
La flore des bords de champs en région méditerranéenne : diversité floristique et réponse aux pratiques agricoles
Title
Field margin flora in the Mediterranean region: floristic diversity and response to agricultural practice
Résumé
Depuis 2012, un réseau de 500 parcelles agricoles est suivi pour mesurer l’impact des pratiques agricoles sur la biodiversité à travers quatre taxons indicateurs, dont la flore des bords de champs. Dans l’ex-région Languedoc-Roussillon, 21 parcelles sont suivies et on fait l’objet en 2017 de deux relevés : l’un par les observateurs habituels du réseau 500 ENI, l’autre par un botaniste. La comparaison des deux types de relevés montrent une sous-estimations de la diversité dans le réseau 500 ENI. Cependant, les modèles visant à expliquer les variations de la diversité et de la composition des communautés montrent que les relevés « observateurs » sont suffisants pour détecter les principaux effets non intentionnels comme l’effet négatif du nombre de traitements herbicides. Au-delà de cette question, cet article décrit la diversité taxonomique et écologique de la végétation des bords de champs dans l’ex-région Languedoc-Roussillon.
Abstract
Since 2012, a network of 500 agricultural fields has been monitored to measure the impact of agricultural practices on biodiversity through four indicator taxa, including field margin flora. In the former Languedoc-Roussillon region, 21 plots are monitored and were surveyed twice in 2017: once by the usual observers of the 500 ENI network, but also by a botanist. The comparison of the two types of surveys shows an underestimation of plant diversity in the 500 ENI network. However, models explaining variations in species richness and community composition showed that the observer surveys are sufficient to detect the main unintended effects, such as the negative effect of the number of herbicide treatments. Beyond this question, this article describes the taxonomic and ecological diversity of field margins vegetation in the former Languedoc-Roussillon region.
1. Introduction
Depuis 2012, le ministère en charge de l’agriculture a mis en place un suivi de la biodiversité des agroécosystèmes pour mesurer l’impact des pratiques agricoles. À l’origine de ce dispositif, l’article L251-1 du Code rural qui indique que « la surveillance biologique du territoire doit suivre l’apparition éventuelle d’effets non intentionnels des pratiques agricoles sur l’environnement ». Pour cela, un réseau de cinq cents parcelles cultivées a été mis en place sur l’ensemble de la France métropolitaine (Corse comprise) avec l’appui scientifique du Muséum national d’histoire naturelle (MNHN), de l’Agence nationale de sécurité sanitaire (Anses), de l’Institut national de recherche pour l’agriculture, l’alimentation et l’environnement (Inrae) et de l’Université de Rennes, et le relais local d’organisations professionnelles agricoles (chambres d’agricultures, Fredon) (Andrade et al., 2021). Sur chaque parcelle, quatre taxons font l’objet d’observations : les vers de terre, échantillonnés à l’intérieur des parcelles, la flore et les coléoptères, observés dans la bordure extérieure des champs, et les oiseaux, à l’échelle du paysage.
Afin que le suivi soit réalisable par des non-spécialistes, des protocoles simplifiés ont été mis au point : les vers de terres sont classés dans quatre groupes fonctionnels, les coléoptères sont regroupés dans quatorze morphogroupes (correspondant aux principales familles de Coléoptères) et une liste fermée de cent-cinquante espèces végétales et trente-cinq oiseaux sont suivis. Les observateurs bénéficient de formations régulières et ont à disposition des guides d’identification simplifiés. Cependant, pour la flore, les observateurs font en réalité les relevés les plus exhaustifs possible, mais avec un certain nombre de lacunes. Il est alors légitime de se demander quelles sont les conséquences de ces lacunes sur l’estimation de la diversité floristique et la capacité de détecter des effets non intentionnels (ENI).
Dans la suite de l’article, nous nous intéresserons plus spécifiquement à la flore des bordures de champ à l’échelle de l’ex-région Languedoc-Roussillon qui a fait l’objet d’une analyse détaillée en 2017 avec des relevés supplémentaires effectués par l’un d’entre nous (GF). À l’échelle de la France métropolitaine, une première étude des données « 500 ENI » a montré une réponse de la flore des bords de champs à la diversité du paysage et à la nature des éléments voisins (présence de haies, boisements ou fossés), à la fréquence de fauche de la bordure, mais aussi à l’intensité de la fertilisation azotée et des traitements herbicides dans la parcelle (Fried et al., 2018). Si la flore des bordures de champ est largement étudiée dans le nord de l’Europe depuis plus de trente ans (Alignier & Baudry, 2015 ; Kleijn & Snoeijing, 1997 ; Marshall, 1989 ; Marshall & Moonen, 2002 ; Tarmi et al., 2009), les connaissances sur les communautés de bords de champ en région méditerranéenne sont bien plus réduites (Bassa et al., 2011, 2012 ; Cirujeda et al., 2019).
L’objectif de cette étude était donc triple : i) décrire la diversité de la flore des bords de champ en région méditerranéenne, ii) mesurer les biais d’observations du réseau 500 ENI en comparant la diversité estimée par les observateurs du réseau et la diversité estimée par un botaniste, iii) analyser la composition et la diversité des communautés en fonction de quelques variables agricoles et paysagères.
2. Matériels et méthodes
2.1. Sites d’étude
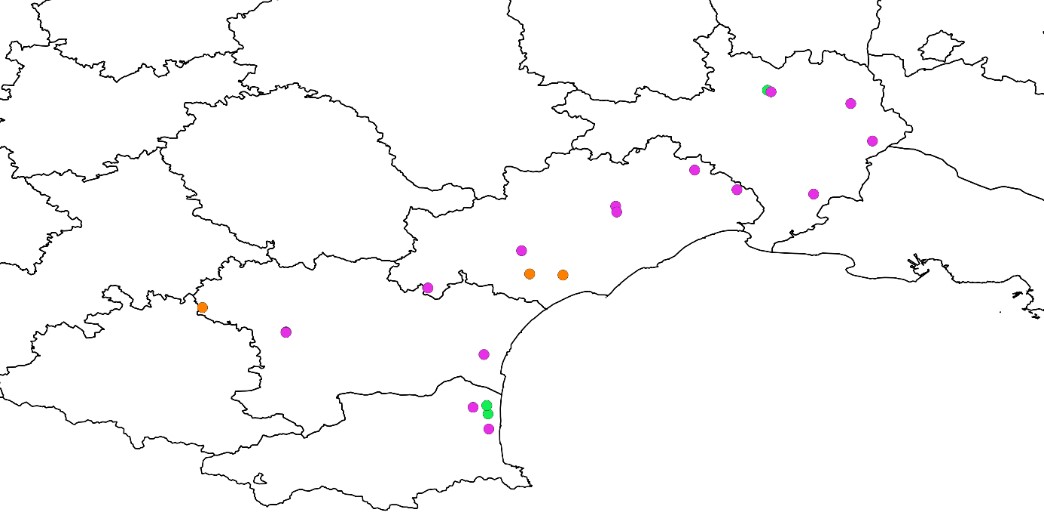
À l’échelle nationale, les cinq cents parcelles suivies tous les ans depuis 2012 se répartissent en environ 350 parcelles de cultures céréalières (blé, maïs), 100 parcelles de vignes et 50 parcelles de cultures maraîchères (Andrade et al., 2021). Dans chaque région, les parcelles ont été choisies pour être représentatives des principaux systèmes de cultures. Dans l’ex-région Languedoc-Roussillon, vingt-et-une parcelles sont suivies dont quinze sont des vignes, trois sont des parcelles de maraîchages et trois sont des céréales (tête de rotation maïs).
Ces vingt-et-une parcelles se distribuent des Pyrénées-Orientales (quatre parcelles) au Gard (cinq parcelles), en passant par l’Aude (quatre parcelles) et l’Hérault (huit parcelles) et couvrent cinq régions naturelles telles que définies dans la Flore de la France méditerranéenne continentale (Tison et al., 2014) (carte 1) : Littoral-sud (cinq parcelles), Lauragais (trois parcelles), Littoral-nord (trois parcelles), Languedoc (huit parcelles) et Rhône (deux parcelles).
2.2. Méthode d’échantillonnage de la flore
Pour chacune des vingt-et-une parcelles, nous avons relevé la flore de trois zones (figure 1) : la bordure du champ suivie par le réseau 500 ENI (notée B), l’intérieur de la parcelle dont la bordure est suivie (noté P) et la bordure du champ voisin le plus proche (notée Bis, si possible en face de la bordure B).
