
Essai de synthèse phytosociologique sur les fourrés sempervirents méditerranéens
Title
A phytosociological synthesis attempt on the Mediterranean evergreen shrubs
Résumé
Dans la synthèse des Pistacio lentisci-Rhamnetea alaterni, classe de fourrés méditerranéens sempervirents, proposée ici sur la base de trente-cinq tableaux synthétiques, on reconnaît la division en trois ordres : les Pistacio lentisci-Rhamnetalia alaterni plutôt basiphiles (avec dix-neuf alliances, dont douze nouvelles et quatre pour la France), les Cytiso villosi-Genistetalia monspessulanae acidiphiles (avec dix alliances, dont six nouvelles et deux en France) et les Rubio tenuifoliae-Pistacietalia terebinthi est-méditerranéens (ordre nouveau, avec quatre alliances, dont deux nouvelles).
Abstract
In the synthesis of Pistacio lentisci-Rhamnetea alaterni, class of evergreen Mediterranean shrubs, proposed here on the basis of thirty five synthetic tables, we recognize the division into three orders: the Pistacio lentisci-Rhamnetalia alaterni rather basiphilic (with nineteen alliances, including eleven new ones and four ones for France), the acidophilous Cytiso villosi-Genistetalia monspessulanae (with ten alliances, including six new ones and two in France), and the east-Mediterranean Rubio tenuifoliae-Pistacietalia terebinthi (a new order, with four alliances, two of which new or validated).
On peut considérer que la végétation arbustive (à nanophanérophytes) d’Europe se répartit entre huit classes, dont sept ont fait l’objet de synthèses récentes pour la France ou au-delà : Betulo pubescentis-Alnetea alnobetulae (de Foucault, 2012), Nerio oleandri-Tamaricetea africanae (de Foucault et al., 2012), Cytisetea scopario-striati (de Foucault et al., 2013a), Franguletea alni (de Foucault & Royer, 2014), Pegano harmalae-Salsoletea vermiculatae (de Foucault, 2015a), Rhamno catharticae-Prunetea spinosae (de Foucault & Royer, 2016) et Salicetea purpureae (de Foucault & Cornier, 2020). La huitième classe, celle des Pistacio lentisci-Rhamnetea alaterni, créée par Julve (1993) mais validée seulement récemment (de Foucault, 2020a), reste donc à traiter. C’est l’objet du présent article, qui suit plus ou moins les modèles précédents, toutefois sans les fiches puisque cette synthèse ne s’inscrit pas vraiment dans le cadre du Prodrome des végétations de France.
1. Méthodes et conventions
Pour effectuer cette synthèse, on a retenu surtout les syntaxons reconnus dans la péninsule Ibérique (d’après le synopsis de Rivas-Martínez et al., 2001, puis des travaux plus récents), en France méditerranéenne (d’après le synopsis de Julve, 1993, puis des travaux postérieurs), en Afrique du Nord méditerranéenne et en Méditerranée orientale (travaux divers). Une fois mis en tableaux, la place de ces syntaxons est évaluée, certains sont déplacés d’un tableau à un autre de façon à construire des tableaux plus homogènes et aussi plus esthétiques. Les alliances classiques sont aussi réévaluées, en tenant compte éventuellement des types nomenclaturaux pour les nommer de façon conforme à l’ICPN 4 (Theurillat et al., 2021). Un tableau synthétique global (tableau 1) est dressé de façon à construire la hiérarchie synsystématique et des tableaux par alliance (tableaux 2 à 35) sont élaborés pour alimenter les colonnes synthétiques du tableau 1 selon une méthode exposée in de Foucault (2020b : 45 et 48). Pour la péninsule Ibérique, la base de données Sivim a été largement utilisée.
Suivant la version 4 de l’ICPN (Theurillat et al., 2021), on a presque systématiquement adopté la nomenclature des taxons actuels, laissant le nom originel lorsque cette correction s’avérait délicate ; en général, le nom originel est rappelé au niveau de la source bibliographique.
On a suivi ici le paradigme synusial (Gillet et al., 1991 ; de Foucault, 2020a) ; étant donné sa taille et sa vigueur, on a conservé Ampelodesmos mauritanicus parmi les arbustes et arbrisseaux. La nomenclature suit Flora Gallica (Tison & de Foucault, 2014) pour les taxons présents en France et Euro+Med Plantbase (http://ww2.bgbm.org/EuroPlusMed/query.asp, qui conserve Calicotome distinct de Cytisus, au contraire de Flora Gallica) pour les autres. Pour gagner de la place dans les tableaux, les termes ‘subsp.’ et ‘var.’ y seront indiqués par le signe *. Les variations autonymes sont réduits à leur initiale (exemple : Rubia peregrina subsp. p. pour Rubia peregrina subsp. peregrina), ainsi que dans le texte. Les taxons juvéniles des essences arborescentes sont indiqués par ‘j’, bien qu’il n’a pas toujours été possible de distinguer clairement les fourrés et les forêts, ainsi que la stratification des phytocénoses dans les travaux classiques ; les taxons plus vigoureux que la normale (par exemple un chaméphyte atteignant exceptionellement la strate nanophanérophytique) seront identifiés par le signe ‘!’. Certains taxons sont considérés au sens large, surtout Rhamnus lycioides s. l., qui incluera aussi Rh. oleoides ; Cistus creticus incluera la subsp. eriocephalus.
Les associations dénommées de manière hétérogène au plan biologique, donc non conformes au paradigme suivi ici, seront identifiés par des guillements entourant leur nom.
On abrègera l’autorité ‘Rivas Mart., T.E. Díaz, Fern.Gonz., Izco, Loidi, Lousã & Penas 2002’ en ‘Rivas Mart. et al. 2002’ ; la locution ‘hors texte’ est abrégée en h. t.
2. Déclinaison de la classe des Pistacio lentisci-Rhamnetea alaterni
PISTACIO LENTISCI-RHAMNETEA ALATERNI Julve ex Julve & B. Foucault in B. Foucault 2020 (Evaxiana 7 : 58)
[syn. : Pistacio lentisci-Rhamnetea alaterni Julve 1993 (Lejeunia, NS, 140 : 109) nom. inval. ; incl. Paliuretea spinae-christi Trijnasić 1978 (Vegetacijska karta SR Hrvatske 1 : 43)]
Typus classis : Pistacio lentisci-Rhamnetalia alaterni Rivas Mart. 1975 (Anales Inst. Bot. Cavanilles 31 (2) : 213) désigné par un des auteurs de la classe (de Foucault, 2020, Evaxiana 7 : 58).
Fourrés méditerranéens sempervirents caractérisés par Smilax aspera, Pistacia lentiscus, P. terebinthus (incl. subsp. palaestina, sans grande valeur selon des travaux récents), Myrtus communis, Osyris alba, Clematis flammula, Asparagus acutifolius, Phillyrea latifolia, Juniperus oxycedrus subsp. o., Prasium majus (quoique rarissime dans l’ordre 2), Olea europaea subsp. e. j, Quercus coccifera, Rhus coriaria, Cytisus laniger… En dehors de Dioscorea communis et Crataegus monogyna, les Pistacio-Rhamnetea alaterni restent bien différenciés des Rhamno-Prunetea spinosae. Cette synthèse montre par ailleurs que Coriaria myrtifolia caractérise plutôt les Rhamno-Prunetea spinosae (plus précisément le Pruno spinosae-Rubion ulmifolii) que la présente classe. Plusieurs taxons appartiennent à des familles eurytropicales qui annoncent les fourrés de latitudes plus basses : Anacardiaceae (Pistacia, Rhus, Searsia), Arecaceae (Chamaerops), Myrtaceae (Myrtus), Smilacaceae (Smilax) (pour plus de détails cf. infra, § 3).
Ces fourrés entretiennent des relations spatiales et temporelles (dynamiques) avec les boisements des Pino halepensis-Quercetea ilicis (de Foucault, 2020a), dont plusieurs taxons caractéristiques apparaissent notés j dans les divers tableaux, les garrigues plus basses des Rosmarinetea officinalis (de Foucault, 2020d) et les landes des Cisto-Lavanduletea stoechadis.
À plusieurs reprises dans des travaux antérieurs, on a déjà évoqué le cas des « macrochaméphytes », végétaux intermédiaires entre chaméphytes et nanophanérophytes qui caractérisent une végétation intermédiaire entre landes et fourrés. C’est par exemple le cas des Pistacio lentisci-Cistetum eriocephali et Charybdido pancratii-Cistetum monspeliensis d’Apulie (Di Pietro & Misano, 2010), du Cistetum monspeliensis des Albères (Julve & Bouteloup, 2018 : 212), du Rhamno saxatilis-Genistetum scorpii (Choisnet, 2019, tableau 12), du Cisto albidi-Genistetum scorpii (de Foucault, 2019b) et d’un matorral ibérique à Coronilla juncea (photo 1), végétations d’environ 8 à 10 décimètres de hauteur (photo 2), ce qui correspond plus ou moins à la hauteur du Quercetum cocciferae (photo 13) classiquement rangé dans les Pistacio-Rhamnetalia alaterni. De Corse, le relevé suivant décrit une végétation de ce type avec Genista salzmannii :
Calvi, près de la zone industrielle de Cantone, N 42° 32’ 11,8’’, E 8° 48’ 21,8’’, 33 m (coordonnées et altitude approximatives), 28 mai 2014, 9 taxons :
- Cistus monspeliensis 4, Pistacia lentiscus 3, Phillyrea angustifolia 2, Genista salzmannii +, Asparagus acutifolius +, Erica scoparia s. +,
- autres taxons : Lavandulea stoechas *s. ! 2, Olea europaea *e. j 2, Fraxinus angustifolia *a. j 1,
alors que la photo 3 en illustre une autre avec Cistus creticus et C. monspeliensis, vue aussi par Paradis et al. (2014, tableau 20 : relevés 1 et 2) : Cistetum monspeliensi-cretici ass. nov. hoc loco, typus nominis hoc loco : relevé 2 du tableau 20 in Paradis et al. (2014, J. Bot. Soc. Bot. France 67 : 66). Il y a sans doute d’autres associations de ce type à décrire, on les laissera de côté (sauf bien sûr le Quercetum cocciferae) dans cette synthèse en attendant qu’elles soient mieux connues. Signalons cependant que Choisnet (2019 : 104) a défini le Lonicero etruscae-Genistion scorpii pour y placer certaines de ces végétations. Les macrochaméphytes seront ici considérés comme caractéristiques de la présente classe, leurs formes juvéniles étant alors considérées comme différentielles au sein des Cisto-Lavanduletea stoechadis et des Rosmarinetea officinalis.
Vers le nord de son aire, cette classe est relayée par les Rhamno catharticae-Prunetea spinosae et les Franguletea alni d’Europe tempérée à subméditerranéenne, avec des alliances de passage comme le Pruno spinosae-Rubion ulmifolii et le Frangulo alni-Pyrion cordatae (de Foucault & Royer, 2014, 2016) ; vers le sud, elle est notamment relayée par les Zizipho loti-Searsietea tripartitae d’optimum steppique saharien et les Kleinio neriifoliae-Euphorbietea canariensis macaronésiens (de Foucault, 2020a), des alliances précisées ci-après assurant parfois les passages chorologiques.
Le tableau synthétique 1 dégage clairement les deux ordres classiques plutôt ouest-méditerranéens (O.1 et O.2) et met en évidence un troisième (O.3). D’ailleurs l’ensemble des deux premiers ordres se différencie très significativement du troisième, plutôt est-méditerranéen, par Rhamnus alaternus subsp. a., Daphne gnidium, Chamaerops humilis, Ampelodesmos mauritanicus, Phillyrea angustifolia, Ph. media, Viburnum tinus, Lonicera implexa, Rubus ulmifolius, Rubia peregrina subsp. longifolia, Teucrium fruticans, Rosa sempervirens, Cistus albidus, C. monspeliensis et Cytisus spinosus (ces deux derniers réputés acidiphiles, mais pourtant assez bien représentés dans certaines alliances du premier ordre), Ulex parviflorus subsp. p., Bupleurum fruticosum, Coronilla glauca, Clematis cirrhosa, Pinus halepensis j, Pinus pinaster j… L’ensemble O.1 + O.2 possède donc la valeur d’une sous-classe, les Eu-Pistacio lentisci-Rhamnenea alaterni ; Rubia peregrina subsp. p. et Rubus ulmifolius se partagent entre cette sous-classe et la partie atlantique à subméditerranéenne des Rhamno-Prunetea spinosae. Pour simplifier, dans la suite, on n’utilisera pas cette unité supérieure, on la rappellera juste en conclusion. Dans le tableau 1, Viburnum tinus, normalement caractéristique de sous-classe, apparaît comme taxon différentiel des Cytiso villosi-Genistetalia monspessulanae ; il s’agit sans doute d’un effet d’échantillonnage et statistique.




Ordre 1. Pistacio lentisci-Rhamnetalia alaterni Rivas Mart. 1975 (Anales Inst. Bot. Cavanilles 31 (2) : 213) ; tableau 1 : colonnes CcPl à Git
Typus ordinis : Asparago albi-Rhamnion oleoidis Rivas Goday ex Rivas Mart. 1975 (Anales Inst. Bot. Cavanilles 31 (2) : 213) désigné par l’auteur de l’ordre (1975, Anales Inst. Bot. Cavanilles 31 (2) : 213).
Fourrés méditerranéens plutôt basiphiles à Rhamnus lycioides s. l., Erica multiflora, Jasminum fruticans, Genista hirsuta subsp. erioclada, Asparagus albus, Juniperus phoenicea subsp. ph., Euphorbia characias subsp. c., Anthyllis cytisoides, Anagyris foetida, Globularia alypum.
Alliance CcPl. Clematido cirrhosae-Pistacion lentisci all. nov. hoc loco ; tableau 1 : colonne CcPl ; tableau 2
[syn. : Oleo-Ceratonion auct. p. p.]
Typus nominis hoc loco : Clematido cirrhosae-Myrtetum communis O. Bolòs & Molin. ex O. Bolòs, Molin. & P. Monts. 1970 (Acta Bot. Barcinon. 5 : 40) [syn. : groupement à Myrtus communis O. Bolòs & Molin. 1958 (Collect. Bot. (Barcelona) 5 (3) : 730)].
Fourrés thermo-méditerranéens des îles Baléares à Ampelodesmos mauritanicus, Rubia peregrina subsp. longifolia, Cneorum tricoccon, Buxus balearica, Clematis cirrhosa (incl. var. balearica ; photo 4), Euphorbia dendroides, Phillyrea media subsp. rodriguezii, Genista majorica, Rhamnus ludovici-salvatoris.
CcPl1. Ampelodesmo mauritanici-Arbutetum unedonis Llorens, Gil & Tébar in Rivas Mart. et al. 2002 (Sivim, consulté le 14/09/2020).
CcPl2. « Aro picti-Phillyreetum rodriguezii O. Bolòs, Molin. & P. Monts. 1970 » (de Bolòs et al., 1970, tableau 3) ; à renommer une fois éclaircies la systématique et la nomenclature au sein du complexe Phillyrea media.
CcPl3. Buxo balearicae-Genistetum majoricae Rivas Mart. in Rivas Mart., M.J. Costa, P. Soriano, Pérez-Badia, Llorens & Roselló 1992 (Rivas-Martínez et al., 1992, tableau 10).
CcPl4. « Cneoro tricocci-Ceratonietum siliquae O. Bolòs in O. Bolòs & Molin. 1958 » (de Bolòs & Molinier, 1958, tableau 2) ; à revoir après caractérisation et séparation de la synusie arborescente : Ceratonio siliquae-Oleetum europaeae B. Foucault 2021 (de Foucault, 2021a), à ranger dans les Pino halepensis-Quercetea ilicis et les Quercetalia rotundifolio-ilicis (de Foucault, 2020a). Dans l’esprit de Guinochet (in Guinochet & Drouineau, 1944 : 25), il paraît en effet évident que l’Oleo–Ceratonion siliquae Braun-Blanq. ex Guin. & Drouineau 1944 correspond à des forêts « climaciques » thermo-méditerranéennes à Olea europaea et Ceratonia siliqua ; il en est de même chez Molinier (1954), puis de Bolòs et Molinier (1958) qui conçoivent la végétation de cette alliance comme remplaçant le Quercetum ilicis, soit clairement deux syntaxons forestiers climaciques homologues. C’est donc à cette alliance qu’il faut rattacher le Ceratonio siliquae-Oleetum europaeae arborescent.
CcPl5. Clematido cirrhosae-Myrtetum communis O. Bolòs & Molin. ex O. Bolòs, Molin. & P. Monts. 1970 nom. mut. hoc loco (de Bolòs & Molinier, 1958, tableau 3 ; de Bolòs et al., 1970, tableau 5 ; sub Clematido balearicae-Myrtetum communis).
CcPl6. Cneoro tricocci-Rhamnetum bourgaeanae Tébar & Llorens 1995 (Sivim, consulté le 14/09/2020).
CcPl7. Rhamno ludovici-salvatoris-Juniperetum turbinatae (Camarasa, Cardona, Masalles, Terradas, E. Velasco & Vigo 1976) Gil, Llorens, Tébar & M.J. Costa 1995 (Sivim, consulté le 14/09/2020) ; ou à placer dans le Juniperion turbinatae ?
CcPl8. Cneoro tricocci-Pistacietum lentisci O. Bolòs & Molin. (1969) 1984 (de Bolòs & Molinier, 1984 : 208).
CcPl9. Smilaco asperae-Ampelodesmetum mauritanici Rivas Mart. in Rivas Mart., M.J. Costa, P. Soriano, Pérez-Badia, Llorens & Roselló 1992 nom. mut. hoc loco (Sivim, consulté le 14/09/2020, sub Smilaco balearicae-Ampelodesmetum mauritanici).
CcPl10. Prasio majoris-Oleetum europaeae O. Bolòs & Molin. 1969 (de Bolòs & Molinier, 1969 : 257).
Il faudrait y ajouter encore le fourré à Buxus balearica relevé à Majorque vers 1 200 m d’altiude par de Bolòs et Molinier (1958 : 739).
Alliance ARo. Asparago albi-Rhamnion oleoidis Rivas Goday ex Rivas Mart. 1975 (Anales Inst. Bot. Cavanilles 31 (2) : 213) ; tableau 1 : colonne ARo ; tableau 3
Typus allianciae : Asparago albi-Rhamnetum oleoidis Rivas Goday in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Anales Inst. Bot. Cavanilles 17 (2) : 320) désigné par l’auteur de l’alliance (1975 : 213).
Fourrés ibériques à Chamaerops humilis (photo 5), Asparagus aphyllus, Rhamnus lycioides s. l., Quercus coccifera… ; cartographie chorologique in Rivas-Martínez (1974 : cartes 2 et 5).
ARo1. Ampelodesmo mauritanici-Chamaeropetum humilis Braun-Blanq., Font Quer, G. Braun-Blanq., Frey, Jansen & Moor 1936 (Sivim, consulté le 13/09/2020).
ARo2. Asparago albi-Rhamnetum oleoidis Rivas Goday in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Sivim, consulté le 13/09/2020).
ARo3. Asparago albi-Quercetum cocciferae Rivas Mart. in Rivas Mart. et al. 2002 (Sivim, consulté le 13/09/2020) ; sans aucun doute à rattacher au précédent, alors prioritaire.
ARo4. Asparago aphylli-Calicotometum villosae Rivas Mart. 1975 (Sivim, consulté le 14/09/2020).
ARo5. Asparago aphylli-Myrtetum communis Rivas Mart., Cantó, Fern.Gonz. & Sánchez-Mata ex J.C. Costa, Lousã & Espírito-Santo 1997 (Sivim, consulté le 14/09/2020).
ARo6. Bupleuro gibraltarici-Pistacietum lentisci Martínez-Parras, Peinado & Alcaraz 1986 (Sivim, consulté le 14/09/2020).
ARo7. Chamaeropo humilis-Juniperetum phoeniceae Rivas Mart. in Alcaraz, T.E. Díaz, Rivas Mart. & P. Sánchez 1989 (Sivim, consulté le 14/09/2020).
ARo8. Chamaeropo humilis-Rhamnetum lycioidis O. Bolòs 1957 (Sivim, consulté le 14/09/2020).
ARo9. Cneoro tricocci-Buxetum balearicae Rivas Goday & Rivas Mart. 1969 (Sivim, consulté le 14/09/2020).
ARo10. « Melico arrectae-Quercetum cocciferae Braun-Blanq., P. Silva & Rozeira 1956 » (Sivim, consulté le 14/09/2020).
ARo11. Phillyreo angustifoliae-Rhamnetum angustifoliae M.J. Costa & Mansanet 1981 (Sivim, consulté le 14/09/2020).
ARo12. Quercetum coccifero-airensis Espírito-Santo in Espírito-Santo, Lousã, J.C. Costa & Capelo 1994 (Sivim, consulté le 14/09/2020).
ARo13. Querco cocciferae-Pistacietum lentisci Braun-Blanq., Font Quer, G. Braun-Blanq., Frey, Jansen & Moor 1936 (Sivim, consulté le 14/09/2020).
ARo14. « Teucrio-Hippocrepidetum valentinae O. Bolòs 1956 » (Sivim, consulté le 14/09/2020).
ARo15. « Melico-Cocciferetum Braun-Blanq., P. Silva & Rozeira 1956 » (Braun-Blanquet et al., 1956, tableau IV).

