
Nouvelles considérations phytosociologiques sur la végétation de la Grèce
Title
New phytosociological considerations on the vegetation of Greece
Résumé
Sur la base de travaux phytosociologiques antérieurs, revus à la lumière du paradigme synusial, et de données nouvelles acquises lors d’un voyage récent, l’auteur apporte des éléments sur la végétation de Grèce continentale et de Crète illustrés grâce à plus d’une soixantaine de photos. Trente-cinq associations originales sont définies, ainsi que dix alliances et un ordre.
Abstract
On the basis of previous phytosociological works, revised in the light of the synusial paradigm, and of new data acquired during a recent trip, the author brings elements on the vegetation of continental Greece and Crete illustrated with more than sixty photographs. Thirty five original associations are defined, as well as ten alliances and one order.
De la mi-mai au début du mois de juin 2022, un voyage en Grèce continentale a permis de parcourir plusieurs sites naturels et historiques. La carte 1 en précise les diverses étapes : Vólos (point de départ pour le Pélion et Makrinitsa), puis Véroia (et Vergina), Thessalonique (Thessaloniki), Flórina, Kalabáka (souvent écrit Kalambaka, dans les Météores, avec la ville voisine Kastraki absente de la carte), Ioánnina, Nafpaktos (absent de la carte, à l’est de Mesolóngion, sur la rive nord du golfe de Corinthe), enfin Athènes (Athens) avec Thrakomakedones et le mont Parnitha. Des données phytosociologiques ont pu être rassemblées pour préparer ce compte-rendu agrémenté de photographies de flore et de végétation. La nomenclature botanique suit Euro+Med PlantBase, le signe * remplace ‘subsp.’ ou ‘var.’ dans les tableaux et les relevés isolés ; les noms des variations autonymes sont abrégés à leur initiale. Les tableaux phytosociologiques détaillés 2 et 8 à 11 sont intégrés au texte alors que les tableaux synthétiques 1 et 3 à 7 sont placés dans un fichier annexe.
1. La végétation forestière
Après un large aperçu de la végétation forestière de la Grèce continentale par Debazac et Mavrommatis (1971), c’est surtout à Barbero et Quézel (1976), puis Gamisans et Hébrard (1979) que l’on doit les apports les plus importants sur ce sujet, avec des relevés et des tableaux réalisés selon la méthode phytosociologique classique. On va le reprendre ici à la lumière du paradigme synusial (de Foucault, 2020). On y a associé les données sur des forêts crétoises issues de la publication de Barbero & Quézel (1980) ; ces données avaient été reprises en 1993 (de Foucault, 1993), mais d’une manière très partielle.
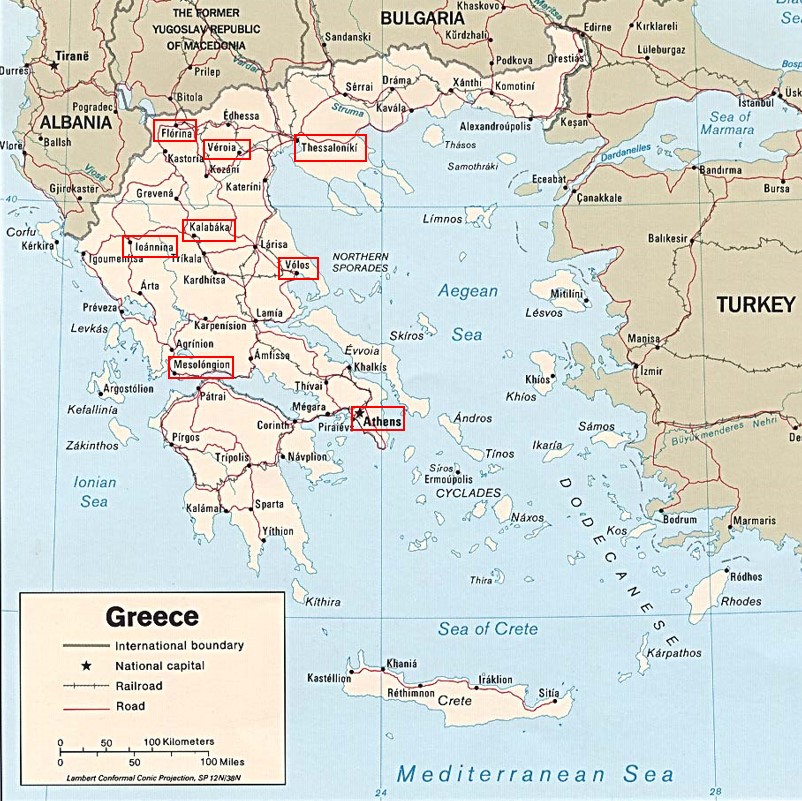
Annexe du tableau 2 – rel. 1 : Athènes, versant nord de l’Acropole ; rel. 2 : Athènes, colline de Philopappos.
* La végétation arborescente
Le tableau 1 synthétise les données sur la végétation arborescente apportées par Barbero et Quézel (1976, syntaxons en BQ) puis Gamisans et Hébrard (1979 ; syntaxons en GH).
Ainsi les colonnes BQ1 et BQ2, partageant Olea europaea subsp. e., décrivent une végétation à rattacher à l’Oleo europaeae-Ceratonion siliquae Braun-Blanq. ex Guin. & Drouineau 1944 (de Foucault, 2021a : 5). L’absence de Quercus ilex dans ces relevés laisse penser qu’il s’agit d’une association différente du Ceratonio siliquae-Oleetum europaeae plutôt ouest-méditerranéen (Guinochet & Drouineau, 1944 ; Molinier, 1954 ; de Bolòs & Molinier, 1958 ; de Foucault, 2021b), développée à une altitude moyenne de 126 m : Oleo europaeae-Pinetum halepensis ass. nov. hoc loco, typus nominis hoc loco : relevé 2 du tableau 2 hoc loco (photo 1). Dans le contexte urbain d’Athènes, cette végétation est souvent pénétrée de taxons exotiques comme Parkinsonia aculeata, des Eucalyptus, des Cupressus…

Les colonnes BQ3 à GH11b du tableau 1 se rapprochent par la combinaison de Quercus coccifera (ici arborescent) et Q. pubescens, moins souvent Cercis siliquastrum et Fraxinus ornus. En particulier, la colonne BQ3 décrit une association de transition entre Oleo-Ceratonion siliquae et Abietion cephalonico-borisii-regis (cf. infra), à une altitude moyenne de 875 m ; elle a été concrètement observée sur les flancs du mont Parnitha, au-dessus d’Athènes :
mont Parnitha, non loin du Regency Casino, N 39° 09’ 09,2’’, E 23° 41’ 12,8’’, 1 074 m, 01 juin 2022, 6 taxons
Quercus coccifera 3, Abies cephalonica (photo 2 ; parfois parasité par Viscum album *abietis) 2, Pinus halepensis 1, Quercus ilex +, Fraxinus ornus *o. +, Cercis siliquastrum +,
typus nominis hoc loco du Pino halepensis-Abietetum cephalonicae ass. nov. hoc loco (photo 3 ; voir aussi photo 32).


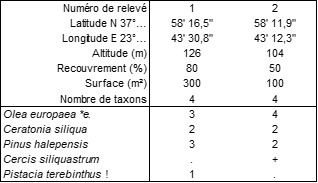
Les colonnes BQ4c et BQ5-8 décrivent le Cercidi siliquastri-Quercetum cocciferae ass. nov. hoc loco, à une altitude moyenne de 430 m, typus nominis hoc loco : composante arborescente du relevé 25 du tableau 4 in Barbero & Quézel (1976, Ecol. Medit. II : 15), en remplaçant Quercus pseudococcifera par Q. coccifera et Quercus brachyphylla par Q. pubescens.
La colonne BQ6-7 se distingue suffisamment des précédentes pour justifier un Cercidi siliquastri-Quercetum ilicis ass. nov. hoc loco à une altitude moyenne de 400 m, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 6 in Barbero & Quézel (1976, Ecol. Medit. II : 19), en remplaçant Acer orientale par A. sempervirens.
La colonne BQ9-10, forte de 54 relevés, décrit une chênaie de Quercus frainetto (photo 4)-châtaigneraie développée à une altitude moyenne de 970 m : Castaneo sativae-Quercetum frainetto ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 5 du tableau 10 in Barbero & Quézel (1976, Ecol. Medit. II : 29).