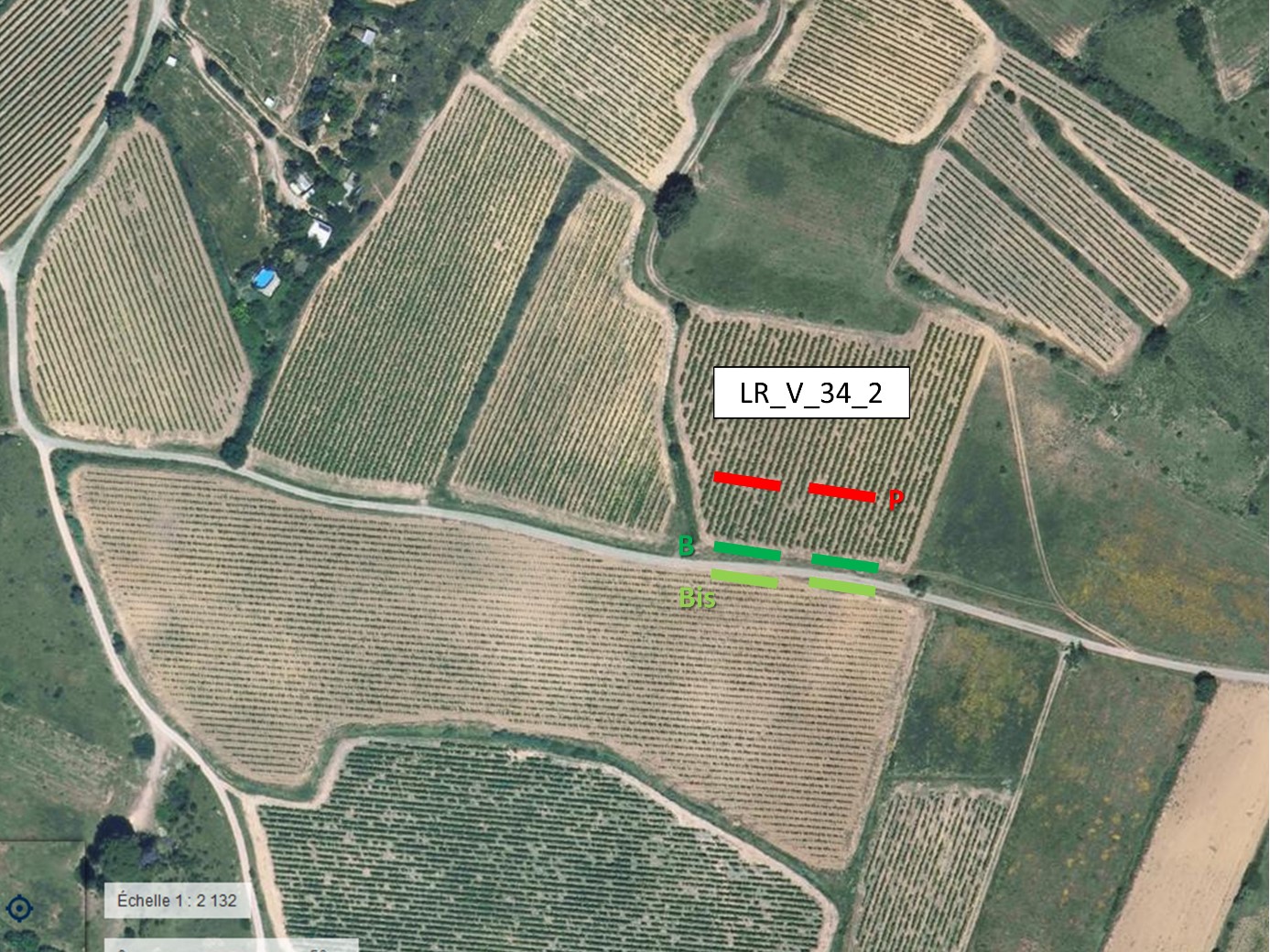
Le relevé floristique dans la bordure est réalisé dans dix quadrats de 1 m2 (un quadrat mesure 2 m de long sur 0,5 m de large ; photo 1) répartis en deux séries de cinq quadrats contigus, les deux séries étant distantes l’une de l’autre d’au moins 30 m quand cela est possible. Seule la présence-absence des taxons dans le quadrat est noté, ce qui conduit pour chaque taxon présent à une note d’abondance de 1 à 10 pour l’ensemble de la bordure. Les relevés effectués par Guillaume Fried entre le 18 avril et le 29 mai 2017 seront appelés « relevés botaniste » par la suite. Les transects B (bordure de référence du réseau 500 ENI) ont par ailleurs été visités par les observateurs de référence du réseau sur la même période ; ils seront par la suite appelés « relevé observateurs ».

2.3. Analyses des données
Pool régional d’espèces
Dans un premier temps, nous avons comparé la liste des taxons observés dans les bordures (B, Bis) et dans les parcelles (P). Puis nous avons réalisé une synthèse des caractéristiques du pool d’espèces des bordures de champ : principales familles, chorologie, statut de fréquence régionale (d’après Tison et al., 2014), spectre biologique (Julve, 1998) et habitats d’origine. Pour ce dernier point, nous nous sommes basés sur les habitats préférentiels indiqués dans la Flore de la France méditerranéenne continentale (Tison et al., 2014).
Diversité locale
Après une description du nombre moyen d’espèces par quadrat (1 m²) et par bordure (10 m²), nous avons comparé la richesse spécifique entre les communautés des bords de champ (B) et les communautés de la parcelle suivie (P) à l’aide d’un test de Student apparié (les données suivant la loi normale et les variances étant égales entre les échantillons comparés). Nous avons de la même manière comparé la richesse spécifique entre le bord de champ de référence (B) et la bordure voisine (Bis). Par ailleurs, nous avons testé l’existence d’une corrélation entre la richesse des bordures (B) et de la parcelle suivie (P), ainsi qu’entre la richesse de la bordure de référence (B) et de la bordure voisine (Bis). Une corrélation de la richesse spécifique entre parcelle et bordure pourrait indiquer une réponse similaire au niveau d’intensité des pratiques agricoles appliquées dans la parcelle, tandis qu’une corrélation de la richesse entre les bordures de deux parcelles différentes (donc gérées différemment) révèlerait plutôt l’effet de la diversité du pool local d’espèces, influencée par les conditions abiotiques locales et/ou la diversité du paysage.
Partitionnement de la diversité
Afin de mieux évaluer l’apport de la flore des bords de champs à la diversité locale et régionale, nous avons suivi l’approche développée par Lande (1996) qui partitionne la diversité totale (γ) entre la diversité à l’intérieur des communautés (α) et de la diversité entre communautés (β). Cela conduit à un modèle de partitionnement additif de la diversité définissant la diversité β comme la différence entre la diversité γ et la diversité α : β = γ – α. Le partitionnement de la diversité fournit la structure avec laquelle la diversité peut être mesurée à différents niveaux d’organisation emboîtés. Pour effectuer ces analyses, nous avons distingué six niveaux d’organisation : (1) le quadrat de 1 m² dans la parcelle cultivée, (2) la parcelle cultivée (Pc) qui regroupe les dix quadrats de 1 m² de la parcelle ; (3) la parcelle étendue (Pe) qui regroupe les relevés de la parcelle (P) et de sa bordure (B) ; (4) enfin, le site local qui regroupe la parcelle étendue à sa bordure (P+B) et le bord de champ voisin (Bis) ; (5) le cinquième niveau correspond au regroupement des sites par région naturelle ; (6) le sixième et dernier niveau correspond à la diversité γ régionale à l’échelle de l’ensemble du réseau de l’ex-région Languedoc-Roussillon. Le partitionnement de la diversité égale : γ = α + βPc+ βPe+ βSite + βrég.
Variations de la richesse spécifique
Pour comprendre l’influence des pratiques agricoles et de l’environnement sur la richesse spécifique des communautés de bords de champ, nous avons utilisé un modèle de régression linéaire mixte. Dans cette analyse, l’unité d’échantillonnage considérée est le bloc de cinq quadrats de 1 m² ; comme il y a deux blocs de cinq quadrats par bordure (séparés par 30 m), nous avons paramétré comme facteur aléatoire l’identité de la parcelle pour tenir compte de la structure emboîtée de l’échantillonnage (deux blocs par parcelle). Avec 42 échantillons, en respectant la règle de dix échantillons pour analyser l’effet d’une variable explicative, nous nous sommes limités à l’analyse de quatre variables : i) le nombre de traitements herbicides dans la parcelle qui représente le niveau d’intensification, ii) la nature de la culture (vigne versus culture annuelle) qui synthétise un ensemble de pratiques distinctes, iii) la largeur de la bordure du champ suivi, iv) la diversité du paysage dans un rayon de 500 m autour de la parcelle, basée sur le calcul de l’indice de diversité de Shannon appliqué aux différents types d’occupation du sol (boisements, prairies, cultures, batis, routes, surface en eaux, etc.). Afin de mesurer un éventuel effet observateur sur la capacité à détecter des effets non intentionnels, nous avons comparé un modèle basé sur la richesse spécifique estimée par un botaniste à un modèle basé sur la richesse spécifique estimée par les observateurs du réseau 500 ENI. Dans le modèle « observateurs », en plus de l’identité de la parcelle, l’identité de l’observateur est également définie comme un facteur aléatoire.
Composition des communautés
Afin d’identifier les déterminants de la composition des communautés, nous avons réalisé plusieurs analyses multivariées. Dans un premier temps, une analyse des correspondances redressée (detrended corresponce analysis, DCA) a été effectuée sur l’ensemble du jeu de données, soit 63 relevés, incluant 21 parcelles (P) et 42 bordures (B et Bis). Dans un second temps, une DCA a été réalisée sur le jeu de données ne contenant que les relevés effectués dans les bordures du réseau 500 ENI (21 bordures B). Les axes de la DCA ont été interprétés en projetant a posteriori les variables explicatives sur la carte des relevés. La seconde DCA, réalisée sur les seules bordures 500 ENI, visait à comparer les données « botaniste » et « observateurs » pour évaluer dans quelle mesure la discrimination des sites selon leur composition était identique ou non le long des premiers axes multivariés.
