
Données phytosociologiques sur les forêts de la Malepère (Aude, France) et leurs lisières
Title
Phytosociological data on the Malepère forests (Aude, France) and their edges
Résumé
Cette contribution à une monographie phytosociologique des forêts de la Malepère (département de l’Aude) et de leurs lisières a notamment permis la description de six associations végétales nouvelles ou validées, ainsi que la définition d’une alliance originale au sein des Trifolio medii-Geranietea sanguinei, l’Inulo conyzae-Galion maritimi. Deux autres associations classiques sont nouvelles pour ce département, quoique hors Malepère.
Abstract
This contribution to a phytosociological monography of the forests of the Malepère (department of Aude, France) and their edges allowed the description of six new or validated plant associations, as well as the definition of an original alliance within the Trifolio medii-Geranietea sanguinei, the Inulo conyzae-Galion maritimi. Two other classic associations are new for this department, but out of Malepère.
1. Introduction
À la suite à de diverses études phytosociologiques menées sur la végétation de la Malepère (de Foucault, 2015a, 2017a, c, 2019a, b, ponctuellement 2020b), on aborde ici l’étude des forêts locales et de leurs lisières. Des publications antérieures sur cette thématique sont disponibles, mais se réduisent souvent à des guides de stations forestières et à des investigations plutôt phytoécologiques (Alquié & Rubio, 2010 ; Lecomte & Moundy, 2011 ; Chabalier & Lecomte, 2013). Il est temps d’entreprendre des études plus strictement phytosociologiques. Selon Alquié & Rubio (2010), 70 % de la surface de la Malepère sont réservés à la forêt (photo 1).

Avant d’entrer dans les détails des résultats obtenus, la considération globale des boisements de la Malepère fait apparaître deux ensembles :
- l’ensemble de la Malepère orientale, sur la rive gauche de l’Aude, avec des bois pas toujours nommés s’étendant sur les communes de Roullens, Preixan, Rouffiac-d’Aude, Cépie, Saint-Martin-de-Villereglan ; l’altitude y est assez faible (souvent en-dessous de 350 m) et l’influence méditerranéenne plus marquée ; les sols paraissent plus basiques que ceux de l’ensemble suivant ;

- l’ensemble de la Malepère centro-occidentale, avec le bois de Caux (Villarzel-du-Razès), le bois de Las Mounjos (Montréal), le bois du Chapitre (Montréal et Arzens)… ; l’altitude y est plus élevée (souvent au-dessus de 300 m, avec au nord du bois de Las Mounjos le mont Naut, point culminant du massif à 442 m ; photo 2) et l’influence atlantique plus marquée ; les sols y paraissent un peu plus acides ; Fagus sylvatica y fait son apparition (col de la Malepère, bois du Chapitre au Saut Cani), ainsi que Carpinus betulus au bois de Las Mounjos.
2. Méthodes et conventions
La méthodologie d’étude des forêts a suivi le paradigme synusial (Gillet et al., 1991 ; de Foucault, 2020b). Chaque relevé est replacé dans l’espace (toponymie, latitude et longitude en ° sexagésimaux, altitude) et le temps (date du relevé, de 2013 à 2020) ; sauf exception, le recouvrement et la surface parcourue ont été notés. Les informations toponymiques et temporelles sont placées en annexe, après la bibliographie. Les relevés ont été classés en quinze tableaux faisant apparaître le nombre total de taxons par relevé, le nombre moyen de taxons par relevé d’un syntaxon élémentaire (Sy E), le nombre d’occurrences des taxons par Sy E. Pour certaines associations, on dressera aussi des cartes chorologiques au niveau communal (fond cartographique communal et cantonnal du Conseil départemental de l’Aude).
La nomenclature des taxons suit Flora Gallica (Tison & de Foucault, 2014). Les noms subspécifiques des taxons autonymes sont réduits à leur initiale pour gagner de la place (exemple : Rubia peregrina subsp. p. au lieu de Rubia peregrina subsp. peregrina) ; pour la même raison, dans les tableaux, le signe * remplace ‘subsp.’ ou ‘var.’. Le signe ‘j’ qualifie un taxon arborescent ou arbustif représenté par des individus juvéniles dans les synusies inférieures, alors que le signe ! désigne un taxon particulièrement exubérant par rapport à sa norme habituelle.
3. La végétation forestière
3.1. Les associations arborescentes
Seulement dix relevés de l’association arborescente la plus fréquente ont été rassemblés (tableau 1) tant elle est homogène sur une grande partie de la Malepère. Il s’agit essentiellement d’une chênaie mixte à Quercus ilex et Q. pubescens ; potentiellement, on devrait aussi y trouver Sorbus domestica, S. torminalis, Prunus avium présents à l’état juvénile dans les strates intraforestières arbustives et herbacées. Elle correspond au Quercetum pubescenti-ilicis (Braun-Blanq. 1936) B. Foucault 2020 (de Foucault, 2020b ; photo 3).

Il existe aussi un boisement plus pionnier où Pinus halepensis, essence héliophile, joue un rôle physionomique important ; il s’agit alors du Pino halepensis-Quercetum ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020 (de Foucault, 2020b ; photo 4).
Au bois du Chapitre (Montréal et Arzens), l’association arborescente est un peu différente, étant surtout constituée de Quercus petraea subsp. p., qui domine Q. ilex, ces deux essences étant accompagnées de Sorbus torminalis, S. domestica, Acer campestre, A. monspessulanum, Fraxinus excelsior, Tilia cordata. Provisoirement, on peut l’interpréter comme une variation à Quercus petraea subsp. p. du Quercetum pubescenti-ilicis.

3.2 Les associations arbustives intraforestières
La végétation arbustive intraforestière la plus fréquemment rencontrée (quatorze relevés, tableau 2) associe Ligustrum vulgare, Viburnum tinus, V. lantana, Arbutus unedo, Rubia peregrina subsp. p., Dioscorea communis, plus rarement Rosa sempervirens ; un lot important de formes juvéniles de taxons potentiellement arborescents s’y associe, surtout Quercus ilex, Sorbus domestica, S. torminalis. Elle se distingue du Roso sempervirentis-Viburnetum tini essentiellement par la rareté ou l’absence de taxons plus héliophiles, dont Lonicera etrusca. Il ne paraît donc pas utile de définir une association originale distincte. On y observe deux variations, l’une typique différenciée par Cornus sanguinea subsp. s., Viburnum lantana, Prunus spinosa, Juniperus communis subsp. c., l’autre différenciée surtout par Arbutus unedo et Erica arborea, semble-t-il plus acidiphile que le Roso-Viburnetum tini arbutetosum unedonis (de Foucault, 2015a).

