
L’utilisation des ligneux en phytosociologie synusiale
Title
The use of woody plants in synusial phytosociology
Résumé
Grâce à une définition précisée des phanérophytes et à l’utilisation de la suite de Fibonacci pour caractériser les hauteurs végétatives modales, il est possible de redéfinir le contenu floristique des classes phytosociologiques phanérophytiques présentes en France et dans les pays mitoyens.
Abstract
Following a new definition of phanerophytes, and taking into account the Fibonacci’s suite in order to characterize the vegetative heights of trees and shrubs, it is possible to redefine the phanerophyte flora of phytosociological classes which are present in France and neighbouring countries.
1. Introduction
La prise en compte des ligneux dans les relevés phytosociologiques synusiaux résulte de choix théoriques et techniques essentiels. Il est tout d’abord utile de rappeler les sept critères utilisables pour délimiter les synusies sur le terrain. Ce sont par ordre d’importance les suivants :
- homogénéité spécifique: une stabilité de la liste des taxons lors de l’exploration est nécessaire, en prenant en compte les écophènes : accomodats/écomorphoses et stades de développement (germination, juvénile, adulte). Cela nécessite aussi une réflexion sur l’aire minimale de chaque synusie relevée, surtout quand elles sont superposées, par exemple dans le cas des forêts, et sur le temps minimal de recherche des espèces dans chaque synusie, qui est souvent lié à leurs tailles ;
- homogénéité spatiale et temporelle: l’espace vital des taxons doit être similaire, en trois dimensions. Ainsi on ne mélangera pas des arbres de trente mètres et des herbes de quinze centimètres ; de même on ne complétera pas des relevés lors de saisons différentes. Chaque type de synusie possède son optimum saisonnier, qui guide le choix de la période de relevé ;
- homogénéité globale des stratégies de vie: cela nécessite en particulier de prendre en compte les types biologiques pour les plantes ou les types nutritionnels pour les animaux ;
- homogénéité phénologique: il conviendra de distinguer par exemple les phénophases vernales et estivales des végétations, ou les phases migratoires estivales ou hivernales pour les animaux ;
- homogénéité dynamique: elle peut être perçue en identifiant les espèces relictuelles, modales et pionnières au moment du relevé ;
- homogénéité du biotope: on vérifiera l’homogénéité du sol et du microclimat de la station, à l’échelle de la synusie. Ainsi par exemple, la canopée et le sous-bois forestier sont très différents en terme de microclimat, ce qui est également vrai pour les lisières et les chablis. Pour les animaux on s’appuiera sur la structure de la végétation d’accueil ;
- homogénéité morphologique : la taille moyenne des espèces doit être similaire, de même que les types morphologiques adaptatifs (voire anatomiques et physiologiques), par exemple les types foliaires.
Pour de plus amples explications on consultera Gillet et al. (1991), Gillet & Julve (2018), de Foucault (2020). On voit que, parmi ces critères, la prise en compte du type biologique est fondamentale. Par rapport aux travaux originels de Raunkiaer (1904, 1934), nous utilisons un modèle relativement différent (tableau 1).