Dans un second temps, afin de relier directement la composition des communautés aux variables agronomiques et environnementales disponibles, nous avons effectué une analyse des redondances (redundancy analysis, RDA). Nous avons utilisé neuf variables explicatives : la température moyenne annuelle (MAT), le niveau de précipitation annuel (AP), la richesse du sol en azote (Azote), le pH du sol (pH), la largeur de la bordure herbacée (Largeur), le nombre de traitement herbicides (N_Trait_Chi), l’indice de diversité de Shannon du paysage (SHDI), le type de culture (vigne, culture annuelle), le type d’habitat adjacent (Culture, Vigne, Fossé, Haie, Friche_Garrigue, Route, Chemin_Talus).
3. Résultats
3.1. Description du pool d’espèces régional
Sur l’ensemble des vingt-et-un sites et des trois zones échantillonnées (soit 604 quadrats de 1 m²), 293 taxons ont été identifiés, dont 278 sont présents dans les bordures de champ. Cent cinquante-neuf espèces (54%) n’ont été observées que dans les bordures, 119 (41%) à la fois dans les bordures et dans les parcelles et seules quinze espèces (5%) n’ont été trouvées que dans les parcelles sans être observées dans les bordures.
Les familles les plus représentées dans les bordures sont les Asteraceae (48 taxons), les Poaceae (35 taxons) et les Fabaceae (31 taxons), suivies des Brassicaceae (11 taxons), des Apiaceae, des Caryophyllaceae et des Rubiaceae (9 taxons). Le tableau 1 liste les vingt taxons les plus répandus dans les 42 bordures (B + Bis) et donne leur fréquence d’occurrence et leur abondance moyenne (nombre moyen de présence dans les 10 quadrats d’une bordure).
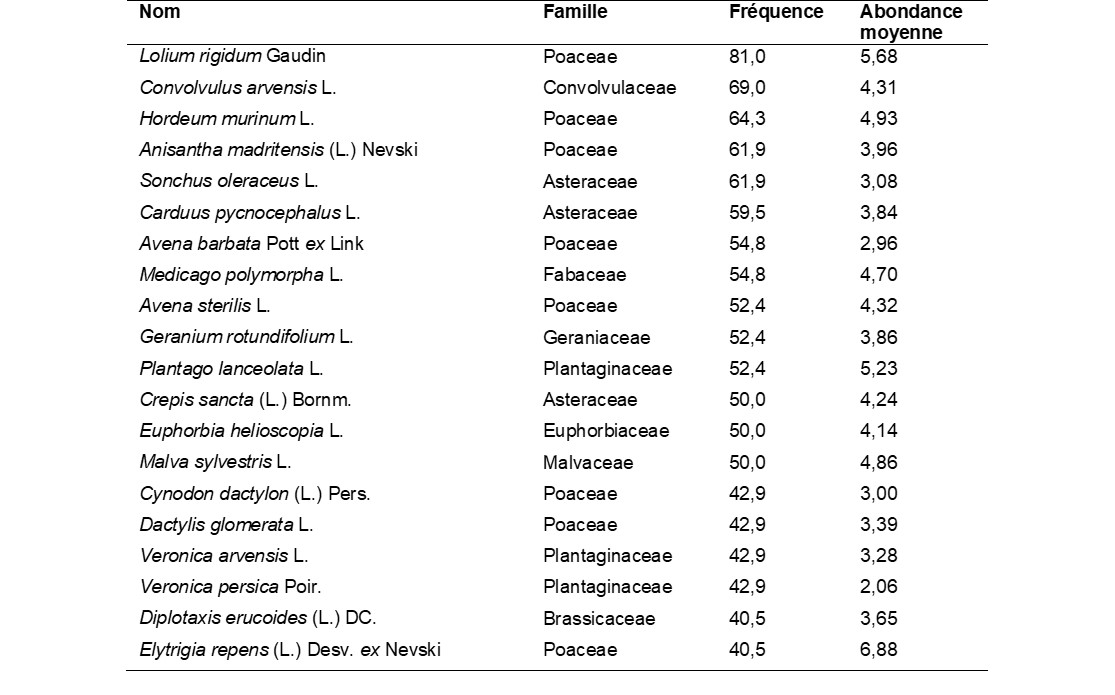
La chorologie du pool d’espèces des bordures de champ est principalement méditerranéenne (21+8+7 = 36%), eurasiatique et européenne méridionale (8+12 = 20%), et dans une moindre mesure holarctique (14%), eurasiatique (12%) et européenne tempérée (8%) (figure 2). La proportion d’espèces exotiques est faible dans ce milieu (3%).
La plupart des espèces des bordures de champ sont des espèces rudérales assez banales (tableau 1). Selon les statuts de la Flore de la France méditerranéenne continentale (Tison et al., 2014), 74 espèces sont très communes (CC), 117 sont communes (C), mais nous avons aussi recensé 37 espèces considérées comme peu fréquentes (PF) et 15 espèces localisées (LO), rares (R) ou très rares (RR) dans la région naturelle correspondante. Il y a donc pas moins d’une espèce sur cinq dans l’ensemble du pool qui présente un certain « intérêt botanique » (figure 3). On peut citer Adonis annua (PF), Cota altissima (PF), Falcaria vulgaris (PF), Fumaria gaillardotii (PF), Hypochaeris glabra (R), Lathyrus annuus (PF), Malva multiflora (R), Tolpis umbellata (PF), Trifolium glomeratum (LO), Trigonella gladiata (PF), Vicia bithynica (R) ou encore Vicia monantha (PF) (voir aussi les espèces illustrées sur la planche 1).

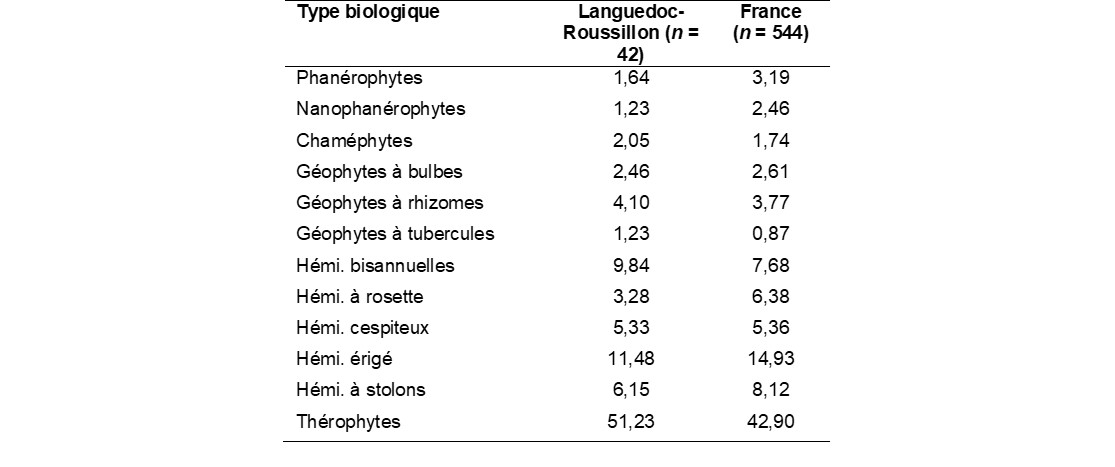
Le spectre biologique montre une prédominance des thérophytes (51%), suivis des hémicryptophytes (36%), des géophytes (8%) et des ligneux (5%). On notera que les bords de champ méditerranéens sont plus riches en espèces annuelles par rapport aux bords de champ de l’ensemble de la France métropolitaine (tableau 2).
