
Les changements nomenclaturaux et taxonomiques récents et nouveautés chez les Ptéridophytes de France
Title
Recent nomenclatural and taxonomic changes and novelties in the Pteridophytes of France
Résumé
Cette note présente les changements nomenclaturaux et taxonomiques qui ont affecté les Ptéridophytes de France depuis 2000. Les principaux arguments sont exposés et les références bibliographiques afférentes indiquées.
Abstract
The note presents the nomenclatural and taxonomic changes concerning the French Pteridophytes since 2000. The main reasons are exposed, and the related bibliographic references mentioned.
1. Introduction
La classification des Ptéridophytes (Lycophytes et Fougères) a fait l’objet d’importantes variations. Cependant, depuis ces trente dernières années, grâce aux résultats acquis par la phylogénie moléculaire et aux nombreuses révisions de familles, genres et groupes d’espèces réalisées dans le monde entier, les familles ont pu être mieux délimitées et sont désormais bien harmonisées, comme le montre la nouvelle classification de PPG I (2016). Ces résultats ont eu pour conséquence une importante évolution de la nomenclature générique et spécifique. Nous présentons ici les changements de noms qui, depuis 2000, affectent les Ptéridophytes de France. Ils sont de deux sortes : d’une part nomenclaturaux, c’est-à-dire liés aux règles du code de nomenclature (Turland et al., 2019), et d’autre part taxonomiques, plus subjectifs car liés à l’évolution des concepts phylogénétiques différemment appréciés selon les auteurs. Sont concernées les familles des Lycopodiaceae, des Isoetaceae, des Ophioglossaceae, des Hymenophyllaceae, des Cystopteridaceae, des Pteridaceae, des Blechnaceae, des Thelypteridaceae et des Dryopteridaceae. Quelques points divers ont également été réglés au sein des Equisetaceae et des Aspleniaceae. Toutes ces modifications sont indiquées dans le récent ouvrage de Prelli & Boudrie (2021).
Le texte ci-dessous reprend les éléments du PowerPoint présenté le 3 octobre 2021 à Montpellier lors du colloque des Convergences botaniques organisé par la Société botanique d’Occitanie. Les références bibliographiques concernant chaque famille sont données en fin de traitement du sujet en question.
Nous profitons également de la présente publication pour corriger quelques erreurs repérées dans le référentiel taxonomique de notre ouvrage cité ci-dessus concernant les taxons Lycopodium lagopus, Asplenium adiantum-nigrum var. silesiacum et Asplenium obovatum subsp. billotii.
2. L’évolution de la classification
Traditionnellement, jusqu’au début du xxie siècle (cf. par exemple Prelli, 2002), on utilisait le terme de « Ptéridophytes » pour désigner les fougères et les plantes alliées, ces dernières comprenant les ordres des Lycopodiales, des Selaginellales, des Isoetales, des Psilotales et des Equisetales comme indiqué sur la figure 1.

La nouvelle classification (figure 2 ; PPG I, 2016 ; voir aussi le résumé proposé par Sessa, 2018) est fondée sur un concept phylogénétique tendant à ne définir, au final et si possible, que des entités monophylétiques, mais tenant compte, évidemment, de la morphologie. Toutefois, cette conception, telle que proposée, a conduit à l’établissement, ou rétablissement, de nombreux « petits » genres. Un autre concept a été proposé (Christenhusz & Chase, 2014), plus global au niveau du regroupement de certaines familles d’Eupolypodes (Aspleniaceae et Dryopteridaceae), mais il ne semble pas faire l’unanimité chez l’ensemble des ptéridologues ni pour le reclassement des herbiers. Quoi qu’il en soit, dans notre ouvrage (Prelli & Boudrie, 2021), nous avons choisi de suivre PPG I, même si ce n’était pas une obligation (PPG I, p. 564 : « This classification is not intended as the final word on lycophyte and fern taxonomy, but rather a summary statement of current hypotheses ») puisque les noms de genres, employés précédemment pour certaines de nos espèces européennes (par exemple dans les genres Lycopodium, Cheilanthes, Blechnum, Stegnogramma…), étant validement publiés, restent utilisables dans une autre conception de la classification, selon les auteurs.
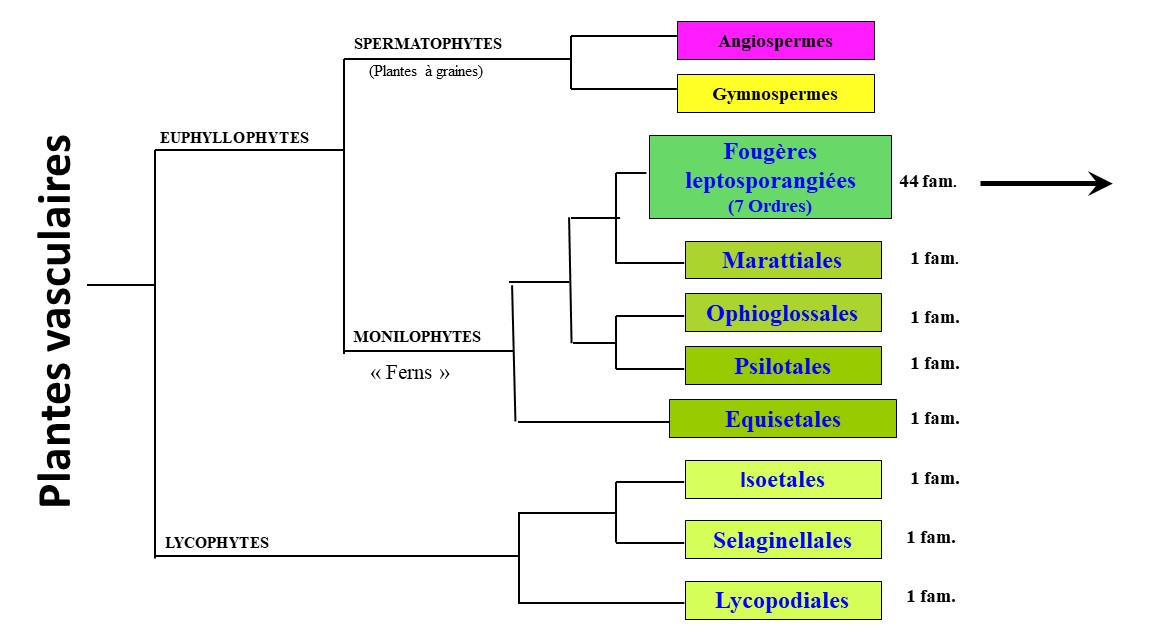
La nouvelle classification de PPG I comprend donc maintenant les Lycophytes, composées de seulement trois ordres (Lycopodiales, Selaginellales et Isoetales), et les Euphyllophytes, divisées d’une part en Monilophytes (Fougères au sens large) et d’autre part en Spermatophytes (Gymnospermes et Angiospermes).
Concernant l’utilisation du terme de « Monilophytes » (signifiant que, dans une section transversale de la tige, la morphologie du xylème rappelle vaguement un collier en forme de chapelet), apparu dès la fin des années 1990 et officialisé par Cantino et al. (2007), ce terme n’est basé sur aucun type et est maintenant préférentiellement remplacé, notamment par les botanistes anglo-saxons, par le terme de « Ferns » [= Fougères au sens large]. Parmi les principaux changements, on notera le passage des Equisetales au sein des Fougères s. l. (Pryer et al., 2001 ; Smith et al., 2006). L’ancienne sous-division des Polypodiophytes équivaut globalement au groupe de familles désormais nommé « Fougères leptosporangiées », mais sans les Ophioglossales (figure 3). L’ancien ordre des Filicales correspond maintenant à l’ordre des Polypodiales, mais sans les Hymenophyllales.