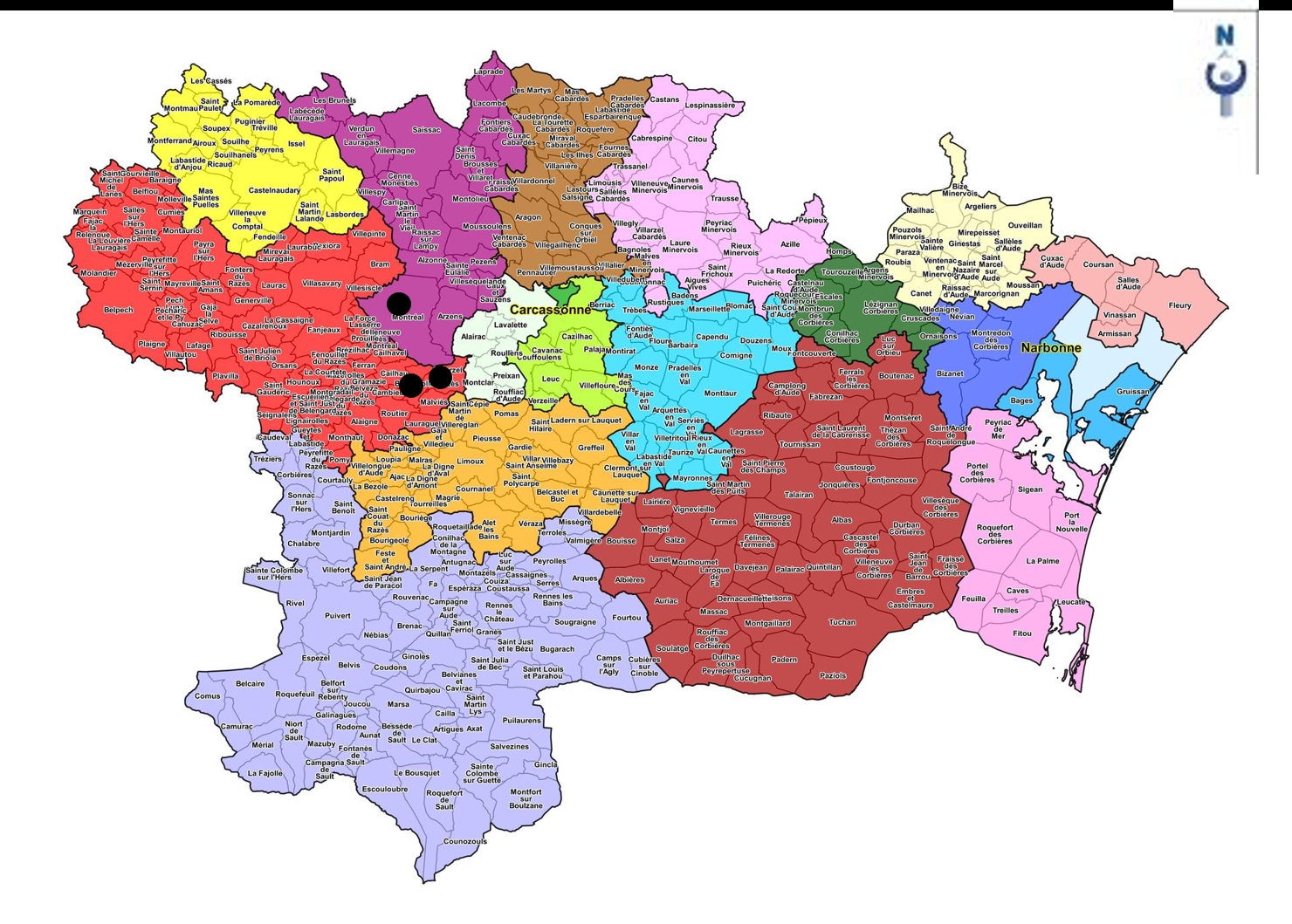
La végétation arbustive intraforestière des bois de la Malepère centro-occidentale est par contre bien distincte de la précédente. Le tableau 3 en rassemble quatorze relevés dispersés, qui combinent essentiellement Ilex aquifolium, Crataegus germanica, C. laevigata, Lonicera periclymenum subsp. p. ; divers arbres y sont représentés à l’état arbustif, surtout Quercus petraea subsp. p., Q. ilex, Prunus avium et Sorbus torminalis. Cette association est assez remarquable, notamment parce qu’elle confirme pleinement l’alliance du Mespilo germanicae-Ilicion aquifolii B. Foucault & Julve ex B. Foucault & J.-M. Royer in B. Foucault 2019. Dans cette unité, ne sont connus que quatre syntaxons, dont trois associations, le Mespilo germanicae-Ilicetum aquifolii B. Foucault 1995, l’Ilici aquifolii-Prunetum spinosae Delelis in B. Foucault & J.-M. Royer 2016 et le Rubo gillotii-Prunetum spinosae J.-M. Royer in B. Foucault & J.-M. Royer 2016, tous les trois plutôt psychrophiles et dépourvus de taxons thermophiles rencontrés ici, comme Rubia peregrina subsp. p., Viburnum tinus, Sorbus torminalis j, Quercus ilex j. Il s’agit donc d’une association très originale qui peut être dénommée Sorbo torminalis-Crataegetum germanicae ass. nov. hoc loco, typus nominis hoc loco : relevé 5 du tableau 3 hoc loco (photo 5). Cette association à Crataegus germanica, Ilex aquifolium, Crataegus laevigata, Lonicera periclymenum subsp. p., Rubia peregrina subsp. p., Dioscorea communis, de jeunes Quercus ilex, Q. pubescens, Sorbus torminalis… accueille en moyenne près de 15 taxons par relevé. Cette association semble exister plus à l’ouest, dans le bois de Pique-Mourre (communes de Cazalrenoux et La Cassaigne ; de Foucault, 2020b : § III, alliance CR). La carte 1 précise la dispersion connue du Sorbo-Crataegetum germanicae (sauf bois de Pique-Mourre, à confirmer).
Le tableau 4 dresse une nouvelle synthèse de cette alliance, avec les cinq syntaxons suivants :
- IPs. Ilici aquifolii-Prunetum spinosae Delelis in Foucault & J.-M. Royer 2016 (d’après de Foucault & Royer, 2016, tableau 4 : colonne 89).
- HM. haie du Morvan (d’après de Foucault & Royer, 2016, tableau 4 : colonne 91).
- RPs. Rubo gillotii-Prunetum spinosae -M. Royer in B. Foucault & J.-M. Royer 2016 (d’après de Foucault & Royer, 2016, tableau 4 : colonne 90).
- MIa. Mespilo germanicae-Ilicetum aquifolii Foucault 1995 ; 1 – typicum (de Foucault, 1995, tableau 28 : colonne A) ; 2 – franguletosum alni B. Foucault 1995 (de Foucault, 1995, tableau 28 : colonne B).
- SCg. Sorbo torminalis-Crataegetum germanicae (le tableau 3 hoc loco).
3.3 Les associations herbacées intraforestières
Au niveau herbacé intraforestier, on a reconnu deux associations ; le tableau synthétique 7 en facilite la comparaison, montrant la présence commune à ces deux syntaxons de Ruscus aculeatus, Daphne laureola, Brachypodium sylvaticum, Euphorbia amygdaloides subsp. a., Carex flacca subsp. f., Hedera helix.

La première association est définie à partir des 38 relevés du tableau 5. En dehors des formes juvéniles de taxons ligneux et de quelques taxons compagnes, elle est surtout caractérisée par la combinaison de Rubia peregrina subsp. p., Daphne laureola, Ruscus aculeatus, Dioscorea communis, Carex flacca subsp. f., C. halleriana, Cephalanthera longifolia ; quelques autres orchidées sont irrégulièrement présentes : Orchis purpurea, Cephalanthera damasonium, Limodorum abortivum, Ophrys insectifera. Il semble que les géophytes, dont justement ces orchidées, soit activement recherchés par les sangliers, qui contribueraient donc à appauvrir cette végétation. Quoi qu’il en soit, on est en présence ici d’une association méconnue et originale, le Cephalanthero longifoliae-Daphnetum laureolae ass. nov. hoc loco, typus nominis hoc loco : relevé 8 du tableau 5 hoc loco (photo 6). Il s’agit d’une association plutôt basiphile, qui paraît pouvoir se ranger dans l’alliance du Dioscoreo communis-Mercurialion perennis (de Foucault, 2020b). Le tableau 5 met en évidence deux variations principales (en dehors des relevés 1 et 2 qui ne se rattachent à aucune des deux) :
- typicum (relevés 3 à 35), incluant le type nomenclatural, différencié par les taxons plutôt basiphiles comme Carex halleriana, Helleborus foetidus, Viburnum lantana j, Hippocrepis emerus e. j, Orchis purpurea, Epipactis microphylla, Cephalanthera damasonium, Limodorum abortivum…, accueillant en moyenne 18,2 taxons par relevé ;
- luzuletosum forsteri subass. nov. hoc loco (relevés 36 à 38), plus acidiphile, différencié surtout par Luzula forsteri, Erica arborea j, Teucrium scorodonia, accueillant en moyenne 22,7 taxons par relevé, typus nominis hoc loco: relevé 36 du tableau 5 hoc loco.
Cette seconde variation, et tout particulièrement le relevé 38, annonce la lande à Calluna vulgaris, Cistus salviifolius, Erica arborea j qui sera décrite plus loin.
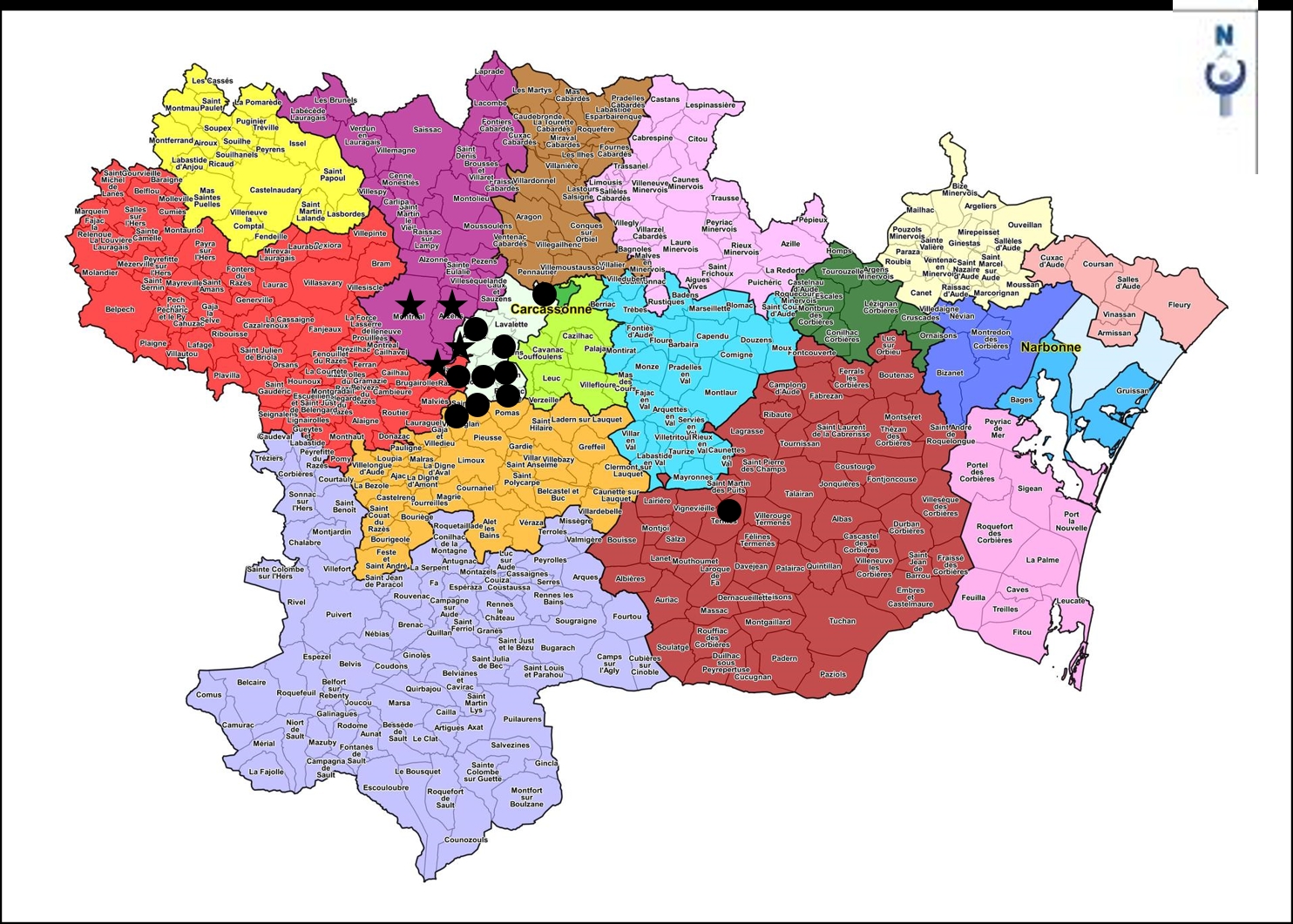
En Malepère, cette association est surtout répartie dans sa partie orientale ; elle a été incidemment rencontrée aussi hors Malepère, plus à l’est, lors d’une sortie de la SESA au printemps 2019 (de Foucault, 2020a). La carte 2 (cercles noirs) donne une idée de sa répartition au niveau communal (incluant la station de Termes du printemps 2019).