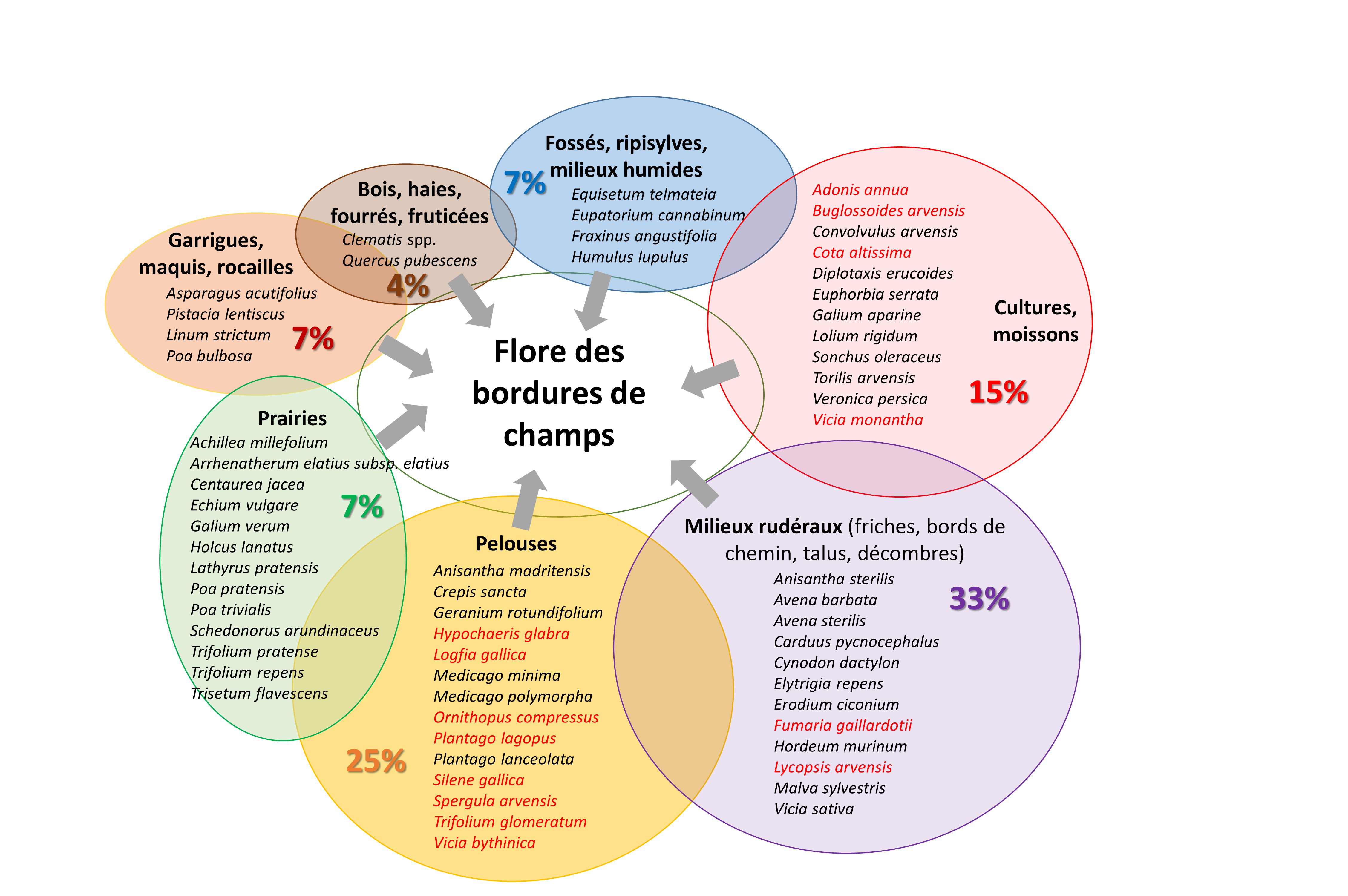
Située à l’interface entre deux parcelles cultivées, entre un champ et un chemin, ou entre un champ et des milieux semi-naturels, la végétation des bords de champ combine des espèces d’origine écologique variée. Bien que directement adjacentes aux bordures, les parcelles cultivées n’apportent que 15% du pool d’espèces (adventices des cultures et messicoles). Le plus grand contingent d’espèces (33%) rassemble des rudérales caractéristiques des friches, des bords de chemin, etc. que l’on trouve souvent aussi en milieu cultivé. Les bords de champs comprennent également beaucoup d’éléments des pelouses xérophiles méditerranéennes (25%) et des garrigues (7%), et dans une moindre mesure un apport des prairies plus mésophiles (7%), des milieux boisés (4%) et des ripisylves (7%) (figure 3).
3.2. Diversité locale et régionale
Variations de la richesse spécifique
À l’échelle des quadrats de 1 m², le nombre d’espèces (richesse spécifique) varie de 3 à 24 avec une moyenne de 11,8 dans les bordures B (parcelles de références 500 ENI). Dans les bordures voisines (Bis) la richesse varie de manière similaire entre 1 et 25 espèces avec une moyenne de 11,1. Dans les parcelles, il n’y a en moyenne que 6,0 espèces par m² avec une variation de 1 à 17 espèces.
À l’échelle des dix quadrats de la bordure, la richesse moyenne est de 31,7 espèces dans les bordures B (parcelles de références 500 ENI) avec un maximum de 51 espèces observées en bordure d’une vigne sur terrain siliceux à Canet-en-Roussillon (figure 4) et un minimum de 14 espèces en bordure d’une culture maraîchère (artichaut) à Torreilles (figure 4). À l’intérieur des parcelles, les dix quadrats totalisent en moyenne 17,1 espèces avec une variation allant de 5 espèces, dans une parcelle de grande culture labourée à Béziers, à 32 dans la même vigne sur terrain siliceux de Canet-en-Roussillon, où la richesse de la bordure est maximale. Dans les bordures voisines, la richesse moyenne est de 33 espèces avec un minimum de 9 espèces en bordure d’une friche à Servas (Gard) et un maximum de 53 espèces à Navacelles (Gard).
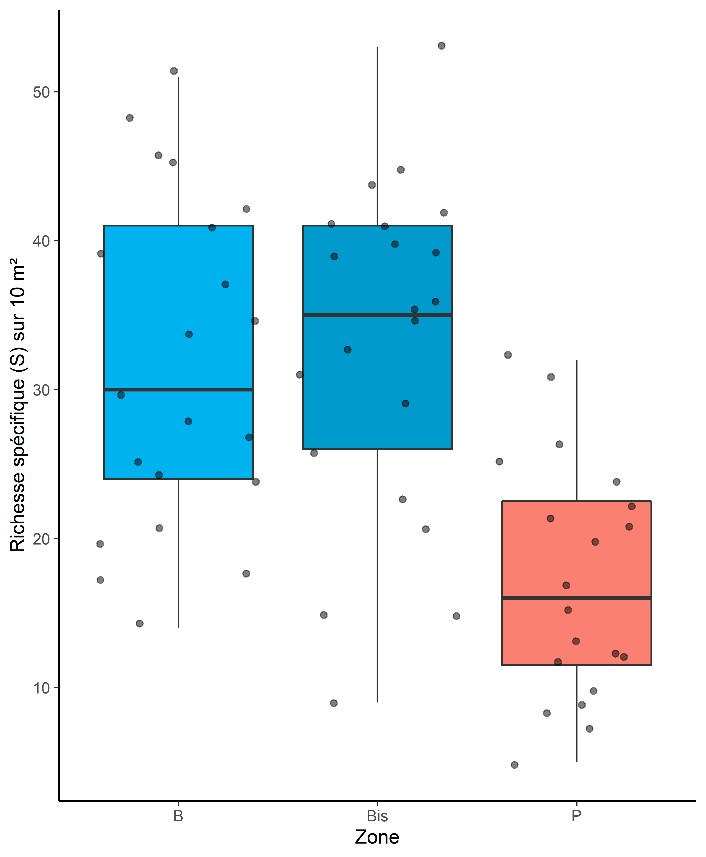
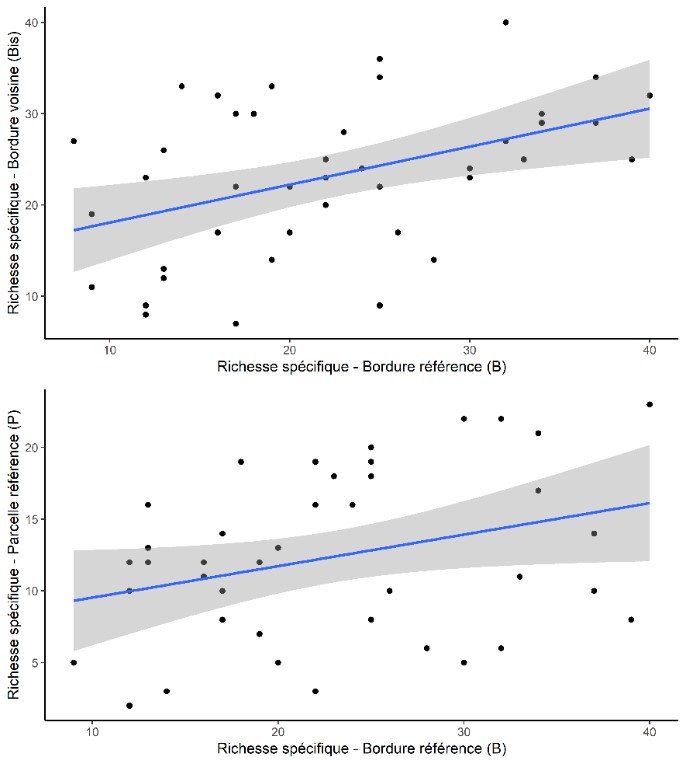
Sans surprise la flore des bordures est significativement plus riche que celle dans les parcelles correspondantes (test-t apparié de Student, t = -6,767, df = 19, P < 0,001 ; figure 4) avec une différence moyenne de 14,1 espèces en plus dans la bordure. Autrement dit, la flore des bordures est en moyenne deux fois plus riche que celle des parcelles. La richesse dans les bordures voisines (Bis) ne montre pas de différence avec la richesse des bordures de référence (test-t apparié de Student, -0,4682, df = 20, P = 0,645 ; figure 4). On trouve une corrélation entre la richesse spécifique de la bordure de référence et celle de la bordure voisine (r = 0,439, P = 0,004 ; figure 5). Dans une moindre mesure, il existe aussi une corrélation entre la richesse dans la bordure de référence et la richesse dans la parcelle (r = 0,319, P = 0,045 ; figure 5).