Les colonnes GH7 et GH11a semblent correspondre à deux variations d’un même Fraxino orni-Quercetum cerridis ass. nov. hoc loco, à une altitude moyenne d’environ 800 m, typus nominis hoc loco de l’association et du typicum (différencié par Quercus coccifera et Ostrya carpinifolia, colonne GH11a) : composante arborescente du relevé 4 du tableau 11 in Gamisans & Hébrard (1979, Doc. Phytosoc., NS, IV hors texte) ; typus nominis hoc loco du quercetosum frainetto subass. nov. hoc loco (différencié par Quercus frainetto, Acer campestre, Sorbus torminalis, colonne GH7) : composante arborescente du relevé 3 du tableau 7 in Gamisans & Hébrard (1979, Doc. Phytosoc., NS, IV hors texte).
L’ensemble des colonnes BQ3 à GH11b paraît correspondre à une alliance originale, le Cercidi siliquastri-Quercion cocciferae all. nov. hoc loco, caractérisée par une combinaison de Quercus coccifera, Q. pubescens, Cercis siliquastrum et Fraxinus ornus, typus nominis hoc loco : Cercidi siliquastri-Quercetum cocciferae nov.
Les colonnes BQ12-13a à GH8 rassemblent des sapinières à Abies cephalonica (photo 2) et A. borisii-regis, essences associées parfois à Juniperus foetidissima, Prunus cocomilia…, boisements souvent developpés au-dessus de 1 200 m. La colonne BQ12-13a correspond à une sapinière peu caractérisée, alors que la colonne BQ13b combine Castanea sativa et Abies borisii-regis à une altitude moyenne de 1 530 m : Castaneo sativae-Abietetum borisii-regis ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 24 du tableau 13 in Barbero & Quézel (1976, Ecol. Medit. II : 38).
La colonne BQ14 décrit une remarquable association forestière de pente à Aesculus hippocastanum à une altitude moyenne de 1 380 m : Aceri monspessulani-Aesculetum hippocastani ass. nov. hoc loco, typus nominis hoc loco : composante arborescente du relevé 2 du tableau 14 in Barbero & Quézel (1976, Ecol. Medit. II : 40).
À une altiude moyenne de 1 600 m, les colonnes BQ16 et BQ25 peuvent s’interpréter comme deux variations d’un unique Junipero foetidissimae-Abietetum cephalonicae ass. nov. hoc loco ; typus nominis hoc loco de l’association et du typicum (colonne BQ16) : composante arborescente du relevé 5 du tableau 16 in Barbero & Quézel (1976, Ecol. Medit. II : 43) ; typus nominis hoc loco du juniperetosum excelsae subass. nov. hoc loco (colonne BQ25), différencié par le taxon éponyme et Acer heldreichii : composante arborescente du relevé 1 du tableau 25 in Barbero & Quézel (1976, Ecol. Medit. II : 65).
La colonne GH8 est originale, malgré le nombre réduit de relevés : Carpino orientalis-Quercetum dalechampii ass. nov. hoc loco, développé à une altitude moyenne de 1 150 m, typus nominis hoc loco : composante arborescente du relevé 1 du tableau 8 in Gamisans & Hébrard (1979, Doc. Phytosoc., NS, IV hors texte).
Ces cinq syntaxons peuvent être réunis dans l’Abietion cephalonico-borisii-regis all. nov. hoc loco, alliance caractérisée par Abies cephalonica, A. borisii-regis, Juniperus foetidissima, Prunus cocomilia, typus nominis hoc loco : Aceri monspessulani-Aesculetum hippocastani nov.
Les colonnes BQ17-18 à GH6 partagent essentiellement Pinus nigra subsp. pallasiana, alors que les essences des alliances précédentes deviennent plus discrètes, à une altitude moyenne de 1 200 m. On peut surtout caractériser l’Abieti cephalonicae-Pinetum pallasianae ass. nov. hoc loco (colonne BQ17-18), typus nominis hoc loco : composante arborescente du relevé 2 du tableau 17 in Barbero & Quézel (1976, Ecol. Medit. II : 48), et l’Abieti borisii-regis-Pinetum pallasianae ass. nov. hoc loco (colonne BQ19), typus nominis hoc loco : composante arborescente du relevé 3 du tableau 19 in Barbero & Quézel (1976, Ecol. Medit. II : 51). Les colonnes GH5 et GH6 semblent correspondre à deux variations d’un même Aceri obtusati-Pinetum pallasianae ass. nov. hoc loco, typus nominis hoc loco de l’association et du typicum (colonne GH5, à environ 1 200 m d’altitude) : composante arborescente du relevé 12 du tableau 5 in Gamisans & Hébrard (1979, Doc. Phytosoc., NS, IV hors texte) ; typus nominis hoc loco du quercetosum pubescentis subass. nov. hoc loco (différencié par Quercus pubescens, Cercis siliquastrum, Acer monspessulanum, colonne GH6, de plus basse altitude : 835 m) : composante arborescente du relevé 1 du tableau 6 in Gamisans & Hébrard (1979, Doc. Phytosoc., NS, IV hors texte). Ces trois associations peuvent être réunis dans le Pinion pallasianae all. nov. hoc loco, alliance essentiellement caractérisée par Pinus nigra subsp. pallasiana, typus nominis hoc loco : Abieti cephalonicae-Pinetum pallasianae nov., syntaxon que l’on peut rapprocher de l’Abietion cephalonico-borisii-regis pour former l’ordre des Abietetalia cephalonico-borisii-regis ord. nov. hoc loco, ensemble globalement caractérisé par Abies cephalonica, A. borisii-regis, Sorbus graeca, Prunus cocomilia, typus nominis hoc loco : Abietion cephalonico-borisii-regis nov. On renonce pour l’instant à rattacher le tout à une classe précise.
Il reste encore l’ensemble formé par les colonnes BQ20b à GH2 correspondant à des hêtraies à Fagus sylvatica et Acer obtusatum développées au-dessus de 1 000 m ; à ce titre on peut le rattacher en extrême irradiation aux Fraxino excelsoris-Quercetea roboris Gillet ex B. Foucault et al. in B. Foucault 2020.
À une altitude moyenne de 1 575 m, on peut définir le Fago sylvaticae-Pinetum heldreichii ass. nov. hoc loco (colonne BQ20b), typus nominis hoc loco : composante arborescente du relevé 6 du tableau 20 in Barbero & Quézel (1976, Ecol. Medit. II : 53). À une altitude moyenne de 1 030 m, on a l’Aceri obtusati-Fagetum sylvaticae ass. nov. hoc loco variant en
- typicum (colonne BQ21a), différencié par Abies borisii-regis et Taxus baccata, typus nominis hoc loco de l’association et du typicum: composante arborescente du relevé 4 du tableau 21 in Barbero & Quézel (1976, Medit. II : 58) ;
- quercetosum pubescentis subass. nov. hoc loco (colonne BQ21b), différencié par le chêne éponyme, Acer campestre et Sorbus graeca, typus nominis hoc loco : composante arborescente du relevé 12 du tableau 21 in Barbero & Quézel (1976, Medit. II : 58).
La colonne BQ23-24 décrit l’Abieti borisii-regis-Fagetum sylvaticae ass. nov. hoc loco à une altitude moyenne de 1 460 m (le calcul de cette moyenne exclut le relevé 12 du tableau 23 où l’altitude de « 100 m » paraît bien aberrante ; sans doute une erreur de saisie), typus nominis hoc loco : composante arborescente du relevé 4 du tableau 24 in Barbero & Quézel (1976, Ecol. Medit. II : 64).
Une alliance originale est requise pour une telle végétation, caractérisée surtout par Fagus sylvatica, Pinus heldreichii, Abies borisii-regis et l’absence ou l’extrême rareté des essences caractérisant le coeur des Fraxino-Quercetea roboris, l’Abieti borisii-regis-Fagion sylvaticae all. nov. hoc loco, typus nominis hoc loco : Aceri obtusati-Fagetum sylvaticae nov. On renonce pour l’instant à la rattacher à un ordre précis.
On peut résumer ainsi le synsystème reconnu pour la végétation forestière arborescente de Grèce continentale :
Pino halepensis-Quercetea ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020
Quercetalia rotundifolio-ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020
Oleo europaeae-Ceratonion siliquae Braun-Blanq. ex Guin. & Drouineau 1944
Oleo europaeae-Pinetum halepensis nov.
?
Cercidi siliquastri-Quercion pubescentis nov.
Abieti cephalonicae-Pinetum halepensis nov.
Cercidi siliquastri-Quercetum cocciferae nov.
Cercidi siliquastri-Quercetum ilicis nov.
Castaneo sativae-Quercetum frainetto nov.
Fraxino orni-Quercetum cerridis nov.
?
Abietetalia cephalonico-borisii-regis nov.
Abietion cephalonico-borisii-regis nov.
Castaneo sativae-Abietetum borisii-regis nov.
Aceri monspessulani-Aesculetum hippocastani nov.
Abieti cephalonicae-Juniperetum foetidissimae nov.
Carpino orientalis-Quercetum dalechampii nov.
Pinion pallasianae nov.
Abieti cephalonicae-Pinetum pallasianae nov.
Abieti borisii-regis-Pinetum pallasianae nov.
Aceri obtusati-Pinetum pallasianae nov.
Fraxino excelsioris-Quercetea roboris Gillet ex B. Foucault et al. in B. Foucault 2020
?
Abieti borisii-regis-Fagion sylvaticae nov.
Fago sylvaticae-Pinetum heldreichii nov.
Aceri obtusati-Fagetum sylvaticae nov.
Abieti borisii-regis-Fagetum sylvaticae nov.
Venons-en aux forêts crétoises. Un premier problème se pose à propos des données de Barbero & Quézel (1980) : l’unité de l’altitude utilisée dans les tableaux de ces auteurs n’est pas vraiment précisée, il semble bien que ce ne soit pas le mètre, mais plutôt la dizaine de mètres ; ainsi, le tableau VIII cite des altitudes allant de 110 à 165, mais d’après le texte décrivant l’Aceri–Berberidetum creticae il s’agit plutôt de 1 100 à 1 650 m. On retiendra donc les altitudes 10 m.
On retrouve tout d’abord l’Oleo–Ceratonion siliquae (tableau 3 : colonnes Ia à IIIc, en moyenne 295 m), avec des associations à conforter, basale (Ia à IVc), à Quercus pubescens (IIa et IIb), à Q. ithaburensis subsp. macrolepis (IIc), à Pinus brutia (IIIa et IIIb), à Cupressus sempervirens (IIIc). Les colonnes Va à VIc (en moyenne 455 m) synthétisent des boisements intermédiaires à Q. pubescens, alors que les colonnes IVb à VIIc rassemblent des forêts plus alticoles (en moyenne 1 010 m) à Acer sempervirens et Cupressus sempervirens, correspondant à l’Aceri sempervirentis-Cupressetum sempervirentis (Barbero & Quézel 1980) B. Foucault 1993 ex 2020 (le relevé type appartient à la colonne VIIb).
* La végétation arbustive intraforestière de Grèce continentale
Le tableau 4 synthétise les données sur la végétation arbustive intraforestière continentale apportées par Barbero et Quézel (1976) ; on a renoncé à faire le même exercice sur les tableaux de Gamisans & Hébrard (1979) car cette végétation y est plus fragmentaire. La flore est ici bien plus diversifiée et bien plus complexe que pour la végétation arborescente ; on se contentera donc de quelques indications et de comparaisons en lien avec une synthèse antérieure (de Foucault, 2021a).
L’ensemble des colonnes BQ1a à BQ8b est caractérisé un fort contingent d’arbustes et arbrisseaux méditerranéens plus ou moins sempervirents. Il se rattache bien aux Pistacio lentisci-Rhamnetea alaterni Julve ex B. Foucault & Julve in B. Foucault 2020. Avec Asparagus aphyllus, Rhamnus lycioides subsp. oleoides, Phlomis fruticosa (photo 5), Hippocrepis emerus subsp. emeroides (photo 6), il se rattache à l’ordre est-méditerranéen des Rubio tenuifoliae-Pistacietalia terebinthi B. Foucault 2020, mais à aucune des alliances connues au sein de cet ordre.


