium par Glebionis coronaria) ;
- colonnes 14 à 19 : autant de variations de l’« Eragrostieto-Zizipheto-Artemisietum campestris Long 1954 » nom. inval., dont la composante thérophytique peut être isolée comme Noneo calycinae-Stipelluletum capensis ass. nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 15 du tableau VIII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Elizaldia violaceae subsp. calycina par Nonea c.).
Mettant à part la colonne 11 rapprochée des Sisymbrietea officinalis, l’ensemble formé par les colonnes 12 à 19 semble posséder la valeur d’une autre alliance à rattacher aussi aux Notoceratetea bicornis et aux Rostrario litoreae-Schismetalia barbati comme on le constate en injectant la synthèse de ces huit syntaxons dans le tableau synthétique 4 (colonne NCd). Elle montre des affinités certaines avec le Sileno oliverianae-Hippocrepidion ciliatae notamment par le groupe sociologique à Hedypnois cretica, mais s’en distingue assez nettement, ce qui justifie la définition du Noneo calycinae-Cutandion divaricatae all. nov. hoc loco, caractérisé par Nonea calycina, Medicago littoralis, Rumex spinosus, Lolium rigidum, Reseda stricta, Nigella arvensis, Matthiola kralikii, Linaria laxiflora, Echium sabulicola subsp. decipiens, Glebionis coronaria, Limonium bonduellei, Onopordum arenarium, Enarthrocarpus clavatus, se différenciant aussi de l’alliance voisine par des taxons en commun avec l’Iflogo spicatae-Silenion arenarioidis et le Lonchophorion capiomontanae, typus nominis hoc loco : le Glebionido coronariae-Anacycletum cyrtolepidioidis nov.
Vers la fin de son mémoire, Long replace les différentes associations qu’il a décrites en relation les unes avec les autres selon les situations. Les figures 1 et 2 refont le même exercice avec les associations actualisées ici et en reprenant les mêmes légendes qu’utilisa l’auteur.
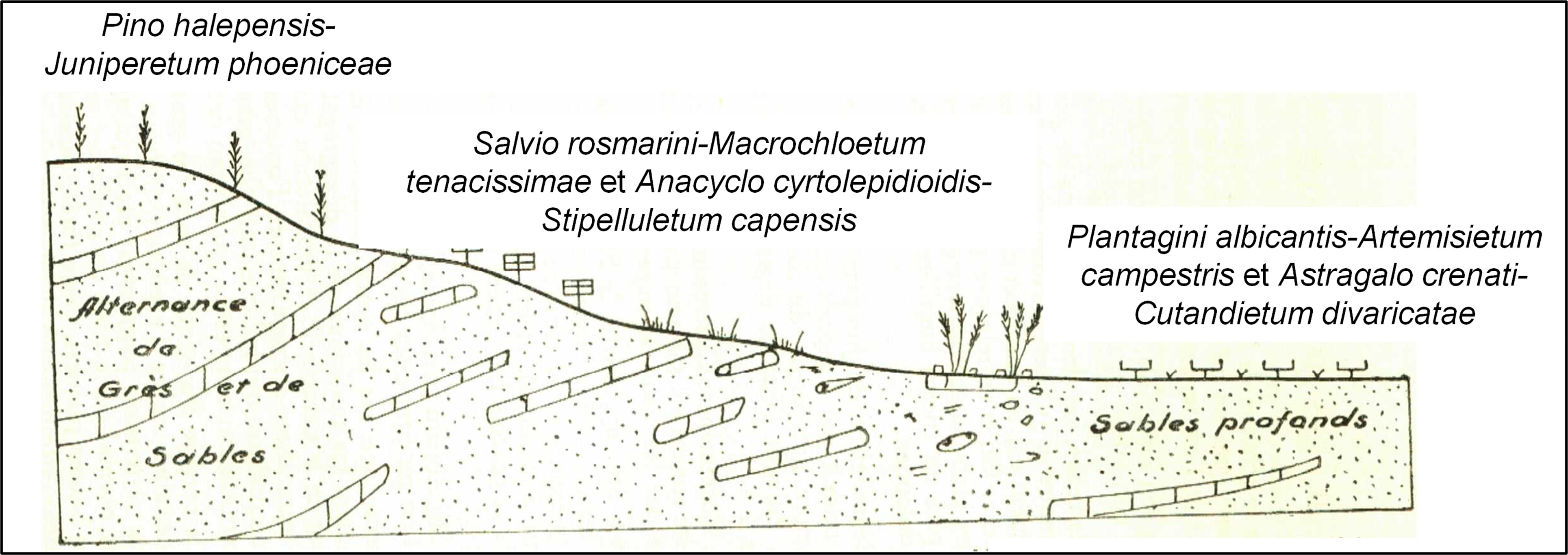
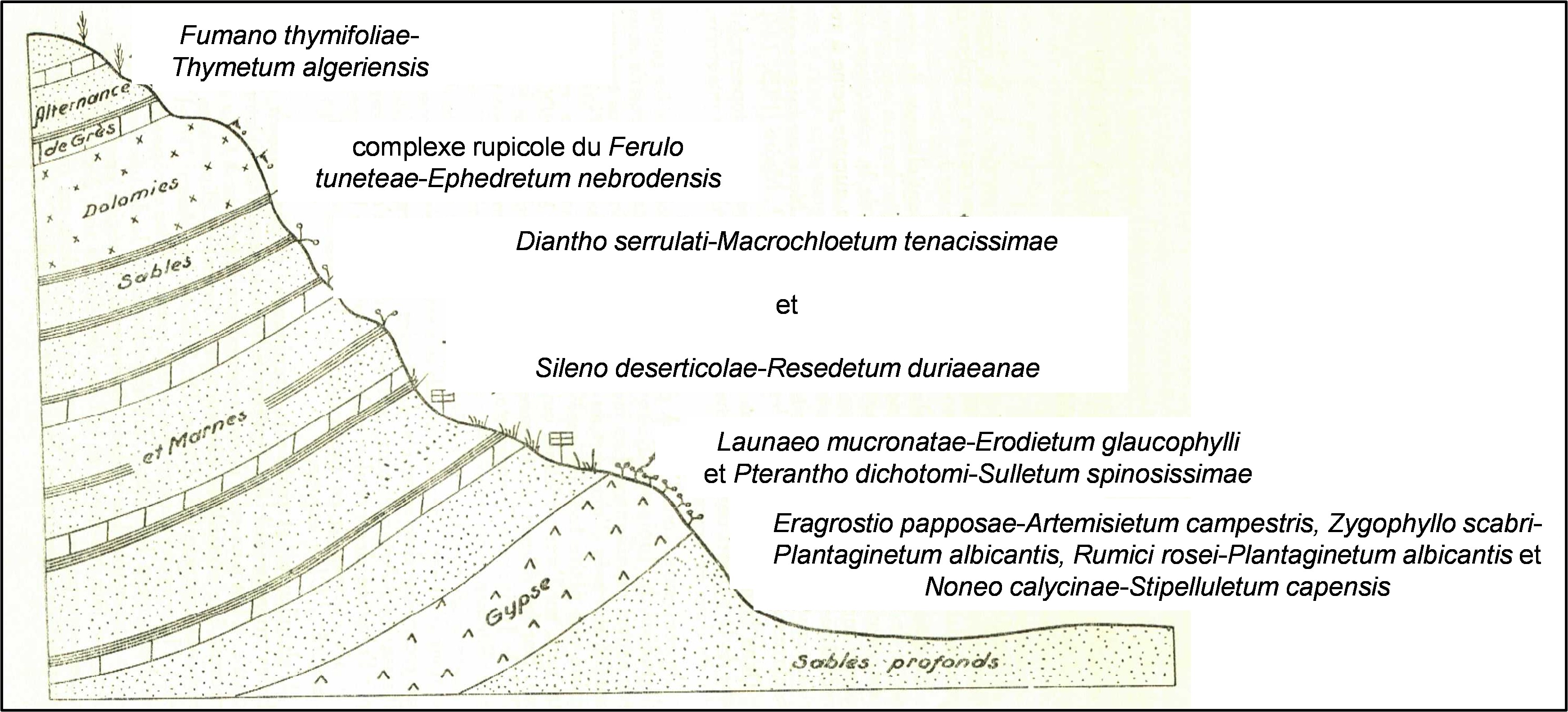
3. La végétation halophile
3.1. La végétation halophile vivace
Le tableau 5 hoc loco synthétise en neuf colonnes cette végétation et fait apparaître une césure majeure entre les colonnes 7 et 8.