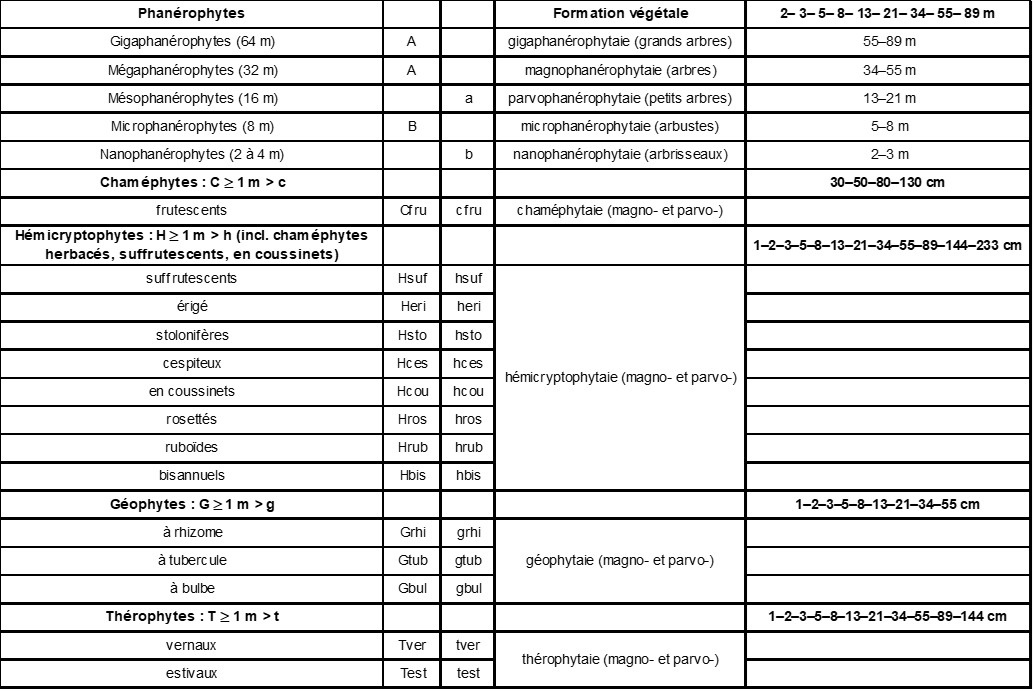
Les plantes aquatiques (-aqua), lianes (-lia), épiphytes (-épi), hémiparasites (-hpar), parasites (-par), carnivores (-car), succulentes (-suc), caducifoliées (-cad), sempervirentes (-semp) constituent des sous-types transversaux indiqués en les séparant du symbole principal par un tiret (ex : C-suc, t-hpar, h-par,
test-hpar, A-lia, C-épi-hpar, test-par, hros-car, A-semp, b-cad).Les deux derniers sous-types (caducifolié et sempervirent) sont surtout indiqués pour les phanérophytes. Si une plante réalise plusieurs types biologiques, on indique les moins fréquents entre parenthèse : ex. heri(test), test(Hbis), etc.
Il convient de noter que les chaméphytes sont restreints aux végétaux ligneux et donc en sont exclus les chaméphytes herbacés, en coussinets et suffrutescents sensu Raunkiaer. Hélophytes et hydrophytes sont répartis dans les groupes reconnus.
Dans la suite, nous nous intéresserons exclusivement aux Phanérophytes, définis comme des ligneux de taille adulte supérieure à deux mètres. Cette catégorie comprend donc les arbres (A), arbustes (B) et arbrisseaux (b). Les arbres possèdent un tronc et un houppier et sont de taille supérieure à huit mètres, les arbustes sont des modèles réduits d’arbre, avec tronc et houppier, sciatolérants et sont liés aux forêts et à leurs lisières, alors que les arbrisseaux sont dépourvus de tronc, mais possèdent plusieurs tiges ligneuses équivalentes, sont héliophiles et ne sont pas liés à des systèmes forestiers, mais sont plutôt pionniers en dynamique primaire ou en superposition dans des systèmes pelousaires ou landeux.