On peut le subdiviser en un premier sous-ensemble (colonnes BQ1a à BQ3) mal caractérisé, où l’on peut distinguer au moins la colonne BQ2 sous le nom de Rubio tenuifoliae-Pistacietum lentisci ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 5 du tableau 2 in Barbero & Quézel (1976, Ecol. Medit. II : 11) en remplaçant Rubia olivieri par R. tenuifolia (photo 18).
Le second sous-ensemble (colonnes BQ4a à BQ8b) est mieux caractérisé et a certainement valeur d’alliance ; mais pour en définir une, il faudrait disposer d’au moins une association valide, ce qui paraît prématuré.
Le vaste ensemble des colonnes BQ9a à BQ24a est difficile à structurer. Une partie assez bien caractérisée est formée par l’ensemble BQ9a à BQ21b où se rencontrent des taxons caractérisant plutôt les Rhamno-Prunetea spinosae (Cornus mas, Clematis vitalba, Lonicera xylosteum, Euonymus europaeus, Ribes uva-crispa, Daphne mezereum, Rosa canina, Crataegus monogyna, Dioscorea communis) et à un degré moindre des taxons orientaux (Hippocrepis emerus subsp. emeroides, Carpinus orientalis j), ainsi que Juniperus oxycedrus subsp. o. Dans la colonne BQ13a apparaît un Lonicera « graeca » dont la nomenclature actuelle ne correspond à rien selon les référentiels récents consultés. On peut au moins y définir l’Euonymo latifolii-Hippocrepidetum emeroidis ass. nov. hoc loco (colonnes BQ14a et BQ14b), typus nominis hoc loco : composante arbustive du relevé 6 du tableau 14 in Barbero & Quézel (1976, Ecol. Medit. II : 40).
On peut retenir cette association pour valider l’Hippocrepido emeroidis-Clematidion vitalbae all. nov. hoc loco, alliance caractérisée par la combinaison de taxons orientaux (Hippocrepis emerus subsp. emeroides, Carpinus orientalis j), de Juniperus oxycedrus subsp. o. et de taxons caractérisant les Rhamno-Prunetea spinosae (Cornus mas, Clematis vitalba, Lonicera xylosteum, Euonymus europaeus, Ribes uva-crispa, Daphne mezereum, Rosa canina, Crataegus monogyna, Dioscorea communis), classe à laquelle se rattache donc cette alliance (mais dans un ordre encore imprécis), typus nominis hoc loco : Euonymo latifolii-Hippocrepidetum emeroidis nov.
* La végétation arbustive intraforestière de Crète
Le tableau 5 synthétise les données sur la végétation arbustive intraforestière de Crète selon les données de Barbero & Quézel (1980), non utilisées dans la synthèse de 2021 (de Foucault, 2021a). Elle relève clairement des Rubio tenuifoliae-Pistacietalia terebinthi B. Foucault 2021.
Globalement les colonnes Ia à VIIa (en moyenne 375 m) correspondent à une alliance qui serait caractérisée par Prunus webbii, Calicotome villosa, Styrax officinalis, Rubus sanctus.
Les colonnes Va à VIc (en moyenne 455 m) rassemblent des fourrés acidiphiles vicariants crétois des Cytiso villosi-Genistetalia monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 ; elles viennent combler un manque qui avait été souligné dans la synthèse de 2021 : « L’équivalent des Cytiso villosi-Genistetalia monspessulanae en Méditerranée orientale paraît plutôt mal connu » (de Foucault, 2021a : 31). Ces fourrés partagent Erica arborea, Cytisus creticus, Arbutus unedo, plus rarement A. andrachne. On peut au moins définir le Cytiso cretici-Ericetum arboreae ass. nov. hoc loco (colonnes Va, VIa et VIb), typus nominis hoc loco : composante arbustive du relevé 4 du tableau VI in Barbero & Quézel (1980, Ecol. Medit. V : 195). Cette association constitue le typus nominis hoc loco du Cytision cretici all. nov. hoc loco, alliance caractérisée par la combinaison de Erica arborea, Cytisus creticus, Arbutus unedo, A. andrachne, et la rareté ou l’absence de nombreux taxons caractérisant ou différenciant l’éventuelle alliance Ia à VIIa (voir tableau 5).
Enfin les colonnes VIIb à VIIIb (en moyenne 1 290 m) rassemblent des fourrés oro-crétois à Berberis cretica, Rhamnus saxatilis subsp. prunifolia, Rosa pulverulenta, plus rarement Zelkova abelicea ; ils relèvent de l’Asparago acutifolii-Berberidion creticae B. Foucault 2021 (de Foucault, 2021a) typifié par l’Aceri sempervirentis-Berberidetum creticae Barbero & Quézel 1980 (Ecol. Medit. V : 202). Relève aussi de cette alliance le Rhamno prunifoliae-Berberidetum creticae Zaffran & B. Foucault in B. Foucault 2021 (de Foucault, 2021c).
* La végétation herbacée intraforestière de Grèce continentale
Le tableau 6 synthétise les données sur la végétation herbacée vivace intraforestière continentale apportées par Barbero & Quézel (1976, syntaxons en BQ), puis Gamisans & Hébrard (1979, syntaxons en GH). La flore est ici encore plus diversifiée et bien plus complexe que pour la végétation arborescente ou arbustive ; on se contentera donc à nouveau de quelques indications et de comparaisons en lien avec une synthèse antérieure (de Foucault, 2020).
Laissant de côté l’ensemble BQ1a à BQ3, différencié par des taxons issus de phryganes des Cisto-Micromerietea julianae (à une altitude de 230 m, sauf l’élément BQ3 à 875 m), Asplenium onopteris, Carex distachya, Ruscus aculeatus… oriente le placement de l’ensemble BQ4a+4b à BQ8b (altitude moyenne de 490 m) dans les Rubio peregrinae-Asplenietea onopteridis B. Foucault 2020, ne renfermant pour l’instant qu’un seul ordre, les Rubio peregrinae-Asplenietalia onopteridis B. Foucault 2020, et quatre alliances ouest- à centre-méditerranéennes. Il paraît prématuré de définir les subdivisions de cette classe pour la Grèce.
On a eu l’occasion d’observer une telle végétation en mai 2022 :
- à Makrinitsa (nord de Vólos, Pélion, bord de la route 4303 ; N 39° 24’ 00,7’’, E 22° 59’ 50’’, 591 m, le 15 mai 2022) : Asplenium onopteris, Campanula patula, Symphytum bulbosum, Lathyrus laxiflorus (photo 7), Buglossoides purpureocaerulea, Cyclamen hederifolium (photo 8), Melica uniflora, Primula vulgaris s. l., Luzula forsteri, Anemone pavonina (photo 9), Arum italicum, Dioscorea communis, Brachypodium sylvaticum, Rubia peregrina, Carex flacca… ;
- dans les Météores, à Kastraki, près du monastère Aghia Trias Asketirion (N 39° 42’ 42,3’’, E 21° 37’ 16,7’’, 320 m, le 22 mai 2022), sous une canopée de Fraxinus ornus, Carpinus orientalis (photo 10), Cercis siliquastrum, Acer monspessulanum/sempervirens : Ruscus aculeatus, Lathyrus laxiflorus, Asplenium onopteris, Dioscorea communis, Carex distachya, depauperata, Cyclamen hederifolium, Hedera helix, Luzula forsteri, Melica uniflora… ;
- dans les Météores, à Kalambaka, sud du Large Boulder (N 39° 42’ 49,8’’, E 21° 37’ 58,7’’, 388 m, le 24 mai 2022, 60 %, 30 m², 8 taxons) : Ruscus aculeatus 3, Asplenium onopteris 2, Carex distachya 2, Asparagus aphyllus 1, Cyclamen hederifolium +, Symphytum bulbosum +, Fraxinus ornus *o. j 1, Quercus coccifera j 1 ;
- dans les Météores, à Kalambaka, sud-ouest d’Aghia Triada (N 39° 42’ 52,8’’, E 21° 38’ 12,4’’, 360 m, le 24 mai 2022, 70 %, 40 m², 14 taxons) : Asplenium onopteris 3, Ruscus aculeatus 3, Symphytum bulbosum 2, Umbilicus rupestris 1, Luzula forsteri 1, Cyclamen hederifolium 1, Carex distachya +, Teucrium divaricatum +, Pseudoturritis turrita +, Fraxinus ornus *o. j 1, Quercus j 1, Q. coccifera j 1, Carpinus orientalis j 1, Clematis flammula j +.