L’ensemble formé par les colonnes 1 à 7 rassemble des syntaxons halophiles à divers degrés :
- colonnes 1 et 2 : l’Halocnemetum strobilaceri sebkharum Long 1954 et l’Arthrocnemetum glauci G. Long 1954, qui sont deux noms illégitimes, le premier parce qu’utilisant un terme écologique, le second parce que c’est un homonyme de l’Arthrocnemetum glauci Braun-Blanq. et al. 1952 franco-méditerranéen ; la comparaison des deux colonnes montre qu’il n’y a finalement pas lieu de les distinguer et qu’on peut les rattacher à un unique Caroxylo tetrandri-Arthrocauletum macrostachyi (G. Long 1954) ass. nov. hoc loco; typus nominis hoc loco : composante vivace (Ch + G) du relevé 3 du tableau XV in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Arthrocnemum glaucum par Arthrocaulon macrostachyum et Salsola tetrandra par Caroxylum tetrandrum) ;
- colonne 3 : « Limoniastreto-Suaedeto-Salsoletum cruciatae Long 1954 » nom. inval., redéfini ici comme Suaedo vermiculatae-Caroxyletum tetrandri ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H + G) du relevé 1 du tableau XVI in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Salsola tetrandra par Caroxylum tetrandrum) ;
- colonne 4 : le Suaedetum fruticosae sebkharum Long 1954 nom. illeg., renommé ici Suaedetum monoico-verae (G. Long 1954) ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H + G) du relevé 2 du tableau XVII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Suaeda fruticosa var. brevifolia par S. monoica et S. fruticosa par S. vera) ;
- colonne 5 : Salicornieto-Sperguletum marginatae Long 1954, valide mais que l’on propose de muter en Spergulario mediae-Salicornietum fruticosae nom. mut. hoc loco pour tenir compte de la dominance de Salicornia fruticosa (incl. S. arabica), lectotypus nominis hoc loco: composante vivace (Ch + H + G) du relevé 2 du tableau XXI in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en remplaçant Spergula marginata par Spergularia media) ;
- colonne 6 : Traganeto-Reaumurietum vermiculatae sebkharum Long 1954 nom. illeg., validé plus simplement en Reaumurio vermiculatae–Traganetum nudati (G. Long 1954) ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H + G) du relevé 14 du tableau XX in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ;
- colonne 7 : Salsoleto-Atriplicetum halimi sebkharum Long 1954 nom. illeg., validé ici en Caroxyletum vermiculato-tetrandri (G. Long 1954) ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H) du relevé 1 du tableau XVIII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en changeant Salsola tetrandra en Caroxylum tetrandrum et S. vermiculata en C. vermiculatum) ;
- colonnes 8 et 9 : Salsoletum vermiculatae-villosae sebkharum Long 1954 nom. illeg. ; la colonne 9 est la seule validée ici en Plantagini albicantis-Caroxyletum vermiculati (G. Long 1954) ass. nov. hoc loco, typus nominis hoc loco: composante vivace (Ch + H + G) du relevé 8 du tableau XIX in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte, en changeant Salsola vermiculata en Caroxylum vermiculatum).
Comme le précise Long en p. 234 de son mémoire, ces associations s’ordonnent le long de gradients de salinité selon la figure 3.