Non synthétisées dans la colonne ARo du tableau 1, les trois associations suivantes, pas spécialement littorales, paraissent assurer des transitions entre cette alliance et le Juniperion turbinatae :
ARo16. Asparago horridi-Juniperetum turbinatae Díez-Garretas, Asensi & Martín 1996 (Sivim, consulté le 14/09/2020).
ARo17. « Phlomido purpureae-Juniperetum turbinatae Capelo, Lousã & J.C. Costa 1994 » (Sivim, consulté le 14/09/2020).
ARo18. Aristolochio baeticae-Arbutetum unedonis Pinto Gomes & Cano 2002 (Pinto Gomes & Paiva, 2005, tableau LXXVII).
Alliance Pa. Periplocion laevigatae Rivas Mart. 1975 nom. mut. hoc loco (Anales Inst. Bot. Cavanilles 31 (2) : 216, sub Periplocion angustifoliae) ; tableau 1 : colonne Pl ; tableau 4
Typus allianciae : Mayteno europaei-Periplocetum laevigatae Rivas Goday & Esteve in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Anales Inst. Bot. Cavanilles 17 (2) : 285-406).
Fourrés substeppiques du Sud-Est ibérique, ainsi que de la côte sud et de quelques îles de Sicile à Lycium intricatum, Asparagus albus, partageant avec l’alliance suivante Periploca laevigata (= P. angustifolia ; photo 6) ; cartographie chorologique in Rivas-Martínez (1974 : cartes 2 et 5) et Brullo et al. (2008 : figure 1).
* Groupe de syntaxons ibériques, faiblement différenciés
Pl1. « Arisaro simorrhini-Tetraclinidetum articulatae Rivas Goday & Rivas Mart. in Rivas Mart. 1975 » (Sivim, consulté le 10/09/2020)
Pl2. Mayteno europaei-Periplocetum laevigatae Rivas Goday & Esteve in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Sivim, consulté le 10/09/2020).
* Groupe de syntaxons siciliens, fortement différenciés
Pl3. Periploco laevigatae-Juniperetum turbinatae Bartolo, Brullo, Minissale & Spampinato 1988 nom. mut. hoc loco (Gianguzzi et al., 2012, tableau 4, sub Periploco angustifoliae-Juniperetum turbinatae).
Pl4. Periploco laevigatae-Euphorbietum dendroidis Brullo, Di Martino & Marcenò 1977 nom. mut. hoc loco (Brullo et al., 2008, tableau 1 : colonnes 3 à 12, sub Periploco angustifoliae-Euphorbietum dendroidis).
Pl5. Calicotomo infestae-Searsietum tripartitae Bartolo, Brullo & Marcenò 1982 nom. mut. hoc loco (Ibid., tableau 1 : colonne 13, sub Calicotomo infestae-Rhoetum tripartitae).
Pl6. Periploco laevigatae-Searsietum tripartitae Brullo, Gianguzzi, La Mantia & Siracusa 2008 nom. mut. hoc loco (Ibid., tableau 1d, sub Periploco angustifoliae-Rhoetum tripartitae).
Pl7. Ephedro fragilis-Pistacietum lentisci Brullo, Guarino & Ronsisvalle 2000 (Ibid., tableau 1 : colonnes 15 et 16).
Pl8. Asparago horridi-Retametum gussonei Brullo, Guarino & Ronsisvalle 2000 (Ibid., tableau 1 : colonne 17).
Avec Lycium intricatum (photo 7), Asparagus albus, Withania frutescens et Ziziphus lotus (photo 8), et très peu de taxons caractérisant les Pistacio-Rhamnetea alaterni, les Mayteno europaei-Ziziphetum loti Fern.Casas 1970 et Ziziphetum loti Rivas Goday & Bellot 1944 paraissent mieux placés dans les Warionio saharae-Withanietalia frutescentis B. Foucault 1993 ex 2020 et les Zizipho loti-Searsietea tripartitae B. Foucault 1993 ex 2020 d’optimum nord-africain steppique ; ces deux syntaxons sont replacés dans le tableau synthétique de cette classe (tableau 5a en RZl1 et RZl2, d’après Sivim, consulté le 10/09/2020), reprenant et complétant le tableau B8 in de Foucault (2020a), montrant qu’une alliance steppique originale peut les rapprocher, le Rubio peregrinae-Ziziphion loti all. nov. hoc loco, caractérisée par la combinaison de Rubia peregrina subsp. p., Retama sphaerocarpa, Lycium intricatum, Asparagus albus, Withania frutescens et Ziziphus lotus, typus nominis hoc loco : Mayteno europaei-Ziziphetum loti Fern.Casas 1970 (Publ. Inst. Biol. Aplic. 49 : 113). Au sein du Withanion adpressae, les nouvelles colonnes Wa3 et Wa4 correspondent respectivement aux Zizipho loti-Rhoetum tripartitae Quézel, Barbero, Benabid & Rivas Mart. 1994 (Quézel et al., 1994, tableau 3) et Adenocarpo bacquei-Buxetum balearicae sensu Quézel et al. 1994 (Phytocoenologia 22 (4) : 546), non Quézel & Barbero 1981 (Quézel et al., 1994, tableau 4 ; cf. infra PJp1) ignorés dans la synthèse de 2020.
Ajoutons à cela que Braun-Blanquet et Maire (1924 : 26, 40, 48, 49, 53, 58, 73, 78, 83, 86, 94) publient plusieurs relevés de fourrés substeppiques du Lycio intricati-Ephedrion altissimae.
D’un autre côté, Quézel et Barbero (1981) décrivent de façon invalide les Ephedro-Juniperetea phoeniceae et les Ephedro-Juniperetalia phoeniceae (art. 2d, 3g, 3o, 5), rassemblant des formations présteppiques à genévriers du Maroc. Aucune alliance n’y est définie. Sur la base de ce tableau 5a (ensemble A.1.4), on propose ici de définir le Polygalo balansae-Juniperion phoeniceae all. nov. hoc loco, typus nominis hoc loco : Retamo dasycarpae-Juniperetum phoeniceae Quézel & Barbero ex B. Foucault, caractérisé par la combinaison de Juniperus phoenicea subsp. ph., J. oxycedrus subsp. o., Polygala balansae, Ephedra fragilis subsp. cossonii , Coronilla ramosissima, Pistacia atlantica j, avec les syntaxons suivants :
PJp1. Adenocarpo bacquei-Buxetum balearicae Quézel & Barbero 1981 (Quézel & Barbero, 1981, tableau 3), assez différent de Wa4 (cf. supra).
PJp2. Groupement rupicole à Warionia saharae-Antirrhinum ramosissimum (Ibid. : 1143-1144).
PJp3. Coronillo ramosissimae-Juniperetum phoeniceae Quézel & Barbero 1981 nom. inval. (Ibid., tableau 1) ; validé ici en Coronillo ramosissimae-Juniperetum phoeniceae Quézel & Barbero ex ass. nov. hoc loco, typus nominis hoc loco de l’association : relevé 4 du tableau 1 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) : h. t.), type du typicum différencié surtout par Retama dasycarpa et Cupressus dupreziana subsp. atlantica j, des zones atlasiques plus internes ; typus nominis hoc loco du searsietosum tripartitae Quézel & Barbero ex subass. nov. hoco : relevé 11 du tableau 1 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) : h. t.), différencié par Searsia tripartita, Asparagus albus, Warionia saharae, des zones atlasiques périphériques soumises aux influences atlantiques.
PJp4. Retamo dasycarpae-Juniperetum phoeniceae Quézel & Barbero 1981 nom. inval. (Ibid., tableau 2), validé ici en Retamo dasycarpae-Juniperetum phoeniceae Quézel & Barbero ex ass. nov. hoc loco, typus nominis hoc loco de l’association : relevé 4 du tableau 2 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) : h. t.).
PJp5 à PJp7. « Bupleuro spinosi-Juniperetum phoeniceae Quézel & Barbero 1981 » (Ibid., tableau 5) ; ce syntaxon paraît trop complexe pour ne former que trois variations d’une même association. Le syntaxon PJp5 (relevés 3 et 5) est une junipéraie à Rhamnus lycioides subsp. atlantica et Asparagus horridus, originale mais deux relevés connus seulement. PJp6 (relevés 1, 2, 4 et 6) est une junipéraie paucispécifique. PJp7 (relevés 7 à 15) est bien mieux défini, floristiquement et statistiquement : Juniperetum thurifero-oxycedri (Quézel & Barbero 1981) ass. nov. hoc loco, typus nominis hoc loco : relevé 10 du tableau 5 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) : h. t.) ; avec Berberis hispanica et Crataegus laciniata, il annonce le Berberido hispanicae-Crataegion laciniatae.
PJp8. Groupement à Genista tridens (Braun-Blanquet & Maire, 1924 : 143).
Cette alliance ne relève plus des Pistacio-Rhamnetea alaterni, on propose de la rattacher aux Zizipho loti-Searsietea tripartitae et aux Warionio saharae-Withanietalia frutescentis, quoique un peu par enchaînement pour les syntaxons PJp1 et PJp4 à PJp8.
Dans la publication de Quézel et Barbero (1981), il reste encore les « Berberido hispanicae-Fraxinetum dimorphae » et « Lonicero arboreae-Cedretum atlanticae » (Quézel & Barbero, 1981 : 1155). Ils paraissent plus proches du Berberido hispanicae-Crataegion laciniatae B. Foucault 1993 ex 2020 et des Crataego laciniatae-Berberidetalia hispanicae B. Foucault 2020 (de Foucault, 2020a). Le tableau 5b (reprenant une partie du tableau B9 in de Foucault, 2020a) synthétise les syntaxons suivants :
BCl1. Roso micranthae-Berberidetum hispanicae (Quézel, Barbero & Benabid 1987) B. Foucault 2020 (de Foucault, 2020a, tableau B9 : colonne LJ8).
BCl2. Lonicero arboreae-Berberidetum hispanicae B. Foucault 2020 (Ibid. : colonne LJ9).
BCl3. Buxo sempervirentis-Juniperetum thuriferae Quézel 1957 (Ibid. : colonne LJ10).
BCl4. « Berberido hispanicae-Fraxinetum dimorphae Quézel & Barbero 1981 » (Quézel & Barbero, 1981, tableau 6), complexe au sein duquel on peut isoler le Junipero thuriferae-Berberidetum hispanicae (Quézel & Barbero 1981) ass. nov. hoc loco, typus nominis hoc loco de l’association et du typicum (différencié par Quercus rotundifolia j et Fraxinus dimorpha j) : relevé 6 du tableau 6 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) h. t.) ; typus nominis hoc loco du rosetosum siculae subass. nov. hoc loco (différencié par Rosa sicula, Rhamnus lycioides subsp. atlantica, Genista scorpius, optimum de Juniperus thurifera) : relevé 9 du tableau 6 in Quézel & Barbero (1981, Bol. Soc. Brot., série 2, 53 (2) h. t.).
BCl5. « Lonicero arboreae-Cedretum atlanticae Barbero, Quézel & Rivas Mart. 1981 » (Barbero et al., 1981, tableau 34).
Alliance RJp. Rhamno myrtifoliae-Juniperion phoeniceae all. nov. hoc loco ; tableau 1 : colonne RJp ; tableau 6
Typus nominis hoc loco : Rhamno myrtifoliae-Juniperetum phoeniceae Molero & Pérez-Raya 1987 (Lazaroa 7 : 306).
Fourrés du Sud-Est ibérique à Rhamnus myrtifolia (photo 9), Juniperus phoenicea subsp. ph., J. oxycedrus subsp. o.
RJp1. Rhamno myrtifoliae-Juniperetum phoeniceae Molero & Pérez-Raya 1987 (‘… myrtifolii-…’ art. 41b ; Sivim, consulté le 10/09/2020).
RJp2. Rhamno lycioidis-Pinetum halepensis (Torres, García-Fuentes, Salazar, Cano & F. Valle 1999) Rivas Mart. in Rivas Mart. et al. 2002 (Torres et al., 1999, tableau 1 sub Junipero phoeniceae-Pinetum halepensis).
RJp3. Rhamno myrtifoliae-Quercetum rotundifoliae Pérez Latorre, Casimiro & Cabezudo 2015 (Pérez Latorre et al., 2015, tableau 15).
Alliance ALn. Arbuto unedonis-Laurion nobilis Rivas Mart., Fern.Gonz. & Loidi 1999 (Itinera Geobot. 13 : 400) ; tableau 1 : colonne ALn ; tableau 7
Typus allianciae : Phillyreo latifoliae-Arbutetum unedonis (A. Velasco 1983) Loidi, Herrera, Olano & Silván 1994 (J. Veg. Sci. 5 (4) : 537) désigné par les auteurs de l’alliance (1999, Itinera Geobot. 13 : 400).
Fourrés acidiclinophiles du nord et du nord-ouest de la péninsule Ibérique, sous influence atlantique, à Lonicera periclymenum subsp. p., Prunus spinosa, Laurus nobilis j, moins souvent Arbutus unedo. La séparation d’avec une éventuelle forêt à Laurus nobilis ne paraît pas clairement établie.
La partie la plus acidiphile de l’alliance initiale (excluant le type) est intégrée ici aux Cytiso villosi–Genistetalia monspessulanae en tant que Pruno lusitanicae-Ericion arboreae (cf. infra, alliance PEa). De même, l’intégration à cette alliance du « Scrophulario laxiflorae-Rhododendretum baetici », de l’extrême sud de la péninsule, donc très éloigné du centre de l’aire de la présente alliance, au sein de la sous-alliance du Rhododendrenion baetici Rivas Mart. & Sánchez-Mata 2001, nous paraît assez peu pertinente ; on propose ici l’intégration de cette association plutôt acidiphile aux Cytiso villosi–Genistetalia monspessulanae (cf. infra, alliance Rb).
ALn1. « Calluno vulgaris-Lauretum nobilis Fern.Prieto, Arbesú & Bueno in T.E. Díaz & Fern.Prieto 1994 » (Sivim, consulté le 10/09/2020) ; voir aussi Rodríguez Guitián et al. (2007).
ALn2. Hedero helicis-Lauretum nobilis Bueno & Fern.Prieto 1991 (Sivim, consulté le 10/09/2020).
ALn3. Glandoro diffusae-Oleetum europaeae Bueno & Fern.Prieto 1991 (Sivim, consulté le 10/09/2020).
ALn4. Phillyreo latifoliae-Arbutetum unedonis (A. Velasco 1983) Loidi, Herrera, Olano & Silván 1994 (Sivim, consulté le 10/09/2020).


ALn5. Smilaco asperae-Arbutetum unedonis T.E. Díaz & Fern.Prieto 1994 (Díaz & Fernández-Prieto, 1994, tableau 7).
ALn6. Dioscoreo communis-Lauretum nobilis Rodríguez Guitián, Romero & Ramil 2007 (Rodríguez Guitián et al., 2007, tableau 5 : relevés 1 à 19).


Alliance EMc. Euphorbio dendroidis-Myrtion communis all. nov. hoc loco ; tableau 1 : colonne EMc ; tableau 8
[syn. : Oleo-Ceratonion siliquae auct. p. p. ; cf. supra, au Ceratonio siliquae-Oleetum europaeae en CcPl4]
Typus nominis hoc loco : Euphorbietum dendroidis Guin. in Guin. & Drouineau 1944 (Recueil des travaux de l’Institut botanique de Montpellier I : 26).
Fourrés ouest-méditerranéens thermophiles à Euphorbia dendroides, Myrtus communis, Cytisus spinosus, Ceratonia siliqua j, Olea europaea subsp. e. j souvent en lien avec les forêts de l’Oleo–Ceratonion siliquae ; cartographie chorologique in Rivas-Martínez (1974 : cartes 2 et 5, sub Oleo-Ceratonion).
EMc1. Calicotomo spinosae-Myrtetum communis Guin. in Guin. & Drouineau 1944 em. O. Bolòs 1962 (Sivim, consulté le 14/09/2020), transition vers l’Ericion arboreae.
EMc2. Chamaeropo humilis-Euphorbietum dendroidis O. Toubal & A. Toubal 1996 (Toubal & Toubal, 1996b, tableau 4).
EMc3. Groupement à Olea europaea-Pistacia terebinthus O. Toubal & A. Toubal 1996 (Ibid., tableau 1).
EMc4. Bupleuro fruticosi-Euphorbietum dendroidis Géhu, Kaabèche & Gharzouli 1992 (Géhu et al., 1992b, tableau 1).

EMc5. Euphorbietum dendroidis Guin. in Guin. & Drouineau 1944 (Lapraz, 1974, tableau 1 : colonnes 1 à 43) ; photo 10.
EMc6. Myrto communis-Pistacietum lentisci (Molin. 1954) Rivas Mart. 1975 (Molinier, 1954, tableau h. t.) ; syn. : Oleo-Lentiscetum provinciale Molin. 1954 (« Oleolentiscetum auct. ») ; voir aussi Braun-Blanquet et al. (1952 : 243), Lapraz (1977), Franquesa (1995, tableau 53) et Noble & Baret (2021).
EMc7. Pistacio lentisci-Rhamnetum alaterni O. Bolòs 1970 (de Bolòs, 1970, tableau VI).
EMc8. Phillyreo latifoliae-Euphorbietum dendroidis Paradis, Hugot & Delage 2005 (Paradis et al., 2005, tableau 1).
EMc9. Asparago albi-Euphorbietum dendroidis Biondi & Mossa 1992 ; a – de Sardaigne (Biondi & Mossa, 1992, tableau 2) ; b – de Corse (Paradis & Piazza, 2001, tableau 6) ; ce syntaxon corse est cependant plus proche de EMc8, lui-même corse, que de EMc9a.
EMc10. Oleo europaeae-Euphorbietum dendroidis Trinajstić 1973 (Trinajstić, 1984, tableau 2).
EMc11. Cisto monspeliensis-Euphorbietum dendroidis (Molin. 1937) Julve 1993 prov. (Molinier, 1937 : 113) des îles d’Hyères ; à étayer sur de bien plus nombreux relevés.
EMc12. Myrto communis-Juniperetum oxycedri Franquesa 1995 (Franquesa, 1995, tableau 50).
Sous le terme de « Oleo-Ceratonion », Brullo et al. (2008, tableau 3) synthétisent quatorze associations siciliennes, qu’on va réduire à douze en écartant l’Asparago acutifolii-Ziziphetum loti Gianguzzi, Ilardi & Raimondo 1996 (colonne 30 du tableau cité), qui paraît mal placé ici, et l’Erico arboreae-Pinetum halepensis (colonne 39), connu de seulement trois relevés et qui va plutôt vers les Cytiso-Genistetalia monspessulanae. Notre tableau 9 décrit un syntaxon de rang alliance rappelant l’Euphorbio dendroidis-Myrtion communis précédent par les deux taxons éponymes de celui-ci, ainsi que Prasium majus, Ceratonia siliqua j, mais nettement différencié par Calicotome infesta subsp. i., Ephedra fragilis, Teucrium fruticans, Rubia peregrina subsp. longifolia, ainsi que par l’absence significative de Ampelodesmos mauritanicus, Cytisus spinosus, Cistus monspeliensis, Clematis flammula, Rubia peregrina subsp. p.
Alliance CTf. Calicotomo infestae-Teucrion fruticantis all. nov. hoc loco ; tableau 1 : colonne CTf ; tableau 9
Typus nominis hoc loco : Salvio fruticosae-Phlomidetum fruticosae Barbagallo, Brullo & Fagotto 1979 (Pubb. Ist. Bot. Univ. Catania 1979 non consulté).
Fourrés thermophiles centre-sud-méditerranéens, surtout connus de Sicile, à Euphorbia dendroides, Myrtus communis, Prasium majus, Ceratonia siliqua j, Calicotome infesta subsp. i., Ephedra fragilis, Teucrium fruticans (photo 11), Rubia peregrina subsp. longifolia.
* Groupe d’associations à Asparagus albus, Euphorbia dendroides, Myrtus communis, Clematis cirrhosa…
CTf1. Euphorbietum dendroidis sensu Brullo, Gianguzzi, La Mantia, & Siracusa 2008, non Guin. in Guin. & Drouineau 1944 (Brullo et al., 2008, tableau 3 : colonnes 1 à 16).
CTf2. Pistacio lentisci-Chamaeropetum humilis Brullo & Marcenò 1985 (Ibid., tableau 3 : colonnes 17 à 20).
CTf3. « Chamaeropo humilis-Sarcopoterietum spinosi Barbagallo, Brullo & Fagotto 1979 » (Ibid., tableau 3 : colonnes 21 et 22), à associer au précédent.
CTf4. Teucrio fruticantis-Rhamnetum alaterni Brullo, Minissale, Scelsi & Spampinato 1993 (Ibid., tableau 3 : colonnes 23 et 24).
CTf5. Myrto communis-Pistacietum lentisci sensu Brullo, Gianguzzi, La Mantia & Siracusa 2008, non (Molin. 1954) Rivas Mart. 1975 (Ibid., tableau 3 : colonnes 25 à 28).
CTf6. Ephedro fragilis-Lycietum europaei Brullo & Marcenò 1985 (Ibid., tableau 3 : colonne 29).
CTf7. Chamaeropo humilis-Quercetum cocciferae Brullo & Marcenò 1985 (Ibid., tableau 3 : colonnes 31 à 33, sub Chamaeropo humilis-Quercetum calliprini).
CTf8. Pyro spinosae-Calicotometum infestae Brullo, Gianguzzi, La Mantia & Siracusa 2008 nom. mut. hoc loco (Ibid., tableau 3 : colonne 34, sub Pyro amygdaliformis-Calicotometum infestae).
CTf9. Salvio fruticosae-Phlomidetum fruticosae Barbagallo, Brullo & Fagotto 1979 (Ibid., tableau 3 : colonnes 35 à 37).
* Autres associations
CTf10. Calicotomo infestae-Juniperetum turbinatae Brullo, Gianguzzi, La Mantia & Siracusa 2008 (Ibid., tableau 3 : colonne 38).
CTf11. « Pistacio lentisci-Pinetum halepensis De Marco, Veri & Caneva 1984 » (Ibid., tableau 3 : colonnes 40 à 43).
CTf12. Hippocrepido emeri-Bupleuretum fruticosi Brullo, Minissale, Scelsi & Spampinato 1993 (Ibid., tableau 3 : colonnes 44 et 45).

Alliance RQc. Rhamno lycioidis-Quercion cocciferae Rivas Goday ex Rivas Mart. 1975 (Anales Inst. Bot. Cavanilles 31 (2) : 208) ; tableau 1 : colonne RQc ; tableau 10
Lectotypus nominis hoc loco (l’alliance ne paraît pas avoir été validée par son auteur) : Quercetum cocciferae Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952 (Les groupements végétaux de la France méditerranéenne : 240) [syn. : Quercetum cocciferae Braun-Blanq. 1925 (Veröff. Geobot. Inst. E.T.H. Stiftung Rübel 3 : 317) nom. nud. inval.].
Fourrés catalans, franco-méditerranéens et sardes peu caractérisés, surtout par Quercus coccifera et Jasminum fruticans ; cette alliance marque en fait la fin de l’extension de la classe vers le nord de son aire, avant le passage aux Rhamno-Prunetea spinosae ; cartographie chorologique in Rivas-Martínez (1974 : carte 5).
* Groupe de syntaxons à Quercus coccifera, Q. rotundifolia j, Q. faginea subsp. f. j, Clematis flammula, Jasminum fruticans, Rhamnus lycioides s. l., Cistus albidus, Euphorbia characias subsp. c., Pistacia terebinthus, Genista scorpius, Lonicera etrusca, Viburnum tinus
RQc1. Asparago acutifolii-Jasminetum fruticantis O. Bolòs 1961 (Sivim, consulté le 14/09/2020).
RQc2. Pyro spinosae-Paliuretum spinae-christi Kuhnholtz-Lordat 1954 (Dieleman, 1970, tableau 1) ; photo 12.
RQc3. « Staehelino dubiae-Rhoetum coriariae Noble & Baret » (Noble & Baret, 2021, tableau 37).
RQc4. Jasmino fruticantis-Phillyreetum angustifoliae Tallon in Noble & Baret 2021 (Ibid., tableau 38 : colonne 5).
RQc5. Quercetum cocciferae Braun-Blanq. ex Braun-Blanq., Roussine & Nègre 1952 [syn. : Junipero oxycedri-Quercetum cocciferae (Braun-Blanq. 1925) B. Foucault & Julve 1991 (Bull. Soc. Bot. Centre-Ouest, NS, 22 : 408) nom. inval. et illeg.] (Noble & Baret, 2021, tableau 38 : colonne 6) ; photo 13.
RQc6. Clematido flammulae-Osyrietum albae O. Bolòs 1962 (Sivim, consulté le 19/09/2020).
RQc7. Crataego monogynae-Quercetum cocciferae Martínez-Parras, Peinado & Alcaraz 1984 (Sivim, consulté le 19/09/2020).
RQc8. Daphno gnidii-Quercetum cocciferae Rivas Mart., Cantó, Fern.Gonz. & Sánchez-Mata in Rivas Mart. et al. 2002 (Sivim, consulté le 19/09/2020).
RQc9. Genistetum valentinae Alcaraz, P. Sánchez, De la Torre, Ríos & J. Alvarez 1991 (Sivim, consulté le 19/09/2020).
RQc10. « Teucrio compacti-Quercetum cocciferae Martínez-Parras, Peinado & Alcaraz 1987 » (Sivim, consulté le 19/09/2020).
RQc11. Genisto occidentalis-Quercetum cocciferae Loidi, Herrera, Olano & Silván 1994 (Loidi et al., 1994, tableau 1 : relevés 48 à 51).
RQc12. Genisto scorpii-Retametum sphaerocarpae Rivas Mart. ex Fuente 1986 (Sivim, consulté le 19/09/2020).
RQc13. Hedero helicis-Cytisetum heterochroi Mateo 1983 (Sivim, consulté le 14/09/2020).
RQc14. « Hyacinthoido hispanicae-Quercetum cocciferae (Rivas Goday in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960) Peinado & Martínez-Parras 1985 » (Sivim, consulté le 19/09/2020).
RQc15. Groupement audois à Crataegus azarolus (de Foucault, 2016 : 138, liste en colonne de gauche et relevé quantifié en colonne de droite).
RQc16. Phillyreo latifoliae-Pistacietum terebinthi Pavón Núñez, Hidalgo Triana & Pérez Latorre 2013 (Pavón Núñez et al., 2013, tableau 1).

* Groupe de syntaxons à Myrtus communis et Clematis cirrhosa, surtout actuellement connus de Corse et Sardaigne
RQc17. Clematido cirrhosae-Pistacietum lentisci Gamisans & Muracciole 1984 (Gamisans & Muracciole, 1984, tableau 13).
RQc18. Pistacio lentisci-Oleetum europaeae Paradis, Féral, Passigny-Hernandez, Nicolau & Carles 2014 nom. illeg. [non Oleo europaeae-Pistacietum lentisci Krause, Ludwig & Seidel 1963) (Paradis et al., 2014, tableau 12).
RQc19. Phillyreo latifoliae-Myrtetum communis Hugot & Panaïotis in Reymann, Panaïotis & Bioret 2017 (Reymann et al., 2017 : 148).
RQc20. « Cyclamino repandi-Oleetum sylvestris » Bacchetta, Bagella, Biondi, Farris, Filigheddu & Mossa 2003 (Bacchetta et al., 2003, tableau 1).
RQc21. Myrto communis-Oleetum europaeae Bacchetta, Bagella, Biondi, Farris, Filigheddu & Mossa 2003 (Ibid., tableau 2).
RQc22. Crataego monogynae-Pistacietum lentisci Biondi, Filigheddu & Farris 2001 (Biondi et al., 2001, tableau 58).