Sur la base des trois derniers relevés/liste, l’association des Météores mérite d’être distinguée sous le nom de Cyclamini hederifolii-Ruscetum aculeati ass. nov. hoc loco, typus nominis hoc loco : relevé 4 hoc loco des Météores ci-dessus (photo 11).

Avec Aremonia agrimonioides, Anemone blanda, Symphytum bulbosum (photo 12), Potentilla micrantha (photo 13), Poa nemoralis, Campanula trachelium, Melica uniflora, l’ensemble BQ9a à BQ25, soit les deux tiers du tableau 6, peut être rapproché des Anemono nemorosae-Caricetea sylvaticae et sans doute des Cyclamini hederifolii-Lathyretalia veneti B. Foucault 2020.


Il se divise en un sous-ensemble BQ9a à BQ10b différencié des suivants par l’absence d’un grand nombre de taxons, dont beaucoup de taxons caractéristiques de classe : Cephalanthera damasonium, C. rubra, Viola gr. riviniana, Neottia nidus-avis, Allium ursinum, Epilobium montanum, Festuca heterophylla, Polystichum aculeatum, Epipactis helleborine, Lathyrus venetus, Sanicula europaea, Primula vulgaris, Lactuca muralis, Limodorum abortivum, Myosotis sylvatica subsp. cyanea, Daphne laureola, Clinopodium grandiflorum.
Le sous-ensemble GH1 à GH11b est remarquablement homogène et bien caractérisé. On peut au moins distinguer :
- le Veronico vindobonensis-Aremonietum agrimonioidis ass. nov. hoc loco (colonne GH5, autour de 1 200 m), typus nominis hoc loco: composante herbacée vivace du relevé 14 du tableau 5 in Gamisans & Hébrard (1979, Phytosoc., NS, IV hors texte) ;
- le Campanulo spatulatae-Lathyretum laxiflori ass. nov. hoc loco (colonne GH7, autour de 750 m), typus nominis hoc loco: composante herbacée vivace du relevé 8 du tableau 7 in Gamisans & Hébrard (1979, Phytosoc., NS, IV hors texte).
Ce sous-ensemble de sous-bois herbacés montagnards du nord-ouest de la Grèce continentale (Épire et Macédoine sud-occidentale) mérite clairement le rang d’alliance : Veronico vindobonensis-Galion laconici all. nov. hoc loco, caractérisée par Galium sylvaticum subsp. laconicum, G. lucidum, Veronica chamaedrys subsp. vindobonensis (ici en limite sud-orientale de son aire plutôt centre-européenne), Campanula spatulata, Ptilostemon strictus, Trifolium patulum, Knautia drymeia subsp. nympharum, Hypericum spruneri, Selinum silaifolium, Lembotropis nigricans, Crepis viscidula, Vogtia microphylla, typus nominis hoc loco : Campanulo spatulatae-Lathyretum laxiflori nov.
Le large sous-ensemble BQ12 à BQ24b (altitude moyenne 1 400 m) est très original et statistiquement bien caractérisé. On peut au moins distinguer
- le Campanulo abietinae-Aremonietum agrimonioidis ass. nov. hoc loco (colonne BQ15), typus nominis hoc loco: composante herbacée vivace du relevé 5 du tableau 15 in Barbero & Quézel (1976, Medit. II : 42) ;
- le Bornmuellero emarginatae-Aremonietum agrimonioidis ass. nov. hoc loco (colonne BQ20b), typus nominis hoc loco: composante herbacée vivace du relevé 7 du tableau 20 in Barbero & Quézel (1976, Medit. II : 53) en remplaçant Petteria emarginata par Bornmuellera emarginata ;
- le Cyclamini repandi-Daphnetum laureolae ass. nov. hoc loco (colonnes BQ21a et BQ21b), typus nominis hoc loco: composante herbacée vivace du relevé 8 du tableau 21 in Barbero & Quézel (1976, Medit. II : 58) ;
- l’Aremonio agrimonioidis-Campanuletum pichleri ass. nov. hoc loco (colonne BQ23), typus nominis hoc loco: composante herbacée vivace du relevé 2 du tableau 23 in Barbero & Quézel (1976, Medit. II : 62), en remplaçant le nom Campanula trichocalycina par C. pichleri, correspondant à une « hêtraie à fougères » (Athyrium filix-femina, Polystichum aculeatum) et à l’association arborescente de l’Abieti borisii-regis-Fagetum sylvaticae nov. (cf. supra).
Tout ce sous-ensemble correspond à une alliance originale oro-balkanique (altitude moyenne 975 m) très richement caractérisée par Cephalanthera damasonium, C. rubra, Neottia nidus-avis, Lathyrus grandiflorus (photo 14), L. laxiflorus (photo 7), L. venetus, Polystichum aculeatum, Epipactis helleborine, Epilobium montanum, Sanicula europaea, Primula vulgaris, Limodorum abortivum, Ferulago sylvatica, Daphne laureola, Clinopodium grandiflorum, Daphne oleoides, Arum italicum, Digitalis lanata, D. ferruginea, Geocaryum cynapioides, Helleborus cyclophyllus, Campanula sparsa, Doronicum orientale (photo 15), Trifolium pignantii, Elymus panormitanus, Anemone blanda, Symphytum bulbosum, Aremonia agrimonioides, Potentilla micrantha, Phlomis samia, Physospermum cornubiense… (voir aussi le tableau 6), l’Aremonio agrimonioidis-Doronicion orientalis all. nov. hoc loco, typus nominis hoc loco : Campanulo abietinae-Aremonietum agrimonioidis nov.