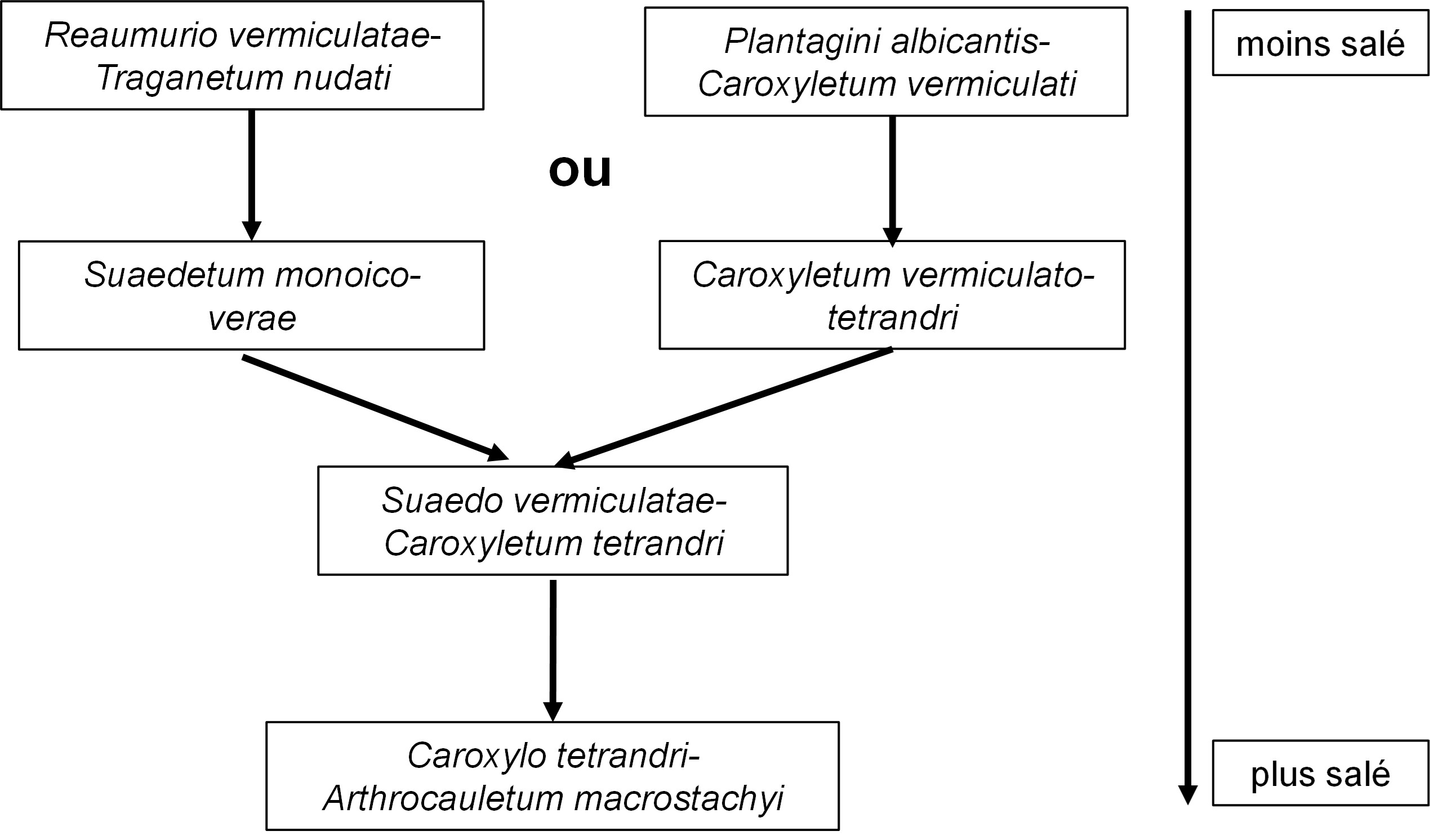
Pour interpréter l’ensemble formé par les colonnes 1 à 7, il convient de le reporter dans la synthèse globale des Salicornietea fruticosae (de Foucault, 2021a, sub Sarcocornietea frutricosae) ; on obtient ainsi le tableau 6 hoc loco, lequel montre que la nouvelle unité (colonne SCt) entre bien dans les Limoniastretalia guyoniani Guin. 1951, sans toutefois se rattacher aux trois alliances qui y sont déjà définies. On peut définir ici le Suaedo monoicae-Caroxylion tetrandri all. nov. hoc loco, caractérisé ou différencié des alliances voisines par Suaeda monoica, Caroxylon vermiculatum, C. tetrandrum et Atriplex halimus (qui y trouvent leur optimum pour la classe), A. glauca, Artemisia herba-alba, Zygophyllum cornutum, Frankenia laevis, Aeluropus lagopoides, typus nominis hoc loco : le Suaedo vermiculatae-Caroxyletum tetrandri nov.