La seconde association caractérise plutôt la Malepère centro-occidentale ; le tableau 6 en rapporte 17 relevés. Ce syntaxon combine essentiellement Lonicera periclymenum subsp. p., Daphne laureola, Ruscus aculeatus, Dioscorea communis, Rubia peregrina subsp. p., Luzula forsteri, Melica uniflora, Teucrium scorodonia ; il est donc bien distinct du Cephalanthero-Daphnetum laureolae précédent (tableau 7) et peut être décrit sous le nom de Melico uniflorae-Ruscetum aculeati ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 6 hoc loco. Il accueille en moyenne environ 20 taxons par relevé. La carte 2 (étoiles noires) dresse l’état de sa dispersion communale connue, les communes de Montclar et Villarzel-du-Razès assurant le relais d’une association à l’autre.
En dépit de l’absence du premier taxon éponyme de l’alliance, cette association possède des affinités floristiques certaines avec le Primulo vulgaris-Ruscion aculeati atlantique (de Foucault, 2020b) ; en témoigne notamment la position de Polystichum setiferum en situation plane.
Au sein de ces boisements à surface largement plane, on peut parfois observer des ruptures de pente ou des ravins (bois du Chapitre au Saut Cani, bois de Las Mounjos au ruisseau de la Combe, « Sabran »), les Cephalanthero longifoliae-Daphnetum laureolae et Melico uniflorae-Ruscetum aculeati sont alors souvent remplacés par l’Asplenio onopteridis-Polystichetum setiferi qui accueille Asplenium onopteris, A. scolopendrium, Polystichum setiferum (de Foucault, 2020b ; photo 7). Le relevé suivant est inédit :
Roullens, « Sabran », N 43° 09’ 22,4’’, E 2° 14’ 50’’, 294 m, 16/05/2020, 40 %, 15 m², 11 taxons
- Asplenium onopteris 2, Polystichum setiferum 2, Dioscorea communis 1, Ruscus aculeatus 1, Rubia peregrina *p. +,
- autres taxons : Hedera helix 2, Rubus 2, Symphytum tuberosum +, Brachypodium sylvaticum +, Acer campestre j +, Ligustrum vulgare j +.
La carte 3 en dresse la répartition connue, entre Malepère, Montagne noire et Corbières.
À titre informatif, on peut enfin présenter ici le relevé d’un sous-bois herbacé sur terrasse au bord du ruisseau dit de Bonnemère ou de Ganès selon les sources topographiques :
Alairac, vers « Ganès », N 43° 10’ 00,9’’, E 2° 14’ 14,6’’, 228 m, 18/05/2020, 80 %, 40 m², 27 taxons
- Pulmonaria affinis 2, Lamium galeobdolon *montanum 2, Melica uniflora 2, Carex flacca *f. 2, Rubia peregrina *p. 2, Euphorbia amygdaloides *a. 1, Viola reichenbachiana 1, Symphytum tuberosum 1, Brachypodium sylvaticum 1, Ruscus aculeatus +, Circaea lutetiana +, Arum italicum +, Iris foetidissima +, Daphne laureola +, Carex pendula +,
- autres taxons : Hedera helix 3, Bromopsis ramosa *r. 1, Smilax aspera +, Laurus nobilis j 1, Corylus avellana j 1, Ligustrum vulgare j 1,Cornus sanguinea *s. j +, Acer campestre j +, Viburnum lantana j + Prunus avium j +, Crataegus monogyna j +, Quercus ilex j +.

Il est original par la présence de P. affinis, de S. aspera rare en Malepère, de Symphytum tuberosum et autres taxons absents des niveaux topographiques supérieurs mieux drainés.
4. La végétation des lisières
4.1 Les lisières arbustives ou manteaux
Les lisières arbustives ont été peu abordées dans les présentes investigations. On rappellera simplement la définition en Malepère de deux associations, le Roso sempervirentis-Viburnetum tini, sur un substrat peu acide, et le Cytiso scoparii-Ericetum arboreae, sur un substrat acidifié (de Foucault, 2015a). Le tableau 8 rapporte huit nouveaux relevés de ce dernier provenant de la Malepère.
4.2 Les lisières herbacées ou ourlets
En dehors des ourlets thérophytiques vernaux (de Foucault, 2017a), qui peuvent entrer en mosaïque avec les suivants, les ourlets préforestiers vivaces ont rarement été abordés. Royer (2020) cite deux associations catalanes susceptibles d’atteindre notre région, le Galio maritimi-Origanetum vulgaris O. Bolòs 1967 et le Chrysanthemo corymbosi-Oryzopsietum paradoxae O. Bolòs 1978, qu’il range dans un groupe d’associations appauvries et marginales au sein du Trifolio medii-Geranienion sanguinei, du Geranion sanguinei, des Antherico ramosi-Geranietalia sanguinei et des Trifolio medii-Geranietea sanguinei.
Dans cette contribution, on présente d’abord deux syntaxons à Galium maritimum. Le premier est défini par les sept relevés du tableau 9 ; à part les nombreux taxons arbustifs et arbrisselés du Roso sempervirentis-Viburnetum tini et quelques chaméphytes liés surtout au Koelerio macranthae-Thymetum vulgaris (de Foucault, 2019a), cet ourlet associe Silene italica, Galium maritimum, Rubia peregrina subsp. p., Origanum vulgare subsp. v., Stachys recta subsp. r., Inula conyzae, Clinopodium vulgare. On en restera ici au niveau d’un groupement à Thymus vulgaris subsp. v. – Silene italica.

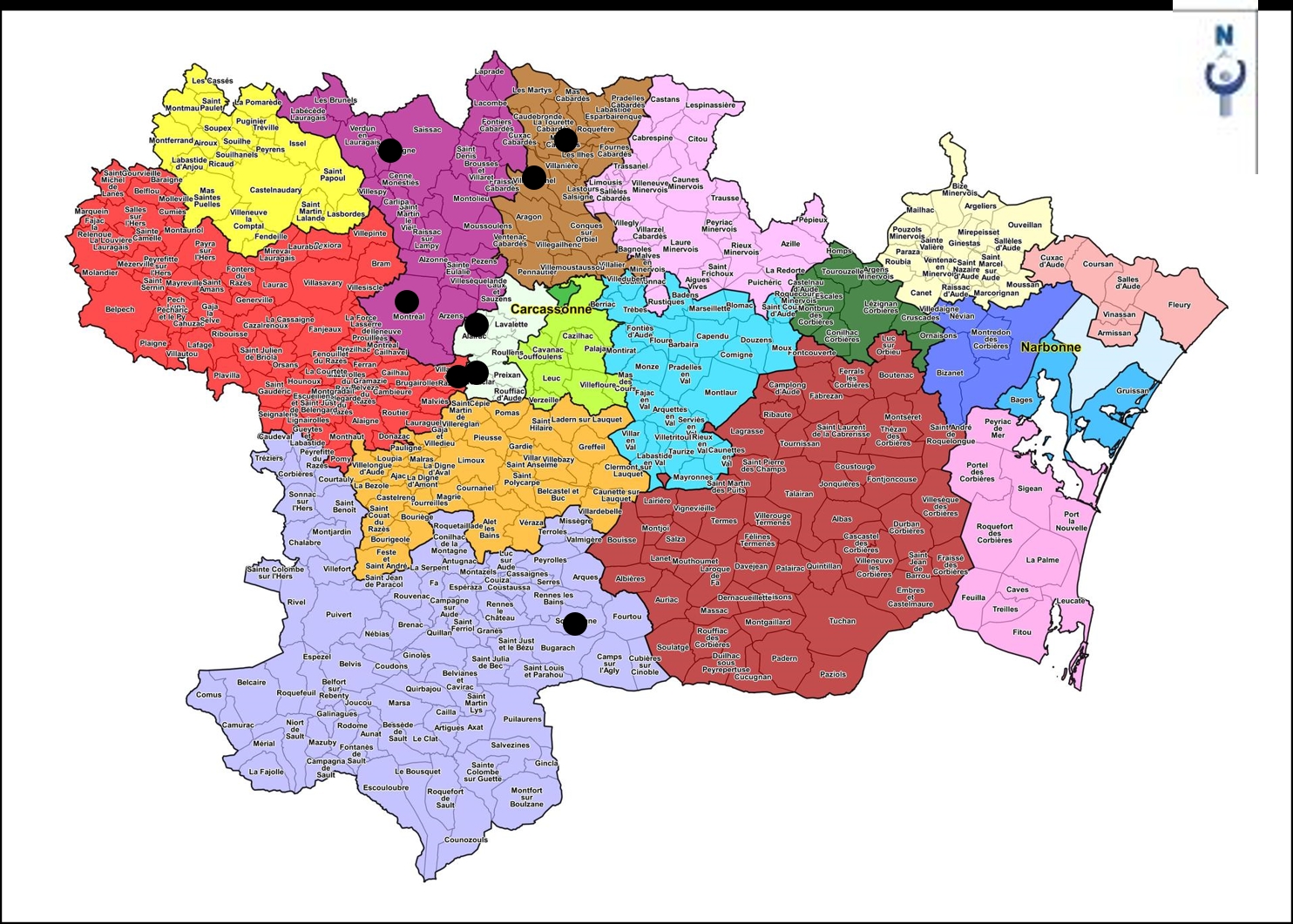
Le second syntaxon est mieux caractérisé, sur la base de dix-neuf relevés/listes (tableau 10) qui partagent Galium maritimum, Euphorbia amygdaloides subsp. a., Silene italica, Campanula rapunculus, Urospermum dalechampii, Epipactis microphylla, Daphne laureola, Rosa sempervirens, Dioscorea communis, Lathyrus latifolius, Inula conyzae, Lonicera etrusca… ; il s’agit d’un syntaxon bien distinct du Galio-Origanetum vulgaris (voir aussi pour comparaison le tableau 11) et défini ici comme Inulo conyzae-Galietum maritimi ass. nov. hoc loco, typus nominis hoc loco : relevé 7 du tableau 10 hoc loco (photo 8). Cet ourlet est assez riche floristiquement puisqu’il accueille en moyenne 35 taxons par relevé (sur 17 relevés, les listes n’étant évidemment pas prises en compte), dont souvent un tiers de taxons ligneux (arbrisseaux, arbustes, arbres) juvéniles. Il constitue le plus souvent l’ourlet de la chênaie verte à Cephalanthero longifoliae-Daphnetum laureolae. La carte 4 en dresse la répartition communale connue (cercles).