Alliance AJp. Amelanchiero ovalis-Juniperion phoeniceae all. nov. hoc loco ; tableau 1 : colonne AJp ; tableau 11
Typus nominis hoc loco : Buxo sempervirentis-Juniperetum phoeniceae Rivas Mart. 1969 (Publ. Inst. Biol. Apl. XLVI, non consulté).
Fourrés fortement xérophiles et souvent inféodés aux fentes des falaises, surtout calcaires, homologues des fourrés plus septentrionaux de l’Amelanchiero ovalis–Buxion sempervirentis au sein des Rhamno-Prunetea spinosae, caractérisés par Juniperus phoenicea subsp. ph., Buxus sempervirens, Amelanchier ovalis subsp. o., Quercus coccifera, Rhamnus saxatilis subsp. s.
AJp1. Jasmino fruticantis-Buxetum sempervirentis O. Bolòs 1973 (de Bolòs, 1973, tableau 6).
AJp2. a – Rhamno lycioidis-Quercetum cocciferae Braun-Blanq. & O. Bolòs 1954 (Sivim, consulté le 19/09/2020) ; b – Rhamno lycioidis-Juniperetum phoeniceae Rivas Mart. & G. López in G. López 1976 (Sivim, consulté le 19/09/2020) ; la comparaison de ces deux colonnes ne montre guère de différence significative entre ces deux syntaxons, de sorte qu’on derait inclure le second dans le premier, prioritaire.
AJp3. Spiraeo obovatae-Quercetum cocciferae Loidi & Fern.Prieto 1986 (Sivim, consulté le 19/09/2020).
AJp4. Rhamno saxatilis-Juniperetum phoeniceae Roselló 1994 (Roselló, 1994, tableau 99).
AJp5. Groupement à Buxus sempervirens (López Vélez, 1996, tableau 12).
AJp6. Amelanchiero ovalis-Ericetum multiflorae Géhu, Biondi & Bournique 1992 (Géhu et al., 1992, tableau 5).
AJp7. Buxo sempervirentis-Juniperetum phoeniceae Rivas Mart. 1969 (incl. Junipero phoeniceae-Amelanchieretum ovalis B. Foucault 1991) ; a – race ibérique différenciée par Quercus rotundifolia j (Sivim, consulté le 14/09/2020) ; b – race franco-méditerranéenne à Pistacia terebinthus, Prunus mahaleb et Quercus ilex j (de Foucault, 1991a : 84 ; de Foucault & Julve, 1991, tableau 6 : relevés 1 à 3, sub Junipero phoeniceae-Amelanchieretum ovalis) ; c – race franco-méditerranéenne à Globularia alypum et Quercus ilex j (Noble & Baret, 2021, tableau 38, colonne 8). Reconnu sous le nom invalide et illégitime de 1991 par de Foucault (2020c : 128) ; cité aussi par Choisnet (2019 : 89) ; photo 14.
AJp8. Junipero phoeniceae-Ficetum caricae B. Foucault & Julve 1991 (de Foucault & Julve, 1991 : 408), connu du seul relevé type ; dérivant du Buxo sempervirentis-Juniperetum phoeniceae sous l’effet de l’eutrophisation des parois calcaires plus naturelles.
Sur le revers sud de la Montagne noire audoise, il existe de même un fourré thermophile de rocaille affleurante dont le tableau 12 rapporte cinq relevés et une liste floristique inédits réalisés en divers points au-dessus de Cabrespine (Aude, le 23 avril et le 10 juin 2017, puis le 29 septembre 2019 ; au-dessus du village, Costo Granié, Mourral Blanc, la Condamine, au-dessus du parking du gouffre, ), entre 360 m et 600 m d’altitude. Il se distingue du Buxo sempervirentis-Juniperetum phoeniceae, présent dans l’Aude surtout au niveau des Corbières calcaires, par Rhamnus saxatilis subsp. s., Bupleurum fruticosum, Lonicera implexa, ainsi que Erica arborea. On le décrit ici sous le nom de Bupleuro fruticosi-Juniperetum phoeniceae ass. nov. hoc loco, typus nominis hoc loco : relevé 2 du tableau 12 hoc loco.
Entre 1 150 et 1 750 m d’altitude, le « Junipero phoeniceae-Pinetum mauretanicae buxetosum sempervirentis » décrit par Gomez Mercado (2011, tableau 77) paraît constituer un relais entre l’Amelanchiero–Juniperion phoeniceae (Juniperus phoenicea subsp. ph., J. oxycedrus subsp. o., Rhamnus myrtifolia) et l’Amelanchiero–Buxion sempervirentis (Lonicera splendida, Rosa canina, R. pouzinii), en lien avec une végétation arborescente à Pinus nigra subsp. salzmannii, Quercus rotundifolia, Q. faginea subsp. f., Sorbus aria, Acer granatense (Quercion lusitanico-rotundifoliae ; de Foucault, 2020a).
À propos de cette alliance à Juniperus phoenicea et Buxus sempervirens, on peut aussi évoquer le « Digitali laciniatae-Buxetum balearicae Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 » (Quézel et al., 1988, tableau 6), à Juniperus phoenicea et Buxus balearica, difficilement classable dans l’ensemble des alliances actuellement connues des Pistacio-Rhamnetalia alaterni (tableau 1 : colonne DBb).
Alliance Jt. Juniperion turbinatae Rivas Mart. 1975 nom. mut. hoc loco (Anales Inst. Bot. Cavanilles 31 (2) : 215, sub Juniperion lyciae) ; tableau 1 : colonne Jt ; tableau 13
Typus allianciae : Rhamno oleoidis-Juniperetum macrocarpae Rivas Mart. 1965 (Anales Inst. Bot. Cavanilles 22 : 399) désigné par l’auteur de l’alliance (1975, Anales Inst. Bot. Cavanilles 31 (2) : 215, sub Rhamno oleoidis-Juniperetum lyciae Rivas Mart. 1965).
Fourrés des arrière-dunes et des falaises exposées du littoral méditerranéen occidental à Juniperus phoenicea subsp. turbinata, J. oxycedrus subsp. macrocarpa et, en Espagne, Corema album !.
Jt1. Asparago acutifolii-Juniperetum macrocarpae Molin. & R. Molin. ex O. Bolòs 1962 (Sivim, consulté le 09/09/2020).
Jt2. « Astragalo tragacanthae-Juniperetum macrocarpae Rivas Mart. & Cantó in Rivas Mart. et al. 2002 » (Sivim, consulté le 09/09/2020).
Jt3. Juniperetum turbinatae O. Bolòs, Molin. & P. Monts. 1970 (Acta Bot. Barcinon. 5 : 36) non Juniperetum turbinatae Molin. ex O. Bolòs 1967 (de Bolòs et al., 1970, tableau 4), renommé ultérieurement Clematido cirrhosae-Juniperetum turbinatae (O. Bolòs, Molin. & P. Monts. 1970) Rivas Mart. 1975.
Jt4. Coremato albi-Juniperetum macrocarpae M.B. Crespo, De la Torre, Alcaraz, Costa & Solanas in Rivas Mart. et al. 2002 (Sivim, consulté le 09/09/2020).
Jt5. Daphno gnidii-Juniperetum transtaganae Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990 nom. mut. hoc loco (Sivim, consulté le 09/09/2020, sub Daphno gnidii-Juniperetum navicularis).
Jt6. Juniperetum turbinatae Molin. ex O. Bolòs 1967 (Sivim, consulté le 09/09/2020).
Jt7. Osyrio lanceolatae-Juniperetum turbinatae Rivas Mart. ex Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990 nom. mut. hoc loco (Sivim, consulté le 09/09/2020, sub Osyrio quadripartitae-Juniperetum turbinatae).
Jt8. Rhamno oleoidis-Juniperetum macrocarpae Rivas Mart. 1965 (Sivim, consulté le 09/09/2020).
Jt9. Rubio longifoliae-Juniperetum macrocarpae Rivas Mart., M.J. Costa & Loidi 1992 (Sivim, consulté le 09/09/2020).
Jt10. Groupement à Juniperus oxycedrus subsp. macrocarpa (Géhu et al., 1987, tableau 7).
Jt11. Smilaco asperae-Juniperetum turbinatae (Géhu, Biondi, Géhu-Franck & Taffetani 1987) Julve 1993 prov. nom. mut. hoc loco (Géhu et al., 1987, tableau 8, sub Smilaco mauritanicae-Juniperetum turbinatae).
Jt12. Querco cocciferae-Juniperetum turbinatae (Rivas Mart. 1975) Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990 (Sivim, consulté le 14/09/2020).
Jt13. Oleo europaeae-Juniperetum turbinatae Arrigoni et al. in De Marco et al. 1985 (Géhu & Biondi, 1994, tableau 62, sub Oleo sylvestris-Juniperetum turbinatae) ; cité aussi par Paradis et al. (2014, tableau 11).
Jt14. Rhamno angustifoliae-Juniperetum turbinatae Rivas Mart. ex Freitag 1971 corr. Alcaraz, T.E. Díaz, Rivas Mart. & P. Sánchez 1989 (Díez Garretas & Asensi, 2013, tableau I : colonne 5).
Jt15. Phillyreo angustifoliae-Juniperetum turbinatae Arrigoni, Nardi & Raffaelli 1985 (Díez Garretas & Asensi, 2013, tableau I : colonne 8) ; cité par Noble & Baret (2021) ; c’est sans doute à cette association qu’il faut rattacher la végétation à Juniperus phoenicea subsp. turbinata observée sur les arrière-dunes languedociennes (de Foucault & Royer, 2019, et photo 15).
Jt16. Euphorbio characiae-Juniperetum turbinatae Biondi, Filigheddu & Farris 2001 (Biondi et al., 2001, tableau 55).
Jt17. Chamaeropo humilis-Juniperetum turbinatae De Marco, Dinelli & Caneva 1985 (Biondi et al., 2001, tableau 56).
Jt18. Ephedro fragilis-Juniperetum macrocarpae Bartolo, Brullo & Marcenò 1982 (Géhu & Géhu-Franck, 1986, tableau 15 ; Géhu et al., 1992b, tableau 11).
Jt19. Ephedro fragilis-Juniperetum turbinatae Géhu & Sadki 1995 (Géhu & Sadki, 1995, tableau 8).
Il est probable que l’Asparago acutifolii-Anthyllidetum barbae-jovis Géhu, Biondi & Bournique 1992 relève aussi de cette alliance, mais la sous-espèce de Juniperus phoenicea n’est pas précisée (Géhu et al., 1992c, tableau 7).
Bien caractérisée écologiquement, cette alliance l’est assez faiblement au plan floristique, si bien qu’il est parfois difficile de classer certains syntaxons à J. phoenicea subsp. turbinata développés en situation non arrière-dunaire (cf. supra, syntaxons ARo16, ARo17 et ARo18, et infra, Oleo europaeae-Paliurion spinae-christi). Par ailleurs, certaines associations pourraient être rattachées à des alliances pas spécialement littorales, ainsi le Clematido cirrhosae-Juniperetum turbinatae (Jt3) dans le Clematido cirrhosae-Pistacion lentisci ou le Phillyreo angustifoliae-Juniperetum turbinatae (Jt15) dans le Rhamno lycioidis-Quercion cocciferae.
D’autres associations non synthétisées ici ont été compilées par Díez Garretas & Asensi (2013).

Alliance AJm. Phillyreo mediae-Juniperion turbinatae all. nov. hoc loco ; tableau 1 : colonne PJt
Typus nominis hoc loco : Pino pinastri-Juniperetum turbinatae Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia 9 (3) : 368, sub Pino pinastri-Juniperetum lyciae).
Fourrés arrière-dunaires du littoral marocain ; alliance proche de la précédente par Juniperus phoenicea subsp. turbinata, J. oxycedrus subsp. macrocarpa, mais s’en distinguant par Osyris lanceolata, Chamaerops humilis, Rubia peregrina subsp. longifolia, Phillyrea media, Olea europaea subsp. e. j.
Synthèse des Pino pinastri-Juniperetum turbinatae Barbero, Quézel & Rivas Mart. 1981 nom. mut. hoc loco (Barbero et al., 1981, tableau 22, sub Pino pinastri-Juniperetum lyciae), Clematido cirrhosae-Juniperetum turbinatae sensu Barbero, Quézel & Rivas Mart. 1981 [Ibid. : 369, sub Clematido cirrhosae-Juniperetum lyciae, non Clematido cirrhosae-Juniperetum turbinatae (O. Bolòs, Molin. & P. Monts. 1970) Rivas Mart. 1975 (cf supra, Jt3 des Baléares) ; à renommer (art. 39), mais sans possibilité de le typifier sur les données publiées, synthétiques] et Rhamno oleoidis-Juniperetum turbinatae Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 nom. mut. hoc loco (Quézel et al., 1988, tableau 9, sub Rhamno rotundifoliae-Juniperetum turbinatae.
Alliance OPs. Oleo europaeae-Paliurion spinae-christi (Biondi, Casavecchia, Biscotti & Pesaresi 2014) stat. nov. hoc loco ; tableau 1 : colonne OPs ; tableau 14
[corresp. syntax. : Oleo europaeae-Paliurenion spinae-christi Biondi, Casavecchia, Biscotti & Pesaresi in Biondi, Casavecchia, Pesaresi, Gangale & Uzunov 2014 (Pl. Biosyst. 148 (4) : 726, sub Oleo sylvestris-Paliurenion spinae-christi)].
Typus nominis hoc loco : Oleo europaeae-Paliuretum spinae-christi Biondi, Casavecchia, Biscotti & Pesaresi 2014 in Biondi, Casavecchia, Pesaresi, Gangale & Uzunov 2014 (Pl. Biosyst. 148 (4) : 726, sub Oleo sylvestris-Paliuretum spinae-christi) désigné par Biondi et al. (2014) pour leur sous-alliance.
Fourrés italiens réalisant le passage des Eu-Pistacio lentisci-Rhamnenea alaterni aux Rubio tenuifoliae-Pistacienea terebinthi à Paliurus spina-christi, Pyrus spinosa, Calicotome spinosa subsp. s., Cytisus spinescens, Rhamnus saxatilis subsp. s., Hippocrepis emerus subsp. emeroides ; cartographie chorologique in Casavecchia et al. (2015, figure 2).
* Groupe d’associations peu différenciées
OPs1. Calicotomo infestae-Paliuretum spinae-christi Casavecchia, Biscotti, Pesaresi & Biondi 2015 (Casavecchia et al., 2015, tableau 2).
OPs2. Oleo europaeae-Paliuretum spinae-christi Biondi, Casavecchia, Biscotti & Pesaresi 2014 nom. mut. hoc loco (Ibid., tableau 1 : colonne 3, sub Oleo sylvestris-Paliuretum spinae-christi).
OPs3. Pistacio terebinthi-Paliuretum spinae-christi Blasi & Di Pietro 2001 (Ibid., tableau 1 : colonne 5).
OPs4. Rhamno saxatilis-Paliuretum spinae-christi Biondi 2001 (Ibid., tableau 1 : colonne 8).
OPs5. Rhamno intermediae-Paliuretum spinae-christi Trinajstić 1996 (Ibid., tableau 1 : colonne 10).
* Groupe d’associations fortement différenciées par Pistacia lentiscus, Prasium majus, Cistus monspeliensis, Hippocrepis emerus subsp. emeroides, Juniperus oxycedrus subsp. macrocarpa, Daphne gnidium, Lonicera implexa, Coronilla valentina, surtout du sud-est de l’Italie
OPs6. « Cyclamino repandi-Paliuretum spinae-christi Biondi 2001 » (Casavecchia et al., 2015, tableau 1 : colonne 4).
OPs7. « Cyclamino repandi-Paliuretum spinae-christi Biondi 2001 » sensu Di Pietro & Misano 2010 (Di Pietro & Misano, 2010, tableau VIII).
OPs8. « Helictotricho convoluti-Pistacietum lentisci Di Pietro & Misano 2010 » (Ibid., tableau IV).
OPs9. Prasio majoris-Pistacietum lentisci Di Pietro & Misano 2010 (Ibid., tableau V).
OPs10. Juniperetum macrocarpo-turbinatae Di Pietro & Misano 2010 (Ibid., tableau VI).
OPs11. Phillyreo latifoliae-Calicotometum infestae Di Pietro & Misano 2010 (Ibid., tableau VII).
OPs12. Pistacio lentisci-Cistetum eriocephali Di Pietro & Misano 2010 (Ibid., tableau IX).
OPs13. « Charybdido pancratii-Cistetum monspeliensis Di Pietro & Misano 2010 » (Ibid., tableau X).
Alliance Jol. Juniperion oxycedro-lagunae Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (Acta Bot. Gallica 154 (1) : 95) ; tableau 1 : colonne Jol ; tableau 15
Typus allianciae : Pistacio terebinthi-Juniperetum lagunae Rodríguez Torres & Cano in Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (Acta Bot. Gallica 154 (1) : 368) désigné par les auteurs de l’alliance (2007, Acta Bot. Gallica 154 (1) : 95).
Fourrés du centre et du sud de la péninsule Ibérique à Juniperus oxycedrus subsp. lagunae (Pau ex C. Vicioso) Rivas Mart., Cytisus striatus subsp. eriocarpus (Boiss. & Reut.) Rivas Mart. (deux taxons ignorés de Euro+Med Plantbase), Cistus ladanifer subsp. l., Quercus rotundifolia j, un peu à la limite des Cytiso-Genistetalia monspessulanae et des Cytisetea scopario-striati.
Jol1. Echinosparto iberici-Juniperetum lagunae Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (Cano et al., 2007, tableau I).
Jol2. Cytiso eriocarpi-Juniperetum lagunae Pinto Gomes & Cano in Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (Ibid., tableau II).
Jol3. Pistacio terebinthi-Juniperetum lagunae Rodríguez Torres & Cano in Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (Ibid., tableau III).
Jol4. « Stipo tenacissimae-Juniperetum lagunae Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 » (Ibid., tableau IV).
Jol5. Groupement à Phlomis purpurea-Juniperus oxycedrus subsp. lagunae (Ibid., tableau V).
On retrouve ce type de végétation en strate arbustive du Smilaco asperae-Quercetum suberis décrit par Pinto Gomes et al. (2003, tableau I).
Les alliances qui suivent, propres au Maghreb, possèdent en commun Tetraclinis articulata j (photo 16) ; en outre, c’est à ce niveau que se trouve l’optimum de Globularia alypum pour la classe (tableau 1). L’Euphorbio bivonae-Searsietum tripartitae décrit par Siab-Farsi et al. (2014, tableau 4, sub Euphorbio bivonae-Rhoetum tripartiti, ‘…Rhusetum’) ne rentre bien dans aucune des six alliances qui suivent.
Alliance PCl. Prasio majoris-Cytision lanigeri all. nov. hoc loco ; tableau 1 : colonne PCl ; tableau 16
Typus nominis hoc loco : Cytision lanigeri-Myrtetum communis B. Foucault & Sadki nov.
Fourrés tunisiens et nord-est-algériens à Prasium majus, Cytisus laniger, Ampelodesmos mauritanicus, Tetraclinis articulata j.
* Groupe de syntaxons à Juniperus phoenicea subsp. ph.
PCl1. Groupement à Quercus coccifera (Vanden Berghen, 1990, tableau 1).
PCl2. Groupement à Tetraclinis articulata j et Juniperus phoenicea (Ibid., tableau 4).
PCl3. Groupement à Rosmarinus officinalis et Juniperus phoenicea (Ibid., tableau 5).
* Groupe de syntaxons à Dioscorea communis, Asparagus acutifolius, Clematis cirrhosa, Rhamnus alaternus subsp. a.
PCl4. Cytiso lanigeri-Myrtetum communis B. Foucault & Sadki ass. nov. hoc loco (Sadki, 1995, tableau III), fourré nord-est-algérien à Myrtus communis, Cytisus laniger, Rubia peregrina subsp. p., Clematis cirrhosa, Pistacia lentiscus, Olea europaea subsp. e. j, Phillyrea angustifolia, Cistus monspeliensis ; typus nominis hoc loco : relevé 14 du tableau III in Sadki (1995, Doc. Phytosoc., NS, XV h. t.).
PCl5. Groupement à Cistus villosus (« Tetraclinido articulatae-Cyclaminetum persici El Hamrouni & R.J. Loisel 1979 » ; El Hamrouni & Loisel, 1979, tableau 1).
PCl6. Prasio majoris-Ericetum multiflorae ass. nov. hoc loco (Guinochet, 1980, tableau 6, sub Prasio-Oleetum Bolòs & Molin. 1969 tetraclinidetosum), typus nominis hoc loco : relevé 15 du tableau 6 in Guinochet (1980, Phytocoenologia 7 : 450).