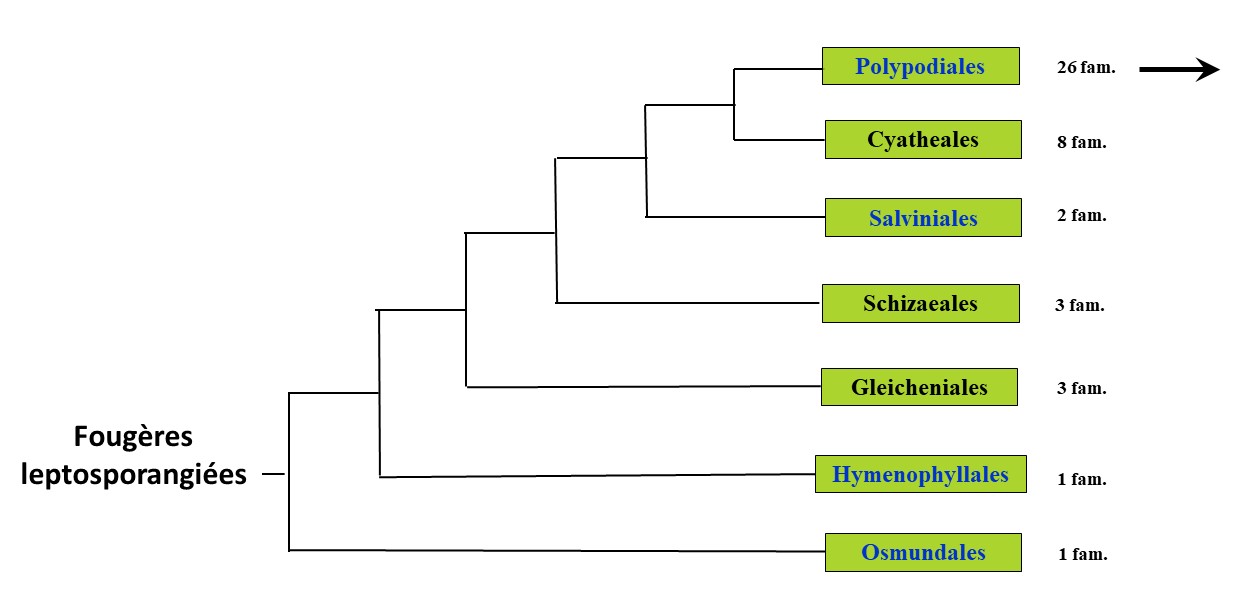
L’ordre des Polypodiales (figure 4) comprend, pour la France, d’une part les deux familles basales des Dennstaedtiaceae et des Pteridaceae et d’autre part les sous-ordres des Eupolypodes, celui des Eupolypodes I (ou Polypodiineae) et celui des Eupolypodes II (ou Aspleniineae). La distinction entre ces deux groupes est fondée sur la vascularisation du pétiole, les Eupolypodes I possédant trois faisceaux vasculaires ou plus, hormis quelques exceptions, comme la sous-famille des Grammitidoideae, et les Eupolypodes II n’en possédant que deux, hormis quelques exceptions comme les Blechnaceae (Rothfels et al., 2012).

Même s’il n’apparaît plus officiellement dans la nouvelle classification, le terme de « Ptéridophytes » (ou « Pteridophyta ») reste largement utilisé par tous les ptéridologues, non seulement dans le langage courant mais également dans les publications, et désigne globalement les « Lycophytes et Fougères ».
L’ensemble des Lycophytes et Fougères comprend actuellement (2022) 14 ordres, 51 familles, 345 genres et environ 12 000 espèces. En 2016, PPG I présentait 337 genres et, depuis cette date, au moins 8 genres nouveaux dont un ancien réhabilité doivent être ajoutés : Adetogramma T.E. Almeida (2017 ; Polypodiaceae), Baja Windham & L.O. George (2019 ; Pteridaceae), Ephemeropteris R.C. Moran & Sundue (2019 ; Athyriaceae), Mickelopteris Fraser-Jenk. (2016 ; Pteridaceae), Mineirella Ponce & Scataglini (2021 ; Pteridaceae), Parrisia Shalisko & Sundue (2019 ; Polypodiaceae), Sahashia Li Bing Zhang & Liang Zhang (2020 ; Ophioglossaceae) et Spicantopsis Nakai (1933 ; Blechnaceae).
Références
Cantino P.D., Doyle J.A., Graham S.W., Judd W.S., Olmstead R.G., Soltis D.E., Soltis P.S. & Donoghue M.J., 2007. Towards a phylogenetic nomenclature of Tracheophyta. Taxon 56 (3) : E1-E44.
Christenhusz M.J.M. & Chase M.W., 2014. Trends and concepts in fern classification. Annals of Botany 113 : 571-594.
PPG I (Schuettpelz E. et al.), 2016. A community-derived classification for extant lycophytes and ferns. Journal of Systematics and Evolution 54 (6) : 563-603.
Prelli R. & Boudrie M., 2021. Les Fougères et plantes alliées d’Europe. Ed. Biotope, Mèze, 528 p.
Pryer K.M., Schneider H., Smith A.R., Cranfill R., Wolf P.G., Hunt J.S. & Sipes S.D., 2001. Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants. Nature 409 : 618-621.
Rothfels C.J., Sundue M.A., Kuo L.-Y., Larsson A., Kato M., Schuettpelz E. & Pryer K.M., 2012. A revised family-level classification for Eupolypod II ferns (Polypodiidae : Polypodiales). Taxon 61 : 515-533.
Sessa E.B., 2018. Evolution and Classification of Ferns and Lycophytes. In H. Fernández (ed.), Current Advances in Fern Research, Department of Organisms and Systems Biology, Oviedo University, Oviedo, Spain : 179-200.
Smith A.R., Pryer K.M., Schuettpelz E., Korall P., Schneider H. & Wolf P.G., 2006. A classification for extant ferns. Taxon 55 : 705-731.
Turland N.J., Wiersema J.H., Barrie F.R., Greuter W., Hawksworth D.L., Heren-Deen P.S., Knapp S., Kusber W.-H., Li D.-Z., Marhold K., May T.W., McNeill J., Monro A.M., Prado J., Price M.J. & Smith G.F. (eds.), Loizeau P.-A., Marder A. & Price M.J. (trad.), 2019. Code international de nomenclature pour les Algues, les Champignons et les Plantes (Code de Shenzhen) adopté par le Dix-Neuvième Congrès international de botanique, Shenzhen, Chine, juillet 2017. Conservatoire et Jardin botaniques de la Ville de Genève, Publication hors-série 19, 228 p., DOI : 10.5281/zenodo.2558299.
3. Modifications dans les différentes familles
3.1. Lycopodiaceae
Depuis Linné (1753) jusque vers le milieu du xxe siècle, les Lycopodes étaient, en général, tous rangés sous le nom de genre Lycopodium À partir des années 1980-1990, les quatre genres cités ci-après sont considérés comme constituant la famille des Lycopodiaceae (Øllgaard, 1987, 1990) : Huperzia Bernh. (photo 1), Phylloglossum Kunze, Lycopodium L. et Lycopodiella Holub (photo 2), dont certains (Lycopodium et Lycopodiella) divisés en sections.


La nouvelle conception des Lycopodes (Wikström & Kenrick, 2001 ; Øllgaard, 2015 ; Field et al., 2016 ; PPG I, 2016) conduit à considérer ces anciennes sections comme des genres à part entière. De ce fait, les ajustements suivants ont été effectués :
- rétablissement du genre Diphasiastrum Holub, de la section Complanata Victorin, avec trois espèces et trois taxons considérés comme des hybrides :
- D. alpinum (L.) Holub
- D. complanatum (L.) Holub
- D. tristachyum (Pursh) Holub (photo 3)
- D. ×issleri (Rouy) Holub [ alpinum × D. complanatum]
- D. ×oellgaardii Stoor, Boudrie, Jérôme, Horn & Bennert [ alpinum × D. tristachyum] (photo 4)
- D. ×zeilleri (Rouy) Holub [ complanatum × D. tristachyum] ;


- établissement d’un nouveau genre, Spinulum, de la section Annotina (Rouy) Holub, et Lycopodium annotinum devient Spinulum annotinum (L.) A. Haines, 2003 [subsp. annotinum] (photo 5).
Lycopodium lagopus ( ex C. Hartm.) Zinserl. ex Kuzen., 1953 : suite à une erreur de copier-coller, la date de publication du basionyme de ce taxon est 1858 (et non 1831), soit Lycopodium clavatum var. lagopus Laest. ex C. Hartm., 1858.