Le tableau 11 tente une nouvelle synthèse du Trifolio medii-Geranienion sanguinei (d’après le tableau 3 de Royer, à paraître, avec l’accord de l’auteur, légèrement modifié : ordre des taxons, nomenclature en accord avec Flora Gallica, colonne 40 compilée selon Sivim consulté le 14 juillet 2019, ajout de nos deux nouveaux syntaxons de la Malepère ; les numéros 28 à 41 sont ceux de la synthèse initiale) :
- Centaureo semidecurrentis-Melampyretum cristati Braque & Loiseau ex J.-M. Royer.
- Odontito chrysanthae–Phyteumatetum teneri Braque & Loiseau ex J.-M. Royer.
- Lithospermo purpurocaerulei–Pulmonarietum longifoliae B. Foucault 2008.
- Trifolio medii-Silaetum silai Braque ex J.-M. Royer.
- Pteroselini austriaci-Trifolietum alpestris van Gils & Keysers ex J.-M. Royer.
- Trifolio medii–Laserpitietum latifolii van Gils & Gilissen 1976 ; a – d’Autriche ; b – de Suisse ; c – de Savoie.
- Bupleuro falcati–Laserpitietum latifolii Billy ex J.-M. Royer.
- Coronillo variae–Vicietum tenuifoliae J.-M. Royer & Rameau 1983 ; a – d’Allemagne ; b – de Bourgogne et Champagne ; c – du Jura français.
- Campanulo bononiensis–Vicietum tenuifoliae Krausch ex T. Müll. 1962.
- Coronillo variae–Peucedanetum alsatici J.-M. Royer, Rameau & Prin 1983 ; a – de Champagne ; b – d’Auvergne.
- Clinopodio vulgaris–Laseretum trilobi Knapp ex T. Müll. in Oberd. 1978.
- Coronillo variae–Brachypodietum pinnati J.-M. Royer & Bidault ex J.-M. Royer 1973.
- Galio maritimi–Origanetum vulgaris O. Bolòs 1967.
- Chrysanthemo corymbosi–Oryzopsietum paradoxae O. Bolòs 1978.
- Groupement à Thymus vulgaris – Silene italica (tableau 9 hoc loco).
- Inulo conyzae-Galietum maritimi B. Foucault (tableau 10 hoc loco).
Ce tableau synthétique montre bien la grande coupure passant entre les colonnes 39 et 40. Il n’est pas tenable de réduire l’ensemble des quatre derniers syntaxons à un simple groupe appauvri et marginal du premier grand ensemble, donc du Trifolio medii-Geranienion sanguinei optimal. Il est clair qu’il faut définir une alliance originale d’ourlets basiphiles à chorologie nord-ouest-méditerranéenne, surtout en dessous de 800 m d’altitude, l’Inulo conyzae-Galion maritimi B. Foucault & J.-M. Royer all. nov. hoc loco, caractérisée ou différenciée des alliances voisines par Galium maritimum, Viola alba, Urospermum dalechampii, Silene italica, Cruciata glabra var. g., auxquels on peut ajouter Medicago hybrida ; typus nominis hoc loco : Inulo conyzae-Galietum maritimi B. Foucault. D’autres relevés d’ourlets audois mais hors Malepère se rapportent aussi à cette alliance : à Villardebelle (de Foucault, 2015b : 197), à Fontjoncouse (de Foucault, 2016 : 140), à la Coume de Félines (de Foucault, 2017b : 191), à Alet-les-Bains (de Foucault et al., 2017 : 196-7). À l’autorité de cette alliance, nous avons associé le nom de notre ami J.-M. Royer en hommage à son travail de synthèse sur la classe.
L’ourlet étudié maintenant et relevé plutôt en Malepère centro-occidentale est assez distinct des précédents et d’ailleurs paraît ne pas se rattacher à l’Inulo conyzae-Galion maritimi. Le tableau 12 en rapporte douze relevés qui partagent Lathyrus niger subsp. n., Prunella hastifolia, Serratula tinctoria, Silene nutans subsp. n., Conopodium majus subsp. m., Pulmonaria longifolia subsp. l., Teucrium scorodonia, Melittis melissophyllum subsp. m., Trifolium medium subsp. m., Oenanthe pimpinelloides… Il s’agit donc d’un ourlet très original, un peu plus acidiphile que l’Inulo conyzae-Galietum maritimi : Prunello hastifoliae-Lathyretum nigri ass. nov. hoc loco, typus nominis hoc loco : relevé 1 du tableau 12 hoc loco (photo 9). À côté du typicum, typifié par le type de l’association, différencié par Rumex acetosa subsp. a., Cardamine pratensis, Festuca heterophylla, Potentilla sterilis, Brachypodium sylvaticum…, accueillant en moyenne 35 taxons par relevé, il existe une sous-association différenciée par Anacamptis pyramidalis var. p., Clinopodium vulgare, Bromopsis erecta subsp. e., Asparagus acutifolius, Inula conyzae, Ruscus aculeatus, Urospermum dalechampii, Galium maritimum, un peu plus thermophile, mais connue par seulement cinq relevés. Chacune de ses deux variations majeures présente une variante plus acidiphile à Genista pilosa subsp. p., Calluna vulgaris, Hypericum pulchrum, Cistus salviifolius, Lathyrus linifolius, Hieracium virgultorum, Cytisus scoparius subsp. s. j, Erica arborea j. Ce bel ourlet, qui correspond aussi à celui incomplètement relevé à Alet-les-Bains (de Foucault et al., 2017 : 199) et dont la carte 4 (étoiles) dresse la chorologie communale connue, semble pouvoir être placé dans le Conopodio majoris–Teucrion scorodoniae au sein des Melampyro pratensis-Holcetea mollis, tendant vers le Teucrio scorodoniae-Trifolienion medii au sein des Trifolio-Geranietea sanguinei (Royer, à paraître). Il semble atteindre le massif gréseux de Rennes-les-Bains comme le montre cette observation rapide (en N 42° 54’ 03,4’’, E 2° 18’ 47,1’’, 338 m) de Lathyrus niger subsp. n., L. linifolius, Lonicera periclymenum subsp. p., Veronica officinalis, Solidago virgaurea subsp. v., Rubia peregrina subsp. p., Teucrium scorodonia, Trifolium medium subsp. m., Brachypodium sylvaticum, Potentilla erecta, Betonica officinalis subsp. o., Prunella hastifolia…

Les variantes acidiphiles des deux variations majeures du Prunello hastifoliae-Lathyretum nigri semblent annoncer un ourlet franchement acidiphile réellement rencontré lors de nos investigations au bois du Chapitre (N 43° 10’ 56,6’’, E 2° 10’ 21,2’’, 309 m, 11/07/2019, 27 taxons) :
- Lonicera periclymenum *p. 3, Avenella flexuosa *f. 3, Luzula sylvatica *s. 3, forsteri 1, Pteridium aquilinum 2, Hieracium virgultorum 2, Melampyrum pratense 1, Teucrium scorodonia 1, Hypericum pulchrum 1, Danthonia decumbens *d. +, Pilosella officinarum +, Solidago virgaurea *v. +,
- jeunes ligneux : Cytisus scoparius *s. j 2, Viburnum tinus j 1, Crataegus germanica j 1, Castanea sativa j 1, Sorbus torminalis j 1, Erica arborea j 1, Prunus avium j 1, Quercus ilex j 1, petraea *p. j +, Ilex aquifolium j +,
- autres taxons : Hedera helix 2, Calluna vulgaris 1, Vinca minor +, Centaurea decipiens +, Agrostis capillaris *c.