Partitionnement de la diversité
Avec une richesse moyenne de α = 6,0 espèces, un quadrat de 1 m² à l’intérieur d’une parcelle cultivée ne représente que 11% de la richesse d’un site et 2,1% de la richesse globale du réseau régional (figure 6). L’ajout des neuf autres quadrats de 1 m² dans la même parcelle augmente en moyenne la richesse de βpc =11,3 espèces (soit + 65%), qui atteint 17,3 espèces au niveau d’une parcelle cultivée. Avec l’ajout de la flore du bord de champ (10 quadrats de 1 m²), la richesse moyenne augmente très nettement de βpe = 22,8 espèces (soit + 57%) pour atteindre 40,2 espèces au niveau de la parcelle étendue (soit les dix quadrats de la parcelle (P) et les dix quadrats de la bordure (B)).

En ajoutant la flore du bord de champ voisin, la richesse moyenne du site passe à 54,5, soit un apport moyen de βsite = 14,3 espèces supplémentaires (+ 26%). Les différences entre sites d’une même région naturelle contribuent à un ajout supplémentaire de βrégion = 90,8 espèces (soit + 62%) pour atteindre 145,3 espèces en moyenne par région naturelle. L’apport d’espèces le plus important s’effectue lorsque l’on passe de l’ensemble des parcelles étendues au sein d’une région naturelle à l’ensemble des parcelles étendues du réseau régional avec un ajout de β = 137,7 espèces (soit + 49%) pour atteindre 283 espèces (pool régional).
Comparaison de la diversité estimée par les observateurs et par un botaniste
Dans les 21 bordures des parcelles de références (B), les observateurs du réseau 500 ENI ont recensé 113 espèces tandis que les relevés effectués par le botaniste (G. F.) sur ces mêmes bordures ont permis de détecter un total de 206 espèces, soit + 45%. Soixante quinze espèces étaient communes aux deux, 131 n’ont été trouvées que par le botaniste et 38 que par les observateurs. Parmi les espèces non détectées par les observateurs, 58% sont rares (fréquence < 5%) et 23% sont assez rares (fréquence < 10%). Parmi les 19% d’espèces assez communes (fréquence > 10%) et mal détectées par les observateurs, on trouve des Poacées (Lolium rigidum confondu avec L. perenne, Bromus hordeaceus, Avena sterilis confondue avec A. barbata), des Asteracées (Carduus pycnocephalus, Crepis foetida subsp. rhoeadifolia, Podospermum laciniatum) ou des espèces discrètes (e.g. Arenaria leptoclados). À l’échelle d’une bordure, les observateurs trouvent en moyenne 13 espèces et le botaniste 31,7 espèces, soit + 59%.
3.3. Facteurs expliquant la diversité floristique des bordures
Le modèle basé sur la richesse spécifique des bordures estimée par le botaniste explique 59,4% des variations (R² conditionnel), dont 36,2% expliqués par les variables fixes (R² marginal). Dans ce modèle, la richesse spécifique du bord de champ est influencée par le type de culture de la parcelle et le nombre de traitements herbicides (tableau 3). Un test de comparaison post-hoc montre que le nombre d’espèces est plus élevé en bordure de vignes (25,3 ± 7,9) qu’en bordure de cultures annuelles (15,1 ± 6,8). Une relation négative est trouvée entre le nombre de traitements herbicides dans la parcelle et le nombre d’espèces dans la bordure. Chaque traitement supplémentaire dans la parcelle correspond en moyenne à 3,33 espèces en moins dans la bordure.
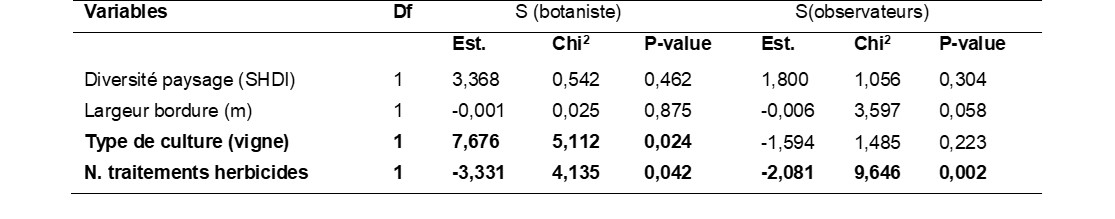
Fait intéressant, même si les observateurs sous-estiment la diversité végétale réelle (-59%), le modèle basé sur leur estimation de la richesse donne des résultas proches du modèle « botaniste » (tableau 3). L’effet le plus fort détecté est une relation négative avec le nombre de traitements herbicides. Comme dans le modèle « botaniste », la diversité du paysage a un effet positif (non significatif) et la largeur de la bordure a un effet quasi nul (non significatif). Seule différence notable, l’absence d’effet du type de culture. Cela peut être mis en lien avec le positionnement des bordures de vigne, souvent en contact avec des friches, pelouses ou garrigues et présentant un plus grand nombre d’espèces méditerranéennes par rapport aux bordures de cultures annuelles (maraîchage, maïs) dont les bordures, en contexte de plaine agricole, sont plus souvent composées d’espèces plus banales, plus facilement identifiées par les observateurs. Il faut toutefois noter qu’il y a un très fort effet « observateur » puisque les variables fixes n’expliquent que 22,7% des variations (R² marginal) alors qu’en prenant en compte les effets aléatoires, dont l’identité de l’observateur, le modèle explique 88,1% des variations (R² conditionnel).
3.4. Composition de la flore des bords de champs
La DCA oppose les relevés sur l’axe 1 selon un double gradient géographique et écologique (figure 7). Les relevés se distribuent par région selon un gradient de continentalité avec le Lauragais et le Rhône dans les coordonnées négatives, le Languedoc au centre et les régions littorales nord et sud dans les cooordonnées positives (figure 7b). Bryonia dioica, Eupatorium cannabinum et Urtica dioica se retrouvent dans les bordures mésophiles des ambiances médioeuropéennes (Lauragais, Rhône), tandis qu’à l’autre extrémité du gradient on trouve des espèces plus xéro-thermophiles comme Plantago lagopus, Centranthus calcitapae ou Phalaris aquatica. L’axe 1 oppose aussi les relevés selon les zones, avec les bordures au centre et l’intérieur des parcelles dans les coordonnées positives (figure 7d). Associée à ce gradient de perturbation, on notera la présence plus significatives d’annuelles (Poa annua, Senecio vulgaris, Capsella bursa-pastoris, Papaver rhoeas) ou de géophytes tolérant le fractionnement (Cirsium arvense, Cynodon dactylon) dans les coordonnées positives, tandis que les hémicryptophytes (Elytrigia repens, Poa pratensis, Potentilla reptans) sont plus nombreuses dans les coordonnées négatives (figure 7a).
L’axe 2 sépare les relevés selon le type de culture (vignes dans les coordonnées positives, cultures annuelles dans les coordonnées négatives ; figure 7d) et le mode de production (agriculture biologique dans les coordonées positives ; figure 7c). On notera que la différence de composition entre les bordures et les parcelles est plus prononcée dans les cultures annuelles que dans les vignes.