Références
Field A.R., Testo W., Bostock P.D.B., Holtum J.A.M., Waycott M,. 2016. Molecular phylogenetics and the morphology of the Lycopodiaceae subfamily Huperzioideae supports three genera: Huperzia, Phlegmariurus and Phylloglossum. Molecular Phylogenetics and. Evolution 94 : 635-657.
Haines A., 2003. The families Huperziaceae and Lycopodiaceae in New England: A taxonomic and ecological reference. Bar Harbor: V.F. Thomas Co., 100 p. [: 85-86].
Linnaeus C., 1753. Species Plantarum, 2. Laurentius Salvius, Stockholm, 1 200 p.
Øllgaard B., 1987. A revised classification of the Lycopodiaceae s. lat. Opera Botanica 92 : 153-178.
Øllgaard B., 1990. Lycopodiaceae. In K.U. Kramer & P.S. Green (eds.), Pteridophytes and Gymnosperms (K. Kubitzki (ed.), The Families and Genera of Vascular Plants. I), Springer-Verlag, Berlin, Heidelberg, New York : 31-39.
Øllgaard B., 2015. Six new species and some nomenclatural changes in neotropical Lycopodiaceae. Nordic Journal of Botany 33 : 186-196.
Wikström N. & Kenrick P., 2001. Evolution of Lycopodiaceae (Lycopsida): Estimating divergence times from rbcL gene sequences by use of nonparametric rate smoothing. Molecular Phylogenetics and. Evolution 19 : 177-186.
3.2. Isoetaceae
Après de nombreuses tergiversations quant à l’application du nom Isoetes setacea [notamment à cause de confusions entre Isoetes echinospora Durieu, I. tenella (Léman) Desv. et I. delilei Rothm.], le problème a pu être enfin réglé et l’historique de ces diverses confusions a été bien détaillé et explicité par Greuter & Troia (2015). La révision du spécimen type de Lamarck [P00565109, in P-LA] d’Isoetes setacea Lam. a montré que la plante correspondait en fait à Isoetes lacustris L. et non à I. echinospora ou à I. delilei. De ce fait, I. setacea Lam. est synonyme d’I. lacustris. En conséquence, l’Isoetes des mares temporaires du maquis méditerranéen, précédemment nommé I. setacea, doit être nommé Isoetes delilei Rothm., 1944 (photo 6).

L’orthographe correcte d’Isoetes duriei Bory est Isoetes durieui Bory (Art. 60.8.(a), Recommandation 60C.3 et, notamment, la note (d) de la Recommandation 60C.4 qui donne l’exemple de l’épithète jussieui; Turland et al., 2019, ainsi que les sites web IPNI et Tropicos).
Isoetes velata Braun, 1849, étant un nom. illeg. à cause de l’inclusion [sous le nom I. velata var. longissima A. Braun, pl. 37 de Braun, 1849 ; photo 7], en tant que variété, du nom légitime antérieur, I. longissima Bory, 1844, mais non basé automatiquement sur le même type (Art. 7.5 ; Turland et al., 2019 ; Troia & Greuter, 2014 : 15-16 ; Troia et al., 2014), deux changements sont intervenus :
- Isoetes longissima Bory, 1844, est le nom correct pour les espèces classées précédemment sous le nom d’ velata (photo 8),
- Isoetes tenuissima Boreau, 1850, est le nom actuellement accepté pour velata subsp. tenuissima (photo 9).


Quelques nouveautés sont à signaler :
- dans le groupe « histrix» avec deux espèces anciennement décrites et réhabilitées (cf. respectivement Troia & Rouhan, 2018 ; Bagella et al., 2014) ; leur statut en tant qu’espèces à part entière reste toutefois à préciser :
- Isoetes delalandei Lloyd, 1852 (de Bretagne),
- Isoetes gymnocarpa (Gennari) A. Braun, 1864 (du sud de la Corse) ;
- dans le groupe « lacustris», une espèce a été décrite :
- Isoetes « creussensis» Lazare & Riba, 2010 [cependant, son statut reste à confirmer quant à son degré de ploïdie car des analyses récentes de spécimens de la station type ont montré la présence de plantes 10x, donc comme lacustris, et non 12x comme supposé ; Abdulhak, 2016] ;
- chez les hybrides d’Isoetes, un premier point a été éclairci avec la description d’un nouvel hybride (Brunton & Rumsey, 2021) – le nom d’I. hickeyi C. Taylor & Luebke utilisé jusque-là pour désigner l’hybride européen entre I. echinospora et I. lacustris s’appliquant en réalité à des taxons nord-américains – :
- Isoetes ×jermyi F. Brunt. & Rumsey, 2021 (I. echinospora × I. lacustris) ;
- un second point a été abordé et concerne le taxon I. brochonii, resté longtemps énigmatique. Les dernières investigations (Brunton et al., 2020) montrent que ce taxon est un hybride du fait de la présence de spores avortées sur les spécimens types de Motelay. Mais, si l’un de ses parents semble bien défini (I. echinospora), le second reste à identifier et pourrait être I. creussensis si ce dernier est confirmé et si I. brochonii est bien 7x (voir remarque ci-dessus et discussion in Prelli & Boudrie, 2021 : 90-91).
- Isoetes ×brochonii Motelay, 1892 ( echinospora × I. « creussensis»).
Références
Abdulhak S., 2016. Notes sur les Isoetes sp. du massif du Carlit, Pyrénées-Orientales. Rapport Conservatoire botanique national alpin, 12 p.
Bagella S., Peruzzi L., Caria M.C. & Filigheddu R., 2014. Unraveling the taxonomy and nomenclature of the Isoetes histrix Bory species complex (Isoetaceae, Lycopodiidae). Turkish Journal of Botany 38 : 383-387.
Brunton D.F., Sokoloff P.C. & Aymerich P., 2020. The taxonomy, status and origin of Isoetes ×brochonii and I. creussensis (Isoetaceae), two Pyrenean endemic taxa. Botany Letters 167 (4) : 391-408.
Brunton D.F. & Rumsey F.J., 2021. Isoetes ×jermyi hyb. nov. (Isoetaceae), a new quillwort (lycophyte) hybrid from western Europe. Botany Letters 168 (4) : 503-511.
Greuter W. & Troia A., 2015. Disentangling Isoetes setacea and removing threats to Isoetes echinospora. Taxon 64 : 811-815.
Lazare J.-J. & Riba S., 2010. Les isoètes de l’Estany de Creussans (Andorre). Journal de botanique de la Société botanique de France 90 : 19-26.
Troia A. & Greuter W., 2014. A critical conspectus of Italian Isoetes (Isoetaceae). Plant Biosystems 148 (1) : 15-16.
Troia A., Raimondo F.M. & Campisi P., 2014. The Isoetes longissima complex (Isoetaceae) in Italy: observations on the morphology of spores and leaves, and taxonomic implications. Phytotaxa 174 (3) : 149-156.
Troia A. & Rouhan G., 2018. Clarifying the nomenclature of some Euro-Mediterranean quillworts (Isoetes, Isoetaceae): Indicator species and species of conservation concern. Taxon 67 : 996-1004.
3.3. Equisetaceae
Dans le sous-genre Hippochaete Milde, regroupant, pour la France, les trois espèces Equisetum hyemale, E. ramosissimum Desf. et E. variegatum Schleich. ex F. Weber & D. Mohr, des hybrides triploïdes ont été mis en évidence dont deux sont présents en Alsace (Lubienski & Bennert, 2006 ; Lubienski et al., 2010 ; Lubienski, 2013). Ils correspondent à la combinaison d’E. hyemale avec chacun de ses hybrides diploïdes avec d’une part E. variegatum et d’autre part E. ramosissimum. Ils peuvent être notés de deux façons différentes, soit en individualisant les espèces en présence, soit en tenant compte des hybrides diploïdes avec E. hyemale :
- Equisetum ×alsaticum (H.P. Fuchs & Geissert) G. Phil. ex Lubienski & Bennert, 2006 (photo 10)
(E. hyemale × E. hyemale × E. variegatum)
(E. hyemale × E. ×mackayi)
Ce taxon était connu sous le nom d’Hippochaete alsatica H.P. Fuchs & Geissert, 1980 et était considéré (par exemple Derrick et al., 1987 : 38), jusqu’aux travaux récents de Lubienski & Bennert (2006), comme synonyme d’E. ×trachyodon [= maintenant E. ×mackayi ; voir plus bas] ;
- Equisetum ×ascendens Lubienski & Bennert, 2010 (photo 11)
(E. hyemale × E. hyemale × E. ramosissimum)
(E. hyemale × E. ×moorei).