Cet ourlet se range bien aussi dans le Conopodio majoris-Teucrion scorodoniae et rappelle beaucoup l’Hyperico pulchri-Melampyretum pratensis, dont il constitue au moins une race subméditerranéenne, peut-être même une association originale vicariante.

4.3 Les landes
Les landes à Ericaceae ont souvent la valeur d’ourlets chaméphytiques ; leur étude peut donc logiquement suivre celle des ourlets herbacés vivaces. C’est aussi l’occasion de tenter ici plus largement la description des landes de la Montagne noire et régions voisines.
À cet effet, le tableau 13 rassemble 42 relevés. On constate que cette lande peut être caractérisée par la combinaison des taxons suivants : Erica cinerea, Cistus salviifolius, Calluna vulgaris, Genista pilosa subsp. p., Rubia peregrina subsp. p., ainsi que des taxons juvéniles annonçant le Cytiso scoparii-Ericetum arboreae : Erica scoparia subsp. s., E. arborea, Cytisus scoparius subsp. s., Arbutus unedo, Quercus ilex. Ces relevés se répartissent en deux syntaxons majeurs (laissant de côté les relevés 1 et 2 plus fragmentaires) :
- l’un (relevés 3 à 13) différencié par Lavandula stoechas subsp. s., Cistus albidus, monspeliensis, paraissant plus thermophile, souvent sur des substrats plus superficiels, accueillant de 13 à 14 taxons en moyenne par relevé ;
- l’autre (relevés 14 à 42), différencié par Lonicera periclymenum p., Rubus ulmifolius, Ulex europaeus subsp. e. j, Pteridium aquilinum, Genista anglica, Centaurea decipiens, Carex flacca subsp. f., C. pilulifera subsp. p., Avenella flexuosa subsp. f., Brachypodium rupestre subsp. r., Lathyrus linifolius, moins thermophile, souvent sur des substrats plus profonds, accueillant le même nombre moyen de taxons par relevé.
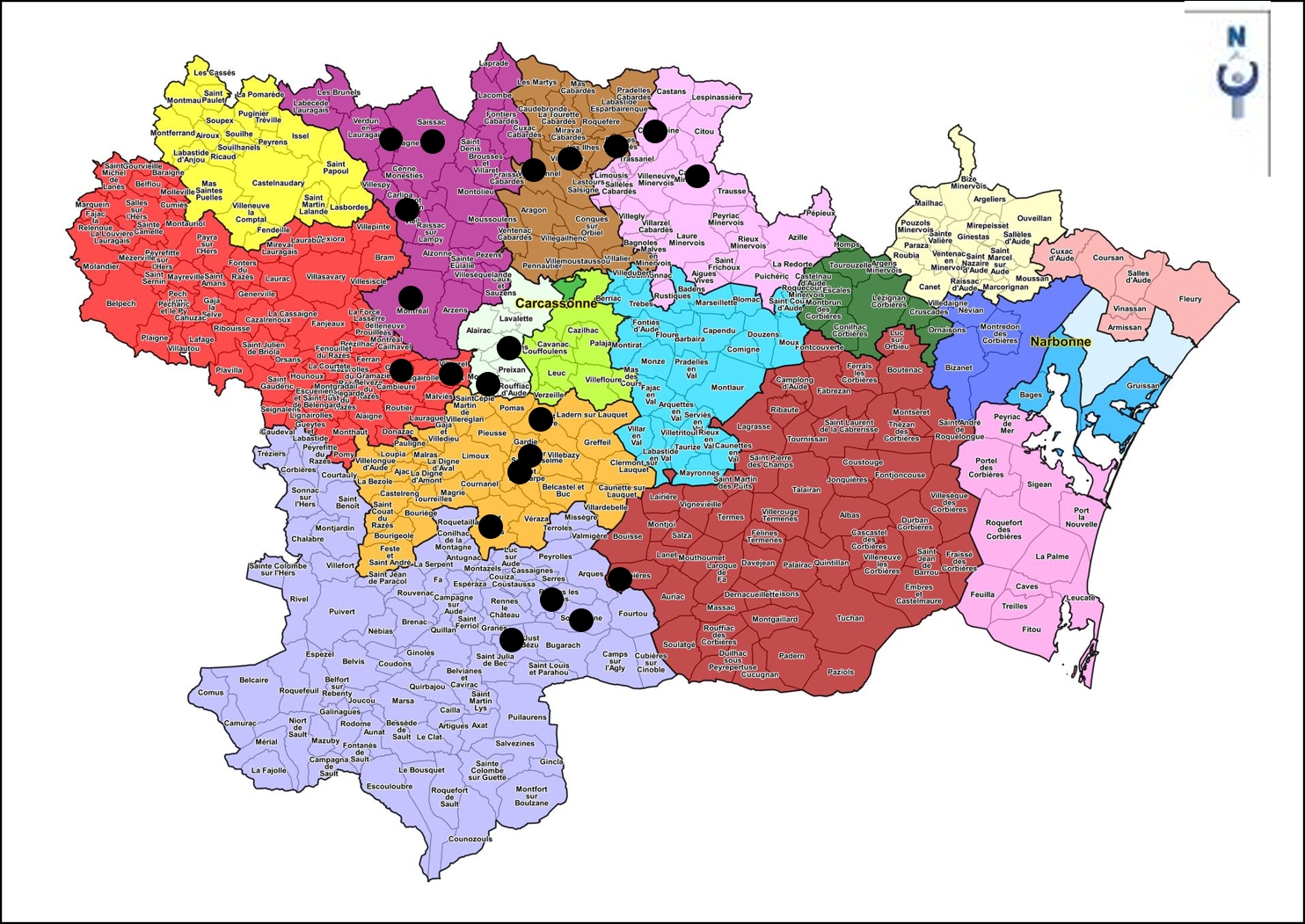
Cette combinaison de taxons place cette lande à la charnière chorologique entre Calluno-Ulicetea minoris et Cisto-Lavanduletea stoechadis ; son caractère modérément méditerranéen amène à la rattacher au Cisto salviifolii-Ericion cinereae Géhu ex Géhu in Bardat et al. 2004. Dans cette alliance sont actuellement décrites deux associations valides : le Festuco juncifoliae-Ericetum cinereae Géhu & Géhu-Franck 1975 (le nom de la fétuque est à corriger en Festuca vasconcensis ; Géhu & Géhu-Franck, 1975 ; type de l’alliance) et le Cisto salviifolii-Ericetum cinereae Guitton et al. 2018. Le Lavandulo stoechadis-Ericetum cinereae cité par Géhu (1975) de la Montagne noire et du Minervois siliceux est un nomen nudum invalide. Malgré la faible présence de Lavandula stoechas dans notre tableau 11, c’est bien à ce syntaxon qu’il convient de rattacher la lande étudiée ici : Lavandulo stoechadis-Ericetum cinereae Géhu ex B. Foucault ass. nov. hoc loco. On pourrait peut-être objecter que ce syntaxon pourrait être scindé en deux, la partie thermophile, surtout liée à la Montagne noire, et la partie moins thermophile à Lonicera periclymenum ; cependant la première existe hors Montagne noire et la seconde existe aussi en Montagne noire, ce sont donc plus des traits édapho-climatiques qui les distinguent. Afin de remplir la condition de l’article 3f de l’ICPN, la première des deux variations citées, dont L. stoechas est différentiel, doit livrer le typus nominis hoc loco : relevé 3 du tableau 13 hoc loco (photo 10). Il en résulte que la première variation correspond au typicum, typifié par le type de l’association, la seconde variation pouvant alors être décrite sous le nom de loniceretosum periclymeni subass. nov. hoc loco, typus nominis hoc loco : relevé 25 du tableau 13 hoc loco. Cette lande est dispersée sur la Montagne noire, le Cabardès, la Malepère acide, le Minervois acide, les Corbières (carte 5), entre 200 et 600 m d’altitude, avec une moyenne de 377 m ; incidemment, elle a aussi été relevée à Alet-les-Bains (de Foucault et al., 2017, tableau 1). Elle est reconnaissable encore parmi les données incomplètes provenant du Tarn (de Foucault, 1994 ; Julve & de Foucault, 1994). Enfin, elle est potentielle au domaine de Peyremale (commune de Montolieu) mais est actuellement réduite à Lavandula stoechas subsp. s., Cistus salviifolius et Erica scoparia subsp. s. j (de Foucault & Klesczewski, 2020). On comprendra alors qu’elle puisse appartenir à plusieurs systèmes et séries phytosociologiques, en lien avec des forêts potentielles et des pelouses variées.

À titre informatif, dans le Lauragais nord-oriental (notamment autour d’Issel, environ 230 m d’altitude ; tableau 14 : relevés 1 à 5), mais aussi à plus haute altitude dans les Corbières (tableau 14 : relevé 6, vers 630 m), il existe une lande moins thermophile dont le statut synsystématique reste pour l’instant indéterminé, tout en se rattachant bien au Cisto salviifolii-Ericion cinereae. Ce tableau 14 permet de la comparer au Lavandulo-Ericetum cinereae précédent (colonne LEc, issue du tableau 13, sauf relevés 1 et 2), qui en retour affine l’originalité de ce dernier.