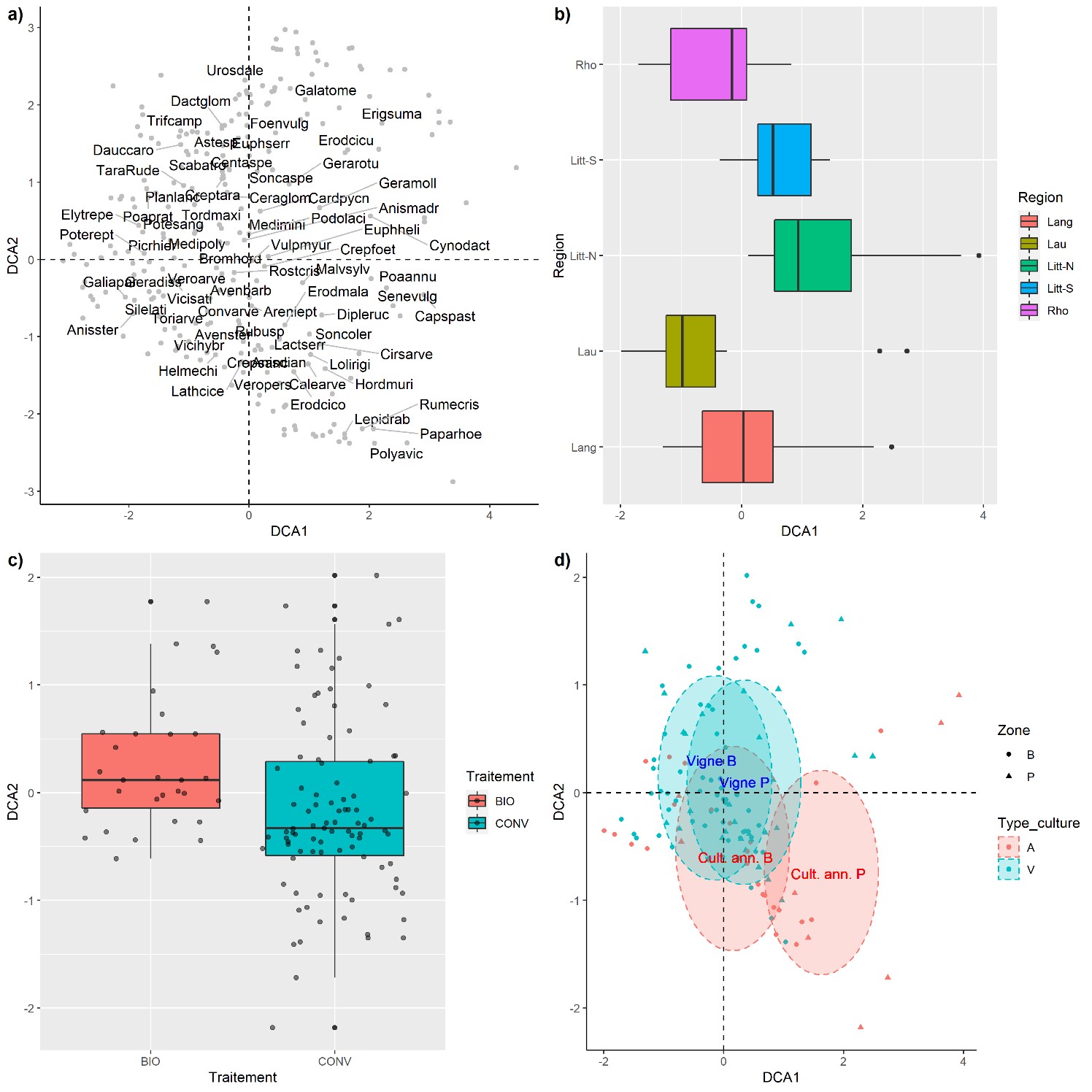
Les deux DCA réalisées sur les bordures 500 ENI avec les données « observateurs » et « botanistes » montrent que les relevés observateurs permettent d’identifier le même gradient principal qu’avec les données botanistes avec une corrélation de r = 0.84 sur le premier axe de la DCA1, mais seulement de r = 0.21 sur le deuxième axe.
La RDA explique 37% des variations dans la composition des bordures de champ (figure 8). L’axe 1 (horizontal, 21,8% de l’inertie) discrimine les parcelles selon un gradient ombrothermique. Les bordures reçevant plus de précipitations (vecteur AP) et avec des températures moyennes annuelles plus faibles (vecteur MAT) sont positionnées dans les coordonnées négatives et hébergent des espèces médio-européennes comme Potentilla reptans, Geranium dissectum ou Poa pratensis. À l’autre extrémité du gradient (températures plus élevées, précipitation plus faible), on trouve des bordures dominées par Galactites tomentosus, Anisantha diandra et Malva sylvestris. La corrélation entre les coordonnées des espèces sur les axes de la RDA et leur valeur indicatrice d’Ellenberg confirme ces résultats. L’axe 1, qui oppose les régions de l’intérieur des terres (Lauragais, vallée du Rhône) aux régions littorales (figure 8c), est corrélé négativement au degré de continentalité des espèces (Ellenberg-C, r = -0,198, P = 0,002 ; tableau 4). Dans une certaine mesure, l’axe 1 oppose également les bordures selon le type d’habitat voisin avec dans les coordonnées négatives des bordures adjacentes à des fossés, talus et chemins dominés par Elytrigia repens, tandis que dans les coordonnées positives les bordures sont voisines de garrigues et de friches, avec des espèces comme Carduus pycnocephalus, Avena barbata ou Foeniculum vulgare. Cela est cohérent avec le fait que cet axe est négativement corrélé au besoin des espèces en humidité édaphique (Ellenberg-H, r = -0,148, P = 0,021 ; tableau 4).
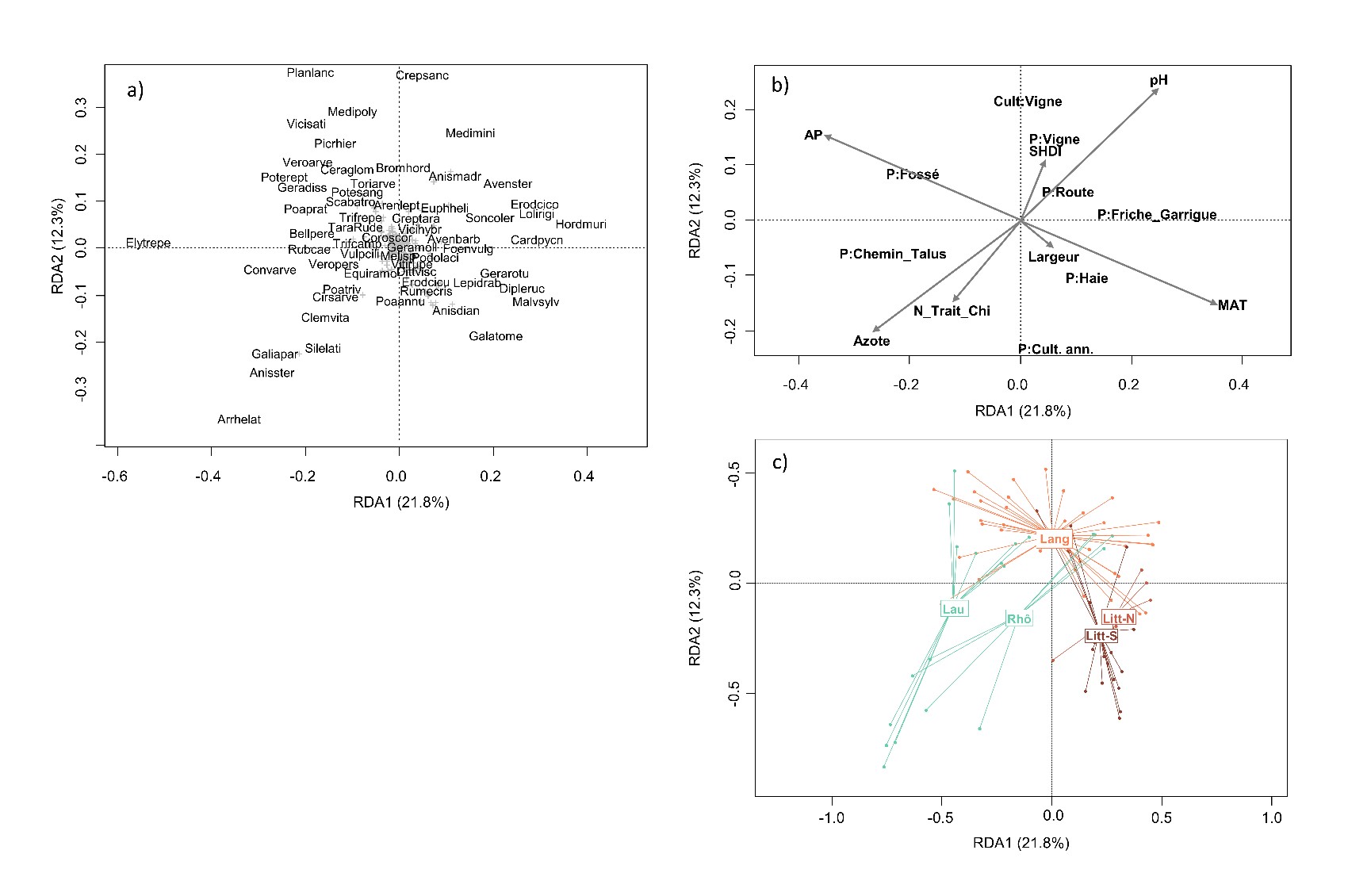
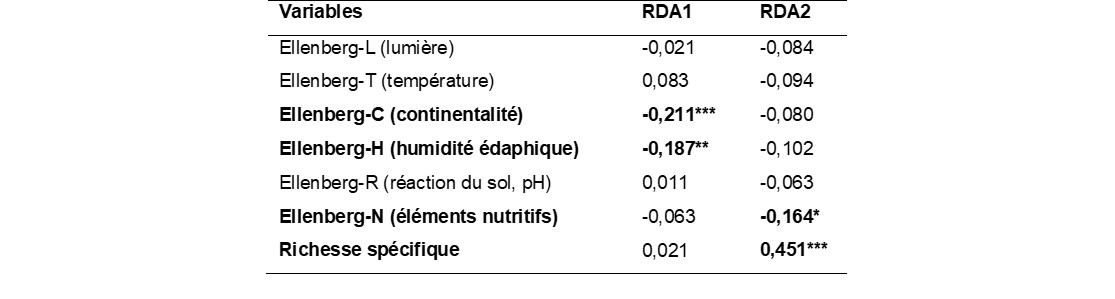
L’axe 2 (12,3% de l’inertie) est davantage lié aux pratiques agricoles. Il oppose les bordures de vigne dans des paysages diversifiés (coordonnées positives) aux bordures de cultures annuelles avec un plus grand nombre de traitements herbicides et des sols plus riches en azote. Les bordures de vigne sont souvent dominées par Crepis sancta, Medicago minima, M. polymorpha, Bromus hordeaceus et Anisantha madritensis. À l’opposé, les bordures de cultures annuelles des plaines agricoles en situation plus intensive sont dominées par Arrhenatherum elatius subsp. elatius, Anisantha sterilis, Galium aparine subsp. aparine et Silene latifolia. Les valeurs indicatrices d’Ellenberg confirment la nature trophique de cet axe 2 avec les espèces nitrophiles qui sont corrélées négativement à l’axe 2 (r = -0,191, P = 0,003).