Le nom correct et la synonymie de l’hybride entre hyemale et E. variegatum, qui était connu sous le nom d’E. ×trachyodon, ont été précisés (Bizot & Pétrement, 2009), l’ajout d’une nothosubsp. autonyme étant dû à l’existence de la nothosubsp. jesupii (avec une autre sous-espèce d’E. hyemale), présente en Amérique du Nord.
- Equisetum ×mackayi (Newman) Brichan, 1842 nothosubsp. mackayi (photo 12)
(E. hyemale subsp. hyemale × E. variegatum subsp. variegatum)
basionyme : Equisetum hyemale var. mackayi Newman, 1842
syn. : Equisetum ×trachyodon (Döll) W.D.J. Koch, 1845
Références
Bizot A. & Pétrement B., 2009. Découverte d’Equisetum ×trachyodon (A. Braun) Koch (Equisetaceae – Pteridophyta) en Belgique à Etalle et modification du taxonyme. Adoxa 61 : 14-21.
Derrick L.N., Jermy A.C. & Paul A.M., 1987. Checklist of European Pteridophytes. Sommerfeltia 6 : i-xx, 1-94.
Lubienski M., 2013. Hybriden der Gattung Equisetum (Equisetaceae, Equisetopsida, Monilophyta) in Europa. Bericht der Institut für Landschafts- und Pflanzenokologie der Universität Hohenheim 22 : 91-124.
Lubienski M. & Bennert H.W., 2006. Equisetum ×alsaticum (Equisetaceae, Pteridophyta) in Mitteleuropa. Carolinea 64 : 107-118.
Lubienski M., Bennert H.W. & Körner S., 2010. Two new triploid hybrids in Equisetum subgenus Hippochaete for Central Europe and notes on the taxonomic value of « Equisetum trachyodon forma Fuchsii » (Equisetaceae, Pteridophyta). Nova Hedwigia 90 (3-4) : 321-341.
3.4. Ophioglossaceae
Dans le genre Ophioglossum, un hybride nouveau a été décrit :
- Ophioglossum ×pseudoazoricum Peruzzi, Pierini, Magrini, Marchetti & Viane, 2015 (photo 13)
(O. azoricum × O. vulgatum)
La révision de la famille des Ophioglossaceae (Hauk et al., 2003) et notamment du genre Botrychium a conduit à un tranfert de genre :
- Botrychium multifidum (S.G. Gmel.) est transféré dans le genre Sceptridium Lyon, 1905, et devient Sceptridium multifidum (S.G. Gmel.) M. Nishida, 1958 (photo 14).
Références
Hauk W.D., Parks C.R. & Chase M.W., 2003. Phylogenetic studies of Ophioglossaceae: evidence from rbcL and trnL-F plastid DNA sequences and morphology. Molecular Phylogenetics and Evolution 28 : 131-151.
Nishida M., 1958. (101) Sceptridium multifidum. In M. Tagawa, Fern miscellany (10), Japanese Journal of Botany 33 (7) : 200-201.
Peruzzi L., Pierini B., Magrini S., Andreucci A., Marchetti D. & Viane R., 2015. Three new hybrids of Ophioglossum (Ophioglossaceae) from Monte Pisano, Tuscany (Central Italy). Plant Biosystems 149 : 737-746.
3.5. Hymenophyllaceae
La révision de la famille des Hymenophyllaceae au niveau mondial (Ebihara et al., 2006) a conduit à la modification suivante :
- Trichomanes speciosum est transféré dans le genre Vandenboschia Copel., 1938, et devient Vandenboschia speciosa (Willd.) G. Kunkel, 1966 (photos 15 et 16).
Le genre Trichomanes L. est, en effet, basé sur Trichomanes crispum L. qui est une espèce d’Amérique tropicale, à caractères morphologiques différents de ceux du genre Vandenboschia (chez Trichomanes, limbe penné, à fausses nervures, à nervation généralement catadrome ; chez Vandenboschia, limbe bipenné ou plus, sans fausses nervures, à nervation anadrome).
Référence
Ebihara A., Dubuisson J.-Y., Iwatsuki K., Hennequin S. & Ito M., 2006. A taxonomic revision of Hymenophyllaceae. Blumea 51 : 221-280.
3.6. Pteridaceae
Le nom de genre Cheilanthes, traditionnellement appliqué aux espèces eurasiatiques, a fait l’objet, auprès du Committee for Pteridophyta du code de nomenclature, d’une proposition de conservation de nom (Panigrahi, 1987) contre l’application du nom de genre Allosorus Bernh., mais qui a été rejetée (Lellinger, 1993), sous prétexte d’insuffisance. Le nom de genre Cheilanthes a, en conséquence, été appliqué aux taxons américains, avec Ch. micropteris Sw. comme type. Toutefois, les taxons eurasiatiques sont restés au nom de Cheilanthes jusqu’à ce que Christenhusz (2012) les transfère sous le nom de genre Allosorus Bernh. Par la suite, Rumsey (2014) établit les combinaisons des taxons hybrides dans ce genre. Plus tard, Fraser-Jenkins et al. (2017a) démontrent que le nom de genre Allosorus Bernh. a été appliqué par erreur aux taxons eurasiatiques. En effet, Allosorus pusillus Bernh., 1806, type du genre Allosorus Bernh., est un nom. illeg. car, d’une part, Bernhardi l’a fondé sur plusieurs espèces mélangées de Pteridaceae, incluant, dans la synonymie, le nom légitime antérieur Pteris acrostica Balb., 1801, et, d’autre part, Allosorus a été typifié par J. Smith en 1875 sur l’espèce Pteris crispa (L.) All. [= Cryptogramma crispa (L.) R. Br. ex Hook.]. Ils établissent également que le nom le plus ancien validement publié pour les espèces eurasiatiques est celui de Oeosporangium Vis., 1867. Se fondant sur ces éléments, Fraser-Jenkins et al. (2017b) ont déposé auprès du Committee for Pteridophyta une nouvelle demande de rejet du nom d’Allosorus en application aux anciens Cheilanthes eurasiatiques, proposition qui a été recommandée sur vote majoritairement favorable (Applequist, 2019). Les diverses combinaisons sous Oeosporangium des espèces (photos 17 à 19) et des hybrides ont été établies par Fraser-Jenkins et al. (2017a), Sáez & Aymerich (2017) et Arana & Moreno Saiz (2020).
La typification du genre Notholaena Br., 1810, a, depuis sa lectotypification initiale par Smith (1875), fait l’objet de nombreux articles (notamment Pichi Sermolli, 1977, 1981, 1983, 1989 ; Tryon, 1956 ; Tryon & Tryon, 1980), controversés. L’épithète marantae a été successivement affectée à plusieurs noms de genre avec les combinaisons Acrostichum marantae L., 1753, Polypodium marantae (L.) Hoffm., 1796, Notholaena marantae (L.) Desv., 1813, Gymnogramma marantae (L.) Mett., 1856, Ceterach marantae (L.) DC., 1865, Cheilanthes marantae (L.) Domin, 1913, Gymnopteris marantae (L.) Ching, 1965, Paraceterach marantae (L.) R.M. Tryon, 1986, Paragymnopteris marantae (L.) K.H. Shing, 1994, Hemionitis marantae (L.) Christenh., 2018, ceci traduisant les difficultés de placement de cette espèce au sein des Pteridaceae. Suite à ces différents placements non satisfaisants, c’est finalement Shing (1993) qui a établi le nouveau nom de genre Paragymnopteris, en le typifiant sur Acrostichum marantae L. On considère actuellement le nom de genre Notholaena R. Br. comme typifié sur l’espèce du Nouveau Monde Pteris trichomanoides L. [= Notholaena trichomanoides (L.) Desv.]. L’historique des typifications et affectations successives et des différents points de vue a été bien résumé par Yatskievych & Smith (2003). Dans le cadre de la révision des Pteridaceae, Rothfels et al. (2008) ont confirmé la restriction des taxons du Nouveau Monde au genre Notholaena R. Br. Ainsi, pour l’Ancien Monde, Notholaena marantae (L.) Desv. a été transféré dans le genre Paragymnopteris K.H. Shing, 1994 (cf. aussi Schuettpelz et al., 2007 ; Zhang & Ranker, 2013) et est devenu Paragymnopteris marantae (L.) K.H. Shing, 1994 [subsp. marantae] (photo 20).