Cette lande accueille parfois Pseudarrhenatherum longifolium, rare taxon patrimonial de l’Aude (Plassart et al., 2016) qui participe aussi à une pelouse acidiphile (Issel, carrefour RD 624 × RD 126, N 43° 22’ 47,4’’, E 1° 58’ 33,1’’, 216 m, 10 taxons) : Ps. longifolium 2, Danthonia decumbens *d. 2, Carex pilulifera *p. 2, C. flacca *f. 2, Serratula tinctoria 2, Lonicera periclymenum *p. 1, Rubia peregrina *p. 1, Brachypodium rupestre *r. 2, Solidago virgaurea *v. 1, Frangula alnus *a. j +. Ces lande et pelouse sont aussi en lien dynamique avec le fourré de l’Erico scopariae-Franguletum alni Géhu & Géhu-Franck 1975, dont trois relevés sont réunis dans le tableau 15 ; c’est la première mention de cette association dans notre département, qui en constitue une des limites orientales.
Dans ce système acide, il existe enfin une molinaie :
Issel, vers carrefour D 624 × D 624Z, sous ligne HT, N 43° 22’ 59,4’’, E 1° 58’ 35,6’’, 240 m, 20/09/2015. 100 %, 200 m², 15 taxons
- Molinia caerulea 3, Erica scoparia *s. j 3, Potentilla erecta +, Serratula tinctoria +, Juncus conglomeratus +, effusus +,
- autres taxons : Erica cinerea 2, Calluna vulgaris 2, Cistus salviifolius +, Carex flacca *f. +, Rubus ulmifolius +, Ulex europaeus *e. j 2, Populus tremula j 1, Quercus pubescens j 1, Frangula alnus *a. j 1.
Elle peut être rattachée à l’Erico scopariae-Molinietum caeruleae B. Foucault 2008, surtout bien développé dans les Landes de Gascogne (de Foucault, 1984).
5. Conclusion
Sur la base d’environ deux cents relevés répartis en une quinzaine de tableaux, cette étude des boisements de la Malepère et de leurs lisières a donc révélé cinq associations nouvelles : le Sorbo torminalis-Crataegetum germanicae, le Cephalanthero longifoliae-Daphnetum laureolae, le Melico uniflorae-Ruscetum aculeati, l’Inulo conyzae-Galietum maritimi et le Prunello hastifoliae-Lathyretum nigri ; elle a validé une association restée longtemps invalide faute de relevés précis, le Lavandulo stoechadis-Ericetum cinereae ; elle a étendu vers l’Aude, mais hors Malepère, l’aire de deux associations devenues classiques, l’Erico scopariae-Franguletum alni et l’Erico scopariae-Molinietum caeruleae. Enfin, elle a permis la définition d’une alliance nouvelle au sein des Trifolio medii-Geranietea sanguinei, l’Inulo conyzae-Galion maritimi. De nouvelles investigations dans les boisements des plaines et collines audoises devraient permettre d’étendre l’aire de ces associations.
Les lignes qui suivent replacent les associations rencontrées à leur place synsystématique (sur la première de ces classes, voir de Foucault, 2020b).
ANEMONO NEMOROSAE-CARICETEA SYLVATICAE Gillet ex Gillet & B. Foucault in B. Foucault 2020
Oxalidetalia acetosellae B. Foucault 2020
Primulo vulgaris-Ruscion aculeati B. Foucault 2020
Melico uniflorae-Ruscetum aculeati B. Foucault ?
Mercurialietalia perennis Gillet ex Gillet & B. Foucault in B. Foucault 2020
Dioscoreo communis-Mercurialion perennis B. Foucault 2020
Cephalanthero longifoliae-Daphnetum laureolae B. Foucault
CALLUNO VULGARIS-ULICETEA MINORIS Braun-Blanq. & Tüxen ex Klika & Hadač 1944
Ulicetalia minoris Quantin 1935
Cisto salviifolii-Ericion cinereae Géhu in Bardat et al. 2004
Lavandulo stoechadis-Ericetum cinereae Géhu ex B. Foucault
FRANGULETEA ALNI Doing ex V. Westh. in V. Westh. & Den Held 1969
Rubetalia plicati H.E. Weber in Pott 1995
Frangulo alni-Pyrion cordatae Herrera et al. 1991
Erico scopariae-Franguletum alni Géhu & Géhu-Franck 1975
PINO HALEPENSIS-QUERCETEA ILICIS B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020
Quercetalia rotundifolio-ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020
Aceri monspessulani-Quercion ilicis B. Foucault & Julve ex B. Foucault & Julve in B. Foucault 2020
Quercetum pubescenti-ilicis (Braun-Blanq. 1936) B. Foucault 2020
PISTACIO LENTISCI-RHAMNETEA ALATERNI Julve ex Julve & B. Foucault in B. Foucault 2020
Cytiso villosi-Telinetalia monspessulanae Rivas Mart. et al. in Rivas Mart. et al. 2002
Ericion arboreae Rivas Mart. (1975) 1987
Cytiso scoparii-Ericetum arboreae B. Foucault 2015
RHAMNO CATHARTICAE-PRUNETEA SPINOSAE Rivas Goday & Borja ex Tüxen 1952
Pyro spinosae-Rubetalia ulmifolii Biondi et al. in Biondi et al. 2014
Pruno spinosae-Rubion ulmifolii O. Bolòs 1954
Roso sempervirentis-Viburnetum tini B. Foucault 2015
Sambucetalia racemosae Oberd. ex H. Passarge in Scamoni 1963
Mespilo germanicae-Ilicion aquifolii B. Foucault & Julve ex B. Foucault & J.-M. Royer in B. Foucault 2019
Sorbo torminalis-Crrataegetum germanicae B. Foucault
SCHEUCHZERIO PALUSTRIS-CARICETEA FUSCAE Tüxen 1937
Caricetalia fuscae W. Koch 1926
Juncion acutiflori Braun-Blanq. in Braun-Blanq. & Tüxen 1952
Serratulo seoanei-Molinienion caeruleae B. Foucault 2008
Erico scopariae-Molinietum caeruleae B. Foucault 2008
TRIFOLIO MEDII-GERANIETEA SANGUINEI T. Müll. 1962
Antherico ramosi –Geranietalia sanguinei Julve ex Dengler in Dengler et al. 2003
Inulo conyzae-Galion maritimi B. Foucault & J.-M. Royer in B. Foucault
Inulo conyzae-Galietum maritimi B. Foucault
Origanetalia vulgaris T. Müll. 1962
Trifolion medii T. Müll. 1962
Teucrio scorodoniae-Trifolienion medii R. Knapp 1976
Prunello hastifoliae-Lathyretum nigri B. Foucault
Annexe du tableau 1 – rel. 1 : Roullens, sous domaine Michaud, 17/04/2017 ; rel. 2 et 3 : id. rel. 1, plus à l’est ; rel. 4 : Roullens, sous « Sabran », 13/10/2013 ; rel. 5 : Roullens, entre les jardins communaux et le chemin du Pinier, 01/05/2017 ; rel. 6 : id. rel. 4, 19/05/2018 ; rel. 7 : Carcassonne, La Cavayère, allée des Écureuils, 05/05/2018 ; rel. 8 : id. rel. 4, 10/05/2014 ; rel. 9 : Roullens, près de la vigne communale, 24/01/2015 ; rel. 10 : Montréal, col de la Malepère, près du carrefour D 43 × D 18, 23/05/2018.
Annexe du tableau 2 – rel. 1 : Roullens, sous « Sabran », 13/10/2013 ; rel. 2 : Roullens, entre les jardins communaux et le chemin du Pinier, 01/05/2017 ; rel. 3 : id. rel. 1, 19/05/2018 ; rel. 4 : id. rel. 1, 10/05/2014 ; rel. 5 : Rouffiac-d’Aude, bord D 43, vers « Gaure », 13/05/2020 ; rel. 6 : Preixan, « Le Cascal », bord de la D 211, 21/05/2020 ; rel. 7 : Roullens, sous domaine Michaud, 17/04/2017 ; rel. 8 : id. rel. 7, plus à l’est ; rel. 9 : id. rel. 7, plus à l’est ; rel. 10 : Roullens, près de la vigne communale, 24/01/2015 ; rel. 11 : Carcassonne, La Cavayère, allée des Écureuils, 05/05/2018 ; rel. 12 : Montclar, bord D 43, entre « Lavax » et « La Soulette », 12/05/2020 ; rel. 13 : Cépie, « Peyret », 13/05/2020 ; rel. 14 : Montclar, sur D 43, est « Bédounet », 13/05/2020.
Annexe du tableau 3 – rel. 1, 2, 3, 4, 5 : Montréal, bois du Chapitre sud, 11/07/2019 ; rel. 6 : Montréal, bois du Chapitre, bord du sentier botanique, 11/07/2019 ; rel. 7 : Montréal, bois du Chapitre, vers le Saut Cani, 11/07/2019 ; rel. 8 : Montréal, bois de Las Mounjos, entre le ruisseau de la Combe et la D 18, 12/08/2019 ; rel. 9 : Montréal, bois de Las Mounjos, sud-ouest du domaine des Jasses, 12/08/2019 ; rel. 10 : Villarzel-du-Razès, bois de Caux, vers « Le Dégo », nord RD 43, en direction de « Lavax », 1208/2019 ; rel. 11 : Brugairolles, bois de Las Mounjos, ouest de « Saint-Georges », 17/08/2019 ; rel. 12 : id. rel. 11, Brugairolles, entre « Saint-Georges » et chemin de Montgrenier ; rel. 13 : Montréal, bord D 18, entre le col de la Malepère et le carrefour D 18 × D 318, 12/05/2020 ; rel. 14 : Villarzel-du-Razès, sous le col de la Malepère, sur D 43, 18/05/2020.