Alliance TQc. Tetraclinido articulatae-Quercion cocciferae (Hadjadj & R.J. Loisel 1999) stat. nov. hoc loco ; tableau 1 : colonne PCl ; tableau 17
[corresp. syntax. : Tetraclinido-Quercenion cocciferae Hadjadj & R.J. Loisel 1999 (Doc. Phytosoc., NS, XIX : 280) nom. inval. (art. 2b, 8)]
Typus nominis hoc loco : Osyrio lanceolatae-Quercetum cocciferae Hadjadj & R.J. Loisel 1999 (Doc. Phytosoc., NS, XIX : 240).
Fourrés nord-ouest-algériens à Rubia peregrina subsp. longifolia, Ampelodesmos mauritanicus, Cistus heterophyllus, C. monspeliensis, Erica multiflora, Genista tricuspidata, Cytisus spinosus, C. laniger, Arbutus unedo, Quercus coccifera, Chamaerops humilis, Tetraclinis articulata j.
TQc1. Osyrio lanceolatae-Quercetum cocciferae Hadjadj & R.J. Loisel 1999 nom. mut. hoc loco (Hadjadj & Loisel, 1999, tableau 3, sub Osyrio quadripartitae-Quercetum cocciferae).
TQc2. Lonicero implexae-Quercetum cocciferae Nègre 1964 ; a – d’après Nègre (1964, tableau 7) ; b – d’après Hadjadj & Loisel (1999, tableau 4) ; quelques différences synfloristiques entre ces deux syntaxons.
TQc3. Oleo europaeae-Tetraclinidetum articulatae Hadjadj & R.J. Loisel 1999 nom. mut. hoc loco (Ibid., tableau 5, sub Oleo sylvestris-Tetraclinidetum articulatae).
TQc4. Junipero oxycedri–Quercetum cocciferae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 6).
TQc5. Tetraclinido articulatae-Quercetum cocciferae Benabid & Fennane 1994 (Benabid & Fennane, 1994, tableau 1.
Alliance GCs. Genisto tricuspidatae-Cytision spinosi B. Foucault & Dahmani all. nov. hoc loco ; tableau 1 : colonne GCs ; tableau 18
[syn. : Genisto tricuspidatae-Calicotomion spinosae Dahmani 1997 (Le chêne vert en Algérie… : 118) nom. ined.]
Typus nominis hoc loco : Calicotomo spinosae-Quercetum rotundifoliae Dahmani & B. Foucault nov.
GCs1. Groupement à Quercus coccifera-Pistacia lentiscus (Miara et al., 2020, tableau F3).
GCs2. Groupement à Quercus rotundifolia-Juniperus oxycedrus subsp. o. (Miara et al., 2012, tableaux 1 et 4 ; Miara et al., 2020, tableaux F4 et M1).
GCs3. Querco rotundifoliae-Arbutetum unedonis Miara, Hadjadj Aoul & Decocq 2020 (Miara et al., 2020, tableau M2), incluant le groupement à Quercus rotundifolia-Arbutus unedo de Miara et al. (2012, tableau 3).
GCs4. Querco suberis-Ampelodesmetum mauritanici Miara, Hadjadj Aoul & Decocq 2020 (Ibid., 2020, tableau M4), incluant le groupement à Quercus suber-Q. faginea subsp. baetica de Miara et al. (2012, tableau 5).
GCs5. Groupement à Pistacia atlantica-Asparagus albus (Miara et al., 2020, tableau M5).
GCs6. « Groupement à Fumana laevis-Tripodion tetraphyllum » (Ibid., tableau M6), incluant le groupement à Tetraclinis articulata-Ampelodesmos mauritanicus de Miara et al. (2012, tableau 2).
GCs7. « Groupement à Rosmarinus eriocalyx et Cistus creticus » (Miara et al., 2020, tableau M7).
GCs8. Calicotomo spinosae-Quercetum rotundifoliae Dahmani & B. Foucault ass. nov. hoc loco [syn. : Calicotomo spinosae-Quercetum rotundifoliae Dahmani 1997 (Le chêne vert en Algérie… : 142) nom. ined.] (Dahmani, 1997, tableau 16) ; validé ici par le relevé GCs8t du tableau 18 hoc loco, composante arbustive du relevé 4 du tableau 16 in Dahmani (1997, Le chêne vert en Algérie… : 143 ; 10 taxons).
Le « Cisto salviifolii–Quercetum rotundifoliae Dahmani 1997 nom. ined. » (Dahmani, 1997, tableau 18), fort de quarante relevés, paraît cependant assez hétérogène, avec au moins une partie plus acidiphile à rapprocher des Cytiso-Genistetalia monspessulanae.
Alliance GPl. Genisto quadriflorae-Pistacion lentisci all. nov. hoc loco ; tableau 1 : colonne GPl ; tableau 19
[corresp. syntax. : Asparago-Rhamnenion oleoidis Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia, NS, 9 (3) : 347), à renommer en Eu-Asparago-Rhamnenion oleoidis]
Typus nominis hoc loco : Junipero turbinatae-Quercetum cocciferae Hadjadj & R.J. Loisel 1999 (Doc. Phytosoc., NS, XIX : 275).
Fourrés du nord-ouest de l’Algérie et du Maroc nord-oriental à Ampelodesmos mauritanicus, Genista quadriflora, Calicotome infesta subsp. intermedia, Globularia alypum, Chamaerops humilis, Quercus rotundifolia j, Tetraclinis articulata j.
GPl1. Calicotomo intermediae-Tetraclinidetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Barbero et al., 1981 : 353 et tableau 15 ; Hadjadj & Loisel, 1999, tableaux 11 et 12) ; Barbero et al. (1981) donnent le même nom aux deux syntaxons décrits par leurs tableaux 14 et 15 ; selon la présente synthèse, celui du tableau 15 apparaît comme distinct donc homonyme illégitime du Calicotomo intermediae-Tetraclinidetum articulatae Barbero, Quézel & Rivas Mart. 1981 retenu ici (cf. infra syntaxon TPa4) ; on le renomme alors (art. 39) comme Calicotomo intermediae-Chamaeropetum humilis (Barbero, Quézel & Rivas Mart. 1981) nom. nov. hoc loco, typus nominis hoc loco : le même que celui proposé par les auteurs du nom rejeté, soit le relevé 1 du tableau 15 in Barbero et al. (1981, Phytocoenologia 9 (3) : 353).
GPl2. Quercetum coccifero-rotundifoliae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 13).
GPl3. Ampelodesmo mauritanici-Tetraclinidetum articulatae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 14).
GPl4. Genisto quadriflorae-Tetraclinidetum articulatae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 15).
GPl5. Junipero turbinatae-Quercetum cocciferae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 18).
GPl6. Ampelodesmo mauritanici-Chamaeropetum humilis Quézel, Barbero, Benabid & Rivas Mart. 1992 (Quézel et al., 1992, tableau 5) ; homonyme illégitime de l’Ampelodesmo mauritanici-Chamaeropetum humilis Braun-Blanq., Font Quer, G. Braun-Blanq., Frey, Jansen & Moor 1936 (cf. supra syntaxon ARo1), renommé ici (art. 39) Genisto quadriflorae–Chamaeropetum humilis (Quézel, Barbero, Benabid & Rivas Mart. 1992) nom. nov. hoc loco, typus nominis hoc loco : relevé 8 du tableau 5 in Quézel et al. (1992, Stvdia Botanica 10 : 81 ; impossible de reprendre le type de l’association initiale, c’est-à-dire le relevé 4, l’un des plus pauvres floristiquement des onze relevés de ce tableau : trois taxons caractéristiques de la classe, contre huit pour celui retenu ici).
Alliance TPa. Tetraclinido articulatae-Pistacion atlanticae Rivas Mart., M.J. Costa & Izco 1986 (Notiz. Soc. Ital. Fitosoc.19 (2) : 76) ; tableau 1 : colonne TPa ; tableau 20
[corresp. syntax. : Pistacienion atlanticae Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia 9 (3) : 352)]
Typus allianciae : Coronillo viminalis-Tetraclinetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia 9 (3) : 355) désigné par les auteurs de l’alliance (1986, Notiz. Soc. Ital. Fitosoc.19 (2) : 84).
Fourrés maghrebins substeppiques des étages thermo-méditerranéen à méso-méditerranéen inférieur sous climat aride à sec continental, à Searsia tripartita, Cistus villosus, Asparagus stipularis, A. altissimus, A. albus, Calicotome infesta subsp. intermedia, Aristolochia baetica, Withania frutescens, Genista tricuspidata, Pistacia atlantica j, Tetraclinis articulata j.
TPa1. Searsio pentaphyllae-Tetraclinidetum articulatae Hadjadj ex Hadjadj & R.J. Loisel 1999 nom. mut. hoc loco (Hadjadj & Loisel, 1999, tableau 16, sub Rhoo pentaphyllae-Tetraclinidetum articulatae) ; syn. : Rhoo pentaphyllae-Tetraclinidetum articulatae Hadjadj 1991 (Ecol. Medit. 17 : 73) nom. inval.
TPa2. Rhamno oleoidis-Tetraclinidetum articulatae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 17).
TPa3. Clematido cirrhosae-Ceratonietum siliquae Barbero, Quézel & Rivas Mart. 1981 (Barbero et al., 1981, tableau 13).
TPa4. Calycotomo intermediae-Tetraclinetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Ibid., tableau 14).
TPa5. Searsio pentaphyllae-Pistacietum atlanticae Barbero, Quézel & Rivas Mart. 1981 nom. mut. hoc loco (Ibid., tableau 16, sub Rhoo pentaphyllae-Pistacietum atlanticae).
TPa6. Coronillo viminalis-Tetraclinidetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Ibid., tableau 17).
TPa7. Phillyreo latifoliae-Oleetum europaeae Barbero, Quézel & Rivas Mart. 1981 nom. mut. hoc loco (Ibid., tableau 18, sub Phillyreo latifoliae-Oleetum sylvestris).
TPa8. Periploco laevigatae-Tetraclinidetum articulatae Benabid 1982 (Fennane, 1986, tableau II).
TPa9. Polygalo balansae-Tetraclinetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Barbero et al., 1981, tableau 19).
TPa10. Ceratonio siliquae-Tetraclinidetum articulatae Fennane 1986 (Fennane, 1986, tableau III).
TPa11. « Rosmarino tournefortii-Tetraclinidetum articulatae Fennane 1986 » (Ibid., tableau IV).
TPa12. Phillyreo mediae-Tetraclinidetum articulatae Fennane 1986 (Ibid., tableau VI).
TPa13. Lonicero implexae-Tetraclinidetum articulatae Fennane 1986 (Ibid., tableau VII).
TPa14. Calicotomo intermediae-Oleetum europaeae Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 nom. mut. hoc loco (Quézel et al., 1988, tableau 8, sub Calicotomo intermediae-Oleetum sylvestris).
TPa15. Pistacio atlanticae-Ziziphetum loti Benabid 1988 (Benabid, 1988, tableau I).
Avec la documentation limitée fournie par l’auteur (défaut de taxonomie fine de certains genres, faible nombre de relevés), il est difficile de placer le Chamaeropo humilis-Asparagetum altissimi Guin. 1980 (Guinochet, 1980, tableau 7).
Alliance Git. Genistion ifniensi-tricuspidatae all. nov. hoc loco ; tableau 1 : colonne Git ; tableau 21
Typus nominis hoc loco : Junipero phoeniceae-Tetraclinidetum articulatae Fennane ex B. Foucault & Fennane nov.
Fourrés de la région de l’arganeraie au Maroc, à Genista ifniensis, G. tricuspidata (incl. subsp. sparsiflora), Periploca laevigata, Asparagus pastorianus, A. albus, Cytisus albidus, Cistus villosus, Globularia alypum, Ceratonia siliqua j, Argania spinosa j, Tetraclinis articulata j.
Git1. Oleo maroccanae-Tetraclinidetum articulatae Fennane ex B. Foucault & Fennane ass. nov. hoc loco [syn. : Oleo salicifoliae-Tetraclinidetum articulatae Fennane 1986 (Bull. Inst. Sci. Rabat 10 : 73) nom. inval.], à Olea europaea subsp. maroccana (= O. salicifolia), Periploca laevigata, Tetraclinis articulata j, Genista tricuspidata, G. ifniensis, Argania spinosa j, Globularia alypum ; Fennane (1986, tableau X, sub Oleo salicifoliae-Tetraclinidetum articulatae) ; typus nominis hoc loco : relevé 139 du tableau X in Fennane (1986, Bull. Inst. Sci. Rabat 10 : 74).
Git2. Junipero phoeniceae-Tetraclinidetum articulatae Fennane ex B. Foucault & Fennane ass. nov. hoc loco [syn. : Junipero phoeniceae-Tetraclinidetum articulatae Fennane 1986 (Bull. Inst. Sci. Rabat 10 : 75) nom. inval.], à Juniperus phoenicea subsp. ph., Osyris lanceolata, Withania frutescens, Periploca laevigata, Phillyrea angustifolia, Chamaerops humilis, Globularia alypum, Tetraclinis articulata j ; Fennane (1986, tableau XII) ; typus nominis hoc loco : relevé 220 du tableau XII in Fennane (1986, Bull. Inst. Sci. Rabat 10 : 77).
Git3. Genisto tricuspidatae-Tetraclinidetum articulatae Fennane ex B. Foucault & Fennane ass. nov. hoc loco [syn. : Genisto sparsiflorae-Tetraclinidetum articulatae Fennane 1986 (Bull. Inst. Sci. Rabat 10 : 73) nom. inval.], à Jasminum fruticans, Chamaerops humilis, Olea europaea subsp. e. j, Cistus villosus, Quercus rotundifolia j, Ceratonia siliqua j ; Fennane (1986, tableau XI, sub Genisto sparsiflorae-Tetraclinidetum articulatae) ; typus nominis hoc loco : relevé 157 du tableau XI in Fennane (1986, Bull. Inst. Sci. Rabat 10).
Ordre 2. Cytiso villosi-Genistetalia monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 nom. mut. hoc loco (Itinera Geobot. 15 : 353, sub Cytiso villosi-Telinetalia monspessulanae) ; tableau 1 : colonnes PEa à HEa
Typus ordinis : Genistion monspessulano-linifoliae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 (Itinera Geobot. 15 : 353, sub Telinion monspessulano-linifoliae), seule alliance subordonnée à cet ordre in Rivas-Martínez et al. (2002, Itinera Geobot. 15 : 353).
Fourrés méditerranéens acidiphiles, souvent riches en Ericaceae, Fabaceae et Cistaceae nanophanérophytiques : Erica arborea, E. scoparia subsp. s., Arbutus unedo, Cistus monspeliensis, C. populifolius subsp. p., C. ladanifer subsp. l., Cytisus grandiflorus, C. spinosus, C. scoparius subsp. s. (optimum dans les Cytisetea scopario-striati), ainsi que Quercus suber j ; ordre initialement placé dans les Cytisetea scopario-striati, sans doute à cause de l’importance des Genisteae ; dans le tableau 1, Viburnum tinus apparaît aussi comme optimal dans cet ordre, il s’agit peut-être d’un effet statistique pour ce taxon normalement caractéristique de classe.
Alliance PEa. Pruno lusitanicae-Ericion arboreae all. nov. hoc loco ; tableau 1 : colonne PEa ; tableau 22
[corresp. syntax. : Arbuto unedonis-Laurenion nobilis (Rivas Mart., Fern.Gonz. & Loidi 1999) Rivas Mart. & Sánchez-Mata 1999 p. p. ; syn. : Arbuto unedonis-Laurion nobilis Rivas Mart., Fern.Gonz. & Loidi 1999 (Itinera Geobot. 13 : 400) p. p. (excluant le type)]
Typus nominis hoc loco : Frangulo alni-Prunetum lusitanicae Lopes, J.C. Costa, Lousã & Capelo in J.C. Costa, Lopes, Capelo & Lousã 2000 (Silva Lusit. 8 (2) non consulté).
Fourrés du nord-ouest et du nord de la péninsule Ibérique (du Minho portugais au Pays basque espagnol), à Prunus lusitanica subsp. l., Frangula alnus subsp. a., Castanea sativa j, partageant avec l’alliance suivante Lonicera periclymenum subsp. p. et subsp. hispanica, Salix atrocinerea, Ilex aquifolium, Quercus robur j, Laurus nobilis j. L’intégration à cette alliance du « Scrophulario laxiflorae-Rhododendretum baetici », de l’extrême sud de la péninsule, au sein de la sous-alliance du Rhododendrenion baetici Rivas Mart. & Sánchez-Mata 2001, ne nous paraît pas satisfaisant ; on tente ici l’intégration de cette association aux Cytiso villosi–Genistetalia monspessulanae (voir alliance suivante).
Le fourré à Prunus lusitanica du Pays basque français, en limite nord extrême d’extension du prunier, entre 400 et 520 m d’altitude, ne relève pas de cette alliance ; décrit sous le nom de Lonicero periclymeni-Prunetum lusitanicae par Raposo et al. (2021), il a été placé dans le Lonicerion periclymeni par ses auteurs, malgré l’absence de taxons caractéristiques des Rhamno-Prunetea spinosae en dehors du premier taxon éponyme de l’association. Le « Polysticho setiferi-Prunetum lusitanicae Barbero, Quézel & Rivas Mart. 1981 » n’en relève pas non plus et d’ailleurs n’entre pas dans les Pistacio-Rhamnetea alaterni (Barbero et al., 1981, tableau 32).
PEa1. Frangulo alni-Arbutetum unedonis T.E. Díaz & Fern.Prieto 1994 (Sivim, consulté le 10/09/2020).
PEa2. « Luzulo henriquesii-Prunetum lusitanicae Honrado, Alves, Lomba, Torres & Barreto 2007 » (Honrado et al., 2007, tableau I), renommé ici Ilici aquifolii-Prunetum lusitanicae (Honrado, Alves, Lomba, Torres & Barreto 2007) ass. nov. hoc loco, à Pyrus cordata, Ilex aquifolium, Frangula alnus subsp. a., Erica arborea, Lonicera periclymenum subsp. p., Prunus lusitanica subsp. l., typus nominis hoc loco : composante arbustive du relevé 5 du tableau I in Honrado et al. (2007, Acta Bot. Gallica 154 (1) : 72).
PEa3. Frangulo alni-Prunetum lusitanicae Lopes, J.C. Costa, Lousã & Capelo in J.C. Costa, Lopes, Capelo & Lousã 2000 (Costa et al., 2015, tableau 2).
PEa4. Morello fayae-Arbutetum unedonis Capelo & Mesquita 1998 nom. mut. hoc loco (Costa et al., 2015, tableau 6, sub Myrico fayae-Arbutetum unedonis).
PEa5. « Omphalodo nitidae-Lauretum nobilis Honrado, P. Alves & F.B. Caldas 2003 » (Rodríguez Guitián et al., 2007, tableau 5 : colonne 32).
PEa6. « Ulici gallii-Arbutetum unedonis Loidi, Herrera, Olano & Silván 1994 » (Sivim, consulté le 10/09/2020).
PEa7. Viburno tini-Prunetum lusitanicae Ladero 1976 (Sivim, consulté le 10/09/2020), irradiation méridionale de l’alliance vers l’Espagne centrale.
PEa8. « Vinco difformis-Lauretum nobilis Capelo, J.C. Costa in J.C. Costa, Lopes, Capelo & Lousã 2000 » (Costa et al., 2015, tableau 5).
PEa9. Bupleuro fruticosi-Arbutetum unedonis J.C. Costa et al. 2002 (Mesquita et al., 2005 : 74).
PEa10. « Holco mollis-Lauretum nobilis Rodríguez Guitián, Romero & Ramil 2007 » (Rodríguez Guitián et al., 2007, tableau 5 : relevés 22 à 30).
PEa11. Groupement à Prunus lusitanica (Braun-Blanquet et al., 1956 : 184).
PEa12. Pruno lusitanicae-Arbutetum unedonis (Aguiar & Capelo 1995) J.C. Costa, Capelo & Lousã in J.C. Costa, Lopes, Capelo & Lousã 2000 (Aguiar & Capelo, 1995, tableau 1, sub Frangulo alni-Arbutetum prunetosum lusitanicae).
Alliance Rb. Rhododendrion baetici (Rivas Mart. & Sánchez-Mata 2001) stat. nov. hoc loco ; tableau 1 : colonne Rb ; tableau 23
[corresp. syntax. : Rhododendrenion baetici Rivas Mart. & Sánchez-Mata 2001 (Lazaroa 21 : 151)]
Typus nominis hoc loco : Rhododendro baetici–Franguletum baeticae Rivas Goday & Rivas Mart. in Rivas Mart. 1964 (Anales Inst. Bot. Cavanilles 22 : 387).
Fourrés acidiphiles à Rhododendron ponticum subsp. baeticum, Frangula alnus subsp. baetica, Rubia peregrina subsp. agostinhoi, Quercus canariensis j, Q. faginea subsp. broteroi j, partageant avec l’alliance précédente Lonicera periclymenum subsp. p. et subsp. hispanica, Salix atrocinerea, Ilex aquifolium, Quercus robur j, Laurus nobilis j, de l’extrême Sud ibérique (cordillière Bétique), avec une extension ponctuelle dans le nord-ouest du Portugal (syntaxon Rb3).
Rb1. Rhododendro baetici–Franguletum baeticae Rivas Goday & Rivas Mart. in Rivas Mart. 1964 (Sivim, consulté le 10/09/2020).
Rb2. Groupement à Bryonia dioica-Rhododendron ponticum subsp. baeticum (Braun-Blanquet et al., 1956 : 190, infrapaginal).
Rb3. Calluno vulgaris-Rhododendretum baetici Honrado, Alves, Lomba, Torres & Barreto Caldas 2007 (Honrado et al., 2007, tableau II) ; déjà repéré par Braun-Blanquet et al. (1956 : 189).
Alliance Ql. Quercion lusitanicae Rothm. 1954 nom. mut. hoc loco (Vegetatio 5-6 : 599, sub Quercion fruticosae) ; tableau 1 : colonne Ql ; tableau 24
[syn. : Quercion fruticosae Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia 9 (3) : 370) nom. illeg.]
Typus allianciae : Frutici-Quercetum taganum Rothm. 1954 (Vegetatio 5-6 : 599) renommé ultérieurement Junipero navicularis-Quercetum lusitanicae (Rothm. 1954) Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990 (Itinera Geobot. 3 : 61).
Fourrés du sud-ouest de la péninsule Ibérique et du Tangérois marocain à Asparagus aphyllus, Juniperus oxycedrus subsp. transtagana (= J. navicularis), Erica australis, Corema album, Quercus lusitanica (= Q. fruticosa) ; cartographie chorologique in Capelo et al. (2001, figure 3).
Ql1. Junipero transtaganae-Quercetum lusitanicae (Rothm. 1954) Rivas Mart., Lousã, T.E. Díaz, Fern.Gonz. & J.C. Costa 1990 nom. mut. hoc loco (Sivim, consulté le 10/09/2020, sub Junipero navicularis-Quercetum lusitanicae).
Ql2. « Senecioni lopezii-Quercetum lusitanicae Rivas Mart. in Rivas Mart. et al. 2002 » (Sivim, consulté le 10/09/2020).
Ql3. Arbuto unedonis-Cistetum populifolii Braun-Blanq., P. Silva & Rozeira 1964 (Braun-Blanquet et al., 1964, tableau VII).
Ql4. Phillyreo angustifoliae-Ericetum scopariae Braun-Blanq., P. Silva & Rozeira 1964 (Ibid., 1964, tableau VIII).
Ql5. Erico scopariae-Quercetum lusitanicae Braun-Blanq., P. Silva & Rozeira 1964 (Ibid., 1964, tableau XII).
Ql6. « Centaureo crocatae-Quercetum lusitanicae Capelo & J.C. Costa in Capelo, J.C. Costa, Lousã & Mesquita 2001 » (Capelo et al., 2001, tableau II).
Ql7. Phillyreo angustifoliae-Quercetum lusitanicae Barbero, Quézel & Rivas Mart. 1981 (Barbero et al., 1981, tableau 23, sub Phillyreo angustifoliae-Quercetum fruticosae).
On pourrait encore en rapprocher une partie du « Lavandulo viridis-Quercetum suberis » décrit par Quinto-Canas et al. (2010, tableau II).
Alliance Ea. Ericion arboreae (Rivas Mart. ex Rivas Mart., M.J. Costa & Izco 1986) Rivas Mart. 1987 (ICONA, serie Técnica, non consulté), incl. Myrtion communis Allier & Lacoste 1980 (Ecol. Medit. 5 : 71) nom. inval. ; tableau 1 : colonne Ea ; tableau 25
[corresp. syntax. : Ericenion arboreae Rivas Mart. ex Rivas Mart., M.J. Costa & Izco 1986 (Notiz. Soc. Ital. Fitosociol. 19 (2) : 82)]
Typus allianciae : Phillyreo angustifoliae-Arbutetum unedonis Rivas Goday & Galiano in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Anales Inst. Bot. Cavanilles 17 (2) : 352) désigné par Rivas-Martínez et al. (1986, Notiz. Soc. Ital. Fitosociol. 19 (2) : 82) pour leur sous-alliance.
Fourrés acidiphiles plus ou moins méditerranéens partageant avec l’alliance suivante Erica scoparia subsp. s., Cistus monspeliensis, Quercus coccifera, Q. ilex j, mais plus pauvres en espèces.
* Groupe de syntaxons fortement différenciés par Lonicera implexa, Myrtus communis, Smilax aspera…, surtout ibériques
– et Cistus albidus, C. populifolius subsp. p., Phillyrea latifolia, Pistacia terebinthus, Quercus rotundifolia j
Ea1. Cytiso heterochroi-Ericetum arboreae López Vélez 1996 (López Vélez, 1996, tableau 6).
Ea2. Cytisophyllo sessilifolii-Bupleuretum fruticosi Rivas Mart. 1969 (Sivim, consulté le 8/09/2020) ; avec Buxus sempervirens, Amelanchier ovalis subsp. o. et moins souvent Juniperus phoeniceae subsp. ph., peut-être mieux dans l’Amelanchiero ovalis-Juniperion phoeniceae ?
Ea3. Erico scopariae-Arbutetum unedonis Ortiz, Amigo & Izco 1991 (Sivim, consulté le 8/09/2020).
Ea4. Ericetum scopario-arboreae Mateo 1983 (Sivim, consulté le 8/09/2020).
Ea5. Phillyreo angustifoliae-Arbutetum unedonis Rivas Goday & Galiano in Rivas Goday, Borja, Esteve, Galiano, Rigual & Rivas Mart. 1960 (Sivim, consulté le 09/09/2020).
Ea6. « Bupleuro rigidi-Arbutetum unedonis Gómez-Mercado 2011 » (Gómez-Mercado, 2011, tableau 82).
Ea7. Erico arboreae-Buxetum sempervirentis Aguiar, Esteves & Penas 1999 (Aguiar et al., 1999, tableau 1)
Ea8. Pistacio lentisci-Juniperetum oxycedri Camarda, Lucchese, E. Pignatti & Pignatti 1995 (Camarda et al., 1995, tableau 10 : relevé 58-68).
– autres, à Cytisus laniger
Ea9. Ampelodesmo mauritanici-Ericetum scopariae O. Bolòs, Molin. & P. Monts. 1970 (Sivim, consulté le 8/09/2020).
Ea10. Erico arboreae-Arbutetum unedonis Molin. 1937 (Molinier, 1937 : 110 et 111 ; Lavagne, 1972, tableau III).
Ea11. Erico arboreae-Arbutetum unedonis sensu Allier & Lacoste 1980 (Allier & Lacoste, 1980, tableau III : relevés 22 à 54), renommé ultérieurement « Pulicario odorae-Arbutetum unedonis Reymann, Panaïotis & Bioret 2017 » et actualisé ici en Myrto communis-Arbutetum unedonis (Allier & Lacoste 1980) nom. nov. hoc loco, typus nominis hoc loco : relevé 33 du tableau III in Allier & Lacoste (1980, Ecol. Medit. 5 : 70) ; reconnaissable aussi chez Paradis & Pozzo di Borgo (2005, tableaux 31 et 32).
Ea12. Myrto communis–Cytisetum villosi ass. nov. hoc loco (Toubal & Toubal, 1996a, tableau II), à Ampelodesmos mauritanicus, Cytisus villosus, C. spinosus, Erica arborea, Arbutus unedo, Myrtus communis, Quercus faginea subsp. f. j, Q. suber j… ; typus nominis hoc loco : composante arbustive et arbrisellée du relevé 3 du tableau II in Toubal & Toubal (1996a, Colloq. Phytosoc. XXIV : 589).
Ea13. Groupement à Cytisus grandiflorus-Ulex europaeus subsp. latebracteatus (Braun-Blanquet et al., 1956, tableau V).
Ea14. « Genisto corsicae-Ericetum multiflorae Lejour & Delbosc in Reymann, Panaïotis & Bioret 2017 » (Reymann et al., 2017 : 150).
Ea15. Junipero oxycedri-Euphorbietum dendroidis (Géhu & Biondi 1994) Paradis & Piazza 2001 (Paradis & Piazza, 2001, tableau 4).
Ea16. Pistacio lentisci-Calicotometum villosae Biondi, Filigheddu & Farris 2001 (Biondi et al., 2001, tableau 59) ; cité de Corse par Paradis et al. (2014, tableau 19).
Ea17. Erico arboreae-Juniperetum turbinatae De Marco, Dinelli & Caneva 1985 corr. Biondi & Bagella 2005 ; a – race italienne à Rubia peregrina subsp. longifolia (toutefois présent aussi en Corse et donc peut-être non significatif), Prasium majus, Rhamnus alaternus subsp. a. (Gianguzzi et al., 2012, tableau 8) ; b – race corse à Erica scoparia subsp. s. et Olea europaea subsp. e. j (Paradis & Pozzo di Borgo, 2005, tableaux 28, 29, sub Myrto communis-Juniperetum turbinatae, 30 et 36, sub Myrto communis-Ericetum scopariae).
* Groupe de syntaxons à Quercus pubescens j, surtout nord-méditerranéens
Ea18. Groupement à Ulex parviflorus subsp. p.-Euphorbia characias subsp. c. (Feliu Xifra, 2003, tableau p. 127), strictement intermédiaire entre les deux groupes de syntaxons.
Ea19. Rubo ulmifolii-Ericetum arboreae Aurière & Reymann in Reymann, Panaïotis & Bioret 2017 (Reymann et al., 2017 : 155).
Ea20. Cisto salviifolii-Cytisetum catalaunici (A. Bolòs & O. Bolòs 1950) O. Bolòs 1956 (de Foucault, 2019a, tableau 4 : synthèse des colonnes A, B, C).
Ea21. Cytiso scoparii-Ericetum arboreae B. Foucault 2015 (de Foucault, 2015b, tableau 4) ; photo 17 ; représente la fin de l’alliance vers le nord de son aire, accueillant alors Cytisus scoparius subsp. s. en provenance des Cytisetea scopario-striati, classe relayant l’Ericion arboreae.
Ea22. Ilici aquifolii-Ericetum scopariae B. Foucault 2021 (de Foucault, 2021b, tableau 1) ; photo 18.
Ea23. Groupement à Buxus sempervirens-Erica scoparia subsp. s. (Choisnet, 2019, tableau 13).
Ea24. « Lavandulo stoechadis-Ericetum scopariae Braun-Blanq. ex Braun-Blanq., Molin. & He. Wagner 1940 » (Braun-Blanquet et al., 1940 : 20) renommé Phillyreo angustifoliae-Ericetum scopariae par Julve (1993), non validable, les données sources étant synthétiques ; ce nom est d’ailleurs illégitime [non Phillyreo angustifoliae-Ericetum scopariae Braun-Blanq., P. Silva & Rozeira 1964 (Agron. Lusit. 23 (4) : 256) prov., cf supra syntaxon Ql4]).
Ea25. Calicotomo spinosae-Ericetum arboreae (Braun-Blanq. in Braun-Blanq., Molin. & He. Wagner 1940) B. Foucault & Julve 1991 (Braun-Blanquet et al., 1940 : 23, sub « association à Cistus crispus et Calicotome spinosa »), non validable, les données sources étant synthétiques.
La monographie de Gauberville et al. (2019) sur les boisements de Pinus nigra subsp. laricio répartit ceux-ci en quatre syntaxons selon notamment un gradient altitudinal. Ceux de plus haute altitude (leurs tableaux 3 et 4 ; moyennes 1 330 m et 1 410 m d’altitude respectivement ; étage montagnard) ont une association arborescente supra-méditerranéenne correspondant au Betulo pendulae-Pinetum laricionis (Gamisans 1975) B. Foucault 2020 (de Foucault, 2020a) et une association intraforestière arbustive se réduisant aux formes juvéniles de celui-ci, donc peu caractérisée. En revanche, ceux de plus basse altitude (tableaux 1 et 2 ; moyennes 700 m et 1 120 m d’altitude respectivement ; étage supraméditerranéen) ont une association arborescente méditerranéo-montagnarde à Pinus nigra subsp. laricio, P. pinaster, Quercus ilex, plus rarement Castanea sativa : Pinetum pinastri-laricionis (Gauberville, Panaïotis, Bioret & Capelo 2019) ass. nov. hoc loco, typus nominis hoc loco : composante A1 du relevé 2 du tableau 1 in Gauberville et al. (2019, Ecol. Medit. 45 (1), on line), à ranger dans les Quercetalia rotundifolio-ilicis et une alliance originale de boisements supraméditerranéens tyrrhéniens, le Querco ilicis-Pinion laricionis all. nov. hoc loco, typifié par le Pinetum pinastri-laricionis (Gauberville, Panaïotis, Bioret & Capelo 2019) nov. (typus nominis hoc loco). L’association intraforestière arbustive nous rapproche de l’alliance étudiée ici, car elle est surtout caractérisée par Erica arborea, Arbutus unedo (surtout tableau 1 de ces auteurs), des formes juvéniles des esences du Pinetum pinastri-laricionis, plus rarement Cytisus villosus, Ilex aquifolium ; il s’agit d’une forme semisciaphile d’un fourré de l’Ericion arboreae sans doute mieux caractérisé dans les clairières et lisières plus éclairées. Quand à la végétation intraforestière herbacée, elle relève clairement des Rubio peregrinae-Asplenietea onopteridis B. Foucault 2020 et du Cyclamino repandi-Asplenion onopteridis B. Foucault 2020 (de Foucault, 2020a).