Sachant que, dans chaque type biologique, il existe de grandes espèces (les magnos) et de petites espèces (les parvos), il est utile de préciser les limites de classes de taille. Notons que ces limites doivent être basées sur la hauteur végétative, seule importante en écologie car la densité modale du feuillage est directement proportionnelle à la fertilité des sols, les inflorescences étant sous la dépendance d’une écologie particulière liée à la pollinisation et à la dispersion des semences ou spores. Ainsi la hauteur végétative d’Arrhenatherum elatius est d’environ trente centimètres alors que sa hauteur générative est de plus d’un mètre. Malheureusement dans les flores est souvent indiquée la hauteur générative et de plus est souvent notée la hauteur la plus fréquemment observée et non point la hauteur atteinte à l’âge adulte sans contrainte de gestion anthropogène, ce qui nuit au centrage phytosociologique des espèces ligneuses en particulier. C’est pourquoi les hauteurs indiquées dans baseflor (Julve, 1998 ff.) pour les ligneux sont les hauteurs maximales mesurées et sont souvent très supérieures aux hauteurs indiquées dans les flores usuelles. La hauteur végétative modale est une propriété fondamentale des associations synusiales. Pour les limites de classes de taille, j’ai longtemps opté pour une loi bilogarithmique : 1-2-4-8-16-32-64-128 qui fonctionne bien sur le terrain pour aider à la délimitation des synusies. Je penche désormais plutôt pour une suite de Fibonacci obtenue en additionnant les deux termes précédents : 0-1-1-2-3-5-8-13-21-34-55-89-144. Cette suite est courante dans le monde végétal pour différentes questions comme le nombre de fleurons d’un capitule ou le nombre de pétales d’une fleur ou d’une inflorescence (marguerite par exemple). Il faut noter que certains termes sont communs, ou proches, entre les deux suites. Donc pour repérer les synusies sur le terrain, on peut s’aider de ces valeurs, mais ce qui est aussi intéressant c’est qu’elles peuvent servir à centrer phytosociologiquement les espèces, c’est-à-dire à discuter l’optimum phytosociologique des espèces, indépendamment de la fréquence de leurs observations dans la nature. Notons que les synusies s’établissent généralement autour d’une valeur de la suite de Fibonacci et que l’on doit relever les taxons qui s’y trouvent, même si ce sont des juvéniles (indiqués juv. dans les relevés).
2. Le tableau des classes tempérées modales et froides
Il est présenté dans la feuille tempéré du fichier Excel en annexe (tableau 2). Seules les espèces indigènes de la flore de France sont indiquées. Le symbole σ montre qu’il s’agit d’unités synusiales. La nomenclature des taxons suit Flora Gallica (Tison & de Foucault, 2014), celle des syntaxons suit baseveg (Julve 1998ff., 2016ff.).
La classe σA Pino cembrae-Piceetea abietis Julve 1993 est une classe boréo-subalpine climacique, de répartition eurosibérienne boréale en zone tempérée froide et également présente dans les étages subalpins des montagnes européennes de la zone tempérée modale. Baseflor en recense pour la France sept taxons caractéristiques dont les hauteurs maximales respectent les valeurs 21-34-55 m de la suite de Fibonacci, ce qui soulève question car la gamme de valeur est trop large pour une seule classe, mais, comme il y a peu d’espèces en France, il n’est pas possible d’y envisager une coupure. Le problème devra être revu en prenant en compte l’ensemble des taxons de l’aire de répartition de la classe.
La classe σA Pinetea sylvestris Julve 2016 comprend des pionniers héliophiles de substrats oligotrophes de répartition eurasiatique, en tempéré froid et modal avec des enclaves en tempéré chaud ibérique. À Pinus sylvestris, dont les nombreux microtaxons semblent avoir peu de valeur, nous avons joint Juniperus communis subsp. communis, petit arbre circumboréal, héliophile et longévif (record à 1 647 années). Là encore, la classe semble hétérogène et devra être revue sur l’ensemble de l’holarctique.
Les σA Fraxino excelsioris–Quercetea roboris Gillet ex Julve 1993, comprennent les arbres climaciques, feuillus caducifoliés, du tempéré modal européen, principalement anémogames et anémochores (seuls les chênes et le merisier sont endozoochores, ce dernier étant également, avec les érables, entomogame).
On y recense trente taxons dont seuls Fagus sylvatica, Acer pseudoplatanus, Fraxinus excelsior, Taxus baccata sont caractéristiques de classe, les autres taxons étant caractéristiques d’unités inférieures, voir le tableau ennexe). La classe est assez homogène en termes de hauteurs (Fibonacci 21-34-55 m). L’If pose problème car c’est un sempervirent qui pourrait inclure cinq taxons différents selon Flora Gallica. Cette espèce fait partie des taxons à questionnement (Julve, 2023), avec Juniperus communis, Ilex aquifolium, Corylus avellana, dont il faudra résoudre le positionnement en prenant en compte la biogéographie historique (aire de répartition actuelle, mais aussi paléohistorique, variation évolutive et écologique).