Annexe du tableau 5 – rel. 1 : Roullens, sous domaine Michaud, 17/04/2017 ; rel. 2 : id. rel. 1, plus à l’est, 17/04/2017 ; rel. 3 : id. rel. 1, plus à l’est ; rel. 4 : Roullens, sous « Sabran », 13/10/2013 ; rel. 5 : Roullens, entre les jardins communaux et le chemin du Pinier, 01/05/2017 ; rel. 6 : Roullens, sud des jardins communaux, 01/05/2017 ; rel. 7 : Roullens, entre les jardins communaux et la vigne communale, 01/05/2017 ; rel. 8 : Villarzel-du-Razès, « Las Coumelos », 06/05/2017 ; rel. 9 : Roullens, « Sabran », 19/05/2018 ; rel. 10 : Carcassonne, La Cavayère, allée des Écureuils, 05/05/2018 ; rel. 11 : Roullens, est du chemin de Baudrigue, 01/06/2019 ; rel. 12 et 13 : Roullens, sud de « Baudrigue » ; rel. 14 : 10/05/2014 ; rel. 15 : id. rel. 1, 03/06/2019 ; rel. 16 : non loin du rel. 15, 03/06/2019 ; rel. 17 : Roullens, sud des jardins communaux, 03/06/2019 ; rel. 18 : Villarzel-du-Razès, vers carrefour RD 36 × RD 53, 11/06/2019 ; rel. 19 : Roullens, sur RD 36, vers « Cazals », 11/06/2019 ; rel. 20 : Villarzel-du-Razès, sur RD 43, vers « Bédounet », 11/06/2019 ; rel. 21 : Roullens, sur D 36, un peu avant « Sabran », 12/05/2020 ; rel. 22 : Saint-Martin-de-Villereglan, chemin des Trois Pechs, nord de « Rives Blanques », 12/05/2020 ; rel. 23 : Rouffiac-d’Aude, sur D 43, vers « Gaure », 13/05/2020 ; rel. 24 : Montclar, sur D 43, base du sentier du Circaète, 13/05/2020 ; rel. 25 : Montclar, chemin de La Salle, vers 6 Pech des Castels, domaine de Saint-André, 13/05/2020 ; rel. 26 à 29 : Roullens, « Sabran », différents points, 16/05/2020 ; rel. 30 : Alairac, chemin du Moulin, NE « Escande », 18/05/2020 ; rel. 31 : sous col de la Malepère, sur D 18, vers PK 9, 1805/2020 ; rel. 32 : Preixan, « Le Cascal » bord D 211, 21/05/2020 ; rel. 33 : Rouffiac-d’Aude, nord-ouest du bourg, 21/05/2020 ; rel. 34 : Rouffiac-d’Aude, « Les Cassagnos », au-dessus du Clos Teisseire, 21/05/2020 ; rel. 35 : Cépie, « Les Mouligneros », chemin des Moulignières, 21/05/2020 ; rel. 36 : Roullens, « Sabran », 19/05/2018 ; rel. 37 : Montclar, sortie nord sud D 43, est « Bédounet », 13/05/2020 ; rel. 38 : Cépie, « Peyret », 12/05/2020.
Annexe du tableau 6 – rel. 1 : Villarzel-du-Razès, vers carrefour D 36 × D 43, 11/06/2019 ; rel. 2 : Montréal, vers domaine des Jasses, 11/06/2019 ; rel. 3 : Montréal, vers « Cammas de Bas », 11/06/2019 ; rel. 4, 5, 6, 7 : Montréal, divers points au bois du Chapitre, 11/06/2019 ; rel. 8 ; col de la Malepère, vers le carrefour D 43 × D 18, 23/05/2018 ; rel. 9 : id. rel. 8, près de l’abri des chasseurs ; rel. 10 : col de la Malepère, carrefour D 43 × D 18, en direction de Montréal, 23/05/2018 ; rel. 11 : Arzens, bord D 43, entre « Le Prunet » et « Aribaud », 23/05/2018 ; rel. 12 : Montréal, bois de Las Mounjos, entre ruisseau de la Combe et D 18, 12/08/2019 ; rel. 13 : Montréal, bois de Las Mounjos, sud-ouest du domaine des Jasses, 12/08/2019 ; rel. 14 : Montréal, nord de « Montgrenier », vers D 18, 17/08/2019 ; rel. 15 : Montréal, bord D 18, entre le col de la Malepère et le carrefour D 18 × D 318, 12/05/2020 ; rel. 16 : Montclar, bord D 43, entre « La Soulette » et « Lavax », 12/05/2020 ; rel. 17 : Villarzel-du-Razès, sous col de la Malepère, bord D 43, vers « Le Dégo », 18/05/2020.
Annexe du tableau 8 – rel. 1 : Montréal, bois du Chapitre, 11/07/2019 ; rel. 2 : Villarzel-du-Razès, 06/05/2017 ; rel. 3 : Montclar, « La Ricatière » est, 28/04/2019 ; rel. 4 : bois de Las Mounjos, entre la RD 18 et le ruisseau de la Combe, croisement entre 2 sentiers forestiers, 12/08/2019 ; rel. 5 : id. rel. 4, nord du ruisseau de la Combe, sud-ouest du domaine des Jasses, 12/08/2019 ; rel. 6 : Villarzel-du-Razès, nord-est « Montgrenier », 17/08/2019 ; rel. 7 : Roullens, vers « Cazals », 13/10/2013 ; rel. 8 : Saint-Martin-de-Villereglan, chemin des Trois Pechs, au nord de « Rives Blanques », 12/05/2020.
Légende du tableau 9 – rel. 1 : Roullens, truffière à la « Ginestière », 31/05/2015 ; rel. 2 : Roullens, bord D 36, 21/06/2012 ; rel. 3 : Roullens, chemin de Louvat, 17/06/2012 ; rel. 4 : sans localisation ni date ; rel. 5 : Roullens, chemin du Garrau, 01/06/2012 ; rel. 6 : id. rel. 3, 17/06/2012 ; rel. 7 : Roullens, Pech Majou, 10/05/2016.
Légende du tableau 10 – rel. 1 : Villarzel-du-Razès, chemin du Dégo, entre « Le Dégo » et « La Soulette », 16/05/2015 ; rel. 2 : Brugairolles, vers carrefour D 18 × chemin menant à « Montgrenier », 06/06/2015 ; rel. 3 : Roullens, vers domaine Michaud, 14/06/2015 ; rel. 4 : liste, Roullens, 12/08/2011 ; rel. 5 et 6 : Roullens, « Sabran », 16/05/2020 ; rel. 7 : Montclar, vers « La Soulette », 16/05/2015 ; rel. 8 : col de la Malepère, vers carrefour D 43 × D 18, 13/06/2015 ; rel. 9 : Rouffiac-d’Aude, vers domaine de Gaure, sur D 43, 26/08/2016 ; rel. 10 : liste, id. rel. 9, 13/11/2015 ; rel. 11 : Roullens, sous domaine Michaud, 03/06/2019 ; rel. 12 : Villarzel-du-Razès, vers « Les Tourtines », sur D 318, 23/05/2018 ; rel. 13 : Roullens, sud des jardins communaux, 03/06/2019 ; rel. 14 : Roullens, prolongement du chemin du Garrau, vers « Rédounasso », 14/05/2017 ; rel. 15 : limite Montclar – Rouffiac-d’Aude, sur D 43, sud « La Soujeole », 13/05/2020 ; rel. 16 : Montclar, chemin de Malfouich, 13/05/2020 ; rel. 17 : Alairac, chemin du Moulin, nord-est « Escande », 18/05/2020 ; rel. 18 : nord-ouest Alairac, sur D 18, sous le col de la Malepère, 18/05/2020 ; rel. 19 : Rouffiac-d’Aude, rue du Gazan, 21/05/2020.
Légende du tableau 12 – rel. 1, 2, 3, 4 : Montréal, bois du Chapitre, 11/06/2019 ; rel. 5 : Montclar, bois de Caux, nord D 43, en allant vers « Lavax », 12/08/2019 ; rel. 6 : Roullens, « Sabran », 13/05/2020 ; rel. 7 : Montclar, bord D 43, entre « La Soulette » et « Lavax », 12/05/2020 ; rel. 8, 9 : id. rel. 1 à 4, côté sud du bois, 11/07/2019 ; rel. 10 : Montclar, bord D 43, entre « Bédounet » et « Touloury », 17/08/2019 ; rel. 11 : Rouffiac-d’Aude, bord D43 entre le domaine de Gaure et « La Soujeole », 13/05/2020 ; rel. 12 : Montclar, nord-est de « Bédounet », 13/05/2020.