Alliance Gml. Genistion monspessulano-linifoliae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 nom. mut. hoc loco (Itinera Geobot. 15 : 353, sub Telinion monspessulano-linifoliae) nom. conserv. ; tableau 1 : colonne Gml ; tableau 26
Typus allianciae : Cytiso baetici-Genistetum monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 désigné in Rivas Mart. et al. 2002 (Itinera Geobot. 15 : 92).
Fourrés acidiphiles thermo-acidiphiles méditerranéens partageant avec l’alliance précédente Erica scoparia subsp. s., Cistus monspeliensis, Quercus coccifera, Q. suber j, Q. ilex j, mais plus riches en taxons dont Genista monspessulana (= Teline monspessulana), G. linifolia (= T. linifolia), Adenocarpus telonensis, Ulex parviflorus subsp. funkii, Cytisus tribracteolatus, C. arboreus subsp. a.
Avec Adenocarpus telonensis, Erica arborea, E. scoparia subsp. s., Ulex parviflorus subsp. funkii, le Pino acutisquameae–Quercetum cocciferae Cabezudo, Nieto & A.V. Pérez 1989 corr. Ladero & Asensi 1999 semble pouvoir se placer dans cette alliance ; toutefois c’est l’holotype du Pino acutisquameae-Juniperion phoeniceae A.V. Pérez & Cabezudo in A.V. Pérez, P. Navas, D. Navas, Y. Gil & Cabezudo 1998 (Acta Bot. Malac. 23 : 153) corr. Rivas Mart. et al. 2002 (Itinera Geobot. 15 : 359) prioritaire sur le Genistion monspessulano-linifoliae ; pour ne pas trop perturber le synsystème, on propose ici la conservation de ce dernier.
Deux groupes d’associations, le Gml4 (Cytiso baetici-Genistetum monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 nom. mut. hoc loco ; Sivim, consulté le 27/09/2020, sub Cytiso baetici-Telinetum monspessulanae) sans rattachement clairement défini, étant réduit à un seul relevé.
* à Juniperus oxycedrus subsp. o.
Gml1. Cytiso baetici-Arbutetum unedonis Nieto-Caldera, Pérez-Latorre & Cabezudo 1990 (Nieto-Caldera et al., 1990, tableau 4).
Gml2. Pino acutisquameae-Quercetum cocciferae Cabezudo, Nieto & A.V. Pérez 1989 corr. Ladero & Asensi 1999 (Cabezudo et al., 1989, tableau 1, sub Pino pinastri-Quercetum cocciferae).
Gml3. Cytiso tribracteolati-Juniperetum oxycedri A.V. Pérez, Galán & Cabezudo in A.V. Pérez, Galán, P. Navas, D. Navas, Y. Gil & Cabezudo 1999 (Sivim, consulté le 8/09/2020).


* à Cytisus spinosus, Asparagus acutifolius, Lonicera implexa, Euphorbia characias subsp. c., Cistus monspeliensis, Quercus ilex j
Gml5. Cytiso villosi-Ericetum arboreae Zéller 1959 (Sivim, consulté le 27/09/2020).
Gml6. Cytiso spinosi-Genistetum linifoliae B. Foucault 2021 (de Foucault, 2021a, tableau 1) ; photo 19.
Gml7. Querco suberis-Genistetum monspessulanae R.J. Loisel 1971 (de Foucault, 2021a, tableau 1 : colonne Qs-Gm).
Gml8. Querco suberis-Genistetum linifoliae R.J. Loisel 1971 (de Foucault, 2021a, tableau 1 : colonne Qs-Gl).
Gml9. Genisto desoleanae-Ericetum arboreae Farris, Zecchi & Filigheddu 2007 (Farris et al., 2007, tableau 13).
Gml10. Genisto monspessulanae-Cytisetum villosi Farris, Zecchi & Filigheddu 2007 (Farris et al., 2007, tableau 14).
Dans les Corbières orientales audoises, au contact d’une lande à Ulex parviflorus subsp. p. j et Lavandula stoechas subsp. s., il existe aussi une végétation encore non décrite relevant de cette alliance dont le tableau 27 rapporte deux relevés en attendant des inventaires plus conséquents (relevé 1 : Villeneuve-les-Corbières, col d’Extrême, côté ouest de la D 611, le 16/03/2016 ; relevé 2 : Thézan-des-Corbières, bois de Donos, côté est de la D 611, le 07/07/2016). Genista monspessulana manque dans ces deux relevés mais est bien présent dans cette région (taxon patrimonial de l’Aude, au même titre qu’Adenocarpus telonensis ; Plassart et al., 2016), alors que G. linifolia en est absent.
Alliance Ft. Flueggeion tinctoriae Rivas Goday 1964 (Vegetación y flórula de la cuenca extremeña del Guadiana : 555, sub Securinegion buxifoliae) ; tableau 1 : colonne Ft ; tableau 28
Typus allianciae : Pyro bourgaeanae-Flueggeetum tinctoriae (Rivas Goday 1964) Rivas Mart. & Rivas Goday 1975 (Colloques internationaux du CNRS 235 : 441).
Alliance très originale de fourrés ibériques mésohygrophiles à Flueggea tinctoria (= Securinega buxifolia), Salix salviifolia, Pyrus bourgaeana, Nerium oleander, Clematis viticella subsp. campaniflora, enrichis en taxons d’origine eurosibérienne (Bryonia dioica, Sambucus nigra, Crataegus monogyna, Rosa micrantha, R. canina), ainsi que de formes juvéniles d’arbres issus de forêts hygrophiles (Alnus glutinosa, Fraxinus angustifolia subsp. a.) et de Quercus rotundifolia, mais plus (surtout le type) ou moins pourvus en taxons caractérisant les Pistacio-Rhamnetea alaterni.
Ft1. Pyro bourgaeanae-Flueggeetum tinctoriae (Rivas Goday 1964) Rivas Mart. & Rivas Goday 1975 (Rivas Goday, 1964, tableau 83, sub Securinegeto-Piretum marianicum).
Ft2. « Oenantho crocatae-Nerietum oleandri Rivas Mart. & Fuente in Fuente, Rufo Nieto, Rodríguez González & Amils Pibernat 2007 » (de la Fuente García et al., 2007, tableau 1), renommé ici Flueggeo tinctoriae-Nerietum oleandri (Rivas Mart. & Fuente in Fuente, Rufo Nieto, Rodríguez González & Amils Pibernat 2007) ass. nov. hoc loco, typus nominis hoc loco : composante nanophanérophytique du relevé 26 du tableau 1 in de la Fuente et al. (2007, Lazaroa 28 : 8).
Alliance GEa. Genisto numidicae-Ericion arboreae all. nov. hoc loco ; tableau 1 : colonne GEa ; tableau 29
Typus nominis hoc loco : Genistetum numidicae O. Toubal 1998 (Itinera Geobot. 11 : 213-226).
Fourrés acidiphiles du Maghreb (Maroc oriental à Tunisie) à Quercus suber j, Rosa sempervirens, Genista numidica, Cytisus spinosus, Ampelodesmos mauritanicus.
GEa1. Groupement à Chamaerops humilis (Wojterski, 1985, tableau 28 : relevés 1 à 6).
GEa2. Groupement à Cistus monspeliensis (Ibid., tableau 28 : relevés 7 à 13).
GEa3. Genistetum numidicae O. Toubal 1998 (Toubal, 1998, tableaux 4 et 5), distinct de GEa8, typifié par le relevé in Toubal (1998, Itinera Geobot. 11 : 218).
GEa4. Groupement à Genista ulicina-G. numidica (Khelifi & Sadki, 1995, tableau V : colonne VII).
GEa5. Groupement à Quercus canariensis j-Cytisus villosus (Braun-Blanquet, 1954 : 184 haut de page et bas de page).
GEa6. Erico arboreae-Quercetum cocciferae Quézel, Barbero, Benabid & Rivas Mart. 1992 (Quézel et al., 1992, tableau 1).
GEa7. Erico arboreae-Myrtetum communis Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 ; a – race rifaine à Aristolochia baetica et Quercus coccifera (Quézel et al., 1988, tableau 1) ; b – race kabyle à Ampelodesmos mauritanicus, Cytisus spinosus et Phillyrea media (Meddour, 2010, tableau 85).
GEa8. Groupement à Genista numidica (Wojterski, 1985, tableau 28), distinct de GEa3.
Alliance TEa. Tetraclinido articulatae-Ericion arboreae all. nov. hoc loco ; tableau 1 : colonne TEa ; tableau 30
Typus nominis hoc loco : Calicotomo intermediae-Tetraclinidetum articulatae Barbero, Quézel & Rivas Mart. 1981 (Phytocoenologia 9 (3) : 350).
Fourrés acidiphiles du Maghreb planitiaire à collinéen à Ampelodesmos mauritanicus, Rubia peregrina subsp. longifolia, Calicotome infesta subsp. intermedia, Erica multiflora, Genista tricuspidata, Cistus ladanifer subsp. l., Tetraclinis articulata j, Olea europaea subsp. e. j.
TEa1. Groupement à Quercus faginea subsp. f. j-Juniperus phoenicea subsp. ph. (Vanden Berghen, 1990, tableau 2).
TEa2. Dioscoreo communis-Quercetum cocciferae Hadjadj & R.J. Loisel 1999 (Hadjadj & Loisel, 1999, tableau 8).
TEa3. Genisto tricuspidatae-Pistacietum lentisci (Hadjadj & R.J. Loisel 1999) nom. nov. hoc loco (Ibid., tableau 7) ; avec Tetraclinis articulata j, Calicotome infesta subsp. intermedia, Genista tricuspidata, Ampelodesmos mauritanicus, Phillyrea angustifolia et l’absence de Aristolochia baetica, Quercus suber j et Clematis cirrhosa, l’Erico arboreae-Myrtetum communis Hadjadj & R.J. Loisel 1999 ne peut être rattaché à l’association de Quézel et al. (1988 ; cf.supra syntaxon A5 et infra syntaxon TEa7) ; il est donc renommé ici (art. 39), typus nominis hoc loco : le même que celui des auteurs pour leur pseudonyme, soit le relevé 86 du tableau 7 in Hadjadj & Loisel (1999, Doc. Phytosoc., NS, XIX : 253).
TEa4. Tetraclinido articulatae-Phillyreetum latifoliae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 9).
TEa5. Calicomo intermediae-Tetraclinidetum articulatae Barbero, Quézel & Rivas Mart. 1981 ericetosum arboreae Hadjadj & R.J. Loisel 1999 (Ibid., tableau 10), élevé ici au rang d’association : Calicotomo intermediae-Ericetum arboreae (Hadjadj & R.J. Loisel 1999) stat. nov. hoc loco, typus nominis hoc loco : le même que celui des auteurs pour leur sous-association, soit le relevé 673 du tableau 10 in Hadjadj & Loisel (1999, Doc. Phytosoc., NS, XIX : 258).
TEa6. Cytiso arborei-Quercetum cocciferae Quézel, Barbero, Benabid & Rivas Mart. 1992 (Quézel et al., 1992, tableau 2).
TEa7. Erico arboreae-Myrtetum communis Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 var. à Lavandula dentata (Quézel et al., 1988, tableau 2) ; la comparaison avec la variante type de cette association (Quézel et al., 1988, tableau 1) placée dans l’alliance précédente montre la présence ici d’Erica multiflora, E. scoparia subsp. s., Tetraclinis articulata j, Ampelodesmos mauritanicus, Chamaerops humilis, Olea europaea subsp. e. et l’absence significative de Daphne gnidium, Aristolochia baetica, Quercus suber j, Rubia peregrina subsp. p. ; il faudra à l’avenir sans doute en faire un syntaxon original.
TEa8. Phillyreo latifoliae-Quercetum cocciferae Quézel, Barbero, Benabid, R.J. Loisel & Rivas Mart. 1988 (Quézel et al., 1988, tableau 3).
TEa9. « Simethido-Ericetum arboreae Nègre 1964 » (Nègre, 1964, tableau 5) ; à renommer, mais impossible à typifier sur les données publiées qui sont synthétiques.
TEa10. Erico arboreae-Quercetum cocciferae sensu Medjadhi & Letreuch-Belarouci 2017 (Doc. Phytosoc., 3e série, 6 : 138) non Quézel et al. 1992 (Medjadhi & Letreuch-Belarouci, 2017, tableau 4) ; se distingue du syntaxon de Quézel et al. (1992 ; cf. supra syntaxon A5) par la présence de Tetraclinis articulata j, Ampelodesmos mauritanicus, Cistus ladanifer subsp. l. et l’absence de Cytisus baeticus, Phillyrea latifolia, Ph. media, Olea europaea subsp. e. j, Cistus albidus.
Alliance Csv. Cytision spinoso-villosi B. Foucault & Meddour all. nov. hoc loco ; tableau 1 : colonne Csv ; tableau 31
Typus nominis hoc loco : Lonicero implexae-Cytisetum villosi Meddour & B. Foucault in B. Foucault 2020 (Evaxiana 7 : 58).
Fourrés acidiphiles de la Kabylie djurdjuréenne, décrits surtout par Meddour (2010), à Cytisus villosus, C. spinosus, Erica arborea, Rubus ulmifolius, l’altitude (jusque 1 400 m) laissant parfois pénétrer des taxons de climat plus tempéré, surtout Crataegus monogyna.
Csv1. Groupement à Juniperus oxycedrus subsp. o.-Cytisus villosus (Meddour, 2010, tableaux 74 et 82).
Csv2. Groupement à Cytisus spinosus-C. villosus (Ibid., tableau 80).
Csv3. Asparago acutifolii-Cytisetum villosi Meddour & B. Foucault in B. Foucault 2020 (Ibid., tableau 83).
Csv4. Lonicero implexae-Cytisetum villosi Meddour & B. Foucault in B. Foucault 2020 (Ibid., tableau 84).
Le tableau 3 de Quézel et al. (1987) décrit un groupement du Haut Atlas oriental (13 relevés, entre 1 180 et 1 500 m d’altitude ; tableau 1 : colonne CCc), à Cistus creticus, Tetraclinis articulata j, Jasminum fruticans, Quercus rotundifolia j, Cytisus arboreus subsp. catalaunicus, Adenocarpus anagyrifolius, Arbutus unedo (mais pas Erica arborea) et divers taxons caractéristiques de la sous-classe et de la classe ; cette association – Cytiso catalaunici-Cistetum cretici ass. nov. hoc loco, typus nominis hoc loco : relevé 4 du tableau 3 in Quézel et al. (1987, Ecol. Medit. 13 (1-2) : 116) – paraît correspondre à une limite altitudinale supérieure des Cytiso villosi-Genistetalia monspessulanae.
Alliance HEa. Hipocrepido emeroidis-Ericion arboreae all. nov. hoc loco ; tableau 1 : colonne HEa ; tableau 32
Typus nominis hoc loco : Phillyreo mediae-Hippocrepidetum emeroidis Barbero & Quézel ex B. Foucault.
Fourrés acidiphiles sud-est-européens à tendance centre-méditerranéenne à Hippocrepis emerus subsp. emeroides, Cistus creticus, Erica arborea, Arbutus unedo, Rubia peregrina subsp. p., Cercis siliquastrum j, Fraxinus ornus subsp. o. j.
HEa1. Groupement à Hippocrepis emerus subsp. emeroides (Barbero & Quézel, 1976, tableau 7, sub « chênaies acidophiles à Arbutus unedo »), validé ici en Phillyreo mediae-Hippocrepidetum emeroidis Barbero & Quézel ex ass. nov. hoc loco, à Hippocrepis emerus subsp. emeroides, Phillyrea media, Clematis flammula, Cotinus coggygria, Quercus coccifera, Q. pubescens j, Pistacia terebinthus, Dioscorea communis, Fraxinus ornus subsp. o. j, Cercis siliquastrum j, typus nominis hoc loco : relevé 3 du tableau 7 in Barbero & Quézel (1976, Ecol. Medit. 2 : 21).
HEa2. « Andrachno-Quercetum ilicis Oberd. 1948 » (Horvat et al., 1974, tableau 11 : colonnes 1 à 6).
HEa3. Cisto-Ericetum arboreae Horvatić 1958 (Ibid., tableau 20 : colonnes 5 à 8).
Le « Genisto-Ericetum manipuliflorae Horvatić 1958 » (Horvat et al., 1974, tableau 20 : colonnes 11 et 12) est relié aux Pistacio-Rhamnetea alaterni par Phillyrea latifolia, Pistacia terebinthus, Smilax aspera, Viburnum tinus, mais n’a guère de lien avec les Cytiso-Genistetalia monspessulanae.
Ordre 3. Rubio tenuifoliae-Pistacietalia terebinthi ord. nov. hoc loco ; tableau 1 : colonnes RJt à ABc
Typus nominis hoc loco : Ceratonio siliquae-Pistacion lentisci Zohary & Orshan 1959 (Vegetatio 18 : 285).
Fourrés centre- à est-méditerranéens caractérisés par un fort déficit de taxons caractérisant ou différenciant la sous-classe précédente ; plus positivement, ils partagent Rubia tenuifolia, Prasium majus, Cytisus laniger, Arbutus andrachne (photo 20), Fraxinus ornus subsp. o. j. Cet ordre entre alors dans une sous-classe distincte des Eu-Pistacio lentisci-Rhamnenea alaterni, les Rubio tenuifoliae-Pistacienea terebinthi subclassis nov. hoc loco, typus nominis hoc loco : Rubio tenuifoliae-Pistacietalia terebinthi nov.
Alliance RJt. Rhamno graecae-Juniperion turbinatae M.J. Costa, Géhu, Peris, Biondi & Arnold 1984 (Doc. Phytosoc., NS, VIII : 373) ; tableau 1 : colonne RJt
[corresp. syntax. : Asparago orientalis-Juniperenion macrocarpae Díez Garretas & Asensi 2013 (Pl. Biosyst. 148 (3) : 51)]
Typus allianciae : Ephedro foemineae-Juniperetum turbinatae Barbero & Quézel 1979 (Doc. Phytosoc., NS, IV : 18) désigné par les auteurs de l’alliance (1984, Doc. Phytosoc., NS, VIII : 368).
Fourrés arrière-dunaires du littoral méditerranéen central à oriental ; alliance rappelant le Juniperion turbinatae ouest-méditerranéen par Juniperus phoenicea subsp. turbinata et J. oxycedrus subsp. macrocarpa mais s’en distinguant par un lot significatif de taxons optimaux en Méditerranée orientale : Asparagus aphyllus subsp. orientalis, Rubia tenuifolia, Rhamnus lycioides subsp. graeca (photo 21), Lycium schweinfurthii, Arbutus andrachne, Daphne gnidioides, ainsi que Cytisus laniger et Prasium majus.
Synthèse des colonnes 38 à 44 du tableau I in Díez Garretas & Asensi (2013) dans la colonne RJt du tableau 1.

18Alliance CsPl. Ceratonio siliquae-Pistacion lentisci Zohary & Orshan 1959 (Vegetatio 18 : 285) ; tableau 1 : colonne CsPl ; tableau 33
[syn. : Ceratonio siliquae-Rhamnion oleoidis Barbero & Quézel 1983 (Colloq. Phytosoc. VIII : 13) nom. inval. et superfl. ; Ceratonio siliquae-Rhamnion oleoidis Barbero & Quézel ex Quézel, Barbero & Akman 1992 (Ecol. Medit. 18 : 83) nom. illeg. et superfl.]
Lectotypus nominis hoc loco : Ceratonio siliquae-Pistacietum lentisci Eig 1946 (Palest. J. Bot., Jerusalem ser. III (4) : 192).