Les types biologiques ne sont pas distribués aléatoirement le long de l’axe 1 (Kruskal-test, P < 0.001). Les thérophytes et les bisannuelles ont des valeurs plus élevées sur cet axe (coordonnées positives) que les phanérophytes, géophytes et hémicryptophytes. Enfin, la richesse spécifique des communautés n’est pas corrélée à l’axe 1 (r = 0,078, P = 0,479) qui est un axe climatique, mais elle est positivement corrélée à l’axe 2 (r = 0,455, P < 0,001) qui est davantage lié aux pratiques agricoles.
4. Discussion
Les objectifs de cette étude étaient d’une part de mieux cerner la diversité et la composition de la flore des bords de champ en région méditerranéenne et d’évaluer comment ses propriétés sont influencées par les pratiques agricoles et l’environnement (climat, sol, paysage). D’autre part, il s’agissait de mesurer les biais éventuels liés aux relevés réalisés par des observateurs qui ne sont pas des botanistes confirmés.
4.1. Biais observateurs : une sous-estimation de la diversité mais une bonne détection des ENI
Le suivi de la biodiversité est devenu un enjeu majeur de notre époque face aux multiples pressions exercées sur les milieux par les activités humaines. Au même moment, les compétences botaniques et naturalistes connaissent une érosion constante (Crisci et al., 2020). Les chercheurs et les botanistes compétents ne sont pas assez nombreux pour assurer un suivi général de la flore commune et ces derniers sont plus volontiers mobilisés dans le suivi des espèces rares ou complexes à identifier et des milieux patrimoniaux. Une alternative est l’utilisation des sciences participatives, adoptées par le MNHN dans le suivi Vigie-Flore, qui fait appel aux botanistes volontaires pour réaliser le suivi des changements d’abondance des espèces végétales les plus communes de France (Martin et al., 2016). Un des avantages des sciences citoyennes est le recueil à moindre coût d’un grand nombre de données à de larges échelles spatiales et temporelles (Pocock et al., 2014). Dans cette stratégie d’échantillonnage, on fait le pari que la masse de données recueillies contrebalance la moindre précision des données générées par des observateurs non spécialistes.
Le réseau 500 ENI présente certaines caractéristiques d’un dispositif de sciences citoyennes avec l’utilisation de protocoles simplifiés et une grande diversité d’observateurs non spécialistes. Il en présente aussi certains biais. Dans un objectif d’inventaire de la diversité, nos résultats montrent clairement une sous-estimation importante de la diversité réelle avec en moyenne seulement 41% des espèces présentes détectées sur 10 m² de bordure et 55% de la diversité régionale sur vingt-et-une bordures échantillonnées. Cette sous-estimation semble en particulier liée à la richesse des bordures de champ méditerranéens car les différences sont moindres en zone médio-européenne lorsque l’on compare la diversité des bords de champ du réseau 500 ENI et celles estimées par des chercheurs botanistes dans les mêmes zones, par exemple dans le Cher (-5,5%), la Côte-d’Or (-5,8%) ou l’Isère (+1,2%) (Fried et al., 2022). Par ailleurs, on ne peut pas reprocher ce biais aux observateurs qui sont censés savoir reconnaître une liste fermée d’une centaine d’espèces non pas dans un objectif de suivi exhaustif de la diversité, mais dans le but de détecter des ENI. De ce point de vue, nos résultats montrent que les effets majeurs, comme l’impact des traitements herbicides sur la richesse spécifique ou les principaux gradients de composition, sont bien détectés par les observateurs même avec une qualité d’observations moindre.
4.2. Spécificités de la végétation des bords de champ en région méditerranéenne
La flore des bords de champs a jusqu’à présent surtout été étudiée dans le nord de l’Europe. Dans ces régions, l’agriculture a été marquée par une intensification plus forte et les bordures de champ ont de ce fait une grande valeur écologique, car ils constituent parfois les derniers vestiges de milieux semi-naturels à l’échelle de paysages entiers (Marshall & Moonen, 2002). L’importance fonctionnelle des bords de champ semble moins reconnue en région méditerranéenne, d’une part car les champs cultivés méditerranéens sont généralement perçus comme moins intensifs, hébergeant encore une flore messicole disparue ailleurs en Europe (Storkey et al., 2012 ; Munoz et al., 2020), d’autre part les effets de bordure (richesse plus forte en bordure) seraient moins importants (Dutoit et al., 2010) et les paysages environnant les agrosystèmes sont plus riches et favorables au maintien d’espèces annuelles (pelouses xérophiles, garrigues).
Si on compare les espèces les plus fréquentes des bordures languedociennes avec celles d’études espagnoles, on retrouve beaucoup d’espèces en commun. Dans une étude réalisée en Catalogne sur soixante bordures (Bassa et al., 2011), Lolium rigidum et Convolvulus arvensis sont également les deux espèces les plus fréquentes et sept autres espèces sont identiques parmi les vingt espèces les plus fréquentes (Avena sterilis, Hordeum murinum, Plantago lanceolata, Dactylis glomerata, Sonchus oleraceus, Cynodon dactylon, Malva sylvestris). Il y aurait donc une certaine homogénéité au sein de la végétation des bordures des champs méditerranéens.
Avec une richesse moyenne de 31,7 espèces, nos résultats confirment que les bords de champ de l’ex-région Languedoc-Roussillon sont les plus diversifiés du réseau 500 ENI, avec ceux de Corse (où les relevés sont effectués par le Conservatoire botanique national de Corse, S = 32,4) et de l’ex-région Midi-Pyrénées (S = 29,9). Ailleurs en France, la richesse est environ deux fois plus faible avec en moyenne quinze espèces sur 10 m² de bordure (minimum en Champagne-Ardennes, S = 10,7 ; maximum en Limousin, S = 21,3). Le même travail de comparaison entre « relevés observateurs 500 ENI » et « relevés botanistes » resterait à réaliser en région PACA où le niveau de diversité devrait être similaire aux deux autres régions méditerranéennes.
L’analyse du partitionnement de la diversité montre que les bordures contribuent de façon importante à la diversité locale dans les plaines viticoles du Languedoc-Roussillon. Une bordure apporte en moyenne 60% de la diversité à l’échelle d’une parcelle étendue (parcelle et sa bordure). Par ailleurs, à l’échelle régionale, 90% des espèces des parcelles sont aussi présentes en bordure, alors que seulement 43% des espèces des bordures sont aussi présentes en parcelles. Comme ailleurs en Europe, les bordures de champs méditerranéens apportent donc une diversité et une originalité floristiques aux paysages agricoles, à l’origine de leur rôle fonctionnel reconnu (Morrison et al., 2017, Solé-Senan et al., 2018). Ici comme ailleurs, leur conservation est donc importante.
4.3. Effets non intentionnels des pratiques agricoles
Le choix de suivre la végétation des bords de champ est lié à sa proximité immédiate des cultures, ce qui en fait le milieu semi-naturel le plus susceptible de subir des effets dits non intentionnels (ENI) des pratiques agricoles, dont la dérive de pesticides sur des espèces non-cibles des traitements. Nos résultats montrent que les bordures méditerranéennes sont également impactées par les pratiques agricoles avec un effet négatif du nombre de traitements herbicides. Cela confirme un résultat obtenu au niveau national (Fried et al., 2018). La richesse plus élevée en bordures de vigne comparée aux bordures de cultures annuelles (céréales ou cultures maraîchères) pourrait s’expliquer par un ensemble de pratiques moins intensives, autres que les herbicides : la fertilisation minérale est moins importante en vignes, le travail du sol est réduit et il existe de plus en plus de vignes à inter-rang enherbé dont la végétation est dans ce cas proche de celle des bordures (comme le montre dans la DCA la position des bords de vigne et des parcelles de vigne). D’après la RDA, les bordures plus eutrophes, dominées par des espèces agrotolérantes comme le brome stérile et le gaillet gratteron, sont également moins diversifiées, un résultat classique souvent lié à l’abondance accrue de ces espèces nitrophiles dont le développement se fait au dépend d’espèces moins compétitives (Kleijn & Snoeijing, 1997).