* La végétation herbacée intraforestière de Crète
Le tableau 7 synthétise les données sur la végétation herbacée vivace intraforestière crétoise apportées par Barbero & Quézel (1980).
L’ensemble Ia à VIc se différencie par Cistus creticus, Origanum onites, Pulicaria odora, Ruscus aculeatus, Teucrium divaricatum… et se sépare en
- un sous-ensemble Ia à Vb (altitude moyenne 355 m), très original et richement différencié par Phlomis lanata, Scutellaria sieberi, Thymbra capitata, Galium fruticosum, Piptatherum caerulescens, Iris unguicularis cretensis, Carex distachya, Phagnalon graecum… ;
- un sous-ensemble VIa à VIc (en moyenne 500 m) à Oenanthe pimpinelloides, Pteridium aquilinum, Clinopodium vulgare, plutôt à valeur d’ourlet.
L’ensemble VIIa à VIIIb est plus orophile (en moyenne 1 164 m) et se différencie par Crepis fraasii, Cirsium morinifolium, Helictochloa agropyroides, Dactylis glomerata subsp. rigida, Phlomis cretica, Melica rectiflora ; on peut au moins distinguer le Crepido fraasii-Bunietum ferulacei ass. nov. hoc loco, typus nominis hoc loco : composante herbacée vivace du relevé 8 du tableau VII in Barbero & Quézel (1980, Ecol. Medit. V : 198), association pouvant tenir lieu de typus nominis hoc loco du Phlomido creticae-Crepidion fraasii all. nov. hoc loco, alliance de végétation herbacée vivace intraforestière oro-crétoise caractérisée par Crepis fraasii, Cirsium morinifolium, Helictochloa agropyroides, Dactylis glomerata subsp. rigida, Phlomis cretica, Melica rectiflora.
Cette végétation originale ne paraît se rattacher ni aux Anemono-Caricetea sylvaticae ni aux Rubio-Asplenietea onopteridis.
2. Les fourrés héliophiles
2.1. Quelques fourrés de corniche selon Quézel (1964)
Dans la publication de Quézel (1964), dans certains tableaux de végétation chasmophytique, on peut remarquer des éléments de végétation arbustive rupicole. Ainsi, dans le Campanuletum aizoo-rupicolae Quézel 1964 (Quézel, 1964, tableau 1), les relevés 5, 10, 12, 14 et 15 permettent la définition d’un tel fourré : Rhamnus sibthorpiana (incl. Rh. guicciardii) V, Cotoneaster tomentosus III, Daphne oleoides II, Rosa pulverulenta II, Rhamnus saxatilis *prunifolia I, Frangula rupestris I, Sorbus aria j I, que l’on peut dénommer Cotoneastro tomentosi-Rhamnetum sibthorpianae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 15 du tableau 1 in Quézel (1964, Vegetatio XII (5-6) hors texte) ; il rappelle les fourrés de l’Amelanchiero ovalis-Buxion sempervirentis dont d’ailleurs Cotoneaster tomentosus est l’un des taxons caractéristiques (de Foucault & Royer, 2016).
Dans les tableaux 15 à 19 (resp. sub Onosmo leptanthi-Scabiosetum taygeteae Quézel 1964, Ribeso uvae-crispae-Galietum lucidi Quézel 1964, Sideritidetum theezantis Quézel 1964, Cirsio cyllenei-Astragaletum cyllenei Quézel 1964, Astragalo rumelici-Marrubietum velutini Quézel 1964, Rindero graecae-Acantholimonetum echini Quézel 1964) et 21 à 23 (resp. sub Astragalo lactei-Convolvuletum cochlearis Quézel 1964, Erysimo parnassi-Minuartietum stellatae Quézel 1964, Thymo hirsuti-Paronychietum chionaeae Quézel 1964), soit 61 relevés, on peut définir le fourré suivant :
- Daphne oleoides IV, Juniperus communis *hemisphaerica III, Prunus prostrata I, Crataegus pycnoloba +, Berberis cretica +, Ribes uva-crispa +, Prunus cocomilia r, Hippocrepis emerus *emeroides r,
- autres taxons : Juniperus foetidissima j +, Abies cephalonica j r,
Junipero hemisphaericae-Daphnetum oleoidis ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 2 du second syntaxon élémentaire du tableau 15 in Quézel (1964, Vegetatio XII (5-6) hors texte).
2.2. Quelques fourrés observés en 2022
Parmi les données plus récentes, on peut déjà évoquer cette végétation relevée sur un coteau rocailleux et fortement exposé à la chaleur dans le Pélion, au nord de Vólos :
Makrinitsa, coteau en bord de route 4303, N 39° 23’ 59,6’’, E 22° 59’ 39,7’’, 631 m, 15 mai 2022, 80 %, 100 m², 13 taxons, photo 16
Rhus coriaria 3, Phlomis fruticosa 2, Spartium junceum 2, Clematis cf. vitalba (non fleuri) 2, Pistacia terebinthus 2, Euphorbia characias *wulfenii 1, Asparagus aphyllus 1, Rhamnus alaternus *a. 1, Rh. lycioides *oleoides +, Hippocrepis emerus *emeroides +, Anagyris foetida (photo 17) +, Rubia tenuifolia (photo 18) +, Quercus coccifera j 1, Platanus orientalis j +.
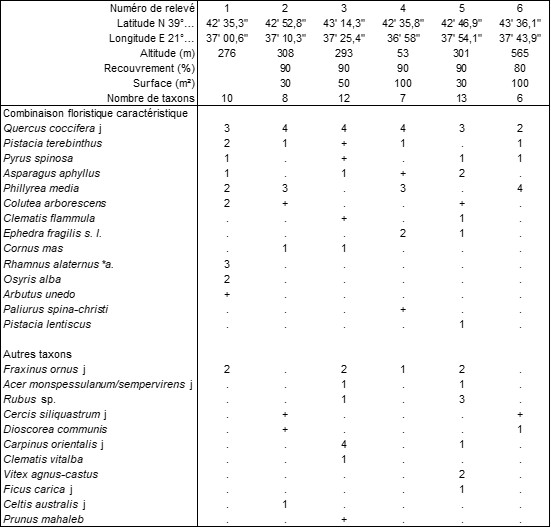
Annexe du tableau 8 (Météores) – rel. 1 : Kastraki, face au Dellas Boutique Hotel, 22 mai 2022 ; rel. 2 : au-dessus de Kastraki, sous “Marmaro”, 23 mai 2022 ; rel. 3 : Kastraki, nord d’Aghion Previna, 22 mai 2022 ; rel. 4 : id. rel. 1, 23 mai 2022 ; rel. 5 : Kalambaka, sud de St. Peter in Chains, 24 mai 2022 ; rel. 6 : Kalambaka, vers le Grand Météore, 25 mai 2022.



Dans les Météores, un fourré xérophile à Quercus coccifera tend à coloniser la base des cheminées de grès coiffées par des monastères (photo 19) ; le tableau 8 en rapporte six relevés. Au chêne kermès, sont souvent associés Pistacia terebinthus, Pyrus spinosa, Asparagus aphyllus, Phillyrea media, Colutea arborescens, Clematis flammula, Ephedra fragilis s. l., Cornus mas, flore renvoyant à nouveau aux Pistacio-Rhamnetea alaterni et aux Rubio-Pistacietalia terebinthi ; la trop forte variabilité sur une aire finalement assez réduite ne permet cependant pas d’envisager une association pour l’instant.

À Flórina (Macédoine sud-occidentale), en marge de pinèdes à Fraxinus ornus, à une altitude moyenne de 770 m, se développe un fourré dont le tableau 9 rapporte six relevés accueillant en moyenne environ huit taxons par relevé. Cette végétation est surtout dominée par Juniperus communis subsp. c. et Rosa canina, accompagnés de taxons juvéniles d’essences potentiellement arborescentes, Fraxinus ornus, Acer tataricum (photo 20), Quercus frainetto : Fraxino orni-Juniperetum communis ass. nov. hoc loco (photo 21), typus nominis hoc loco : relevé 5 du tableau 9 hoc loco. On notera la présence de Syringa vulgaris (photo 22), ici en situation spontanée.