Références
Pour Oeosporangium
Applequist W.L., 2019. Report of the Nomenclature Committee for Vascular Plants: 70. Taxon 68 (4) : 850-851.
Arana M.D. & Moreno Saiz J.C., 2020. New combinations and typification of Mediterranean Oeosporangium (Pteridaceae: Cheilanthoideae). Phytotaxa 433 (3) : 190-194.
Christenhusz M.J.M., 2012. Pteridaceae. In W. Greuter & E. von Raab-Straube (eds.), Euro+Med Notulae, 6 [Notulae ad floram euro-mediterraneam pertinentes 29], Willdenowia 42 : 283-285.
Fraser-Jenkins C.R., Gandhi K.N., Kholia B.S. & Benniamin A., 2017a. An annotated checklist of Indian pteridophytes, I (Lycopodiaceae to Thelypteridaceae). Dehra Dun, Inde, 572 p. [Oeosporangium : 253-266]
Fraser-Jenkins C.R., Smith A.R. & Windham M.D., 2017b. (2509) Proposal to reject the name Allosorus (Pteridaceae). Taxon 66 (2) : 517-518.
Lellinger D.B., 1993. Report of the Committee of Pteridophyta, 13, (822) Pteridaceae, (867) Cheilanthes. Taxon 42 (1) : 119-120.
Panigrahi G., 1987. Proposal to delete Allosorus Bernhardi, nom. rej., and to emend typification of Cheilanthes Swartz, nom. cons. (Pteridophyta). Taxon 36 (1) : 167-168.
Rumsey F.J., 2014. Some name changes in the Pteridaceae of Macaronesia and Europe – with validation of hybrid names in the genus Allosorus Bernh. Fern Gazette 19 (7) : 275-279.
Sáez L. & Aymerich P., 2017. New nomenclatural combinations in vascular plants. Orsis 31 : 31-35.
Visiani R. (de), 1867. Adunanza del giorno 25 marzo 1867 – Il m.e. Roberto de Visiani comunica una illustrazione della Cheilanthes Szovitsii T. et M. – Gen. Oeosporangium, Vis. Atti del Reale Istituto Veneto di Scienze, Lettere ed Arti III (12) : 656-665, tav. X, XI.
Pour Paragymnopteris
Brown R., 1810. Prodromus Florae Novae Hollandiae et Insulae Van-Diemen, 1. Richard Taylor & Sons, London.
Pichi Sermolli R.E.G., 1977. Tentamen pteridophytorum genera in taxonomicum ordinem redigendi. Webbia 31 (2) : 313-512.
Pichi Sermolli R.E.G., 1981. Report of the committee for Pteridophyta. Taxon 30 (1) : 161-163.
Pichi Sermolli R.E.G., 1983. Fragmenta Pteridologiae, VIII. Webbia 37 (1) : 111-140.
Pichi Sermolli R.E.G., 1989. Again on the typification of the generic name Notholaena R. Brown. Webbia 43 (2) : 301-310.
Rothfels C.J., Windham M.D., Grusz A.L., Gastony G.J. & Pryer K.M., 2008. Toward a monophyletic Notholaena (Pteridaceae): Resolving patterns of evolutionary convergence in xeric-adapted ferns. Taxon 57 : 712-724.
Schuettpelz E., Schneider H., Huiet L., Windham M.D. & Pryer K.M., 2007. A molecular phylogeny of the fern family Pteridaceae: Assessing overall relationships and the affinities of previously unsampled genera. Molecular Phylogenetics and Evolution 44 : 1172-1185.
Shing K.H., 1993 (“1994”). A new genus, Paragymnopteris Shing, separated from Gymnopteris Bernh. Indian Fern Journal 10 : 226-231.
Smith J., 1875. Historia Filicum: an Exposition of the Nature, Number; and Organography of Ferns. MacMillan & Co., London.
Tryon R.M., 1956. A revision of the American species of Notholaena. Contributions from the Gray Herbarium 179 : 1-106.
Tryon R.M. & Tryon A.F., 1980. Proposal to conserve Notholaena with the type Pteris trichomanoides. Taxon 29 (1) : 160-161.
Yatskievych G. & Smith A.R., 2003. Typification of Notholaena R. Br. (Pteridaceae). Taxon 52 (2) : 331-336.
Zhang G. & Ranker T.A., 2013. Paragymnopteris. In Z. Wu, P.H. Raven & D.Hong (eds.), Flora of China, Beijing (Science Press) & St. Louis (Missouri Botanical Garden Press), 2–3 (Lycopodiaceae through Polypodiaceae) : 235-237.
3.7. Cystopteridaceae
Précédemment classés soit dans la famille des Athyriaceae, soit dans celle des Woodsiaceae ou d’autres, les genres Cystopteris Bernh. et Gymnocarpium Newman, ainsi que le genre asiatique Acystopteris Nakai, appartiennent maintenant à la famille des Cystopteridaceae, récemment décrite (Shmakov, 2001). Ce statut a été confirmé par les travaux de Rothfels (2012) et Rothfels et al. (2013).
Concernant plus particulièrement Cystopteris fragilis (L.) Bernh., cette entité est considérée comme l’un des groupes les plus complexes parmi les Ptéridophytes et requiert une révision complète, actuellement en cours (Tennant, 2017 ; Hanušová et al., 2019 ; Ekrt et al., 2021). Du fait de l’existence de plusieurs cytotypes à la fois chez fragilis (4x, 5x, 6x, 8x), chez C. alpina (4x, 5x, 6x, 8x), chez C. dickieana (4x, 6x) et chez C. diaphana (6x au moins) et du fait de l’absence de taxon diploïde, non identifié à ce jour en Europe, il est préférable de parler, pour l’instant, du « complexe Cystopteris fragilis s.l.» dont les différents membres sont temporairement établis, pour certains, au rang de sous-espèces :
- Cystopteris fragilis (L.) Bernh. subsp. fragilis : comme indiqué ci-dessus, ce taxon regroupe plusieurs cytotypes. Il reste à établir à quel cytotype se rapporte le type de la subsp. fragilis;
- Cystopteris dickieana Sim est considéré maintenant, de façon temporaire, comme une subsp. de C. fragilis [C. fragilis subsp. dickieana (R. Sim) Hyl., 1945]. Le caractère des spores rugueuses (photo 23), utilisé jusqu’à récemment pour distinguer C. dickieana de C. fragilis aux spores échinulées (photo 24), ne semble plus être considéré comme discriminant. En effet, la plante type d’Ecosse (photo 21) apparaît comme une entité distincte des Cystopteris à spores rugueuses et à morphologie du limbe classique des C. fragilis, comme ceux du Massif central français ou des Pyrénées (photo 22). Le statut de ces Cystopteris à spores rugueuses reste à régler (Dyer et al., 2000) ;
- Cystopteris fragilis var. huteri ex Milde est également considéré comme une sous-espèce [C. fragilis subsp. huteri (Hausm. ex Milde) Prada & Salvo, 1995].
Références
Dyer A.F., Parks J.C. & Lindsay S., 2000. Historical review of the uncertain taxonomic status of Cystopteris dickieana R. Sim (Dickie’s Bladder Fern). Edinburgh Journal of Botany 57 (1) : 71-81.
Ekrt L., Košnar J., Rothfels C.J., Hanušova K, Hornych O. & Urfus T., 2021. Cytogenetic, geographical, spore type and plastid haplotype data reveal cryptic patterns of species diversity in the cosmopolitan Cystopteris fragilis complex (Polypodiopsida: Cystopteridaceae). Botanical Journal of the Linnean Society 199 (3) : 728-739.