Fourrés est-méditerranéens à Rubia tenuifolia, Rhamnus lycioides subsp. graeca, Cytisus laniger, Pistacia terebinthus, Prasium majus et différenciés par des formes juvéniles de taxons arborescents tels que Ceratonia siliqua, Quercus coccifera (pouvant être arborescent en Méditerranée orientale, alors qu’il reste plutôt arbustif en Méditerranée occidentale), Pinus brutia.
La définition du Ceratonio siliquae-Rhamnion oleoidis par Barbero & Quézel 1983 puis sa validation par Quézel et al. (1992a) d’une part ignorent le nom d’alliance donné bien antérieurement par Zohary & Orshan en 1959, d’autre part sont erronées car le type désigné, le « Ceratonio siliquae-Pistacietum palaestinae Zohary & Orshan 1959 », n’existe pas, les deux auteurs cités n’ayant évoqué que trois formes du Ceratonio siliquae-Pistacietum lentisci, dont la plus typique ayant Eig (1946) comme autorité.
La photo 22 illustre un aspect de la végétation de cette alliance pour le Liban.
CsPl1. Groupement à Smilax aspera-Rubia tenuifolia (Stefan et al., 2019, tableau 5 : quatre premiers relevés).
CsPl2. Calicotomo villosae-Rhamnetum punctatae Stefan, Barhoun, Maatouk & B. Foucault 2019 (Ibid., tableau 5 : sept derniers relevés).
CsPl3. Pistacio terebinthi-Cytisetum lanigeri B. Foucault, El Beyrouthy & Arnold 2013 (de Foucault et al., 2013b, tableau 2).
CsPl4. Pistacio terebinthi-Rhamnetum graecae Géhu, Uslu & M.J. Costa 1992 nom. mut. hoc loco (Géhu et al., 1992d, tableau 25, sub Pistacio palaestinae-Rhamnetum graecae).
CsPl5. Rubio tenuifoliae-Pistacietum lentisci Géhu, M.J. Costa & Uslu 1990 (Géhu et al., 1990, tableau 23 ; Géhu et al., 1992d, tableau 26).
CsPl6. Dorystaechadi hastatae-Oleetum europaeae Kurt, Ketenoğlu, Akman, Özdeniz, Şekerciler, Bölükbaşi & Özbey 2015 (Kurt et al., 2015, tableau 1 : relevés 1 à 13).
CsPl7. Rubio tenuifoliae-Pistacietum lentisci sensu Géhu, Apostolidès, Géhu-Franck & Arnold 1992 (Géhu et al., 1992a, tableau 30) ; avec Asparagus aphyllus, Juniperus phoenicea subsp. turbinata, Anagyris foetida dans une variation, l’absence de Quercus coccifera, Cytisus laniger, Olea europaea subsp. e. j, Daphne gnidioides, Pistacia terebinthus…, ce syntaxon doit être manifestement distingué du CsPl5 [Rubio tenuifoliae-Pistacietum lentisci Géhu, M.J. Costa & Uslu 1990 (Doc. Phytosoc., NS, XII : 214, ‘Rubio tenuis…’ art. 41a)] : Asparago aphylli-Pistacietum lentisci (Géhu, Apostolidès, Géhu-Franck & Arnold 1992) nom. nov. hoc loco, à Pistacia lentiscus, Asparagus aphyllus, Anagyris foetida, Juniperus phoenicea subsp. turbinata, Prasium majus, Rhamnus lycioides subsp. graeca, Rubia tenuifolia, typus nominis hoc loco : relevé 6 du tableau 30 in Géhu et al. (1992a, Colloq. Phytosoc. XIX : 580).
CsPl8. Cisto cretici-Arbutetum andrachnes Kaya, Cansaran & Yıldırım 2010 (Kaya et al., 2010, tableau VII).
CsPl9. Paliuretum adriaticum Horvatić 1963 (Horvat et al., 1974, tableau 20 : colonnes 9 et 10), à renommer (art. 34).
CsPl10. « Querco boissieri-Carpinetum orientalis Kaya, Cansaran & Yıldırım 2010 » (Kaya et al., 2010, tableau V).
CsPl11. Paliuro spinae-christi-Pistacietum terebinthi Hamzaoğlu, Duran & Menemen 2002 (Hamzaoğlu et al., 2002, tableau 3).
CsPl12. « Phillyreo mediae-Pinetum brutiae Kaya, Cansaran & Yıldırım 2010 » (Kaya et al., 2010, tableau VI).
CsPl13. Ceratonio siliquae-Pistacietum lentisci Eig 1946 (Zohary & Orshan, 1959 : 288).
CsPl14. Ceratonio siliquae-Pistacietum lentisci orientale Zohary & Orshan 1959 (Zohary & Orshan, 1959 : 294), à renommer sur la base de relevés précis nécessaires à sa validation complète.
CsPl15. Rubio tenuifoliae-Euphorbietum dendroidis Géhu, M.J. Costa & Uslu 1988 (Géhu et al., 1988, tableau 1).
CsPl16. Phlomido fruticosae-Euphorbietum dendroidis Theocharopoulos, Pantera, Fotiadis & Papadopoulos 2020 (Theocharopoulos et al., 2020, tableau 1 : colonne 11).
CsPl17. Ceratonio siliquae-Pistacietum lentisci arenarium Eig 1939 (Zohary & Orshan, 1959 : 293), à renommer sur la base de relevés précis nécessaires à sa validation complète.
Alliance RBs. Pistacio terebinthi-Buxion sempervirentis all. nov. hoc loco ; tableau 1 : colonne PBs ; tableau 34
Typus nominis hoc loco : Pistacio terebinthi-Buxetum sempervirentis B. Foucault, Gerasimidis, Fotiadis & Panajiotidis nov.
Le tableau 34 synthétise les groupements à Buxus sempervirens-Phillyrea media (tableau 1 des auteurs ; ici codé comme PBs1), plus xérophile, et à Platanus orientalis-Buxus sempervirens (tableau 2 des auteurs ; PBs2), plus hygrophile, de Gerasimidis et al. (2006) qui les placent respectivement au sein des Rhamno-Prunetea spinosae et des « Populetea albae ». Ces auteurs les ont décrits du mont Olympe, en Grèce, entre 120 et 170 m d’altitude. De façon plus moderne, le premier correspond bien à un fourré, le second plutôt à un boisement de Platanus orientalis avec un sous-bois arbustif à Buxus sempervirens. Quoi qu’il en soit, ils relèvent bien des Pistacio-Rhamnetea alaterni, avec dans le second cas une introgression de taxons caractérisant les Rhamno-Prunetea spinosae comme c’est souvent le cas en position alluviale. Avec onze relevés, le premier peut être validé sous le nom de Pistacio terebinthi-Buxetum sempervirentis B. Foucault, Gerasimidis, Fotiadis & Panajiotidis ass. nov. hoc loco, à Osyris alba, Buxus sempervirens, Juniperus oxycedrus subsp. o., Pistacia terebinthus, Asparagus acutifolius, Phillyrea latifolia, Quercus coccifera, Crataegus monogyna, Carpinus orientalis j, Fraxinus ornus subsp. o. j, typus nominis hoc loco : relevé 6 du tableau 1 in Gerasimidis et al. (2006, Proceedings of IV Balkan Botanical Congress : 303), dynamiquement lié à des boisements à Carpinus orientalis et Fraxinus ornus (Fraxino orni-Carpinetum orientalis Trinajstić & B. Foucault in B. Foucault 2020 ?). Pour information, le second groupement est associé à un boisement hygrophile à Platanus orientalis, Juglans regia, Vitis vinifera subsp. sylvestris, Ficus carica relevant du Liquidambaro orientalis-Platanion orientalis (de Foucault, 2020a). Le Pistacio terebinthi-Buxion sempervirentis se caractérise par une combinaison floristique à Pistacia terebinthus, Buxus sempervirens, Asparagus acutifolius, Phillyrea latifolia, Quercus coccifera, Crataegus monogyna, Fraxinus ornus j.
Alliance ERr. Ephedro aphyllae-Retamion raetam nov. ; tableau 1 : colonne ERr ; tableau 35
Typus nominis hoc loco : cf. infra.
Le tableau 35 synthétise quatre syntaxons de la façade méditerranéenne d’Israël décrits de façon invalide par Danin et Orshan (1999) ; ce sont
ERr1. « Ephedro aphyllae-Helianthemetum stipulati Danin & Solomehsch in Danin & Orshan 1999 » (Danin & Orshan, 1999, tableau 45 : syntaxon DMH01) ; sans doute une forme du suivant.
ERr2. « Retamo raetam-Echiochiletum fruticosi Danin & Solomehsch in Danin & Orshan 1999 », « Plantagini sarcophyllae-Helianthemetum stipulati Danin & Solomehsch in Danin & Orshan 1999 » et « Trifolio palaestini-Helianthemetum stipulati Danin & Solomehsch in Danin & Orshan 1999 » (Danin & Orshan, 1999, tableau 45 : syntaxons DMH02 et DMH03, tableau 46 : DMH04), globalement réunis et validés ici en Ephedro aphyllae-Retametum raetam Danin & Solomehsch ex ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du syntaxon DMH02 du tableau 45 in Danin & Orshan (1999, Vegetation of Israel, I : 249).
ERr3. « Pistacio lentisci-Helianthemetum stipulati Danin & Solomehsch in Danin & Orshan 1999 » (Danin & Orshan, 1999, tableau 46 : syntaxon DMH05), validé ici en Rubio tenuifoliae-Retametum raetam Danin & Solomehsch ex ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 2 du syntaxon DMH05 du tableau 46 in Danin & Orshan (1999, Vegetation of Israel, I : 253).
ERr4. Pistacio lentisci-Calicotometum villosae Danin & Solomehsch in Danin & Orshan 1999 nom. inval. (Danin & Orshan, 1999, tableau 46 : syntaxon DMH06), validé ici en Pistacio lentisci-Calicotometum villosae Danin & Solomehsch ex ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 3 du syntaxon DMH06 du tableau 46 in Danin & Orshan (1999, Vegetation of Israel, I : 253) ; forme plus riche en espèces du Calicotometum villosae Eig 1946 (Eig, 1946 : 194).
Ces quatre syntaxons peuvent être réunis au sein d’une alliance est-méditerranéenne originale, l’Ephedro aphyllae–Retamion raetam all. nov. hoc loco, caractérisée par Retama raetam, Rubia tenuifolia, Rhamnus lycioides subsp. graeca, Ephedra foeminea, E. aphylla, Rhamnus alaternus subsp. a. (qui paraît peu représenté par ailleurs dans les Rubio tenuifoliae-Pistacietalia terebinthi ; tableau 1), Asparagus aphyllus, A. horridus, typus nominis hoc loco : Ephedro aphyllae-Retametum raetam Danin & Solomehsch ex B. Foucault ; cette alliance correspond à une partie du Retamo raetam-Phlomidion brachyodontis Eig 1938 (Eig, 1946 : 205), surtout chaméphytique.
Aux quatre syntaxons précédents, on pourrait encore ajouter un fourré associé à un boisement de Pistacia atlantica dans des steppes plus continentales (Danin & Orshan, 1999, tableau 32, syntaxon VDN02 : relevés 1, 2, 4, 6 à 9, 13, soit huit relevés, sub « Pistacio atlanticae-Chiliadenetum iphionoidis ») : Ephedra aphylla V, Rhamnus disperma IV, Asparagus horridus II, Colutea istria II, Prasium majus I, Retama raetam I, et sans doute des Pistacia atlantica j, Ephedro aphyllae-Rhamnetum dispermae ass. nov. hoc loco, typus nominis hoc loco : composante arbustive du relevé 2 du tableau 32, syntaxon VDN02 in Danin & Orshan (1999, Vegetation of Israel, I : 200). Le Periplocetum aphyllae Danin & Solomehsch in Danin & Orshan 1999 se réduit à deux arbustes, Periploca aphylla V (sur sept relevés) et Retama raetam IV (Danin & Orshan, 1999, tableau 31).
Ajoutons à cela qu’on trouve dans cette même publication la reconnaissance du Retamo raetam-Calligonetum comosi Danin & Solomeshch in Danin & Orshan 1999 (Vegetation of Israel, I : 303 et tableau 57), reconnu ultérieurement et indépendamment (de Foucault, 2020a : 65) ; comme le syntaxon de 1999 est invalide, le nom complet de cette association reste Retamo raetam-Calligonetum comosi (Quézel 1965) Claisse & B. Foucault ex B. Foucault 2020, avec toutefois une race est-méditerranéenne différenciée par Ephedra aphylla.
Alliance RBs. Asparago acutifolii-Berberidion creticae all. nov. hoc loco (tableau 1 : colonne ABc)
Typus nominis hoc loco : Aceri sempervirentis-Berberidetum creticae Quézel & Barbero 1980 (Ecol. Medit. 5 : 202).
Des montagnes de Crète, Quézel et Barbero (1980, tableau VIII) ont décrit l’Aceri sempervirentis-Berberidetum creticae, à Berberis cretica, Crataegus monogyna (incl. C. azarella), Rosa pulverulenta, Asparagus acutifolius, Pyrus spinosa, ainsi que Acer sempervirens j, Quercus coccifera j, et dans des sous-associations Zelkova abelicea et Cupressus sempervirens j d’un côté, Aristolochia sempervirens d’un autre. Cette association a été revue plus tard (de Foucault, 1993a, tableau 8) avec Amelanchier ovalis subsp. cretica. Comme on le voit, les liens synfloristiques avec les Pistacio-Rhamnetea alaterni et les Rubio tenuifoliae-Pistacietalia terebinthi sont ténus, mais pas nuls. On propose de ranger cette association dans une alliance alticole (1 000 à 1 650 m d’altitude) de cet ordre est-méditerranéen.
L’équivalent des Cytiso villosi-Genistetalia monspessulanae en Méditerranée orientale paraît plutôt mal connu. Des bords méridionaux de la mer Noire, en Turquie, Korkmaz et al. (2008) décrivent plusieurs syntaxons à Erica arborea et parfois Arbutus unedo, mais sans autre lien synfloristique avec les Pistacio-Rhamnetea alaterni. Le tableau 36 synthétise six syntaxons nanophanérophytiques ; il décrit clairement une alliance originale à distribution pontico-caucasienne caractérisée par Vaccinium arctostaphylos, Rhododendron ponticum subsp. p., Rh. luteum, Ilex colchica, Smilax excelsa, Daphne pontica, le Vaccinio arctostaphyli-Rhododendrion pontici all. nov. hoc loco, typus nominis hoc loco : Ilici colchicae-Rhododendretum pontici (Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008) nov.
VRp1. « Calluno vulgaris-Arbutetum unedonis Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008 » (Korkmaz et al., 2008, tableau II), renommé ici Rubo sancti-Arbutetum unedonis (Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008) ass. nov. hoc loco, à Arbutus unedo, Erica arborea, Rhododendron ponticum subsp. p., Vaccinium arctostaphylos, Rubus sanctus ; typus nominis hoc loco : composante nanophanérophyique du relevé 34 du tableau II in Korkmaz et al. (2008, Acta Bot. Gallica 155 (4) : 499).
VRp2. « Carpino betuli-Populetum tremulae Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008 » (Ibid., tableau III).
VRp3. « Hedero helicis-Castaneetum sativae Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008 » (Ibid., tableau IV) ; surtout différenciés par des formes juvéniles d’arbres, les syntaxons VRp2 et VRp3 ne sont peut-être que deux variations d’une même association à préciser.
VRp4. « Rubo caucasici-Quercetum hartwissianae Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008 » (Ibid., tableau V), renommé ici Ilici colchicae-Rhododendretum pontici (Korkmaz, Yalçın, Güray Kutbay, Berk & Bilgin 2008) ass. nov. hoc loco, à Rhododendron ponticum subsp. p., Vaccinium arctostaphylos, Ilex colchica, Quercus hartwissiana j, Dioscorea communis, Pyrus communis (sous-espèce indiquée subsp. communis, mais probablement plutôt subsp. pyraster ou subsp. caucasica), Smilax excelsa, Sorbus umbellata s. l. j, Rubus caucasicus ; typus nominis hoc loco : composante nanophanérophytique du relevé 4 du tableau V in Korkmaz et al. (2008, Acta Bot. Gallica 155 (4) : 503).
VRp5. Groupement à Rhododendron caucasicum (Korotkov, 1995, tableau 3).
VRp6. Groupement à Rubus caucasicus (Ibid., tableau 2).
La photo 23 illustre une végétation de cette alliance observée dans les monts Kaçkar, au niveau de Ayder, à l’est de Trabzon (Turquie).