À Nafpaktos, dans le fond des cours d’eau temporaire, on peut observer un fourré hygrophile se rattachant aux Nerio oleandri-Viticetalia agni-casti (Nerio oleandri-Tamaricetea africanae) :
Nafpaktos, bord de la route 0127, nord du Ktel Bus Terminal, N 38° 24’ 23,4’’, E 21° 50’ 28,8’’, 64 m, 30 mai 2022, 70 %, 30 m², 5 taxons
Vitex agnus-castus 3, Nerium oleander 2, Phlomis fruticosa 2, Ficus carica j, Platanus orientalis j 1,
nouveau représentant du Platano orientalis-Viticetum agni-casti B. Foucault 1993 ex 2009 (photo 23). Cette association avait été définie de manière invalide dans une publication antérieure consacrée à la Crète (de Foucault, 1993) sous le nom de Myrto communis-Viticetum agni-casti.
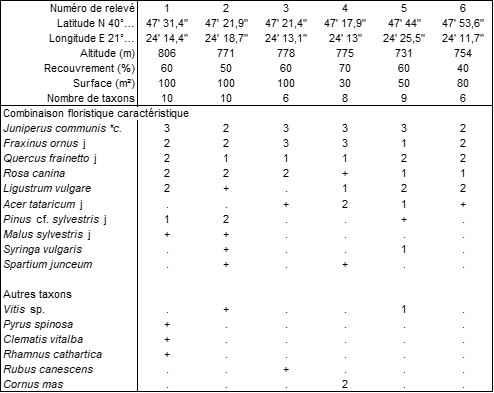
Annexe du tableau 9 (au-dessus de Flórina, Macédoine sud-occidentale) – rel. 1 à 4 : au-dessus de l’hôtel Philippion, 21 mai 2022 ; rel. 5 et 6 : au-dessus de l’hôtel Phaidon & Spa, 21 mai 2022.

Au mont Parnitha, dans l’arrière-pays d’Athènes, au-dessus d’une belle phrygane d’altitude à Cerastium candidissimum, Thymelaea tartonraira, Euphorbia myrsinites, Hypericum empetrifolium, Scutellaria rubicunda, Sideritis scardica, S. syriaca… (photos 24 à 28), avec la thérophyte Brizochloa humilis (proche des Briza, mais à épillets non pendants et lemmes à base atténuée, non cordée) sur sol rocailleux, se développe un fourré correspondant au relevé suivant :
mont Parnitha, non loin du Regency Casino, N 39° 09’ 09,2’’, E 23° 41’ 12,8’’, 1 074 m, 01 juin 2022, 75 %, 200 m², 16 taxons
- arbustes caractérisques : Quercus coccifera 4, Crataegus pycnoloba (photo 29) 2, Juniperus oxycedrus *o. 2, Rosa 1, Pistacia terebinthus 1, Phillyrea media 1, Berberis cretica (photo 30) +, Asparagus aphyllus +, Rhus coriaria +,
- autres taxons : Abies cephalonica j 2, Pinus halepensis j 1, Sorbus umbellata j (photo 31) 1, Cistus creticus ! +, Quercus ilex j +, Fraxinus ornus *o. j +, Cercis siliquastrum j +,
typus nominis hoc loco du Crataego pycnolobae-Quercetum cocciferae ass. nov. hoc loco (photo 32).









La redescente du mont Parnitha vers Thrakomakedones a donné lieu à quelques observations sur une phrygane de plus basse altitude, avec Fumana thymifolia, Anthyllis hermanniae (subsp. h. selon Brullo & Giusso del Galdo, 2006 ; photo 33), Cistus creticus (photo 34), Ptilostemon chamaepeuce (photo 35), Campanula cf. anchusifolia, Satureja thymbra, Hypericum triquetrifolium (photo 36), H. empetrifolium, Teucrium divaricatum (photo 37), Thymbra capitata (photo 38), Sarcopoterium spinosum (photo 39).







3. Les lisières herbacées
3.1. Les lisières herbacées vivaces
Les lisières herbacées, ou ourlets, vivaces ont été peu observées. La plus originale est celle qui a été parcourue à Flórina, vers 700 m d’altitude, avec Daphne blagayana (photo 40), Achillea coarctata, Vincetoxicum cf. fuscatum (photo 41), Silene italica, Hypericum barbatum (photo 42), Fritillaria graeca, Trifolium alpestre, Pilosella cymosa, Bupleurum falcatum, Euphorbia cyparissias, Ranunculus psilostachys (photo 43), Cytisus hirsutus, Geranium sanguineum, Potentilla micrantha, Lathyrus niger, Brachypodium sylvaticum, Genista carinalis (photo 44), Clinopodium vulgare…, qui peut sans problème se rattacher aux Trifolio medii-Geranietea sanguinei, sans qu’il soit possible de préciser les unités de rang inférieur.





3.2. Les lisières herbacées thérophytiques
Il existe aussi des lisières herbacées thérophytiques à développement optimum vernal, caractérisées par Cardamine hirsuta, C. graeca (photo 45), Theligonum cynocrambe, Sedum cepaea, Veronica cymbalaria, Geranium lucidum et G. purpureum, Arabidopsis thaliana, Anisantha sterilis, des petits Campanula malheureusement restés indéterminés (photo 46), Rhagadiolus edulis (photo 47), parfois Cruciata pedemontana (photo 48)… L’une d’elles a été décrite par Oberdorfer (1953-54, tableau 25) sous le nom de Geranio lucidi-Sedetum cepaeae, présent en France sous une race dépourvue de Cardamine graeca (de Foucault, 1988, 2009). Cette association a notamment été observée à Flórina ; Horvat et al. (1974 : 187) la reprennent aussi dans leur synthèse phytosociologique.




Une seconde, inédite, est caractérisée par Chaerophyllum nodosum ; le tableau 10 en rapporte six relevés. Il décrit une végétation assez distincte du Chaerophyllo nodosi-Alliarietum petiolatae Rivas Mart. & Mayor in Fuentes 1986 ouest-européen (de la Fuentes, 1985 publié 1986 ; de Foucault, 2017), car elle possède une tonalité méditerranéenne plus affirmée. Elle a été observée dans les Météores, à une altitude moyenne de 395 m, elle est développée sur sol assez profond et accueille en moyenne neuf thérophytes par relevé. On peut la décrire sous le nom de Rhagadiolo edulis-Chaerophylletum nodosi ass. nov. hoc loco, typus nominis hoc loco : relevé 5 du tableau 10 hoc loco (photo 49).