Hanušová K., Čertner M., Urfus T., Koutecký P., Košnar J., Rothfels C.J., Jarolímová V., Ptáček J. & Ekrt L., 2019. Widespread co-occurrence of multiple ploidy levels in fragile ferns (Cystopteris fragilis complex; Cystopteridaceae) probably stems from similar ecology of cytotypes, their efficient dispersal and inter-ploidy hybridization. Annals of Botany 123 : 845-855.
Rothfels C.J., 2012. Phylogenetics of Cystopteridaceae: Reticulation and Divergence in a Cosmopolitan Fern Family. PhD Thesis, Duke University, Durham, North Carolina, USA, 224 p.
Rothfels C.J., Windham M.D. & Pryer K.M., 2013. A Plastid Phylogeny of the Cosmopolitan Fern Family Cystopteridaceae (Polypodiopsida). Systematic Botany 38 (2) : 295-306.
Shmakov A.I., 2001. Synopsis of the ferns of Russia. Turczaninowia 4 : 36-72.
Tennant D.J., 2017. Cystopteris fragilis subsp. huteri, a taxon new to the British Isles and related taxa in the Cystopteris fragilis complex: Cystopteridaceae (Polypodiopsida). Fern Gazette 20 (6) : 255-266.
3.8. Aspleniaceae
Plusieurs ajustements ont été effectués :
- comme déjà mentionné depuis longtemps (Kramer & Viane, 1990), les genres Ceterach, 1804, et Phyllitis Hill, 1757, sont inclus dans le genre Asplenium L., 1753 (voir aussi Van den Heede et al., 2003 ; Schneider et al., 2004 ; Xu et al., 2020) ;
- forme serpentinicole d’Asplenium adiantum-nigrum (photo 25) : l’étude détaillée des différents travaux de Milde (1856, 1858, 1865, 1867) nous conduit aux conclusions suivantes (étude réalisée en collaboration avec W. Greuter & R. Viane, pers. 2022) :
- Asplenium adiantum-nigrum subsp. silesiacum Milde, Jahresber. Schles. Ges. Vaterl. Cult. 33 : 93, 1856, [nom. inval. (Art. 35.2) en 1856, mais validé (Art. 33.1) en 1858] ;
- Asplenium silesiacum Milde, Actorum Acad. Caes. Leop.-Carol. Nat. Cur. 26 (2) : 605, t. 45, fig. 114, 1858, [nom. inval. (Art. 36.1), nom provisoire non accepté par l’auteur (cf. p. 608)] ;
et
-
- Asplenium adiantum-nigrum silesiacum Milde, Nov. Actorum Acad. Caes. Leop.-Carol. Nat. Cur. 26 (2) : 609, 1858, [nom alternatif valide (Art. 36.3), ici considéré comme la validation de la combinaison de Milde de 1856 (Art. 33.1 et 35.2, Ex. 5)] ;
- Asplenium adiantum-nigrum onopteris (L.) Heufl. var. silesiacum (Milde) Milde, Sporenpfl. 28, 1865.
À première lecture, le statut subspécifique du nom onopteris n’apparaît pas clair à cause d’erreurs typographiques dans le format de sa citation. Cependant, l’analyse approfondie du travail de Milde montre que le classement infraspécifique se révèle en fait clair et cohérent : en effet, les noms des sous-espèces sont précédés d’un sous-titre centré et les épithètes sont écrites en lettres espacées et précédées par une lettre capitale. Les épithètes variétales sont également écrites en lettres espacées, mais précédées par un chiffre arabe. L’apparente confusion dans le texte de Milde résulte d’une double erreur typographique :
1. en page 27, le sous-titre “3. Subspecies” a été placé en-dessous du paragraphe commençant par “C. Onopteris v. Heufler l.c.” alors qu’il aurait dû être placé avant ;
2. en conséquence, p. 28, un nouveau sous-titre “4. Subspecies” a été inséré ; celui-ci n’ayant aucune raison d’être doit être supprimé.
En considérant ces deux corrections, le classement reprend son ordre normal et l’indication “2. Silesiacum” correspond à la publication de ce nom comme variété, classée dans “3. Subspecies” “Onopteris v. Heufler l.c.”. Ces erreurs ont été corrigées par la suite dans Milde (1867).
- Asplenium adiantum-nigrum [subsp. onopteris] silesiacum (Milde) Milde, Fil. Eur. 88, 1867, [comb. superfl., séquence recopiée d’après Milde, Sporenpfl. 1865] ;
- Asplenium adiantum-nigrum var. silesiacum (Milde) Viane & Reichst., Candollea 49 (1) : 283, 1994, [comb. superfl., identique à celle de Milde (1865)] ;
- Asplenium adiantum-nigrum silesiacum (Milde) Fraser-Jenk. & Parris, Indian J. For. 43 : 201, 2021, [comb. superfl., identique à celle de Milde (1858)].
En conclusion, la forme serpentinicole d’Asplenium adiantum-nigrum doit être nommée Asplenium adiantum-nigrum var. silesiacum (Milde) Milde, Sporenpfl. 28, 1865 [basionyme : Asplenium adiantum-nigrum L. subsp. silesiacum Milde, Nov. Actorum Acad. Caes. Leop.-Carol. Nat. Cur. 26 (2) : 609, 1858]. Le statut nomenclatural de cette forme et ses noms d’auteurs, restés incertains jusqu’au début des années 2000, ont pu être établis en 2006 avec l’aide de F. Tronchet (MNHN, Paris) et de R. Viane (données non publiées à l’époque), et ont été officialisés dans notre ouvrage (Prelli & Boudrie, 2021 : 497). Compte tenu de l’analyse effectuée en 2022, deux corrections sont à effectuer dans le référentiel taxonomique de notre ouvrage, p. 497 :
- Asplenium adiantum-nigrum var. silesiacum (Milde) Milde, 1865 (et non 1867),
- basionyme : Asplenium adiantum-nigrum subsp. silesiacum Milde, 1858 (et non Asplenium silesiacum Milde, 1859).
Les noms d’auteurs corrects d’A. obovatum billotii sont (F.W. Schultz) O. Bolòs, Vigo, Masalles & Ninot, 1990 (et non pas 1992 comme indiqué par erreur in Prelli & Boudrie, 2021 : 498). La combinaison établie par M. Kerguélen en 1998 est superflue car celle de Bolos et al. (1990 : 1213) est bien validement publiée et complète avec le basionyme et sa référence. On ne sait pour quelle raison M. Kerguélen a cru bon d’établir une nouvelle combinaison qui, de ce fait, est un isonyme postérieur à ignorer (Art. 6.3, Note 2 du code de nomenclature, Turland et al., 2019 ; comm. pers. W. Greuter, 2020).
Le lectotype d’Asplenium foreziense Legrand ex Magnier a été désigné et les noms d’auteurs corrects confirmés (Boudrie & Viane, 2012).
Les lectotypes, respectivement, d’Asplenium fontanum jahandiezii Litard. [= Asplenium jahandiezii (Litard.) Rouy] et d’Asplenium trichomanes sublusus pachyrachis Christ [= A. trichomanes subsp. pachyrachis (Christ) Lovis & Reichst.] ont été désignés (Boudrie & Viane, 2021).
Parmi les hybrides :
- le statut des hybrides d’Asplenium ×pagesii, 1910 (A. foreziense × A. trichomanes) a été réglé par l’établissement de deux nothosubsp. selon la subsp. d’A. trichomanes impliquée dans la combinaison hybride (Boudrie et al. 2003) :
- Asplenium ×pagesii nothosubsp. guichardii (Litard.) Viane, Boudrie, Rasbach & K. Rasbach, 2003 (photo 26) (A. foreziense × A. trichomanes subsp. trichomanes), triploïde ;
- Asplenium ×pagesii nothosubsp. pagesii (photo 27) (A. foreziense x A. trichomanes subsp. quadrivalens), tétraploïde.
- Asplenium ×aran-tohanum Alejandre & M.J. Escal., 2005 ( obovatum subsp. billotii × A. trichomanes subsp. quadrivalens) a été découvert en 2005 en France (Chagneau, 2009).
Références
Bolòs O. (de), Vigo J., Masalles R.M. & Ninot J.M., 1990. Flora Manual dels Països Catalans. Ed. Pòrtic s.a., Barcelona, 1 233 p.