3. Quelques aspects des structures systématiques arbustives
À la fin de la synthèse sur la végétation forestière arborescente (de Foucault, 2020a, partie A § IV), on a tenté de dégager quelques « structures formelles systématiques » au niveau famille, autrement dit des catégories abstraites de végétation (dites F-structures systématiques) caractérisées par des combinaisons originales de familles végétales (de Foucault, 1987). On propose de réaliser une synthèse équivalente pour la végétation arbustive connue actuellement.
Pour cela, on va considérer les dix classes suivantes, avec leur abréviation : Betulo-Alnetea viridis (BAv), Franguletea alni (Fa), Cytisetea scopario-striati (Css), Rhamno-Prunetea spinosae (RPs), Salicetea purpureae (Sp), Nerio-Tamaricetea africanae (NTa), Kleinio-Euphorbietea canariensis (KEc), Zizipho-Searsietea tripartitae (ZSt), Erico-Morelletea fayae (EMf) et Pistacio-Rhamnetea alaterni (PRa), parfois en considérant en plus leurs variations majeures (ordres ou groupes d’alliances, reconnaissables par un chiffre arabe en fin de matricule). La diversité des genres et espèces est ensuite nivelée au niveau supérieur des familles (délimitation et nomenclature selon APG IV ; Angiosperm Phylogeny Group, 2016) pour établir un tableau brut des combinaisons de familles. En remplaçant les présences en chiffres romains par leur équivalent en chiffres arabes (V → 5…, + étant remplacé par 0,5, r par 0), on élabore un second tableau brut quantitatif qui peut être alors soumis à une analyse en composantes principales (ACP) afin d’en extraire des informations pertinentes. Cette ACP a été réalisée grâce au logiciel libre Past. Les sources utilisées sont reprises dans l’encadré ci-dessous.
Les figures 1a et 1b donnent les résultats selon les trois premiers axes factoriels qui extraient globalement un peu plus de 56 % de l’information contenue dans le tableau initial.
L’examen du gradient le long de l’axe 1 montre clairement un déterminisme climatique : les classes plutôt tempérées dans la partie droite, les classes plutôt méditerranéennes et steppiques vers la gauche. Les variations de classes sont le plus souvent rapprochées, à l’exception toutefois de BAv1 placé près des RPs ; il s’agit du Sorbo mougeotii-Lonicerion alpigenae, alliance justement de transition altitudinale entre les Rhamno-Prunetea spinosae (RPs) et le cœur des Betulo-Alnetea viridis (BAv).
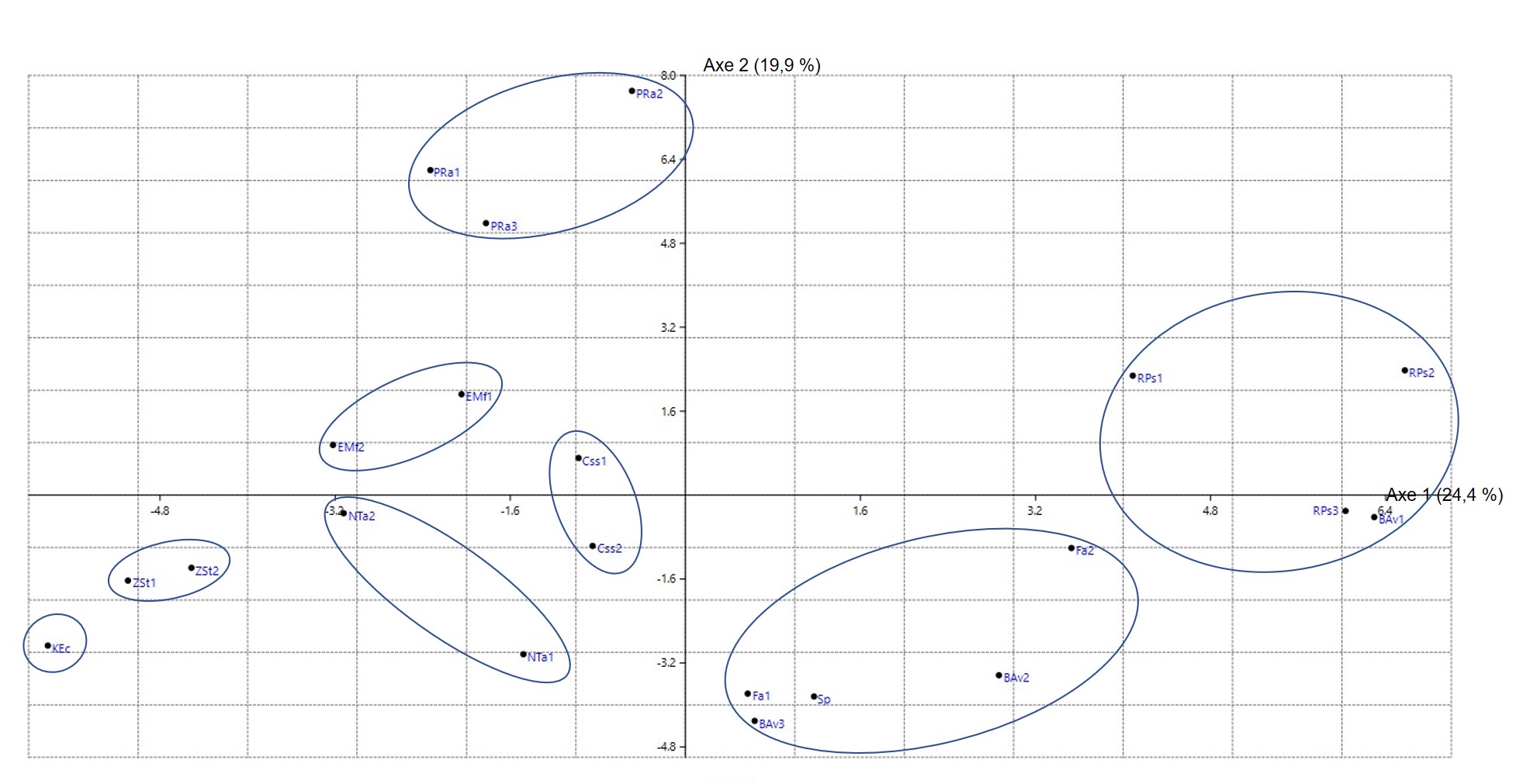
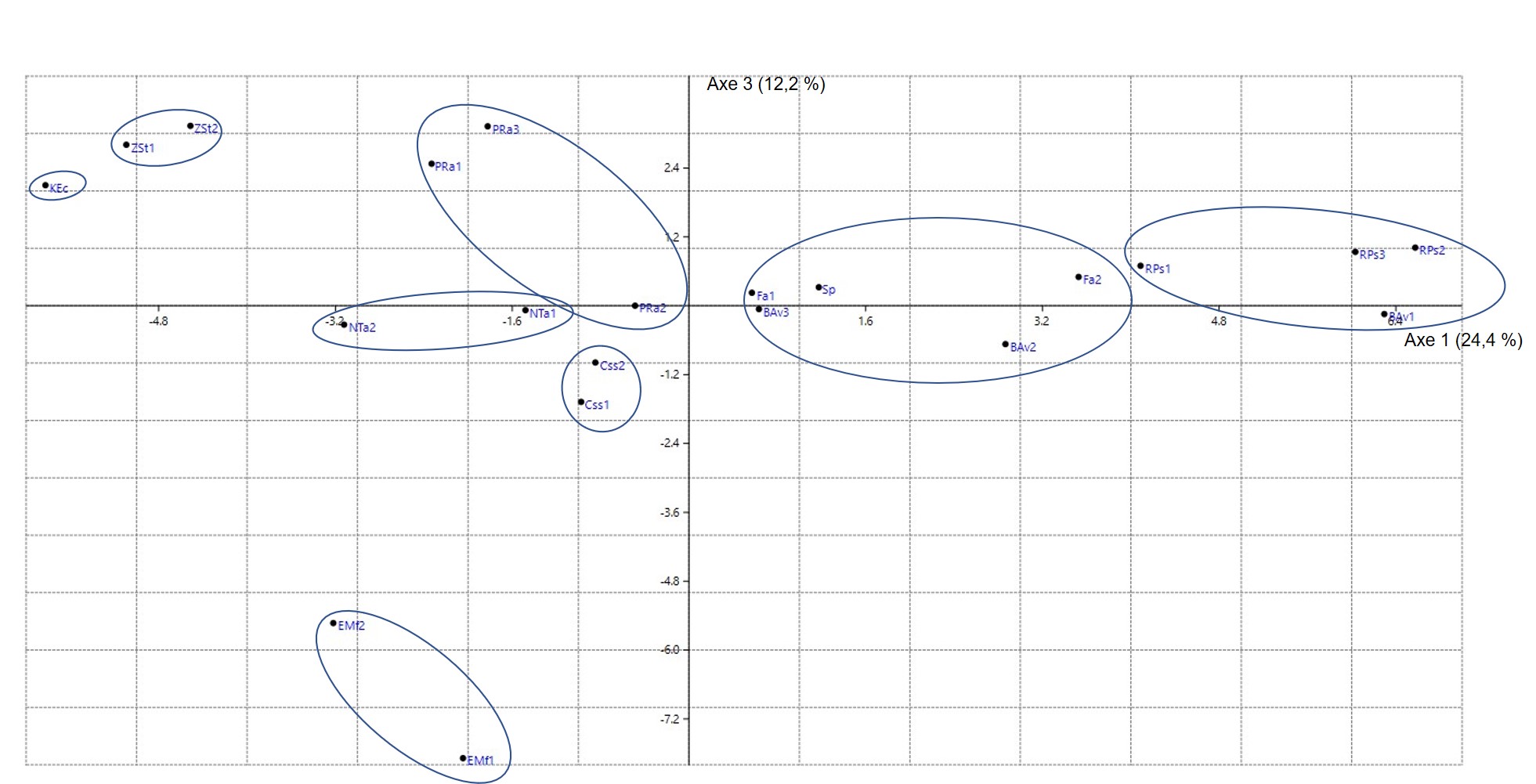
Pour aller plus loin dans l’information acquise par cette ACP, on va s’appuyer sur le tableau 37 qui dresse l’ordination optimale du tableau brut initial selon l’axe 1 de l’ACP. Les syntaxons rassemblés ici partagent les Rhamnaceae, Fabaceae, Rubiaceae, Solanaceae. Une coupure majeure passe entre les colonnes NTa2 et ZSt1 ; l’ensemble de droite est caractérisé par les Apocynaceae et des Asteraceae (surtout Warionia), et réunit les Zizipho-Searsietea tripartitae et les Kleinio-Euphorbietea canariensis, la F-structure associée à cette dernière classe ayant déjà été décrite sous le nom de S(Asclepiadaceae-Euphorbiaceae) (de Foucault, 1993a), à actualiser en S(Apocynaceae-Euphorbiaceae) suite à l’intégration des Asclepiadaceae dans les Apocynaceae. Quant aux Zizipho-Searsietea tripartitae, ils combinent les Anacardiaceae, Asparagaceae, Ephedraceae, Rhamnaceae et Fabaceae, F-structure originale nouvelle que l’on peut dénommer S(Ephedraceae-Anacardiaceae).
À gauche de la séparation NTa2/ZSt1, diverses F-structures se dégagent, partageant les Rosaceae, Ranunculaceae (par le genre Clematis), Rosaceae j et Pinaceae j. Les NTa recouvrent les Nerio-Tamaricetea africanae et se caractérisent essentiellement par les Tamaricaceae, secondairement par les Rosaceae : S(Rosaceae-Tamaricaceae) ; la variation NTa2 (Nerio oleandri-Viticetalia agni-casti) se différencie de NTa1 (Tamaricetalia africanae) par les Asparagaceae, Smilacaceae, Lamiaceae (Vitex), Apocynaceae.
L’ensemble ‘Css + EMf’ se différencie par la rareté des Caprifoliaceae, Thymelaeaceae et Fagaceae j. L’ensemble Css recouvre les Cytisetea scopario-striati et se caractérise par une relative pauvreté familiale, ainsi que par l’optimum des Fabaceae, avec la variation Css1 (six alliances ibéro-atlantiques, dont l’Ulici europaei-Cytision striati) différenciée par les Ericaceae et Fagaceae j. L’ensemble EMf recouvre les Erico arboreae-Morelletea fayae macaronésiens ; il est fortement différencié par les Ericaceae, Aquifoliaceae, Plantaginaceae, Myricaceae, Lamiaceae, Hypericaceae, Primulaceae (genre Heberdenia) et Lauraceae j (liens dynamiques avec les laurisylves), avec la variation EMf1 différenciée par les Thymelaeaceae, Campanulaceae (Canarina) et Rhamnaceae j.
L’ensemble ‘RPs + BAv + Fa + Sp’ se distingue essentiellement par les Salicaceae et les Betulaceae j. Le sous-ensemble ‘RPs + BAv’ est caractérisé par les Betulaceae et Grossulariaceae. Les Rhamno-Prunetea spinosae (RPs) sont recouverts par une F-structure déjà reconnue en 1987, les S(Oleaceae-Caprifoliaceae), caractérisée par la combinaison des Caprifoliaceae, Cornaceae, Adoxaceae, Oleaceae, Celastraceae ; elle présente des variations à Dioscoreaceae (RPs1, soit les Pyro spinosae-Rubetalia ulmifolii), à Buxaceae, Berberidaceae et Cupressaceae (RPs2, soit les Prunetalia spinosae), la troisième différenciée négativement (RPs3, soit les Sambucetalia racemosae). Les Betulo-Alnetea viridis (BAv) se différencient surtout négativement vis-à-vis de la classe précédente ; on peut définir la S(Caprifoliaceae-Salicaceae), qui présente une variation BAv1 déjà évoquée plus haut (Sorbo mougeotii-Lonicerion alpigenae), à Adoxaceae, Fabaceae j et Sapindaceae (Acer) j, une variation BAv2 (Alnion viridis, Salicion silesiacae, Pruno petraeae-Sorbion aucupariae, Salicion waldsteinianae) peu différenciée et une variation BAv3 (Salicion pentandrae) encore moins différenciée (disparition des Caprifoliaceae, Thymelaeaceae, Grossulariaceae, Rosaceae, Rhamnaceae, Fabaceae, Fagaceae j, un degré supplémentaire d’appauvrissement floristique selon des contraintes climatiques et écologiques).
Fa et Sp se rapprochent par les Salicaceae j et l’optimum des Betulaceae j. Fa (Franguletea alni) est peu caractérisé, variant en Fa1 (Salicetalia auritae, à Myricaceae) et Fa2 (Rubetalia plicati, à Caprifoliaceae, Rosaceae, Fagaceae j, Fabaceae j). Sp (Salicetea purpureae) se réduit surtout aux Salicaceae, Rosaceae et Oleaceae : S(Rosaceae-Salicaceae).
Il reste enfin la F-structure attachée aux Pistacio-Rhamnetea alaterni (RPa), caractérisée par les Anacardiaceae, Asparagaceae, Smilacaceae, Cupressaceae (Juniperus), Myrtaceae, Dioscoreaceae, Oleaceae, Santalaceae, Cistaceae, Arecaceae (Chamaerops) et Cupressaceae j, l’absence des trois dernières familles en PRa3 (soit les Rubio tenuifoliae-Pistacietalia terebinthi) n’est peut-être pas significative, la végétation est-méditerranéenne étant bien moins connue que celle des régions ouest-méditerranéennes. Dans ce lot de familles, on en reconnaît certaines d’optimum tropical (ou à dispersion eurytropicale) qui témoignent de la position de relais de cette classe entre la végétation tempérée et la végétation tropicale. On définit ici la nouvelle S(Smilacaceae-Anacardiaceae), qui varie, outre PRa3 déjà discuté, en PRa1 (Pistacio-Rhamnetalia alaterni) différencié négativement et PRa2 (Cytiso-Genistetalia monspessulanae) à Ericaceae et Adoxaceae (Viburnum tinus, peut-être non significatif comme précisé plus haut).
La végétation arbustive tropicale qui remplace les précédentes vers les basses latitudes est encore très peu décrite. On peut citer le Jasmino dichotomi-Zanthoxyletum zanthoxyloidis B. Foucault et al. 2000 (de Foucault et al., 2000, tableau 8) du littoral togolais, à Oleaceae, Rutaceae, Lauraceae, Salicaceae, Rubiaceae, Malvaceae, Apocynaceae, Capparaceae, Chrysobalanaceae, Dichapetalaceae, Ehretiaceae, Annonaceae, Lamiaceae, Passifloraceae, Connaraceae, Asteraceae, Meliaceae j. La composition familiale globale de fourrés guadeloupéens de basse altitude (de Foucault, 1991b, tableau 10 : niveau 1) combine les Primulaceae, Anacardiaceae, Lauraceae, Rhamnaceae, Rutaceae, Capparaceae, Scrophulariaceae, Cactaceae, Apocynaceae, Celastraceae, Solanaceae, Sapindaceae, Annonaceae, Salicaceae, Cucurbitaceae, Simaroubaceae, Boraginaceae, Fabaceae, Poygonaceae, Bignoniaceae, Piperaceae, Malpighiaceae, Rubiaceae, Asteraceae, Myrtaceae, Euphorbiaceae, Melastomataceae, Clusiaceae, Araliaceae. On reconnaît là quelques familles citées pour les S(Smilacaceae-Anacardiaceae).
4. Conclusion
Sur la base d’environ trois cents syntaxons (associations ou groupements), cette synthèse portant sur le sud-ouest de l’Europe et le Maghreb, avec quelques données moins conséquentes vers la Méditerranée orientale, offre une révision significative des Pistacio lentisci-Rhamnetea alaterni. La liste qui suit synthétise la hiérarchie obtenue (dix-neuf alliances pour le premier ordre, dix pour le deuxième, cinq pour le troisième). Les unités présentes en France sont surlignées en gris (quatre alliances pour le premier ordre, deux pour le second). Pour faciliter la recherche des tableaux correspondants, on a indiqué leur numéro en fin de ligne, cette liste pouvant alors avoir valeur d’index.
PISTACIO LENTISCI-RHAMNETEA ALATERNI Julve ex Julve & B. Foucault in B. Foucault 2020 (tableau 1)
Eu-Pistacio lentisci-Rhamnenea alaterni (tableaux 2 à 32)
Pistacio lentisci-Rhamnetalia alaterni Rivas Mart. 1975 (tableaux 2 à 21)
Clematido cirrhosae-Pistacion lentisci nov. (tableau 2)
Asparago albi-Rhamnion oleoidis Rivas Goday ex Rivas Mart. 1975 (tableau 3)
Periplocion laevigatae Rivas Mart. 1975 (tableau 4)
Rhamno myrtifoliae-Juniperion phoeniceae nov. (tableau 6)
Arbuto unedonis-Laurion nobilis Rivas Mart., Fern.Gonz. & Loidi 1999 (tableau 7)
Euphorbio dendroidis-Myrtion communis nov. (tableau 8)
Calicotomo infestae-Teucrion fruticantis nov. (tableau 9)
Rhamno lycioidis-Quercion cocciferae Rivas Goday ex Rivas Mart. 1975 (tableau 10)
Amelanchiero ovalis-Juniperion phoeniceae nov. (tableau 11)
Juniperion turbinatae Rivas Mart. 1975 (tableau 13)
Phillyreo mediae-Juniperion turbinatae nov.
Oleo sylvestris-Paliurion spinae-christi (Biondi, Casavecchia, Biscotti & Pesaresi 2014) nov. (tableau 14)
Juniperion oxycedro-lagunae Cano, Rodríguez-Torres, Pinto Gomes, García-Fuentes, Torres, Salazar, Ruiz-Valenzuela, Cano-Ortiz & Montilla 2007 (tableau 15)
Prasio majoris-Cytision lanigeri nov. (tableau 16)
Tetraclinido articulatae-Quercion cocciferae (Hadjadj & R.J. Loisel 1999) nov. (tableau 17)
Genisto tricuspidatae-Cytision spinosi B. Foucault & Dahmani nov. (tableau 18)
Genisto quadriflorae-Pistacion lentisci nov. (tableau 19)
Tetraclinido articulatae-Pistacion atlanticae Rivas Mart., M.J. Costa & Izco 1986 (tableau 20)
Genistion ifniensi-tricuspidatae nov. (tableau 21)
Cytiso villosi- Genistetalia monspessulanae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 (tableaux 22 à 32)
Pruno lusitanicae-Ericion arboreae nov. (tableau 22)
Rhododendrion baetici (Rivas Mart. & Sánchez-Mata 2001) nov. (tableau 23)
Quercion lusitanicae Rothm. 1954 (tableau 24)
Ericion arboreae (Rivas Mart. ex Rivas Mart., M.J. Costa & Izco 1986) Rivas Mart. 1987 (tableau 25)
Genistion monspessulano-linifoliae Rivas Mart., Galán & Cantó in Rivas Mart. et al. 2002 (tableau 26)
Flueggeion tinctoriae Rivas Goday 1964 (tableau 28)
Genisto numidicae-Ericion arboreae nov. (tableau 29)
Tetraclinido articulatae-Ericion arboreae nov. (tableau 30)
Cytision spinoso-villosi B. Foucault & Meddour nov. (tableau 31)
Hippocrepido emeroidis-Ericion arboreae nov. (tableau 32)
Rubio tenuifoliae-Pistacienea terebinthi nov. (tableaux 33 à 35)
Rubio tenuifoliae-Pistacietalia terebinthi nov. (tableaux 33 à 35)
Rhamno graecae-Juniperion turbinatae M.J. Costa, Géhu, Peris, Biondi & Arnold 1984
Ceratonio siliquae-Pistacion lentisci Zohary & Orshan 1959 (tableau 33)
Pistacio terebinthi-Buxion sempervirentis nov. (tableau 34)
Ephedro aphyllae-Retamion raetam nov. (tableau 35)
Asparago acutifolii-Berberidion creticae nov.
Associées à ce schéma synsystématique, quatre F-structures systématiques originales ont été aussi reconnues (tableau 37). Il sera intéressant de les tester dans l’avenir au niveau de l’empire holarctique.
Il reste à espérer que cette synthèse, sans être évidemment exhaustive, facilitera la dénomination des syntaxons déjà décrits, la détection des homonymes illégitimes et la reconnaissance de syntaxons nouveaux, qu’elle stimulera aussi des études phytosociologiques modernes sur les fourrés sempervirents est-méditerranéens encore trop peu connus. Pour les recherches futures, on doit encore recommander de mieux séparer la végétation forestière proprement dite et la végétation des lisières arbustives ; comme l’écrivent Barbero & Quézel (1983 : 10), « [il] convient de remarquer que […] la distinction entre les associations strictement forestières et celles relatives aux groupements de manteau et de lisière n’a pas été effectuée avec suffisamment de rigueur phytosociologique, ainsi que nous en ont convaincus les recherches entreprises au Maroc en coopération avec S. Rivas-Martínez, où l’individualisation d’unités se rattachant à ces deux types de structures est non seulement nécessaire à la compréhension de l’édifice phytosociologique mais peut-être surtout de sa dynamique et de sa valeur phytoécologique. ». La séparation des formes adultes et des formes juvéniles (symbole j) des essences d’arbres est indispensable à cette distinction.
On peut ajouter qu’il en est de même en ce qui concerne la distinction entre landes et fourrés : trop souvent les chaméphytes et les nanophanérophytes relevant des Ericaceae et des Cistaceae sont mélangés dans les relevés. Les landes sont dominées par des chaméphytes, auxquels peuvent s’associer des formes juvéniles (j) des taxons normalement arbustifs ou arborescents et des taxons herbacés vivaces, les thérophytes formant des communautés indépendantes, qui furent cependant longtemps mêlées aux landes (cf. Braun-Blanquet et al., 1940 ; Braun-Blanquet et al., 1952). Les fourrés sont dominés par des nanophanérophytes (incluant les taxons grimpants), auxquels peuvent s’associer des taxons normalement sous-arbustifs sous des formes vigoureuses (symbole !) et des taxons normalement arborescents sous des formes juvéniles (j).
En présentant le Polygalo balansae-Juniperion phoeniceae, on a évoqué son rattachement aux Warionio saharae-Withanietalia frutescentis et aux Zizipho loti-Searsietea tripartitae par « enchaînement » ; dans un article récent précédent, on a évoqué la même situation pour le Trifolio thalii-Ranunculion aurasiaci vis-à-vis des Sagino saginoidis-Nardetalia strictae (de Foucault, 2021c). Ce phénomène intéressant se retrouve en systématique végétale, les familles végétales par enchaînement les plus classiques étant les Ranunculaceae et les Rosaceae (Spichiger et al., 2002). Ce n’est pas la première fois qu’un parallélisme entre idiosystématique et synsystématique est souligné, c’est ainsi le cas des variations parallèles (de Foucault, 1994). Ce parallélisme peut d’ailleurs être étendu à l’« ethnosystématique », c’est-à-dire la systématique des formes créées par les hommes ; à titre d’exemple, il existe au moins deux types de barrière traditionnelle de prairie présentant chacune une variation type et une variation où la barrière est encadrée par deux murets de granite, le montant pivot étant pris dans une pierre percée (de Foucault, 1991c). Ces concepts d’enchaînement et de variation parallèle s’avèrent être des invariants transdisciplinaires, que l’on devrait donc retrouver dans d’autres démarches systématiques.
Bibliographie
Aguiar C. & Capelo J., 1995. Anotação sobre a posição dos medronhais do Alto Minho. In Notas do Herbário da Estação Florestal Nacional (LISFA), I. Silva Lusitana 3 (1) : 123-125.
Aguiar C., Esteves A. & Penas A., 1999. As comunidades de Buxus sempervirens do Sector Lusitano-Duriense. Quercetea 1 : 177-185.
Allier C. & Lacoste A., 1980. Maquis et groupements végétaux de la série du chêne vert dans le bassin du Fango (Corse). Ecologia Mediterranea 5 : 59-82.
Angiosperm Phylogeny Group, 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181, 1-20.
Bacchetta G., Bagella S., Biondi E., Farris E., Filigheddu R. & Mossa L., 2003. Su alcune formazioni a Olea europaea L. var. sylvestris Brot. della Sardegna. Fitosociologia 40 (1) : 49-53.
Barbagallo C., Brullo S. & Fagotto F., 1979. Boschi di Quercus ilex L. del territorio di Siracusa e principali aspetti di degradazione. Pubblicazioni dell’Istituto Botanico dell’Università di Catania : 1-24.
Barbero M. & Quézel P., 1976. Les groupements forestiers de Grèce centro-méridionale. Ecologia Mediterranea 2 : 3-86.
Barbero M. & Quézel P., 1979. Contribution à l’étude des groupements forestiers de Chypre. Documents phytosociologiques, NS, IV : 9-33.
Barbero M., Quézel P. & Rivas-Martínez S., 1981. Contribution à l’étude des groupements forestiers et préforestiers du Maroc. Phytocoenologia 9 (3) : 311-412.
Barbero M. & Quézel P., 1983. Le problème des manteaux forestiers des Pistacio-Rhamnetalia alaterni en Méditerranée orientale ; essai syntaxonomique. Colloques phytosociologiques VIII, Les lisières forestières : 9-20.
Benabid A., 1988. Contribution à l’étude des groupements non forestiers du Moyen Atlas central (Maroc). Bulletin de l’Institut scientifique de Rabat 12 : 79-98.
Benabid A. & Fennane M., 1994. Connaissances sur la végétation du Maroc : phytogéographie, phytosociologie et séries de végétation. Lazaroa 14 : 21-97.
Biondi E. & Mossa L., 1992. Studio fi tosociologico del promontorio di Capo S. Elia e dei Colli di Cagliari (Sardegna). Documents phytosociologiques, NS, XIV : 1-44.
Biondi E., Filigheddu R.S. & Farris E., 2001. Il paesaggio vegetale della Nurra (Sardegna nord-occidentale). Fitosociologia 38 (2), suppl. 2 : 3-105.
Biondi E., Casavecchia S., Pesaresi S., Gangale C. & Uzunov C., 2014. New syntaxa for the prodrome of Italian vegetation. Plant Biosystems 148 (4) : 723-727.
Bolòs O. (de), 1970. À propos de quelques groupements végétaux observés entre Monaco et Gênes. Vegetatio 21 : 49-73.
Bolòs O. (de), 1973. La vegetación de la Serreta Negra de Fraga. Memorias de la Real Academia de Ciencias de Barcelona 42 (6) : 269-313.
Bolòs O. (de) & Molinier R., 1958. Recherches phytosociologiques dans l’île de Majorque. Collectanea Botanica (Barcelona) 5 (3) : 699-865.
Bolòs O. (de) & Molinier R., 1969. Vue d’ensemble de la végétation des îles Baléares. Vegetatio 17 : 251-270.
Bolòs O. (de), Molinier R. & Montserrat P., 1970. Observations phytosociologiques dans l’île de Minorque. Acta Botanica Barcinonensis 5 : 1-150.
Bolòs O. (de) & Molinier R., 1984. Vegetation of the Pityusic Islands. In H. Kuhbier, J.A. Alcover & G. Arellano (eds.), Biogeography and ecology of the Pityusic Islands, Dr. W. Junk, Den Haag : 185-221.
Braun-Blanquet J., 1925. Die Brachypodium ramosum-Phlomis lychnitis-Assoziation der Roterdeböden Südfrankreichs. Veröffentlichungen des Geobotanischen Institutes der E.T.H. Stiftung Rübel 3 : 304-320.
Braun-Blanquet J., 1953-54. Irradiations européennes dans la végétation de la Kroumirie. Vegetatio IV : 182-194.
Braun-Blanquet J. & Maire R., 1924. Études sur la flore et la végétation marocaines ; comptes-rendus des herborisations de la Société botanique de France, session du Maroc, 1921. Mémoires de la Société des sciences naturelles du Maroc VIII, 1re partie : 3-239.
Braun-Blanquet J., Molinier R. & Wagner H., 1940. Classe Cisto-Lavanduletea (landes siliceuses à cistes et lavandes). Prodrome des groupements végétaux 7 : 1-51.
Braun-Blanquet J., Roussine N. & Nègre R., 1952. Les groupements végétaux de la France méditerranéenne. CNRS, Paris, 297 p.
Braun-Blanquet J., Pinto da Silva A.R. & Rozeira A., 1956. Résultats de deux excursions géobotaniques à travers le Portugal septentrional et moyen, II – Chênaies à feuilles caduques [Quercion occidentale] et chênaies à feuilles persistantes [Quercion fagineae] au Portugal). Agronomia Lusitana 18 (3) : 167-234.
Braun-Blanquet J., Pinto da Silva A.R. & Rozeira A., 1964. Résultats de trois excursions géobotaniques à travers le Portugal septentrional et moyen, III – Landes à cistes et Ericacées (Cisto-Lavanduletea et Calluno-Ulicetea). Agronomia Lusitana 23 (4) : 229-313.
Brullo S., Gianguzzi L., La Mantia A. & Siracusa G., 2008. La classe Quercetea ilicis in Sicilia. Bollettino dell’Accademia Gioenia di Scienze Naturali 41 (369) : 1-80.
Cabezudo B., Nieto J.M. & Pérez-Latorre A., 1989. 4 – Contribución al conocimiento de la vegetación edafófilo-serpentinícola del sector Rondeño (Málaga, España). Acta Botanica Malacitana 14 : 291-294.
Camarda I., Lucchese F., Pignatti S. & Pignatti S., 1995. La vegetazione dell’area Pantaleo-Gutturu Mannu-Punta Maxia-Monte Arcazu nel Sulcis Iglesiente (Sardegna sud-occidentale). Webbia 49 (2) : 141-177.
Cano E., Rodríguez-Torres A., Pinto Gomes C., García-Fuentes A., Torres J.A., Salazar C., Ruiz-Valenzuela L., Cano-Ortiz A. & Montilla R.J., 2007. Analysis of the Juniperus oxycedrus L. communities in the centre and south of the Iberian peninsula (Spain and Portugal). Acta Botanica Gallica 154 (1) : 79-99.
Capelo J.H., Costa J.C., Lousã M. & Mesquita S., 2001. A aliança Quercion fruticosae Rothmaler 1954 em. Rivas-Martínez, Lousã, T.E. Díaz, Fernández-González & J.C. Costa 1990. Quercetea 3 : 99-110.
Casavecchia S., Biscotti N., Pesaresi S. & Biondi E., 2015. The Paliurus spina-christi dominated vegetation in Europe. Biologia, sect. Botany, 70 (7) : 879-892.
Choisnet G., 2019. Approche phytosociologique et symphytosociologique des végétations et des paysages du Bas-Vivarais (nord du bassin méditerranéen français). Thèse de doctorat d’État, Université de Bretagne occidentale, Brest, 347 p. et annexes.
Costa J.C., Lopes M.C., Capelo J. & Lousã M., 2000. Sintaxonomia das comunidades de Prunus lusitanica L. subsp. lusitanica no ocidente da Península. Silva Lusitana 8 (2) : 253-263.
Costa J.C., Monteiro-Henriques T., Bingre P. & Dalila Espírito-Santo D., 2015. Warm-temperate forests of central Portugal: a mosaic of syntaxa. In E.A. Box & K. Fujiwara, Warm-temperate deciduous forests around the northern hemisphere, Geobotany Studies, Springer, Bern : 97-117.
Costa M., Géhu J.-M., Peris J.B., Biondi E. & Arnold N., 1984. Sobre la vegetación termomediterranea litoral de la isla de Chipre. Documents phytosociologiques, NS, VIII : 365-376.
Dahmani M., 1997. Le chêne vert en Algérie. Syntaxonomie, phytoécologie et dynamique des peuplements. Thèse de doctorat de l’université des sciences et technologies H. Boumediene, Alger, 383 p.
Danin A. & Orshan G., 1999. Vegetation of Israel, I – Desert and coastal vegetation. Backhuys Publishers, Leiden, 346 p.
Di Pietro P. & Misano G., 2010. Shrubland and garrigue vegetation in the « Gravine » gorges (Apulia region, south-eastern Italy). Acta Botanica Gallica 157 (2) : 195-229.
Díaz T.E. & Fernández-Prieto J.A., 1994. La vegetación de Asturias. Itinera Geobotanica 8 : 243-520.
Díez Garretas B. & Asensi A., 2013. The coastal plant communities of Juniperus macrocarpa in the Mediterranean region. Plant Biosystems 148 (3) : 429-438.
Dieleman P., 1970. Le Pyreto-Paliuretum spinae-christi Kuhnholtz-Lordat 1954 aux environs de Montpellier. Proceedings, Koninklijke Nederlandse Akademie van Wetenschappen, ser. C, 73 (3) : 254-270.
Eig A., 1939. The vegetation of the light soils belt of the coastal plain of Palestine. Palestine journal of botany, Jerusalem series 1 : 255-308.
Eig A., 1946. Synopsis of the phytosociological units of Palestine. Palestine journal of botany, Jerusalem series III (4) : 183-246.
El Hamrouni A. & Loisel R., 1979. Contribution à l’étude de la tétraclinaie tunisienne : les groupements des djebels Bou Kornine et Ressas. Ecologia Mediterranea 4 : 133-139.
Farris E., Zecchi S. & Filigheddu R., 2007. Phytosociological study of the shrub and pre-forest communities of the effusive substrata of NW Sardinia. Fitosociologia 44 (2) : 55-81.
Feliu Xifra P., 2003. Corologiá y vegetación de la Albera. Thèse de doctorat, Université de Barcelone.
Fennane M., 1986. Contribution à l’étude phytosociologique des tétraclinaies du Maroc. Bulletin de l’Institut scientifique de Rabat 10 : 57-78.
Fernández Casas J., 1970. Notas sobre vegetación. Publacciones del Instituto de Biologia Aplicada 49 : 111-120.