Nous avons déjà exposé les arguments qui aboutissent à l’individualisation des σa Crataego monogynae-Coryletea avellanae Julve 2023 (voir Julve, 2023). Cette classe comprend de petits arbres (Fibonacci 13-21 m), une trentaine de taxons surtout entomogames et endozoochores (exception les saules), caducifoliés, se développant en sous-bois des forêts du tempéré modal. Elles peuvent également, plus rarement, acquérir leur autonomie (corylaies orophiles ou hyper-atlantiques, pommeraies kazaks), ce qui se comprend en prenant en compte leurs histoires paléogéographiques.
Les σB Rhamno catharticae-Prunetea spinosae Rivas Goday & Borja Carbonell ex Tüxen 1962 devront être réexaminés en synusial et compris de manière plus restreinte. Dans la conception présente, c’est une classe arbustive assez homogène (Fibonacci 5-8 m) constituée de taxons entomogames et endozoochores, caducifoliés, sciatolérants, liés aux lisières ou sous-bois des forêts tempérées modales. Elle comprend également une trentaine de taxons.
Les σb Rubo ulmifolii-Rosetea caninae Julve ined. ont déjà été évoqués dans plusieurs publications, mais n’ont pas fait l’objet d’une déclaration officielle. Il s’agit d’une classe d’arbrisseaux héliophiles, entomogames et endozoochores, se développant en dynamique primaire sur des systèmes rocheux, de landes ou de pelouses oligotrophiles. Elle est très homogène (Fibonacci 2-3 m). Sur les pelouses secondaires en voie d’abandon et de recolonisation ligneuse, il peut y avoir télescopage avec la classe arbustive précédente, ce qui a pu obscurcir leur séparation spatiale, pourtant convaincante sur de nombreuses stations d’observations. Elle comprend une soixantaine de taxons pami lesquels de nombreuses Rosaceae et Fabaceae. Il faut y inclure les grandes ronces arbrisseaux, mais pas les petites, majoritaires, qui entrent dans la classe Lonicero periclymeni-Rubetea plicati Haveman, Schaminée & Stortelder in Haveman 1997.
Les σb Cytisetea scopario-striati Rivas Mart. 1974 sont également une classe d’arbrisseaux d’origine ibérique qui devra être revue. L’examen du tableau de synthèse dans de Foucault et al. (2013), en enlevant les chaméphytes et les herbacées, nous montre qu’il n’y a quasiment rien de commun entre les unités ibériques et franco-britannico-allemandes, et que donc il faudra restreindre le concept de cette classe à la partie ibérique (où il y a très peu de Cytisus scoparius transgressif d’ailleurs). Les unités du tempéré modal s’inscriront très bien dans les Rubo ulmifolii-Rosetea caninae.
3. Le tableau des classes hygrophiles de bois tendres, les saulaies et petits ligneux orophiles
La feuille saules (tableau 3) présente d’abord la classe σA Betuletea pendulo-pubescentis Julve 2016. Elle comprend des arbres à bois tendres, peu longévifs et à croissance rapide, pionniers en dynamique primaire ou secondaire (Julve & Vitte, 2019), anémogames et anémochores, qui s’implantent en mosaïques, en lisières ou en clairières des forêts climaciques du tempéré froid et du tempéré modal en Europe et en Asie septentrionale. La classe est assez homogène en taille (Fibonacci 21-34 m).
Les σA Salici albae-Populetea nigrae Julve 2016 sont plus franchement hydrophiles, dans les vallées alluviales, plus ou moins régulièrement en crues perturbatrices. Elle rassemble aussi des bois tendres, saules et des peupliers de taille également assez homogène (Fibonacci 21-34 m). La répartition est européenne occidentale, en tempéré modal et tempéré chaud.
Les σBb Salicetea purpureae subsp. purpureae Moor 1958 devraient être conçus en synusial. Ces saules arbustifs d’environ huit mètres forment des lisières des σA Salici albae-Populetea nigrae, parfois aussi des saulaies arbustives pionnières de vallées alluviales graveleuses dans le cours amont des rivières à courant fort, en particulier en montagne. On voit tout de suite que, dans la liste, une espèce ne convient pas : Myricaria germanica est un arbrisseau beaucoup plus petit que les saules et moins lignifié. Mais comme il est seul dans sa catégorie, nous l’avons provisoirement rattaché à cette classe, pour les valeurs inférieures à un mètre.