Le petit ensemble formé par les colonnes 8 et 9 est assez distinct des unités précédentes ; en remarquant la présence significative de Peganum harmala et Caroxylon vermiculatum (= Salsola vermiculata), on peut plutôt tester son placement dans les Pegano harmalae-Salsoletea vermiculatae Braun-Blanq. & O. Bolòs 1958, muté en Pegano harmalae-Caroxyletea vermiculati nom. mut. hoc loco, dont une synthèse assez large est aussi disponible (de Foucault, 2015) et reprise dans le tableau 7 hoc loco. Dans cette synthèse, on a déjà précisé que quatre des cinq ordres qui sont rattachés à cette classe par les phytosociologues espagnols y induisent une forte hétérogénéité. Pour le présent propos, on se focalisera donc sur le cœur de la classe, à savoir les Salsolo vermiculatae-Peganetalia harmalae Braun-Blanq. & O. Bolòs 1958, muté en Caroxylo vermiculati-Peganetalia harmalae nom. mut. hoc loco, profitant de cette actualisation pour muter tous les noms des alliances en fonction du référentiel nomenclatural suivi ici. On injecte d’une part les colonnes 8 et 9 du tableau 5, d’autre part la colonne 14 du tableau 1 qui toutes possèdent des affinités avec cette classe. On voit tout de suite que ces trois dernières colonnes s’intègrent assez bien au Caroxylo vermiculati-Peganion harmalae Braun-Blanq. & O. Bolòs 1954, l’alliance type des Caroxylo vermiculati-Peganetalia harmalae. Mais surtout, on s’aperçoit qu’une césure majeure passe entre les colonnes 14 et SSm et que, finalement, cet ordre doit être divisé en deux, l’un gardant le nom de 1958, l’autre pouvant être dénommé Sodo oppositifoliae-Suaedetalia verae ord. nov. hoc loco, caractérisé ou différencié du premier par Soda oppositifolia, Sonchus tenerrimus, Suaeda vera, Caroxylon genistoides, Atriplex glauca, Zygophyllum creticum, Malva arborea, Limbarda crithmoides subsp. c., Beta vulgaris subsp. maritima, typus nominis hoc loco : le Sodo oppositifoliae-Suaedion mollis Rigual 1972 (Publ. Inst. Est. Alicantinos 2 (1) : 97, sub Salsolo-Suaedion) ; on y a ramené aussi l’Haloxylo salicornici-Atriplicion glaucae Rivas Goday & Rivas Mart. ex Rigual 1972 corr. Alcaraz, P. Gómez, De la Torre, Ríos & J. Alvarez 1991 (colonne HAg) placé dans les Helichryso stoechadis-Santolinetalia squarrosae Peinado & Martinez Parras 1984 dans la synthèse de 2015, dont il n’est pas le type nomenclatural. On remarquera enfin que les Caroxylo vermiculati-Peganetalia harmalae ne sont donc actuellement pas connus de France.
Le mémoire de G. Long ne fait guère apparaître de végétation halophile plutôt hémicryptophyto-géophytique susceptible de se rattacher aux Juncetea maritimi Tüxen & Oberd. 1958 ; seul un relevé est cité en p. 201. Selon une synthèse de 2021, avec Aeluropus lagopoides, il se rapprocherait de l’Aeluropodo lagopoidis-Juncion rigidi B. Foucault 2021 de l’Égypte et du Sud tunisien (de Foucault, 2021b).
3.2. La végétation halophile thérophytique
Le tableau 8 hoc loco synthétise en onze colonnes cette végétation occupant les ouvertures des sansouires des sebkhas et fait apparaître une césure majeure entre les colonnes 6 et 7.