Foucault B. (de), 1987. Nouvelles recherches sur les structures systématiques végétales : caractérisation, ordination, signification. Phytocoenologia 15 (2) : 159-199.
Foucault B. (de), 1991a. Introduction à une systémique des végétations arbustives. Documents phytosociologiques, NS, XIII : 63-104.
Foucault B. (de), 1991b. Nouvelles considérations sur la végétation de la Guadeloupe (Antilles françaises). Phytocoenologia 19 (4) : 445-478.
Foucault B. (de), 1991c. Un chapitre de technologie rurale : la barrière traditionnelle de prairie, typologie et dynamique. Mémoires de la Société linnéenne Nord-Picardie II : 1-107.
Foucault B. (de), 1993a. Remarques sur la végétation du Maroc et de l’Algérie. Bulletin de la Société botanique du Centre-Ouest, NS, 24 : 267-285.
Foucault B. (de), 1993b. II – Données phytosociologiques. In B. de Foucault & J.-P. Cornu, Itinéraire botanique d’un groupe de Linnéens en Crète, 29 avril-13 mai 1991 (2e partie). Bulletin de la Société linnéenne Nord-Picardie 11 : 85-105.
Foucault B. (de), 1994. Extension à la phytosociologie d’un concept botanique : la variation parallèle. Candollea 49 : 121-127.
Foucault B. (de), 2012. Contribution au prodrome des végétations de France : les Betulo carpaticae-Alnetea viridis Rejmánek in Huml, Lepš, Prach & Rejmánek 1979. Journal de botanique de la Société botanique de France 60 : 47-68
Foucault B. (de), 2015a. Contribution au prodrome des végétations de France : les Pegano harmalae-Salsoletea vermiculatae Braun-Blanq. & O. Bolòs 1958. Journal de botanique de la Société botanique de France 72 : 41-59.
Foucault B. (de), 2015b. Quelques associations arbustives nouvelles pour la Malepère (Aude centrale). Bulletin de la Société botanique du Centre-Ouest, NS, 46 : 244-260.
Foucault B. (de), 2016. Données phytosociologiques collectées lors de sorties du groupe botanique de la SESA durant l’année 2015. Bulletin de la Société d’études scientifiques de l’Aude CXV : 137-145.
Foucault B. (de), 2019a. Quelques données phytosociologiques sur les landes et les fourrés éricoïdes des Albères (département des Pyrénées-Orientales, France). Bulletin de la Société botanique du Centre-Ouest, NS, 50 : 366-371.
Foucault B. (de), 2019b. Contribution à la connaissance phytosociologique de pelouses, prairies et garrigues de la Malepère (département de l’Aude, France). Evaxiana 5 : 156-186.
Foucault B. (de), 2020a. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : 5-248.
Foucault B. (de), 2020b. Approche théorique et pratique du tableau phytosociologique. Botanique 6 : 37-51.
Foucault B. (de), 2020c. Données phytosociologiques collectées lors de sorties du groupe botanique de la Sesa durant l’année 2019. Bulletin de la Société d’études scientifiques de l’Aude CXIX : 95-102.
Foucault B. (de), 2020d. Sur la synsystématique des Rosmarinetea officinalis ouest-méditerranéens. Carnets botaniques 4 : 1-27 et 26 tableaux. DOI : https://doi.org/10.34971/s9d8-qj43
Foucault B. (de), 2021a. Apports de la 155e session extraordinaire de la Société botanique de France à la connaissance de la végétation provençale. Journal de botanique de la Société botanique de France 93 : 48-72.
Foucault B. (de), 2021b. Quelques aspects de la végétation forestière et de lisière du Bas-Vivarais. Carnets botaniques 49 : 1-5 + deux tableaux. DOI : https://doi.org/10.34971/P9D3-D685.
Foucault B. (de), 2021c. Miscellanées phytosociologiques : quelques syntaxons nouveaux, validés, complétés ou corrigés. Carnets botaniques 44 : 1-23 + 28 tableaux. DOI : https://doi.org/10.34971/AKGG-T733.
Foucault B. de) & Julve Ph., 1991. Données phytosociologiques sur la dix-septième session de la SBCO en Languedoc-Roussillon ; réflexions sur les associations arborescentes méditerranéennes. Bulletin de la Société botanique du Centre-Ouest, NS, 22 : 391-420.
Foucault B. (de), Akpagana K., Batawila K. & Bouchet Ph., 2000. Contribution à une étude phytosociologique du littoral sableux du Togo. Acta Botanica Gallica 147 (4) : 333-344.
Foucault B. (de), Bensettiti F., Noble V. & Paradis G., 2012. Contribution au prodrome des végétations de France : les Nerio oleandri-Tamaricetea africanae Braun-Blanq. & O. Bolòs 1958. Journal de botanique de la Société botanique de France 58 : 41-54.
Foucault B. (de), Lazare J.-J. & Bioret F., 2013a. Contribution au prodrome des végétations de France : les Cytisetea scopario-striati Rivas-Mart. 1975. Journal de botanique de la Société botanique de France 64 : 69-90.
Foucault B. (de), El Beyrouthy M. & Arnold-Aspotolidès N., 2013b. Résultats d’investigations phytosociologiques récentes au Liban central. Acta Bot. Gallica: Botany Letters 160 (1) : 65-75.
Foucault B. (de) & Royer J.-M., 2014. Contribution au prodrome des végétations de France : les Franguletea alni Doing ex V. Westh. in V. Westh. & den Held 1969. Journal de botanique de la Société botanique de France 66 : 83-106.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Foucault B. (de) & Royer J.-M., 2019. Quelques données phytosociologiques sur le bas Languedoc à propos de la session de la SBF en avril 2018. Journal de botanique de la Société botanique de France 87 : 83-96.
Foucault B. (de) & Cornier Th., 2020. Contribution au prodrome des végétations de France : les Salicetea purpureae et les Populo albae-Salicetea albae. Documents phytosociologiques, 3e série, 13 : 277-360.
Franquesa i Codinach T., 1995. El paisatge vegetal de la península del Cap de Creus. Arxius de la secció de ciéncias; Institut d’Estudis Catalans CIX : 1-628.
Fuente García V. (de la), Rufo Nieto L., Rodríguez González N. & Amils Pibernat R., 2007. Los adelfares del suroeste de la Península Ibérica. Lazaroa 28 : 5-14.
Gamisans J. & Muracciole M., 1984. La végétation de la Réserve naturelle de la presqu’île de Scandola (Corse) ; étude phytosociologique et cartographie au 1/10 000e. Ecologia Mediterranea X (3-4) : 159-205.
Gauberville Ch., Panaïotis Ch., Bioret F. & Capelo J., 2019. Analyse phytosociologique et phytoécologique des formations forestières à pin laricio de Corse (Pinus nigra J.F. Arnold subsp. laricio Maire). Ecologia Mediterranea 45 (1) : 85-103, + 4 tableaux on line.
Géhu J.-M. & Géhu-Franck J., 1986. Données synsystématiques et synchorologiques sur la végétation du littoral tunisien, de Bizerte à Gabès ; I – La végétation psammophile. Documents phytosociologiques, NS, X (2) : 127-155.
Géhu J.-M., Biondi E., Géhu-Franck J. & Taffetani F., 1987. Données sur la végétation maritime du littoral oriental de la Corse. In M.J. del Arco & W. Wilpret, V Jornadas de fitosociologia, Vegetación de riberas de agua dulce, II : 363-391.
Géhu J.-M., Costa M. & Uslu T. 1988. Aperçu synécologique d’Euphorbia dendroides sur le littoral méridional de la Turquie. Documents phytosociologiques, NS, XI : 607-612.
Géhu J.-M., Costa M. & Uslu T. 1990. Analyse phytosociologique de la végétation littorale des côtes de la partie turque de l’île de Chypre dans un souci conservatoire. Documents phytosociologiques, NS, XII : 203-234.
Géhu J.-M., Apostolidès N., Géhu-Franck J. & Arnold K., 1992a. Premières données sur la végétation littorale des îles de Rhodos et de Karpathos (Grèce). Colloques phytosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 545-582.
Géhu J.-M., Kaabèche M. & Gharzouli R., 1992b. Obsevations phytosociologiques sur le littoral kabyle de Bejaia à Djijel. 1992. Documents phytosociologiques, NS, XIV : 305-322.
Géhu J.-M., Biondi E. & Bournique C., 1992c. Glanures phytosociologiques sur les côtes de Provence. Colloques phyosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 147-157.
Géhu J.-M., Uslu T. & Costa M., 1992d. Apport à la connaissance phytosociologique du littoral sud de la Turquie méditerranéenne. Colloques phyosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 591-622.
Géhu J.-M. & Biondi E., 1994. Végétation du littoral de la Corse. Essai de synthèse phytosociologique. Braun-Blanquetia 13 : 1-149.
Géhu J.-M. & Sadki N., 1995. Remarques de phytosociologie et de synchorologie comparées sur le littoral algérois. Documents phytosociologiques XV : 341-357.
Gerasimidis A., Fotiadis G. & Panajiotidis S., 2006. Contribution to the phytosociological study of low-altitude Buxus sempervirens (Buxaceae) formations (Mt Olympos, Greece). Proceedings of IV Balkan Botanical Congress, Sofia : 301-307.
Gianguzzi L., Ilardi V., Caldarella O., Cusimano D., Cuttonaro P. & Romano S., 2012. Phytosociological characterization of the Juniperus phoenicea L. subsp. turbinata (Guss.) Nyman formations in the Italo-Tyrrhenian Province (Mediterranean Region). Plant Sociology 49 (2) : 3-28.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Gómez-Mercado F., 2011. Vegetación y flora de la Sierra de Cazorla. Guineana 17 : 1-481.
Guinochet M., 1980. Essai sur quelques syntaxons des Cisto-Rosmarinetea et des Quercetea ilicis d’Algérie et de Tunisie. Phytocoenologia 7 : 436-466.
Guinochet M. & Drouineau G., 1944. Notes sur la végétation et le sol aux environs d’Antibes (Alpes-Maritimes). Recueil des travaux de l’Institut botanique de Montpellier I : 22-40.
Hadjadj S., 1991. Les peuplements de Tetraclinis articulata sur le littoral d’Oran (Algérie). Ecologia Mediterranea 17 : 63-78.
Hadjadj-Aoul S. & Loisel R., 1999. Thuya de Berbérie (Tetraclinis articulata (Vahl) Masters). Les peuplements forestiers et préforestiers. Documents phytosociologiques, NS, XIX : 229-285.
Hamzaoğlu E., Duran A. & Menemen Y., 2002. The vegetation of the Kazankaya canyon (Yozgat, Çorum). Gazi Üniversitesi Fen Bilimleri Enstitüsü Dergisi 15 (4) : 795-808.
Honrado J., Alves P., Lomba Â., Torres J. & Barreto Caldas F., 2007. Ecology, diversity and conservation of relict laurel-leaved mesophytic scrublands in mainland Portugal. Acta Botanica Gallica 154 (1) : 63-77.
Horvat I., Glavac V. & Ellenberg H., 1974. Vegetation Südosteuropas. Geobotanica Selecta IV : 1-767.
Julve Ph., 1993. Synopsis phytosociologique de la France (communautés de plantes vasculaires). Lejeunia, NS, 140 : 1-160.
Julve Ph. & Bouteloup R., 2018. Contribution à l’étude phytosociologique synusiale du massif des Albères. Bulletin de la Société de botanique du nord de la France 71 (1-4) : 191-234.
Kaya Ö.F., Cansaran A. & Yıldırım C., 2010. A syntaxonomical investigation of forest and pseudomaquis on transitional area in the central Black Sea region (Amasya, Turkey). Acta Botanica Gallica 157 (3) : 469-482.
Khelifi H. & Sadki N., 1995. Esquisse géobotanique des régions de Collo, Skikda et Annaba (Nord-Est algérien). Colloques phytosociologiques XXIII, Large area vegetation surveys : 593-615.
Korkmaz H., Yalçın E., Güray Kutbay H., Berk E. & Bilgin A., 2008. Contribution to the knowledge of the syntaxonomy and ecology of macchie and forest vegetation in Paphlagonia, North Anatolia, Turkey. Acta Botanica Gallica 155 (4) : 495-512.
Korotkov K., 1995. Caucasus dark coniferous forest phytosociology. Documents phytosociologiques, NS, XV : 273-297.
Krause W., Ludwig W. & Seidel F., 1963. Zur Kenntnis der Flora und Vegetation auf Serpentinstandorten des Balkans; 6 – Vegetationsstudien in der Umgebung von Mantoudi (Euböa). Botanische Jahrbücher 82 (4) : 337-403.
Kurt L., Ketenoğlu A.O., Akman Y., Özdeniz E., Şekerciler F., Bölükbaşi A. & Özbey B.G., 2015. Syntaxonomic analysis of the preforest and forest vegetation in the thermo- and eu-Mediterranean zone around Antalya Gulf, Turkey. Turkish Journal of Botany 39 : 487-498.
Lapraz G., 1974. Les vestiges de l’Oleo-Ceratonion du cap Ferrat. Riviera scientifique (4) : 81-94.
Lapraz G., 1977. Note sur l’Oleo-Lentiscetum du littoral de la Côte d’Azur entre Nice et Menton. Riviera scientifique (2) : 29-46.
Lavagne A., 1972. La végétation de l’île de Port-Cros, notice explicative au 1/5000 du Parc national. Parc national de Port-Cros et Université de Provence Marseille Saint-Charles, 31 p. et une carte.
Loidi J., Herrera M., Olano J.M. & Silván F., 1994. Maquis vegetation in the eastern Cantabrian coastal fringe. Journal of Vegetation Science 5 (4) : 533-540.
López Vélez G., 1996. Flora y vegetación del macizo del Calar del Mundo y sierras adyacentes del sur de Albacete. Instituto de estudios albacetenses de la Exma, Diputación de Albacete, 520 p.
Meddour R., 2010. Bioclimatologie, phytogéographie et phytosociologie en Algérie : exemple des groupements forestierset préforestiers de la Kabylie djurdjuréenne. Thèse de doctorat d’État, Université Mouloud Mammeri, Tizi Ouzou, 397 p. et annexes.
Medjadhi B. & Letreuch-Belarouci A., 2017. Typologie phytosociologique des formations forestières et préforestières des monts des Trara (Ouest algérien). Documents phytosociologiques, 3e série, 6 : 132-171.
Mesquita S., Arsénio P., Lousã M., Monteiro Henriques T. & Costa J.C., 2005. Sintra vegetation and landscape. Quercetea 7 : 65-81.
Miara M.D., Hadjadj Aoul S. & Ait Hammou M., 2012. Analyse phytoécologique et syntaxonomique des groupements végétaux dans le massif de Guezoul-Tiaret (NO Algérie). Bulletin de la Société botanique du Centre-Ouest, NS, 43 : 279-316.
Miara M.D., Hadjadj Aoul S. & Decocq G., 2020. Diversité phytosociologique et séries dynamiques de la végétation de l’Atlas tellien occidental de Tiaret (N-O Algérie). Ecologia Mediterranea 46 (1) : 97-116.
Molero Mesa J. & Pérez Raya F., 1987. Estudio fitosociológico de los sabinares de Juniperus phoenicea L. en el sector Malacitano-Almijarense (provincia corológica Bética). Lazaroa 7 : 301-306.
Molinier R., 1937. Les îles d’Hyères, étude phytosociologique. Annales de la Société d’histoire naturelle de Toulon 21 : 91-129.
Molinier R., 1954. Les climax côtiers de la Méditerranée occidentale. Vegetatio 4 : 284-308.
Nègre R. (dir.), 1964. Carte de la végétation de l’Algérie au 1/50 000e, feuille de Tipasa. Institut de cartographie de la végétation d’Algérie, Alger.
Nieto-Caldera J.M., Pérez-Latorre A.V. & Cabezudo B., 1990. Datos sobre la vegetación silicicola de Andalucia, I. Acta Botanica Malacitana 15 : 179-192.
Noble V. & Baret J., 2021. Catalogue des végétations du département des Bouches-du-Rhône. Conservatoire botanique national méditerranéen, Porquerolles, à paraître.
Paradis G. & Piazza C., 2001. Euphorbia dendroides en Corse : chorologie et phytosociologie. Bulletin de la Société botanique du Centre-Ouest, NS, 32 : 147-178.
Paradis G., Hugot L. & Delage A., 2005. Compléments à la connaissance de la répartition et de la phytosociologie d’Euphorbia dendroides en Corse. Bulletin de la Société botanique du Centre-Ouest, NS, 36 : 83-94.
Paradis G. & Pozzo di Borgo M.-L., 2005. Étude phytosociologique et inventaire floristique de la réserve naturelle des Tre Padule de Suartone (Corse). Journal de botanique de la Société botanique de France 30 : 27-103.
Paradis G., Féral C., Passigny-Hernandez C., Nicolau J. & Carles S., 2014. Paysage végétal de la vallée du Baraci et du nord du golfe de Valinco (sud-ouest de la Corse) : phytosociologie et carte de la végétation. Journal de botanique de la Société botanique de France 67 : 9-105.
Pavón Núñez M., Hidalgo Triana N. & Pérez Latorre A.V., 2013. Aportaciones al conocimiento de las comunidades de Pistacia terebinthus L. y de Acer monspessulanum L. en el sur de la Península Ibérica. Lagascalia 33 : 299-311.
Pérez Latorre A.V., Navas P., Navas D., Gil Y. & Cabezudo B., 1998. Datos sobre la flora y vegetación de la Serranía de Ronda (Málaga, España). Acta Botanica Malacitana 23 : 149-191.
Pérez Latorre A.V., Galán de Mera A. & Cabezudo B., 2000. La vegetación caracterizada por Rhododendron ponticum L. en Andalucía (España). Una complicada historia nomenclatural para una realidad fitocenológica. Acta Botanica Malacitana 25 : 198-205.
Pérez Latorre A.V., Casimiro Soriguer-Solanas F. & Cabezudo B., 2015. Flora y vegetación de la Sierra de Alcaparaín (Málaga, España). Acta Botanica Malacitana 40 : 107-156.
Pinto-Gomes C.J., Ladero Alvarez M., Gonçalves P.C., Mendes S. & Carmo Lopes M., 2003. Smilaco asperae-Quercetum suberis: um novo sobreiral réliquo do Alto Tejo. Quercetea 4 : 23-29.
Pinto Gomes C.J. & Paiva Ferreira R., 2005. Flora e vegetação do Barrocal Algarvia (Tavira, Portimão). Comissão de Coordenação e Desenvolvimento regional de Algarve, 354 p.
Quézel P., Barbero M. & Akman Y., 1978. L’interprétation phytosociologique des groupements forestiers dans le bassin méditerranéen oriental. Documents phytosociologiques, NS, II : 329-352.
Quézel P. & Barbero M., 1980. La végétation forestière de Crète. Ecologia Mediterranea 5 : 175-210.
Quézel P. & Barbero M., 1981. Contribution à l’étude des formations présteppiques à Juniperus arborescents du Maroc. Boletim da Sociedade Broteriana, série 2, 53 (2) : 1137-1160.
Quézel P., Barbero M. & Benabid A., 1987. Contribution à l’étude des groupements forestiers et préforestiers du Haut Atlas oriental (Maroc). Ecologia Mediterranea 13 (1-2) : 107-117.
Quézel P., Barbero M., Benabid A., Loisel R. & Rivas-Martínez S., 1988. Contribution à l’étude des groupements préforestiers et des matorrals rifains. Ecologia Mediterranea 14 : 77-122.
Quézel P., Barbero M. & Akman Y., 1992a. Typification de syntaxa décrits en région méditerranéenne orientale. Ecologia Mediterranea 18 : 81-87.
Quézel P., Barbero M., Benabid A. & Rivas-Martínez S., 1992b. Contribution à l’étude des groupements forestiers et préforestiers du Maroc oriental. Stvdia Botanica 10 : 57-90.
Quézel P., Barbero M., Benabid A. & Rivas-Martínez S., 1994. Le passage de la végétation méditerranéenne à la végétation saharienne sur le revers méridional du Haut Atlas oriental (Maroc). Phytocoenologia 22 (4) : 537-582.
Quinto-Canas R., Vila-Viçosa C., Meireles C., Paiva Ferreira R., Martínez-Lombardo M., Cano A. & Pinto-Gomes C., 2010. A contribute to the knowledge of the climatophilous cork-oak woodlands from Iberian southwest. Acta Botanica Gallica 157 (4) : 627-637.
Raposo M., Pinto Gomes C. & Lazare J.-J., 2021. Phytosociologocal analysis of the communities of Prunus lusitanica in France. Acta Botanica Croatica 80, à paraître.
Reymann J., Panaïotis Ch. & Bioret F. (coord.), 2016 (2017). Prodrome des végétations de Corse. Documents phytosociologiques, 3e série, 4 : 1-175.
Rivas Goday S., 1964. Vegetación y flórula de la cuenca extremeña del Guadiana. Publ. Diputac. Provinc. Badajoz, Badajoz, 777 p.
Rivas Goday S., Borja J., Esteve F., Galiano E.F., Rigual A. & Rivas-Martínez S., 1959 (publié 1960). Contribución al estudio de la Quercetea ilicis hispánica. Conexión de las comunidades hispánicas con Quercus lusitanica s.l. y sus correlaciones con las alianzas de Quercetalia ilicis, Quercetalia pubescentis y Quercetalia robori-petraeae. Anales del Instituto Botánico A.J. Cavanilles 17 (2) : 285-406.
Rivas-Martínez S., 1964 (publié 1965). Esquema de la vegetación potencial y su correspondencia con los suelos de la España peninsular. Anales del Instituto Botánico A.J. Cavanilles 22 : 343-404.
Rivas-Martínez S., 1969. Vegetatio Hispaniae, Notula I. Publicacciones del Instituto de Biologia Aplicada XLVI : 5-34.
Rivas-Martínez S., 1975. La vegetación de la clase Quercetea ilicis en España y Portugal. Anales del Instituto Botánico A.J. Cavanilles 31 (2) : 205-259.
Rivas-Martínez S., 1987. Mapa de series de vegetación de España. ICONA, serie Técnica, 268 p. + 30 mapas.
Rivas-Martínez S. & Rivas Goday S., 1975. Schéma syntaxonomique de la classe Quercetea ilicis dans la péninsule Ibérique. Colloques internationaux du CNRS 235 (La flore du bassin méditerranéen : essai de systématique synthétique) : 431-445.
Rivas-Martínez S., Costa M. & Izco J., 1986. Sintaxonomía de la clase Quercetea ilicis en el Mediterráneo Occidental. Notiziario delle Società Italiana di Fitosociologia19 (2) : 71-98.
Rivas-Martínez S., Lousã M., Díaz T.E., Fernández-González F. & Costa J.C., 1990. La vegetación del sur de Portugal (Sado, Alentejo y Algarve). Itinera Geobotanica 3 : 5-126.
Rivas-Martínez S., Costa M., Soriano P., Pérez Badia M.R., Llorens L. & Roselló J., 1992. Datos sobre el paisaje vegetal de Mallorca e Ibiza (Islas Baleares, España). Itinera Geobotanica 6 : 5-98.
Rivas-Martínez S., Fernández-González F. & Loidi J., 1999. Checklist of plant communities of Iberian Peninsula, Balearic and Canary Islands to suballiance level. Itinera Geobotanica 13 : 353-451.
Rivas-Martínez S., Fernández-González F., Loidi J., Lousã M. & Penas A., 2001. Syntaxonomical checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica 14 : 5-341.
Rivas-Martínez S. & Sánchez-Mata D., 2001. Precisiones sintaxonómicas sobre las alisedas aljíbicas. Lazaroa 21 : 150-152.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal. Addenda to the syntaxonomical checklist of 2001. Itinera Geobotanica 15 : 5-432.
Rodríguez Guitián M.A., Romero Franco R. & Ramil Rego P., 2007. Caracterización ecológica y florística de las comunidades lauroides del occidente de la Cornisa Cantábrica (Noroeste ibérico). Lazaroa 28 : 35-65.
Roselló R., 1994. Catálogo florístico y vegetación de la comarca natural del Alto Mijares (Castellón). Publ. Diputac. Castellón, Castellón, 650 p.
Rothmaler W., 1954. Vegetationsstudien im Nordwestspanien. Vegetatio 5-6 : 595-601.
Sadki N., 1995. Étude des groupements à Olivier et Lentisque de la région d’Annaba (Nord-Est algérien). Essai phytosociologique. Documents phytosociologiques, NS, XV : 253-271.
Siab-Farsi B., Khelifi H. & Kadid Y. 2014. Les maquis littoraux de la région algéroise : syntaxonomie et phytodiversité. Documents phytosociologiques, 3e série, 1 : 418-425.
Sivim, Sistema de Información de la Vegetación Ibérica y Macaronésica (www.sivim.info/sivi/).
Spichiger R.-E., Savolainen V., Figeat M. & Jeanmonod D., 2002. Botanique systématique des plantes à fleurs ; une approche phylogénétique nouvelle des Angiospermes des régions tempérées et tropicales. Presses polytechniques et universitaires romandes, Lausanne, 413 p.
Stephan J.M., Barhoun J., Maatouk E., de Foucault B. & Safi S., 2019. Syntaxonomy of open oak forests on basalt formations in North Lebanon. Lebanese Science Journal 20 (3) : 391-412.
Theocharopoulos M., Pantera A., Fotiadis G. & Papadopoulos A., 2020. Vegetation types with Quercus ithaburensis subsp. macrolepis in Greece. Ecologia Mediterranea 46 (1) : 17-40.
Theurillat J.-P., Willner W., Fernández-González F., Bültmann H., Čarni A., Gigante D., Mucina L. & Weber H., 2021. International Code of Phytosociological Nomenclature, 4th version. Applied Vegetation Science, https://doi.org/10.1111/avsc.12491.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Torres J.A., García-Fuentes A., Salazar C., Cano E. & Valle F., 1999. Caracterización de los pinares de Pinus halepensis Mill. en el sur de la Península Ibérica. Ecologia Mediterranea 25 (2) : 135-146.
Toubal O., 1998. Le Prunetum aviae, le Genistetum numidicae et le Chamaeropetum humilii, trois associations qui caractérisent la végétation numidienne (Edough, N.-E. Algérie). Itinera Geobotanica 11 : 213-226.
Toubal O. & Toubal A., 1996a. Diversité et dynamisme de la végétation de l’Edough (N.-E. Algérie), relation avec la lithologie. Colloques phytosociologiques XXIV, Fitodinamica : 585-599.
Toubal O. & Toubal A., 1996b. L’Euphorbietum dendroidis, faciès rupicole de l’Oleo–Lentiscetum sur substrat rocheux du littoral, micachischistes et calcaire métamorphique (cap de Garde, O. Begrat, N.-E. Algérie). Colloques phytosociologiques XXIV, Fitodinamica : 757-769.
Trinajstić I., 1978. Razred Paliuretea Trin., class. nov. Vegetacijska karta SR Hrvatske 1 : 43.
Trinajstić I., 1984. Vegetacija sveze Oleo-Ceratonion Br.-Bl. u jadranskom primorju Jugoslavije. Acta Botanica Croatica 43 : 167-173 (en croate).
Vanden Berghen C., 1990. Observations sur la végétation du djebel Bou Korbous (Cap Bon, Tunisie septentrionale). Documents phytosociologiques, NS, XII : 123-136.
Wojterski T., 1985. Guide de l’excursion internationale de phytosociologique en Algérie du Nord. Institut national agronomique, El Harrach, 274 p.
Zohary M. & Orshan G., 1959. The maquis of Ceratonia siliqua in Israël. Vegetatio 8 (5-6) : 285-297.
Remerciements
Merci à la Bibliothèque botanique et phytosociologique du Conservatoire botanique national de Bailleul, à Carlos Neto (Instituto de Geografia e Ordenamento do Território, Lisboa, Portugal), à Jorge Capelo (Instituto Nacional de Investigação Agrária, Unidade Estratégica de Investigação e Serviços de Sistemas Agrários e Florestais e Sanidade Vegetal, Oeiras, Portugal), à Rachid Meddour (Université Mouloud Mammeri, Tizi Ouzou, Algérie) et à Vincent Gaudillat (cellule Habitats, UMS 2006 Patrimoine naturel, MNHN) pour l’aide à la recherche de publications.