Les σBb Salici appendiculatae-Alnetea alnobetulae sont une classe orophile, montagnarde à subalpine, européenne occidentale des zones tempérées modales. Il s’agit d’une vingtaine d’espèces de tailles assez homogènes (Fibonacci 2-3-5 m) qui devraient être comparées aux associations montagnardes des Rhamno-Prunetea.
Les σb Franguletea alni Doing 1962 sont une classe d’arbrisseaux hygrophiles à amphibies, assez homogène (Fibonacci 2-3-5 m). plutôt planitiaire-collinéenne et tempérée modale.
Enfin les σb Salicetea bicolori-lapponum Julve 2003 sont une classe d’arbrisseaux boréo-subalpins des montagnes de la zone tempérée modale et tempérée froide. Le climat froid réduit fortement leur taille et les fait tendre vers l’habitus sous-arbrisseau des chaméphytaies en climat très froid. Ils sont tous entomogames et anémochores. Il faut remarquer que la suite de Fibonacci peut également se décliner en décimètres pour les valeurs inférieures à un mètre. Il faudra peut-être découper cette classe en séparant les vrais arbrisseaux des sous-arbrisseaux à comparer aux saules nains arctico-alpins.
4. Le tableau des classes méditerranéennes
La feuille méditerranéen (tableau 4) présente les σA Pinetea nigrae Julve 2016, classe oroméditerranéenne à conifères anémogames et anémochores, de répartition orophile périméditerranéenne occidentale. Elle est relativement homogène (Fibonacci 21-34-55).
Les σA Quercetea fagineo-pubescentis Julve 2017 sont une classe sub-supraméditerranéenne, périphérique à la Méditerranée septentrionale et occidentale, du Maghreb et de l’Espagne à la Turquie, Syrie et jusqu’en Iran. Les arbres y sont souvent marcescents. Elle comprend en France une trentaine d’espèces, mais est bien plus riche sur la totalité de son aire. L’écologie reproductive se rapporte à deux principaux types : anémogame/anémochore et entomogame/endozoochore. Les tailles sont assez hétérogènes (Fibonacci 13-21-34-55) et laissent présager la possible existence d’une classe de petits arbres, mais qui n’apparaît pas encore clairement.
Les σA Pino halepensis-Quercetea ilicis (Braun-Blanq. 1947) B. Foucault & Julve 1991 sont une classe périméditerranéenne de climat tempéré chaud, sauf Libye et Égypte qui sont tropicales. Rappelons que le climat méditerranéen se caractérise par quatre saisons et la possibilité de gel, comme toutes les zones tempérées, et une moyenne annuelle de température d’environ 15 °C, alors que les zones tropicales n’ont pas de saison thermique, pas de gel (en plaine !) et une moyenne annuelle d’environ 20-25 °C. La classe rassemble les associations du mésoméditerranéen et du thermoméditerranéen. Les arbres y sont sempervirents et de taille homogène (Fibonacci 21-34). Elle comprend en France une quinzaine d’espèces, généralement anémogames. La classe constitue des forêts sempervirentes mixtes (Gymnospermes/Angiospermes), mais les conifères sont plutôt thermoméditerranéens et pionniers, brûlent facilement, alors que les Angiospermes sont plutôt mésoméditerranéens et climaciques, résistent mieux au feu, ce qui laisse envisager la possibilité d’une future séparation en deux classes, les Pinetalia halepensis Biondi et al. 2014 ayant déjà été élevées au rang d’ordre.