Boudrie M., Rasbach H., Rasbach K. & Viane R., 2003. Nouvelles données cytologiques et nomenclaturales sur les hybrides entre les fougères Asplenium foreziense et Asplenium trichomanes : Asplenium ×guichardii Litard. et Asplenium ×pagesii Litard. (Aspleniaceae, Pteridophyta). Acta Botanica Gallica 150 (2) : 195-211.
Boudrie M. & Viane R.L.L., 2012. Lectotypification, orthographe et auteurs d’Asplenium foreziense Legrand ex Magnier (Aspleniaceae, Pteridophyta). Actes du colloque Les Fougères d’Alsace, d’Europe et du Monde, Strasbourg, 3-4 octobre 2009. Société botanique d’Alsace : 113-128.
Boudrie M. & Viane R., 2021. Notes nomenclaturales relatives aux Aspleniaceae (Pteridophyta), II. Journal de botanique de la Société botanique de France 96 : 2-7.
Chagneau D., 2009. Un nouvel hybride pour la ptéridoflore française découvert à Mauves-sur-Loire (Loire-Atlantique). E.R.I.C.A. 22 : 33-36.
Fraser-Jenkins C.R. & Parris B., 2021. Pteridophytes of Turkey, A Revised Check-List and their Relationships to Europe and the West Himalaya. Indian Journal of Forestry 43 (3) : 193-214.
Kerguélen M., 1998. Quelques problèmes posés par une révision de la flore de France. E.R.I.C.A. 10 : 7-16.
Kramer K.U. & Viane R.L.L., 1990. Aspleniaceae. In K.U. Kramer & P.S. Green (eds.), Pteridophytes and Gymnosperms, Springer-Verlag, Berlin : 52-56.
Milde J., 1856. Ueber einige neue, in Schlesien beobachtete farne. Jahresbericht der Schlehschen Gesellschaft Vaterländische Kultur 33 : 92-95 (“1855”).
Milde J., 1858. Die Gefäss-Cryptogamen in Schlesien Preussischen und Österreichischen Antheils. Novorum Actorum Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum 26 (2) : 371-767, pl. 31-55 (‘1857’) [Asplenium, pp. 568-609].
Milde J., 1865. Die Höheren Sporenpflanzen Deutschland’s und der Schweiz, 10 – Asplenium Smith [A. adiantum-nigrum], Verlag von Arthur Felix, Leipzig : 20-30.
Milde J., 1867. Filices Europae et Atlantidis, Asiae Minoris et Siberiae, 24 – Asplenium adiantum-nigrum, sumptibus A. Felix, Lipsaie [Leipzig] : 85-88.
Reichstein T., Viane R., Rasbach H. & Schneller J., 1994. Asplenium adiantum-nigrum L. subsp. yuanum (Ching) Viane, Rasbach, Reichstein & Schneller, stat. nov., and the status of A. woronowii Christ (Aspleniaceae, Pteridophyta). Studies in Asplenium for « Flora Iranica« , 6. Candollea 49 (1) : 281-328.
Schneider H., Russell S.J., Cox C.J., Bakker F., Henderson S., Gibby M. & Vogel J.C., 2004. Chloroplast phylogeny of asplenioid ferns based on rbcL and trnL and trnL-F spacer sequences (Polypodiidae, Aspleniaceae) and its implications for the biogeography. Systematic Botany 29 : 260-274.
Van den Heede C.J., Viane R.L.L. & Chase M.W., 2003. Phylogenetic analysis of Asplenium subgenus Ceterach (Pteridophyta: Aspleniaceae) based on plastid and nuclear ribosomal ITS DNA sequences. American Journal of Botany 90 : 481-495.
Xu K.-W., Zhang L., Rothfels C.J., Smith A.R., Viane R., Lorence D., Wood K.R., Chen C.-W., Knapp R., Zhou L., Lu N.T., Zhou X.-M., Wei H.-J., Fan Q., Chen S.-F., Cicuzza D., Gao X.-F., Liao W.-B. & Zhang L.-B., 2020. A global plastid phylogeny of the fern genus Asplenium (Aspleniaceae). Cladistics 36 : 22-71.
3.9. Blechnaceae
La révision récente de la famille des Blechnaceae au niveau mondial (Gasper et al., 2016a, 2016b) a conduit à éclater la famille en trois sous-familles et 24 genres comme suit :
- sous-famille des Woodwardioideae : Anchistea, Lorinseria, Woodwardia;
- sous-famille des Stenochlaenoideae : Salpichlaena, Stenochlaena, Telmatoblechnum;
- sous-famille des Blechnoideae : Austroblechnum, Blechnidium, Blechnopsis, Blechnum, Brainea, Cleistoblechnum, Cranfillia, Diploblechnum, Doodia, Icarus, Lomaria, Lomaridium, Lomariocycas, Neoblechnum, Oceaniopteris, Parablechnum, Sadleria,
Or, le type du genre Blechnum est représenté par Blechnum occidentale L. qui est une espèce néotropicale possédant, notamment, des frondes stériles et fertiles isomorphes (photos 28 et 29).
Ainsi, parmi les 24 genres actuellement établis, seuls Woodwardia et Struthiopteris sont présents en France. En conséquence, le Blechnum spicant européen ne pouvant plus être (selon le concept établi par Gasper et al., loc. cit.) rattaché au genre Blechnum, c’est le nom de genre le plus ancien, validement publié, qui lui est appliqué, soit Struthiopteris Scop., 1754. Blechnum spicant (L.) Roth devient donc Struthiopteris spicant (L.) Weiss, 1770 [var. spicant] (photo 30).
Références
Gasper A.L. (de), de Oliveira Dittrich V.A., Smith A.R. & Salino A., 2016a. A classification for Blechnaceae (Polypodiales: Polypodiopsida): new genera, resurrected names, and combinations. Phytotaxa 275 : 191-227.
Gasper A.L. (de), Almeida T.E., de Oliveira Dittrich V.A., Smith A.R. & Salino A., 2016b. Molecular phylogeny of the fern family Blechnaceae (Polypodiales) with a revised genus-level treatment. Cladistics 33 (4) : 429-446.
3.10. Thelypteridaceae
La révision récente (Kuo et al., 2020) du genre Stegnogramma Blume, 1828, a conduit à restreindre ce genre à des espèces de l’Est asiatique et à séparer les espèces d’Europe et d’Afrique dans le genre Leptogramma J. Sm., 1842. En conséquence, l’espèce présente en Europe et en Macaronésie, connue jusqu’à présent sous le nom de Stegnogramma pozoi (Lag.) K. Iwats. [syn. : Cyclosorus pozoi (Lag.) C.M. Kuo], est transférée dans le genre Leptogramma et devient Leptogramma pozoi (Lag.) Heywood, 1961 (photos 31 à 33). Cette modification est confirmée par le travail récent de mise à jour de la classification des Thelypteridaceae au niveau mondial (Fawcett & Smith, 2021).
Références
Fawcett S. & A.R. Smith, 2021. A Generic Classification of the Thelypteridaceae. Sida, Botanical Miscellany 59, BRIT Press, Fort Worth Botanic Garden, Botanical Research Institute of Texas, USA, I-X, 102 p.
Kuo L.-Y., Chang Y.-H., Huang Y.-H., Testo W., Ebihara A., Rouhan G., Quintanilla L.G., Watkins J.E. Jr., Huang Y.-M. & Li F.-W., 2020. A global phylogeny of Stegnogramma ferns (Thelypteridaceae): generic and sectional revision, historical biogeography and evolution of leaf architecture. Cladistics 36 (2) : 164-183.