Légende du tableau 13 (les communes sont dans l’Aude, sauf rel. 5, 19, 20, 31, 33 et 34) – rel. 1 : Saint-Polycarpe, vers le « Bois Thérèse », 04/11/2019 ; rel. 2 : Sougraigne, chemin des sources de la Salz, 23/02/2014 ; rel. 3 : nord Villanière, sud de « Carrus », 05/05/2019 ; rel. 4 : Fournes-Cabardès, « Sériès », sous D 401, 12/03/2014 ; rel. 5 : Félines-Minervois (Hérault), est « Labéouradou », vers « Les Graves », 25/03/2015 ; rel. 6 : Roullens, sous « Cazals, 15/03/2014 ; rel. 7 : Caunes-Minervois, « La Boriette », 10/04/2014 ; rel. 8 : Villardonnel, 27/05/2015 ; rel. 9 : Villarzel-du-Razès, nord-est de « Montgrenier », 17/08/2019 ; rel. 10 : Villarzel-du-Razès, vers « Le Dégo », sous D 43, 15/12/2013 ; rel. 11 : Cailhau, forêt domaniale, vers « À Jordenac », 18/05/2014 ; rel. 12 : Montclar, sud de « La Ricatière », 03/07/2015 ; rel. 13 : Villar-Saint-Anselme, sud « Ganibert », 31/10/2015 ; rel. 14 : Roullens, entre le cimetière de le domaine Michaud, 13/10/2013 ; rel. 15 : limite Rennes-les-Bains – Rennes-le-Château, bord de la D 14, 24/05/2020 ; rel. 16 : Albières, sous le col du Paradis, 24/10/2015 ; rel. 17 : Villarzel-du-Razès, bois de Caux, sous « Le Dégo », 18/05/2014 ; rel. 18 : Cabrespine, « Belle-Bruyère », 24/09/2014 ; rel. 19 : vers Réalmont (Tarn), 06/05/2014 ; rel. 20 : Le Truel (Aveyron), vers « La Romiguière », 06/05/2014 ; rel. 21 : Albières, est « Mangoso », sur D 613, 09/03/2014 ; rel. 22 : pech de Bugarach, relevé in Braun-Blanquet & Susplugas (1937 : 678) ; rel. 23 : Saint-Hilaire, col de l’Arc près de « Crausse Rabassié », 15/05/2014 ; rel. 24 : Saint-Martin-le-Vieil, « Les Passérouques », 07/10/2014 ; rel. 25 : id. rel. 16, 24/10/2015 ; rel. 26 : id. rel. 11, 18/05/2014 ; rel. 27 : est Saissac, sur D 103, 17/05/2015 ; rel. 28 : id. rel. 16, 24/10/2015 ; rel. 29 : Rennes-les-Bains, au-dessus de « La Branque », 04/10/2018 ; rel. 30 : Montréal, ouest du bois du Chapitre, non loin de la bergerie, 11/06/2019 ; rel. 31 : id. rel. 20, 06/05/2014 ; rel. 32 : Saint-Just-et-le Bézu, forêt communale (coordonnées GPS non précisées), 16/04/2014 ; rel. 33 et 34 : Vaudreuille (Haute-Garonne), « Lesperou », sur D 79J, 20/09/2015 ; rel. 35 : id. rel. 11, 18/05/2014 ; rel. 36 : Villarzel-du-Razès, entre « Le Dégo » et « La Soulette »16/05/2015 ; rel. 37, 38 et 39 : Cailhau, forêt communale, partie nord, 26/10/2015 ; rel. 40 : Villemagne, site Natura 2000 Vallée du Lampy, 15/06/2019 ; rel. 41 : Montréal, bois des Las Mounjos, entre le ruisseau de la Combe et la D 18, 12/08/2019 ; rel. 42 : id. rel. 13, 31/10/2015.
Légende du tableau 14 – rel. 1 : Issel, vers croisement D 624 × D 126, sous une ligne HT, 27/05/2018 ; rel. 2 et 3 : Issel, « Le Faba », 27/05/2018 ; rel. 4 : Labécède-Lauragais, centre de vol à voile de la Montagne noire, près de la D 334, 11/06/2018 ; rel. 5 : Issel, vers carrefour D 624 × D 624Z, sous une ligne HT, 20/09/2015 ; rel. 6 : Albières, sur D 613, col du Paradis, 09/03/2014.
Annexe du tableau 15 – rel. 1 : Issel, vers carrefour RD 624 × RD 126, 27/05/2018 ; rel. 2 : Issel, « Les Mousques Basses », vers carrefour RD 624 × RD 624Z, 20/09/2015 ; rel. 3 : Issel, « Le Faba », 27/05/2018.
Bibliographie
Alquié A. & Rubio S., 2010. Document d’objectifs du site Natura 2000 « FR9101452 Massif de la Malepère ». Chambre d’agriculture de l’Aude, Carcassonne, 151 p.
Braun-Blanquet J. & Susplugas J., 1937. Reconnaissance phytogéographique dans les Corbières. Bulletin de la Société botanique de France 84 : 669-685.
Chabalier J.-Ch. & Lecomte B., 2013. Guide des stations forestières du Razès, de la Piège, de la Malepère et des confins du Razès et de la Piège. CRPF Languedoc-Roussillon, 91 p.
Foucault B. (de), 1984. Systémique, structuralisme et synsystématique des prairies hygrophiles des plaines atlantiques françaises. Thèse de doctorat d’État, Rouen, 675 p.
Foucault B. (de), 1994. La région de Burlats et le Sidobre. Bulletin de la Société botanique du nord de la France 47 (4) : 17-18.
Foucault B. (de), 1995. Contribution à une monographie phytosociologique de la Hague (Manche, France). Bulletin de la Société botanique du nord de la France 48 (4) : 45-90.
Foucault B. (de), 2015a. Quelques associations arbustives nouvelles pour la Malepère (Aude centrale). Bulletin de la Société botanique du Centre-Ouest, NS, 46 : 244-260.
Foucault B. (de), 2015b. Vers un renouveau de la phytosociologie audoise ? Compte rendu de la sortie phytosociologique du groupe botanique de la SESA. Bulletin de la Société d’études scientifiques de l’Aude CXIV : 193-198.
Foucault B. (de), 2016. Données phytosociologiques collectées lors de sorties du groupe botanique de la SESA durant l’année 2015. Bulletin de la Société d’études scientifiques de l’Aude CXV : 137-145.
Foucault B. (de), 2017a. Quelques ourlets thérophytiques de l’Aude. Botanique 2 : 23-36.
Foucault B. (de), 2017b. Données phytosociologiques collectées lors des sorties du groupe botanique de la SESA durant l’année 2016. Bulletin de la Société d’études scientifiques de l’Aude CXVI : 187-196.
Foucault B. (de), 2017c. Un nouveau syntaxon arbustif pour la Malepère (Aude) : le Coriario myrtifoliae-Spartietum juncei. Bulletin de la Société botanique du Centre-Ouest, NS, 48 : 360-365.
Foucault B. (de), 2019a. Contribution à la connaissance phytosociologique de pelouses, prairies et garrigues de la Malepère (département de l’Aude, France). Evaxiana 5 : 156-186.
Foucault B. (de), 2019b. Quelques aspects de la végétation commensale des cultures de la Malepère (Aude, France). Evaxiana 5 : 187-195.
Foucault B. (de), 2020a. Données phytosociologiques collectées lors de sorties du groupe botanique de la SESA durant l’année 2019. Bulletin de la Société d’études scientifiques de l’Aude CXIX : à paraître.
Foucault B. (de), 2020b. Éloge du paradigme synusial : alternative à la classification phytosociologique de la végétation forestière européenne, ouest-asiatique et nord-africaine. Evaxiana 7 : à paraître.
Foucault B. (de), Beaufils L., Paris A. & Puig S., 2017. La sortie phytosociologique du 8 mai 2016 à Alet-les-Bains. Bulletin de la Société d’études scientifiques de l’Aude CXVI : 196-201.
Foucault B. (de) et Klesczewski M., 2020. Contribution à une monographie phytosociologique du domaine de Peyremale (commune de Montolieu, Aude). Carnets botaniques, en préparation.
Foucault B. (de) & Royer J.-M., 2016. Contribution au prodrome des végétations de France : les Rhamno catharticae-Prunetea spinosae Tüxen 1962. Documents phytosociologiques, 3e série, 2 : 150-343.
Géhu J.-M., 1975. Essai pour un système de classification phytosociologique des landes atlantiques françaises. Colloques phytosociologiques II, La végétation des landes d’Europe occidentale : 361-377.
Géhu J.-M. & Géhu-Franck J., 1975. Contribution à l’étude phytosociologique des landes du sud-ouest de la France. Colloques phytosociologiques II, La végétation des landes d’Europe occidentale : 75-87.
Gillet F., de Foucault B. & Julve Ph., 1991. La phytosociologie synusiale intégrée : objets et concepts. Candollea 46 : 315-340.
Julve Ph. & de Foucault B., 1994. Phytosociologie synusiale dans le Tarn. Bulletin de la Société botanique du nord de la France 47 (4) : 23-47.
Lecomte B. & Moundy P.-J., 2011. Typologie des stations forestières de la Malepère, du Razès, de la Piège et des confins du Razès et de la Piège, 1. Alcina, Fédération Aude Claire et Hélène Chevallier Consultante.
Plassart C., Barreau D. & Andrieu F., 2016. Atlas de la flore patrimoniale de l’Aude. Biotope, Mèze, 432 p.
Royer J.-M., 2020. Contribution au prodrome des végétations de France : les Trifolio medii-Geranietea sanguinei T. Müll. 1962. Documents phytosociologiques, 3e série, 13, sous presse.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.