Viennent ensuite trois classes qui ont été mélangées historiquement sous le concept de σBb Pistacio lentisci-Rhamnetea alaterni subsp. alaterni Julve 1993. Il faut reconnaître que les végétations ligneuses méditerranéennes sont très perturbées et ouvertes, ce qui complique singulièrement leur analyse. Des milliers d’années de dent du mouton et le stress hydrique estival, le décapage des sols déjà superficiels, les feux récurrents ne permettent que difficilement la restructuration des formations ligneuses. L’étude des hauteurs végétatives modales permet d’envisager :
- une classe de petits arbres, σa Pistacio terebenthi-Phillyreetea latifoliae Julve nov. prov. hoc loco, formée de petits arbres homogènes (Fibonacci 8-13), endozoochores, mais hétérogènes du point de vue pollinisation, sempervirents ou caducifoliés, mixtes au sens Angiospermes/Gymnospermes. Les classes mixtes sont souvent intermédiaires géographiques entre des classes pures du point de vue morphologie foliaire adaptative, la morphologie foliaire adaptative étant une caractéristique fondamentale des différents types forestiers climaciques du globe. La classe compte une vingtaine de taxons en France ;
- une classe arbustive, σB Myrto communis-Rhamnetea alaterni Julve nov. prov. hoc loco, formée d’arbustes méditerranéens homogènes (Fibonacci 5-8), plutôt sempervirents (sauf quelques exceptions) et sclérophylles, plutôt entomogames et endozoochores. La classe compte également une vingtaine de taxons en France ;
- une classe d’arbrisseaux, très homogène (Fibonacci 2-3), les σb Pistacio lentisci-Rhamnetea lycioidis Julve nov. prov. hoc loco, sempervirents ou caducifoliés, entomogames et plutôt endozoochores. La classe compte encore une vingtaine de taxons en France et ces convergences numériques ne peuvent pas être fortuites.
Notons que nous avons provisoirement intégré les lianes ligneuses ou semi-ligneuses, dans les différentes classes, mais que l’avenir permettra peut-être de les séparer en synusies autonomes, à l’image des σa–lia Hederetea helicis Julve 2005, dont on a pu observer l’expression sur n’importe quel support, arbre, arbuste, falaise, paroi, mur, poteau… et dont les draperies semi-héliophiles colonisent plutôt les forêts secondaires ou les lisières des bois, tout comme en zone tropicale. À ce sujet il convient de bien séparer les lianes enracinées au sol et grimpant sur des supports des véritables épiphytes enracinés dans les microsols des bases de branches ou des fissures de troncs.
5. Conclusion
L’utilisation des hauteurs maximales des ligneux, qui semblent suivre une suite de Fibonacci, nous a permis une réflexion nouvelle sur le contenu et les limites des classes phytosociologiques phanérophytiques présentes en France et dans les pays mitoyens. Elle sert aussi de critère pour délimiter les synusies ligneuses sur le terrain.
Bibliographie
Foucault B. (de), 2020. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest–asiatique et nord–africaine. Evaxiana 7 : 5-248.
Foucault B. (de), Lazare J.-J. & Bioret F., 2013. Contribution au prodrome des végétations de France : les Cytisetea scopario-striati Rivas-Mart. 1975. Journal de botanique de la Société botanique de France 64 : 69-90
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Gillet F. & Julve Ph., 2018. The integrated synusial approach to vegetation classification and analysis. Phytocoenologia, Special Issue on Vegetation Classification Approaches, Phytocoenologia 48 : 141-152.
Julve Ph., 1998 ff. baseflor, index botanique, écologique et chorologique de la Flore de France [date de la version utilisée]. Programme Catminat. http://www.tela–botanica.org.
Julve Ph., 1998 ff. baseveg, répertoire synonymique des unités phytosociologiques de France [date de la version utilisée]. Programme Catminat. http://www.tela–botanica.org.
Julve Ph., 2016 ff. (1998 ff). baseveg World vegetation database [date de la version utilisée]. Programme Catminat. http://www.tela–botanica.org.
Julve Ph., 2023. Ce que peut apporter l’analyse d’un bois bombardé. Bulletin de la Société de botanique du nord de la France 76 : 88-100.
Julve Ph. & Vitte M., 2019. Des synusies aux terroirs : un modèle dynamique et des échelles spatiales. Actes du colloque international de Brest, septembre 2014, La phytosociologie paysagère, des concepts aux applications. Documents phytosociologiques, 3e série, 9 : 46-56.
Raunkiaer Ch., 1904. Om biologiske typer, med Hensyn til Planternes Tilpasning til at overle ugunstige Aarister. Botaniker Tidsskrift 26.
Raunkiaer Ch., 1934. The life forms of plants and statistical plant geography; being the collected papers of Ch. Raunkiaer. Oxford Clarendon Press. 632 p.
Tison J.-M. & de Foucault B. (coords), 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1 196 p.