3.11. Dryopteridaceae
Les quatre subsp. de Dryopteris affinis présentes en France, établies par C.R. Fraser-Jenkins (1980, 1983, 1986, 1987, 1988, 1996a, 1996b), ont été considérées en 2007 par lui-même comme des espèces ( affinis, D. borreri, D. cambrensis et D. pseudodisjuncta) et, en conséquence, les anciennes variétés ont été considérées comme des sous-espèces (voir aussi, pour informations complémentaires, le travail de Trewren, 2014). Il en est de même pour les combinaisons hybrides du groupe complexa. Sont donc présents en France les taxons suivants composant le complexe « D. affinis aggr. » (pour les synonymies, voir Prelli & Boudrie, 2021 : 502, 503, 504) :
- Dryopteris affinis (Lowe) Fraser-Jenk., 1979, taxon 2x (photo 34)
- subsp. affinis
- subsp. punctata Fraser-Jenk., 2007
- Dryopteris borreri (Newman) Newman ex & Tavel, 1937, taxon 3x (photo 35)
- Dryopteris cambrensis (Fraser-Jenk.) Beitel & W.R. Buck, 1988, taxon 3x (photo 36)
- subsp. cambrensis
- subsp. distans (Viv.) Fraser-Jenk., 2007
- subsp. insubrica (Oberh. & Tavel ex Fraser-Jenk.) Fraser-Jenk., 2007
- subsp. pseudocomplexa Fraser-Jenk., 2007
- Dryopteris pseudodisjuncta (Tavel ex Fraser-Jenk.) Fraser-Jenk., 2007, taxon 3x (photo 37)
Note : L’espèce Dryopteris lacunosa S. Jess., Zenner, Chr. Stark & Bujnoch, 2011, décrite par Jessen et al. (2011) d’après une plante de Suisse, a été mentionnée par ces auteurs dans les départements français de l’Aveyron, du Gard et du Tarn. Or, Fraser-Jenkins et al. (2021 : 83) ont démontré que ce taxon avait été établi à un rang trop élevé et ne correspondait qu’à une forme déjà connue de D. borreri et, de ce fait, était synonyme de ce dernier.
Concernant les hybrides entre D. affinis et D. filix-mas, sont présents en France (à noter que, en France, le groupe D. affinis n’est impliqué qu’avec D. filix-mas dans des combinaisons hybrides) :
- Dryopteris ×complanata Fraser-Jenk., 2007 ( filix-mas × D. pseudodisjuncta)
- Dryopteris ×complexa Fraser-Jenk., 1987 nothosubsp. complexa ( affinis subsp. affinis × D. filix-mas)
- Dryopteris ×convoluta Fraser-Jenk., 2007 ( cambrensis × D. filix-mas)
- convoluta (D. cambrensis subsp. insubrica × D. filix-mas)
- Dryopteris ×critica (Fraser-Jenk.) Fraser-Jenk., 2007 ( borreri × D. filix-mas)
Dryopteris submontana / mindshelkensis (photos 38 et 39) : lors de l’étude, dans les années 1990, d’un spécimen d’Afghanistan de Dryopteris mindshelkensis, espèce jusque-là considérée comme endémique d’Asie centrale, Fraser-Jenkins (1997) a constaté que ce spécimen (et d’autres, par la suite, de plusieurs pays d’Asie centrale ; C.R. Fraser-Jenkins, pers. 2021) correspondait en fait à la même espèce que l’espèce européenne Dryopteris submontana (Fraser-Jenk. & Jermy) Fraser-Jenk., 1977. Étant donné que le nom le plus ancien validement publié est Dryopteris mindshelkensis Pavlov, 1954, décrit d’après une récolte du Kazakhstan (à Mindshelke, monts Karatau), cette épithète a priorité sur submontana.
Un hybride nouveau a été mis en évidence en France :
- Dryopteris ×alpirsbachensis Freigang, Zenner, Bujnoch, S. Jess. & Magauer ( carthusiana × D. remota) a été découvert dans les Pyrénées centrales (Haute-Garonne) en 2012 et décrit ultérieurement (Freigang et al., 2017) d’après un spécimen d’Allemagne. Il a également été découvert récemment dans les monts du Lyonnais (photo 40). Cet hybride est pentaploïde (5x) et probablement apogame.
Références
Fraser-Jenkins C.R., 1977. Three species in the Dryopteris villarii aggr., Pteridophyta, Aspidiaceae. Candollea 32 (2) : 305-319.
Fraser-Jenkins C.R., 1980. Dryopteris affinis: a new treatment for a complex species in the European Pteridophyte flora. Willdenowia 10 : 107-115.
Fraser-Jenkins C.R., 1983. Dryopteris affinis. In J. Dostál, T. Reichstein, C.R. Fraser-Jenkins & K.U. Kramer, Illustrierte Flora von Mittel-Europa, ed. 2, 1 (1) Pteridophyta, Paul Parey, Berlin, Hamburg : 142-148.
Fraser-Jenkins C.R., 1986. A classification of the genus Dryopteris (Pteridophyta, Dryopteridaceae). Bulletin of the British Museum of Natural History (Botany) 14 (3) : 183-218.
Fraser-Jenkins C.R., 1987. Taxonomic and Nomenclatural, notes 5, Dryopteris. In L.N. Derrick, A.C. Jermy & A.M. Paul (eds.), Checklist of European Pteridophytes, Sommerfeltia 6 : x-xiii.
Fraser-Jenkins C.R., 1988. Some comments on the nomenclature of Dryopteris. Indian Fern Journal 5 (1-2) : 69-77.
Fraser-Jenkins C.R., 1996a. Dryopteris affinis subsp. pseudodisjuncta (Oberh. & Tavel ex Fraser-Jenk.) Fraser-Jenk., comb. nov. In B.Jonsell (ed.), Lectotypifications and new combinations for Flora Nordica vol. 1 (Lycopodiaceae-Papaveraceae). Nordic Journal of Botany 16 (1) : 4.
Fraser-Jenkins C.R., 1996b. A reaffirmation of the taxonomic treatment of Dryopteris affinis (Dryopteridaceae: Pteridophyta). Fern Gazette 15 (3) : 77-81.
Fraser-Jenkins C.R., 1997. New Species Syndrome in Indian Pteridology and the Ferns of Nepal. International Book Distributors, Dehra Dun, India : 10-11, 134.
Fraser-Jenkins C.R., 2007. The species and subspecies in the Dryopteris affinis group. Fern Gazette 18 (1) : 1-26.
Fraser-Jenkins C.R., Kholia B.S. & Kandel D.R., 2021 (2020). Recent Changes and Corrections in the Taxonomy of Indian Pteridophytes, part 2. Indian Journal of Forestry 43 (1) : 58-86.
Freigang J., Zenner G., Bujnoch W., Jessen S. & Magauer M., 2017. Dryopteris ×alpirsbachensis, hybr. nov., erster Nachweis der Naturhybride zwischen Dryopteris carthusiana und Dryopteris remota (Dryopteridaceae, Pteridophyta). Kochia 10 : 11-33.
Jessen S., Bujnoch W., Zenner G. & Stark C., 2011. Dryopteris lacunosa, eine neue Art des Dryopteris affinis-Aggregats (Dryopteridaceae, Pteridophyta). Kochia 5 : 9-31.
Павлов Н.В., 1954. Hовые Растения Казахстанской Флоры, IV. Вестник Акад. Наук Казах. ССР. 8 : 129, Рнс. 31. [= Pavlov N.V., 1954. Nouvelles plantes de la flore du Kazakhstan, IV. Vestnik Akad. Nauk Kazak. SSR 8 : 129, f. 31.]
Trewren K., 2014. Some taxa within the Dryopteris affinis complex. British Pteridological Society, special publication 13, 48 p.
Remerciements
Nous exprimons nos sincères remerciements à MM. Jean-Charles Bertier (F-42 Chalmazel-Jeansagnières), Arnaud Bizot (F-08 Hannogne-Saint-Martin), Marc Castera (F-64 Pau), Jean-François Christians (F-69 Rillieux-la-Pape), Christopher R. Fraser-Jenkins (Cascais, Portugal), Roger Golding (Bowness on Solway, Cumbria, Grande-Bretagne), Werner Greuter (Orto Botanico, Herbarium Mediterraneum, Università degli Studi di Palermo, Palermo, Italie), Warren D. Hauk (Denison University, Granville, OH, USA), Éric Mosnier (F-63 Combronde), Benjamin Øllgaard (University of Aarhus, Aarhus, Danemark), Rémy Prelli (F-22 Lamballe), Alexander Shmakov (Altaï State University, Barnaul, Russie), Frédéric Tronchet (ex-MNHN, F-75 Paris) et Ronald Viane (Serskamp, Belgique) pour leur aide et leurs conseils, la transmission de certains documents et l’utilisation de certaines photos.