Avec Frankenia pulverulenta, Sphenopus divaricatus, Limonium echioides et Parapholis incurva, l’ensemble formé par les six premières colonnes se rattache assez clairement aux Frankenietalia pulverulentae Rivas Mart ex Castrov. & Porta 1976 ; ce sont
- colonnes 1 à 3 : associées en tant que végétation thérophytique au Caroxylo tetrandri-Arthrocauletum macrostachyi et au Suaedetum monoico-verae; celle-ci paraît toutefois insuffisamment caractérisée pour donner lieu à un statut d’association ;
- colonne 4 : associée au Suaedo vermiculatae-Caroxyletum tetrandri et définie ici comme Trigonello maritimae-Filaginetum mareoticae nov. hoc loco, typus nominis hoc loco: composante thérophytique (T) du relevé 8 du tableau XVI in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ;
- colonne 5 : associée au Reaumurio vermiculatae–Traganetum nudati mais réduite à trois relevés ;
- colonne 6 : associée au Spergulario mediae-Salicornietum fruticosae, mais floristiquement peu caractérisée ;
- colonne 7 : associée à une partie du Caroxyletum vermiculato-tetrandri et de transition vers l’ensemble suivant.
Tout cet ensemble possède la valeur d’une alliance et, pour l’interpréter, il faut le comparer au reste des Frankenietalia pulverulentae, ce que réalise le tableau 9 hoc loco en reprenant une partie d’une synthèse antérieure (de Foucault & Bioret, 2010), avec le Frankenion pulverulentae Rivas-Mart. ex Castrov. & Porta 1976 (colonne Fp), le Polypogonion subspathacei Gamisans 1990 (Ps) et le Limonion avei Brullo 1988 (La). On voit que la quatrième colonne (SSd) ne peut se rattacher à celles-ci et mérite le rang d’alliance : Sphenopodo divaricati-Spergularion diandrae all. nov. hoc loco, caractérisée ou différenciée des autres de l’ordre par Spergularia diandra, Reichardia tingitana, Rostraria salzmannii, Beta macrocarpa, Mesembryanthemum nodiflorum, Limonium echioides, plus quelques thérophytes des steppes déjà décrites (Schismus barbatus, Anacyclus monanthos subsp. cyrtolepidioides…), typus nominis hoc loco : le Trigonello maritimae-Filaginetum mareoticae nov. Il est possible que cette alliance atteigne aussi l’Algérie nord-orientale comme le montre le relevé 3 du tableau 3 in Kaabèche et al. (1995).
L’ensemble des colonnes 8 à 11 correspond à une végétation de steppes plus faiblement halophiles, où le cortège des thérophytes des Notoceratetea bicornis est bien plus marqué que celui des Frankenietalia pulverulentae. La mieux caractérisée est assurément la colonne 10, reliée au Marrubio alyssi-Peganetum harmalae : Silybo eburnei-Mantisalcetum duriaei ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique (T) du relevé 7 du tableau XII in Long (1954, Ann. Serv. Bot. Agron. Tunisie 27 hors texte) ; avec Silybum eburneum, des Centaurea à involucres épineux (C. sicula, C. melitensis), Carthamus lanatus, peut-être Mantisalca duriaei, plus rarement Carduus getulus et Onopordum arenarium, on peut se demander si une eutrophisation encore plus marquée ne mènerait pas à une association des Carthametalia lanati Brullo in Brullo & Marcenò 1985 et des Onopordetea acanthii Braun-Blanq. 1964. Cet ensemble possède la valeur d’une alliance oligohalonitrophile bien caractérisée par Plantago lagopus, Spergularia diandra, Linaria laxiflora, Malva parviflora, Mantisalca duriaei, Anacyclus clavatus, Beta macrocarpa, Diplotaxis erucoides, Volutaria tubuliflora, Asteriscus aquaticus, Hordeum murinum (cf. subsp. leporinum), Astragalus hamosus, Centaurea melitensis, Silene vivianii, Silybum eburneum, Spergulario diandrae-Plantaginion lagopodis all. nov. hoc loco, typus nominis hoc loco : le Silybo eburnei-Mantisalcetum duriaei nov. L’injection de cet ensemble dans le tableau synthétique 4 (colonne SPl) montre que cette alliance s’intègre bien aux Rostrario litoreae-Schismetalia barbati déjà cités, non loin du Noneo calycinae-Cutandion divaricatae.