Les analyses multivariées (DCA, RDA) montrent que la composition dépend d’abord de la localisation géographique des bordures selon un gradient climatique opposant les bordures franchement méditerranéennes (températures élevées, peu de précipitations) et des bordures plus médio-européennes dans le Lauragais et le Gard rhodanien. Les éléments directement voisins de la bordure (fossés, haies, garrigues) expliquent également une part importante des variations de composition (Blaix & Moonen, 2020). Après les influences climatiques et paysagères, les pratiques agricoles discriminent également la flore, notamment à travers les différences entre vigne (culture pérenne) et cultures annuelles et suivant un gradient d’intensification des pratiques.
5. Conclusion
Le suivi de la flore par le réseau 500 ENI sous-estime la diversité réelle des communautés végétales des bords de champs méditerranéens mais semble être en mesure de détecter les principaux effets non intentionnels des pratiques agricoles. Celles-ci, notamment l’intensité du désherbage chimique, affectent négativement la végétation des bords de champs méditerranéens. Cet habitat linéaire mériterait une meilleure conservation car il contribue fortement à la diversité floristique et aux fonctionnalités associées dans les paysages viticoles. Par ailleurs, bien que les bords de champs méditerranéens n’hébergent pas d’espèces à fort enjeu patrimonial, lorsqu’ils sont en contact avec des milieux ouverts voisins (pelouses xérophiles, notamment silicicoles), ils peuvent héberger des espèces intéressantes (e.g. Trifolium bocconei, T. hirtum… ; photo en fin de texte) qui sont parfois aussi en déclin dans leur habitat principal. Deux orientations sont donc à préconiser : 1) la protection des habitats semi-naturels devrait inclure une réflexion sur l’extension des mesures de protection jusque dans les agrosystèmes voisins, en particulier les bords de champ lorsque cela est pertinent (fonctionnement en méta-population), et 2) les efforts actuels de transition agro-écologique de la viticulture devraient sensibiliser davantage les viticulteurs sur les nombreux rôles fonctionnels des bordures de champ (favoriser les auxiliaires, limiter l’érosion, contribution au maintien des pollinisateurs, etc.) afin de les inciter à les maintenir dans un bon état fonctionnel.

Bibliographie
Aavik T., Augenstein I., Bailey D., Herzog F., Zobel M. & Liira, J., 2008. What is the role of local landscape structure in the vegetation composition of field boundaries? Applied Vegetation Science 11 (3) : 375-386, doi:10.3170/2008-7-18486.
Aavik T. & Liira J., 2009. Agrotolerant and high nature-value species, plant biodiversity indicator groups in agroecosystems. Ecological Indicators 9 (5) : 892-901, doi:http://dx.doi.org/10.1016/j.ecolind.2008.10.006.
Aavik T. & Liira J., 2010. Quantifying the effect of organic farming, field boundary type and landscape structure on the vegetation of field boundaries. Agriculture, Ecosystems & Environment 135 (3) : 178-186.
Alignier A. & Baudry J., 2015. Changes in management practices over time explain most variation in vegetation of field margins in Brittany, France. Agriculture, Ecosystems & Environment 211 : 164-172, doi:http://dx.doi.org/10.1016/j.agee.2015.06.008.
Andrade C., Villers A., Balent G., Bar-Hen A., Chadoeuf J., Cylly D., Cluzeau D., Fried G., Guillocheau S., Pillon O., Porcher E., Tressou J., Yamada O., Lenne N., Jullien J. & Monestiez P., 2021. A real-world implementation of a nationwide, long-term monitoring program to assess the impact of agrochemicals and agricultural practices on biodiversity. Ecology and Evolution 11 (9) : 3771-3793, doi:https://doi.org/10.1002/ece3.6459.
Bassa M., Boutin C., Chamorro L. & Sans F.-X., 2011. Effects of farming management and landscape heterogeneity on plant species composition of Mediterranean field boundaries. Agriculture, Ecosystems & Environment 141 (3–4) : 455-460, doi:http://dx.doi.org/10.1016/j.agee.2011.04.012.
Bassa M., Chamorro L., José-María L., Blanco-Moreno J.M. & Sans, F.-X., 2012. Factors affecting plant species richness in field boundaries in the Mediterranean region. Biodiversity and Conservation 21 (4) : 1101-1114, doi:10.1007/s10531-012-0245-y.
Blaix C. & Moonen A.-C., 2020. Structural field margin characteristics affect the functional traits of herbaceous vegetation. PLOS ONE : 15 (9), e0238916. doi:10.1371/journal.pone.0238916.
Cirujeda A., Pardo G., Marí A. I., Aibar J., Pallavicini Y., González-Andújar J. L. & Solé-Senan X.O., 2019. The structural classification of field boundaries in Mediterranean arable cropping systems allows the prediction of weed abundances in the boundary and in the adjacent crop. Weed Research 59 (4) : 300-311. doi:https://doi.org/10.1111/wre.12366.
Crisci J.V., Katinas L., Apodaca M.J. & Hoch P.C., 2020. The End of Botany. Trends in Plant Science 25 : 1173-1176.
Dutoit Th., Gerbaud É., Buisson É. & Tatoni Th., 2010. Les lisières de champs sont-elles des refuges pour les espèces messicoles dans le territoire du parc naturel régional du Luberon ? Courrier scientifique du Parc naturel régional du Luberon 9 : 58-69.
Fried G., Villers A. & Porcher E., 2018. Assessing non-intended effects of farming practices on field margin vegetation with a functional approach. Agriculture, Ecosystems & Environment 261 : 33-44, doi:https://doi.org/10.1016/j.agee.2018.03.021.
Fried G., Le Corre V., Rakotoson T., Buchmann J., Germain T., Gounon R., Royer H., Biju-Duval L., Felten E., Vieren E. & Chauvel B., 2022. Impact of new management practices on arable and field margin plant communities in sunflower crops, with an emphasis on the abundance of Ambrosia artemisiifolia (Asteraceae). Weed Research (in press)
Julve Ph., 1998 ff. Baseflor. Index botanique, écologique et chorologique de la flore de France. http://perso.wanadoo.fr/philippe.julve/catminat.htm.
Kleijn D. & Snoeijing G.I.J., 1997. Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34 (6): 1413-1425.
Lande R., 1996. Statistics and Partitioning of Species Diversity, and Similarity among Multiple Communities. Oikos 76 (1) : 5-13, doi:10.2307/3545743.
Martin G., Porcher E. & Machon N., 2016. Quels enjeux pour l’implication des botanistes bénévoles dans les suivis participatifs de biodiversité en France ? In N.R. Rakotoarisoa, S. Blackmore & B. Riera (eds), Botanists of the twenty-first century, roles, challenges and opportunities, éditions Unesco, 69-77.
Marshall E., 1989. Distribution patterns of plants associated with arable field edges. Journal of Applied Ecology 26 (1) : 247-257.
Marshall E.J.P. & Moonen A.C., 2002. Field margins in northern Europe: their functions and interactions with agriculture. Agriculture, Ecosystems & Environment 89 (1) : 5-21, doi:http://dx.doi.org/10.1016/S0167-8809(01)00315-2.
Morrison J., Hernández Plaza E., Izquierdo J. & González-Andújar J.L., 2017. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: which biotic and abiotic factors are important? Agriculture, Ecosystems & Environment 247 : 216-224.
Munoz F., Fried G., Armengot L., Bourgeois B., Bretagnolle V., Chadoeuf J., Mahaut L., Plumejeaud C., Storkey J., Violle C. & Gaba S., 2020. Ecological Specialization and Rarity of Arable Weeds: Insights from a Comprehensive Survey in France. Plants 9 : 824.
Pocock M.J.O., Chapman D.S., Sheppard L.J. & Roy, H.E., 2014. Choosing and Using Citizen Science: a guide to when and how to use citizen science to monitor biodiversity and the environment. Centre for Ecology & Hydrology, 28 p.
Solé-Senan X.O., Juárez-Escario A., Conesa J.A. & Recasens J., 2018. Plant species, functional assemblages and partitioning of diversity in a Mediterranean agricultural mosaic landscape. Agriculture, Ecosystems and Environment 256 : 163-172.
Storkey J., Meyer S., Still K.S. & Leuschner C., 2012. The impact of agricultural intensification and land-use change on the European arable flora. Proceedings of the Royal Society, B, rspb20111686.
Tarmi S., Helenius J., & Hyvönen T., 2009. Importance of edaphic, spatial and management factors for plant communities of field boundaries. Agriculture, Ecosystems and Environment 131 (3-4) : 201-206, doi:10.1016/j.agee.2009.01.016.
Tison J.‑M., Jauzein Ph. & Michaud H., 2014. Flore de la France méditerranéenne continentale. Naturalia Publications, Turriers, 2078 p.
Remerciements
Le réseau 500 ENI est piloté par le ministère en charge de l’agriculture et financé par l’Office français de la bodiversité (OFB) dans le cadre du plan Ecophyto. L’analyse des données a été effectuée dans le cadre des projets Ecophyto GT STATS 500 ENI (piloté par Pascal Monestiez entre 2015 et 2018) et GT STEP 500 ENI (co-piloté par Benoît Ricci et Guillaume Fried entre 2019 et 2022), également financés par l’OFB. Les auteurs tiennent à remercier les agriculteurs partenaires du réseau au sein de l’ex-région Languedoc-Roussillon pour la mise à disposition de leurs parcelles et pour les informations sur leurs pratiques.