4. Quelques associations orophiles d’après Quézel (1964)
En parcourant les tableaux sur la végétation des hautes montagnes de Grèce publiés par Quézel (1964), on peut mettre en évidence quelques associations particulières. Tout d’abord les relevés 2 et 6 du tableau 15 de cet auteur (sub Onosmo leptanthi-Scabiosetum taygeteae Quézel 1964), les tableaux 18 (Astragalo rumelici-Marrubietum velutini Quézel 1964), 19 (Rindero graecae-Acantholimonetum echini Quézel 1964), 21 (Astragalo lactei-Convolvuletum cochlearis Quézel 1964), 28 (Plantagini capitatae-Trifolietum alpestris Quézel 1964) et les relevés 5 et 8 du tableau 30 (Nepeto nudae-Astragaletum cephalonici Quézel 1964) offrent une documentation de 25 relevés sur l’association thérophytique orophile suivante : Cerastium brachypetalum *tauricum IV, Cynosurus echinatus II, Arenaria serpyllifolia II, Veronica arvensis I, Filago arvensis I, Saxifraga tridactylites +, Malcolmia graeca *bicolor +, Draba verna r, soit l’Arenario serpyllifoliae-Cerastietum taurici ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 5 du tableau 30 in Quézel (1964, Vegetatio XII (5-6) hors texte).
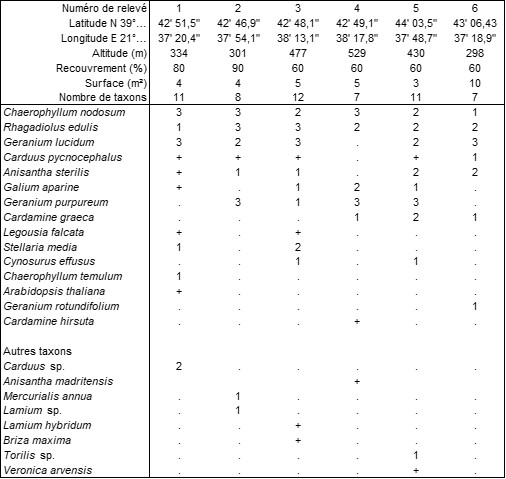
Annexe du tableau 10 (Météores) – rel. 1 : au-dessus de Kastraki, sous “Pyxari”, 23 mai 2022 ; rel. 2 : Kalambaka, sud de St. Peter in Chains, 24 mai 2022 ; rel. 3 : Kastraki, au pied d’Aghia Triada, 24 mai 2022 ; rel. 4 : id. rel. 3, est d’Aghia Triada, 24 mai 2022 ; rel. 5 : Kastraki, sous Ypapanti et Varlaam, 25 mai 2022 ; rel. 6 : Kastraki, sous Agios Georgios Mandilas, 25 mai 2022.
D’autre part, les huit relevés du tableau 16 (sub Sideritidetum theezantis Quézel 1964) permettent de décrire la communautés suivante : Viola parvula IV, Veronica verna IV, Malcolmia graeca *bicolor IV, décrite comme une autre association thérophytique orophile, le Violo parvulae-Malcolmietum bicoloris ass. nov. hoc loco, typus nominis hoc loco : composante thérophytique du relevé 5 du tableau 16 in Quézel (1964, Vegetatio XII (5-6) hors texte), que l’on peut rapprocher de l’alliance elle-même orophile du Sedion annui B. Foucault 2019 dont V. parvula est caractéristique (de Foucault, 2019).
En p. 310 de cette même publication, le Pinguiculo hirtiflorae-Aquilegietum amaliae Quézel 1964 est clairement une association des Adiantetea capilli-veneris, association omise dans la synthèse de cette classe (de Foucault, 2015a). Tant par la grassette que par l’ancolie, on peut remarquer la vicariance de cette végétation de Méditerranée orientale avec celle du Pinguiculion longifoliae Fern. Casas 1970 (écrit par erreur Schoeno-Caricion brachystachyos dans le tableau 1 de la synthèse de 2015), mais on ne peut la rattacher à cette unité ouest-européenne. En revanche, si on en rapproche l’Adianto capilli-veneris-Pinguiculetum hirtiflorae dont l’autorité apparaît soit comme Krause et al. 1963 corr. Deil 1989, soit comme Stevanović & Bulić 1989 selon les sources (on n’a pas encore eu accès aux données synfloristiques à son propos ; Krause et al., 1963 : 376), on peut définir une alliance est-méditerranéenne vicariante, l’Adianto capilli-veneris–Pinguiculion hirtiflorae all. nov. hoc loco, notamment caractérisée par Pinguicula hirtiflora, typus nominis hoc loco : Pinguiculo hirtiflorae-Aquilegietum amaliae Quézel 1964 (Vegetatio XII (5-6) : 320) ; il paraît utile d’ajouter A. capillus-veneris à P. hirtiflora pour nommer cette alliance afin de distinguer cette végétation des bas-marais des Scheuchzerio-Caricetea nigrae tels que le Soldanello pindicolae-Pinguiculetum hirtiflorae Quézel 1967 (Quézel, 1967 : 208 ; Horvat et al., 1974 : 654). Remarquons que cette situation est isomorphe à celle de Pinguicula grandiflora dans les Pyrénées audoises et le nord-ouest de la péninsule Ibérique, qui participe tant à l’Adianto capilli-veneris-Pinguiculetum grandiflorae B. Foucault 2019 qu’au Carici mairei-Pinguiculetum grandiflorae B. Foucault 2019 ou au Pinguiculo grandiflorae-Caricetum lepidocarpae Rivas Mart. et al. 1984, ces deux derniers rattachés au Caricion davallianae (Rivas-Martínez et al., 1984 ; de Foucault, 2019).
À propos des Adiantetea capilli-veneris, mais hors Grèce, signalons une association ibérique décrite postérieurement à la synthèse de 2015 et relevant bien du Pinguiculion longifoliae, l’Adianto capilli-veneris-Pinguiculetum saetabensis M.B. Crespo, M.Á. Alonso, J.E. Oltra & Mart.-Azorín 2019 (Crespo et al., 2019).
5. La végétation liée aux activités humaines
5.1. Les fourrés rudéraux
Dans beaucoup de régions du monde, au voisinage des activités humaines, il existe souvent des fourrés de recolonisation des zones délaissées, abandonnées…, par exemple le Fraxino excelsioris-Sambucetum nigrae B. Foucault 1991, le Ribeso uvae-crispae-Sambucetum racemosae B. Foucault 1991 pour la France, le Nicotiano glaucae-Ricinetum communis B. Foucault 2013 au Maghreb, les Calotropido procerae-Ricinetum communis B. Foucault 2020 et Calotropido giganteae-Ricinetum communis B. Foucault in B. Foucault & J.-M. Royer 2016 tropicaux, le Leucaeno leucocephalae-Broussonietum papyriferae B. Foucault 2020 à Taiwan, le Calliandro calothyrsi-Sambucetum javanicae B. Foucault in B. Foucault & J.-M. Royer 2016 d’Indonésie… Le Fico caricae-Sambucetum nigrae B. Foucault 2015, décrit du midi de la France (de Foucault, 2015b), se retrouve bien dans les collines et basses montagnes de Grèce, par exemple à Makrinitsa et dans les Météores (photo 50).

En dehors de cette situation, il existe des fourrés plus thermophiles (photo 51), mais ils combinent essentiellement des formes juvéniles d’espèces potentiellement arborescentes : Ailanthus altissima, Celtis australis, Ficus carica, Acer negundo, Robinia pseudacacia, rarement Nicotiana glauca, de sorte qu’il paraît difficile de décrire le fourré correspondant, qui s’avère n’être qu’une forme juvénile d’une forêt plus évoluée dominée par l’Ailanthe.

5.2. La végétation anthropophile des murs
Les villes historiques de Grèce continentale sont souvent bien pourvues en murs anciens, ruines, remparts… qui sont colonisés par des chasmophytes anthropophiles, dont Parietaria judaica qui permet de rattacher cette végétation aux Parietarietea judaicae (de Foucault, 2014).
Ainsi les remparts de Thessalonique et de Nafpaktos sont colonisés par une association à Capparis spinosa correspondant au Capparietum rupestris O. Bolòs & Molin. 1958, muté ici en Capparietum spinosae. Le câprier éponyme y forme souvent de longues guirlandes pendantes (photo 52). Le relevé réalisé au port de Nafpaktos
(N 38° 23’ 30,8’’, E 21° 49’ 43,3’’, 29 mai 2022, 8 taxons) a donné : Cymbalaria muralis 3, Parietaria judaica 2, Reichardia picroides 2, Allosorus guanchicus 2, Capparis spinosa 1, Phagnalon saxatile 1, Umbilicus horizontalis 1, Ficus carica j +.

Plus originale est la végétation observée à plusieurs endroits de Ioánnina, capitale de l’Épire ; le tableau 11 en rapporte neuf relevés, qui partagent, outre P. judaica, Campanula versicolor (photo 53), Hedera helix, de jeunes Ficus carica. En outre, dans la forme typique, plus thermophile, on a relevé Umbilicus horizontalis (photo 54), Alyssum saxatile (photo 55), Micromeria juliana (photo 56). On peut la décrire sous le nom de Campanulo versicoloris-Parietarietum judaicae ass. nov. hoc loco, typus nominis hoc loco : relevé 9 du tableau 11 hoc loco (photo 57). Le relevé 1 décrit une variation exposée au nord accueillant un taxon souvent inféodé aux sous-bois herbacés à luminosité atténuée, Umbilicus erectus (photo 58). Cette association a été observée dans d’autres villes de Grèce continentale, mais la Campanule éponyme n’a été déterminée que tardivement, lors de sa floraison. Selon Horvat et al. (1974 : 552), des stations primaires de C. versicolor seraient des falaises rocheuses à Onosma frutescens dont la végétation relèverait du Campanulion versicoloris et des Onosmetalia frutescentis.






Il est particulièrement intéressant de rapprocher cette présence de Campanula versicolor en position secondaire sur des murs, ou parfois aussi sur des roches en place mais alors en contexte anthropisé (relevé 7 du tableau 11), de la présence sur murs d’autres Campanulaceae comme Trachelium caeruleum dans le Cymbalario muralis-Trachelietum caerulei Rivas Mart. 1969 (photo 59) ou Petromarula pinnata sur les murs de certaines villes de Crète (observation personnelle en 1991, non évoquée dans le compte-rendu ultérieur ; de Foucault, 1993).
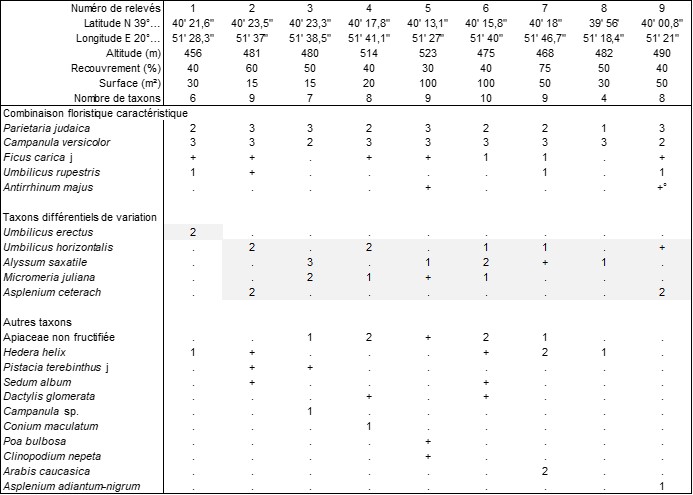
Annexe du tableau 11 (Ioánnina, les 27 et 28 mai 2022) – rel. 1 : rempart nord-ouest du Kastro, vers l’embarcadère ; rel. 2 : mur de la citadelle d’Aslan Pacha ; rel. 3 : mur de la tour byzantine ; rel. 4 : rempart au bord du Vissarionos Makri, vers Its Kale ; rel. 5 : mur à Plateia, le long de Konstantinou Karamanli ; rel. 6 : vers Ekthese Argyrotechnias ; rel. 7 : sur la roche en place près de Dionisiou Filosophou ; rel. 8 : mur du parking est de Plateia Kologiou ; rel. 9 : nord du musée archéologique, dans le parc Litharitsia.