4. Conclusion
Au-delà de l’importante contribution qu’a apportée cette publication de Long de 1954 pour la connaissance de la végétation centro-tunisienne, elle a permis d’étoffer la structure de six classes phytosociologiques : Lygeo sparti-Macrochloetea tenacissimae (ajoutant une alliance, le Deverro scopariae-Thymion algeriensis), peut-être Pergulario tomentosae-Pulicarietea crispae (ajoutant le Centaureo dimorphae-Atractylidion cardui), les Notoceratetea bicornis (ajoutant le Sileno oliverianae-Hippocrepidion ciliatae, le Noneo calycinae-Cutandion divaricatae et le Spergulario diandrae-Plantaginion lagopodis), les Salicornietea fruticosae (ajoutant le Suaedo monoicae-Caroxylion tetrandri), les Pegano harmalae-Caroxyletea vermiculati (ajoutant les Sodo oppositifoliae-Suaedetalia verae), les Saginetea maritimae (ajoutant le Sphenopodo divaricati-Spergularion diandrae).
Bibliographie
Foucault B. (de), 2009. Contribution au prodrome des végétations de France : les Cardaminetea hirsutae Géhu 1999. Journal de botanique de la Société botanique de France 48 : 49-70.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Sisymbrietea officinalis Korneck 1974. Journal de botanique de la Société botanique de France 58 : 55-116.
Foucault B. (de), 2015. Contribution au prodrome des végétations de France : les Pegano harmalae-Salsoletea vermiculatae Braun-Blanq. & O. Bolòs 1958. Journal de botanique de la Société botanique de France 72 : 41-59.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021a. Contribution à une synthèse des Sarcocornietea fruticosae. Carnets botaniques 59 : 1-19 + 17 tableaux, https://doi.org/10.34971/8h0c-0b94.
Foucault B. (de), 2021b. Contribution à une synthèse des Juncetea maritimi. Carnets botaniques 68 : 1-30 + 22 tableaux, https://doi.org/10.34971/nbat-mh09.
Foucault B. (de), 2024. Nouvelles interprétations phytosociologiques des subéraies marocaines. Carnets botaniques 228 : 1-8, https://doi.org/10.34971/9BFQ-G158.
Foucault B. (de) & Bioret F., 2010. Contribution au prodrome des végétations de France : les Saginetea maritimae Westhoff et al. 1962. Journal de botanique de la Société botanique de France 50 : 59-83.
Foucault B. (de) & Lazare J.-J., 2020. Contribution à une synthèse phytosociologique des steppes d’Afrique du Nord (hors Lygeo sparti-Stipetea tenacissimae). Botanique 6 : 53-116.
Foucault B. (de) & Noble V., 2023. Contribution à une synthèse des Lygeo sparti-Macrochloetea tenacissimae. Carnets botaniques 149 : 1-17, https://doi.org/10.34971/V632-0572.
Kaabèche M., Gharzouli R. & Géhu J.-M., 1995. Observations phytosociologiques sur le Tell et les hauts plateaux de Sétif (Algérie). Documents phytosociologiques, n. s., XV : 117-125.
Lemée G., 1953. Contribution à la connaissance phytosociologique des confins saharo‑marocains. Les associations à thérophytes des dépressions sableuses et limoneuses non salées et des rocailles aux environs de Beni‑Ounif. Vegetatio 4 : 137‑154.
Long G., 1954. Contribution à l’étude de la végétation de la Tunisie centrale. Annales du service botanique et agronomique de Tunisie 27 : 1-388 et XXI tableaux.
Rigual A., 1972. Flora y vegetación de la provincia de Alicante (El paisaje vegetal alicantino). Publicationes del Instituto de Estudios Alicantinos 2 (1) : 1‑403.
Rivas-Martínez S., Fernández González F., Loidi J., Lousã M. & Penas A., 2001. Syntaxonomical checklist of vascular plant communities in Spain and Portugal to association level. Itinera Geobotanica 14 : 5-341.