5.3. Une végétation messicole
Non loin de Véroia, à Vergina, vers le carrefour route 5305 × route de Vergina, un champ de céréales localisé
N 40° 29’ 35,5’’, E 22° 18’ 54,9’’, 83 m a donné lieu au relevé suivant :
Papaver rhoeas 3, Hypecoum imberbe 2 (photo 60), Bunias erucago +, Delphinium consolida (photo 61) +, Glebionis coronaria 1, Buglossoides arvensis 1, Viola gr. arvensis +, Anthemis cotula 2, Scandix pecten-veneris 1, Medicago sp. 1, Misopates orontium 1, Lolium rigidum 2, Rapistrum rugosum +, Bifora testiculata + (photo 62),
qui peut se rattacher à l’alliance méditerranéenne du Roemerion hybridae.



5.4. Les friches nitrophiles
Séduit par la flore plutôt naturelle, au contraire d’Oberdorfer (1953-54), on a peu regardé la végétation herbacée nitrophile, souvent riche en « chardons » divers. Signalons simplement la friche observée au-dessus du lit de rivière asséché colonisé par le Platano-Viticetum agni-casti (photo 23), friche à Dittrichia viscosa, Hyparrhenia sinaica, Oloptum miliaceum, Verbascum sinuatum… accompagnés de taxons des phryganes comme Stachys cretica (photo 63), Genista acanthoclada (photo 64), Sarcopoterium spinosum, Teucrium gr. polium…, ces derniers à considérer comme témoins relictuels d’une phrygane initiale plus naturelle ici dégradée par les influences humaines (photo 65). Cette végétation peut se rattacher à l’Hyparrhenion hirtae Braun-Blanq., P. Silva & Rozeira 1956 (de Foucault & Jasprica, 2019).



6. Conclusion
Cet apport à la connaissance de la végétation grecque a donc permis de reconnaître trente-cinq associations végétales, dix alliances nouvelles (Cercidi siliquastri-Quercion cocciferae B. Foucault, Abietion cephalonico-borisii-regis B. Foucault, Pinion pallasianae B. Foucault, Abieti borisii-regis-Fagion sylvaticae B. Foucault, Hippocrepido emeroidis-Clematidion vitalbae B. Foucault, Cytision cretici B. Foucault, Aremonio agrimonioidis-Doronicion orientalis B. Foucault, Veronico vindobonensis-Galion laconici B. Foucault, Phlomido creticae-Crepidion fraasii B. Foucault, Adianto capilli-veneris–Pinguiculion hirtiflorae B. Foucault), ainsi que l’ordre des Abietetalia cephalonico-borisii-regis B. Foucault. Cet apport vient notamment compléter pour cette partie de l’Europe la synthèse précédente sur la végétation forestière européenne selon le paradigme synusial (de Foucault, 2020). Il reste à espérer que cette contribution pourra relancer les études phytosociologiques en Méditerranée orientale et tout particulièrement dans les Balkans.
Bibliographie
Barbero M. & Quézel P., 1976. Les groupements forestiers de Grèce centro-méridionale. Ecologia Mediterranea II : 3-86.
Barbero M. & Quézel P., 1980. La végétation forestière de Crète. Ecologia Mediterranea V : 175-210.
Bolòs O. (de) & Molinier R., 1958. Recherches phytosociologiques dans l’île de Majorque. Collectanea Botanica (Barcelona) 5 (3) : 699-865.
Brullo S. & Giusso del Galdo G., 2006. Taxonomic remarks on the Anthyllis hermanniae L. (Fabaceae, Faboideae) specis complex of the Mediterranean flora. Novon 16 : 364-314.
Crespo Villalba M.B., Alonso Vargas M.Á., Oltra Benavent J.E. & Martínez Azorín M., 2019. Adianto-Pinguiculetum saetabensis (Pinguiculion longifoliae): una nueva asociación rupícola setabense. Flora Montiberica 75 : 8-10.
Debazac E.-F. & Mavrommatis G., 1971. les grandes divisions écologique de la végétation forestière en Grèce continentale. Bulletin de la Société botanique de France 118 : 429-452.
Foucault B. (de), 1988. Contribution à la connaissance phytosociologique des corniches rocheuses de la vallée de l’Argenton, entre Argenton-Château et Massais (Deux-Sèvres). Bulletin de la Société botanique du Centre-Ouest, NS, 19 : 39-64.
Foucault B. (de), 1993. II – Données phytosociologiques. In B. de Foucault & J.-P. Cornu, Itinéraire botanique d’un groupe de Linnéens en Crète, 29 avril-13 mai 1991 (2e partie). Bulletin de la Société linnéenne Nord-Picardie 11 : 85-105.
Foucault B. (de), 2009. Contribution au prodrome des végétations de France : les Cardaminetea hirsutae Géhu 1999. Journal de botanique de la Société botanique de France 48 : 49-70.
Foucault B. (de), 2014. Contribution au prodrome des végétations de France : les Parietarietea judaicae Rivas-Mart. in Rivas Goday 1964. Acta Botanica Gallica: Botany Letters 161 (4) : 403-427.
Foucault B. (de), 2015a. Contribution au prodrome des végétations de France : les Adiantetea capilli-veneris Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952. Acta Bot. Gallica: Botany Letters 162 (4) : 375-403.
Foucault B. (de), 2015b. Quelques associations arbustives nouvelles pour la Malepère (Aude centrale). Bulletin de la Société botanique du Centre-Ouest, NS, 46 : 244-260.
Foucault B. (de), 2017. Quelques ourlets thérophytiques de l’Aude. Botanique 2 : 23-36.
Foucault B. (de), 2019. Données phytosociologiques collectées lors des sorties du groupe botanique de la SESA durant l’année 2018. Bulletin de la Société d’études scientifiques de l’Aude CXVIII : 176-186.
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2021a. Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens. Carnets botaniques 51 : 1-42 + 37 tableaux, https://doi.org/10.34971/D6BN-RG49.
Foucault B. (de), 2021b. Apports de la 155e session extraordinaire de la Société botanique de France à la connaissance de la végétation provençale. Journal de botanique de la Société botanique de France 93 : 62-86.
Foucault B. (de), 2021c. Miscellanées II : syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 70 : 1-22 + 13 tableaux, https://doi.org/10.34971/3t1m-6445.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Foucault B. (de) & Jasprica N., 2019. Sur une classe de végétation méconnue en France : les Dittrichietea viscosae Trinajstić, B. Foucault & Jasprica cl. nov. Botanique 5 : 155-170.
Fuentes V. (de la), 1985 publié 1986. Vegetación orófila del occidente de la provincia de Guadajara (Espana). Lazaroa 8 : 123-219.
Gamisans J. & Hébrard J.-P., 1979. À propos de la végétation des forêts d’Épire et de Macédoine grecque occidentale. Documents phytosociologiques, NS, IV : 289-341.
Guinochet M. & Drouineau G., 1944. Notes sur la végétation et le sol aux environs d’Antibes (Alpes-Maritimes). Recueil des travaux de l’Institut botanique de Montpellier I : 22-40.
Horvat I., Glavac V. & Ellenberg H., 1974. Vegetation Südosteuropas. Geobotanica Selecta IV : 1-767.
Krause W., Ludwig W. & Seidel F., 1963. Zur Kenntnis der Flora und Vegetation auf Serpentinstandorten des Balkans. 6 – Vegetationsstudien in der Umgebung von Mantoudi (Euböa). Botanisch Jahrbüscher 82 (4) : 337-403.
Molinier R., 1954. Les climax côtiers de la Méditerranée occidentale. Vegetatio IV : 284-308.
Oberdorfer E., 1953-54. Über Unkrautgesellschaften der Balkanhalbinsel. Vegetatio IV : 379-411
Quézel P., 1964. La végétation des hautes montagnes de la Grèce méridionale. Vegetatio XII (5-6) : 289-385.
Quézel P., 1967. La végétation des hauts sommets du Pinde et de l’Olympe de Thessalie. Vegetatio XIV : 127-228.
Rivas-Martínez S., Díaz T.E., Fernández Prieto J.A., Loidi J & Penas A., 1984. La vegetación de la alta montaña cantábrica: Los Picos de Europa. Ediciones Leonesas, León, 295 p.
Remerciements
Ils s’adressent à Pierre Coulot pour la vérification des déterminations de certaines Fabaceae.


