
Complément à l’étude phytosociologique du littoral du Languedoc-Roussillon : les dunes de Sainte-Marie-la-Mer à Argelès-sur-Mer ; comparaison avec un système arrière-dunaire atlantique
Title
Complement to the phytosociological study of the Languedoc-Roussillon coast: the dunes from Sainte-Marie-la-Mer to Argelès-sur-Mer; comparison with an Atlantic rear dune system.
Résumé
Des données nouvelles sur la végétation arrière-dunaire du Roussillon confirment la définition de l’Erodio bipinnati-Vulpietum fasciculatae, du Lobulario maritimae-Ephedretum distachyae et de l’Alkanno matthioli-Corynephoretum canescentis, nom toutefois illégitime à remplacer par celui prioritaire de Diantho pyrenaici–Corynephoretum canescentis. On définit comme nouveaux le Malcolmio ramosissimae-Trifolietum cherleri et le Sileno gallicae-Malcolmion ramosissimae au sein des Malcolmietalia ramosissimae et des Tuberarietea guttatae. Une comparaison avec le système arrière-dunaire du Morbihan est aussi effectuée. Une synthèse sur le Corynephorion canescentis est enfin proposée, avec trois nouvelles alliances : le Carici arenariae-Corynephorion canescentis, le Sesamoido spathulifoliae-Jasionion montanae et l’Alkanno matthioli-Corynephorion canescentis.
Abstract
Some new data on the Roussillon dune vegetation confirm the definition of Erodio bipinnati-Vulpietum fasciculatae, Lobulario maritimae-Ephedretum distachyae and Alkanno matthioli-Corynephoretum canescentis, illegitimate name to be replaced by Diantho pyrenaici-Corynephoretum canescentis. The Malcolmio ramosissimae-Trifolietum cherleri and Sileno gallicae-Malcolmion ramosissimae in Malcolmietalia ramosissimae and Tuberarietea guttatae are defined as new. A comparison with the Morbihan dune system is also carried out. A synthesis on Corynephorion canescentis is also proposed, with three new alliances: the Carici arenariae-Corynephorion canescentis, the Sesamoido spathulifoliae-Jasionion montanae and the Alkanno matthioli-Corynephorion canescentis.
Dans une publication précédente (de Foucault, 2017), je présentais le résultat d’investigations phytosociologiques menées sur le littoral languedocien, entre Port-la-Nouvelle (Aude) et Sainte-Marie-la-Mer (Pyrénées-Orientales) ; j’y confirmais notamment une association vivace arrière-dunaire, le Lobulario maritimae-Ephedretum distachyae, validais l’Alkanno matthioli-Corynephoretum canescentis et définissais une association thérophytique arrière-dunaire nouvelle, l’Erodio bipinnati-Vulpietum fasciculatae. De là, il était tentant de poursuivre les investigations plus au sud, donc de Sainte-Marie-la-Mer à Argelès-sur-Mer (Pyrénées-Orientales), jusqu’au pied des falaises de la Côte Vermeille. Les résultats de ces nouvelles recherches sont présentés ici et comparés aux résultats précédents.
Dans son ensemble, le littoral sableux s’étirant de Port-la-Nouvelle à Argelès-sur-Mer se caractérise par des sables quartzeux, plutôt acides, déposés par les fleuves côtiers descendant du massif pyrénéen (Têt, Tech) ; à ce titre il est assez différent du littoral de sable coquillier s’étendant vers le nord-est à partir de Narbonne.
La nomenclature des taxons de France suit Flora Gallica (Tison & de Foucault, 2014) ; pour la synthèse des Malcolmietalia qui finira cette contribution, la nomenclature des taxons absents de France est celle de Euro+Med Plantbase (http://ww2.bgbm.org/EuroPlusMed/query.asp). Les noms des sous-espèces autonymes sont réduits à leur initiale pour gagner de la place ; pour la même raison, dans les tableaux et leur annexe, * remplace « subsp. » ou « var. ».
1. La dune blanche
La première ceinture vivace dunaire correspond à la dune blanche, colonisée par une végétation ouverte, qui s’avère être physionomiquement plus une « agropyraie » qu’une classique « ammophilaie ». Le tableau 1 en rassemble huit relevés nouveaux et décrit l’Echinophoro spinosae-Elytrigietum junceae (Braun-Blanq. 1933) Géhu 1988 nom. mut. hoc loco déjà reconnu dans l’étude de 2017 (sub Echinophoro spinosae-Elymetum farcti). Signalons la rencontre du rare Stachys maritima à Saint-Cyprien-Plage, pratiquement en ville (photo 1).
Sous l’effet des perturbations anthropiques, au milieu des espèces pérennes peut se structurer une végétation thérophytique à Cakile maritima subsp. m. et Xanthium orientale subsp. italica.

2. La végétation vivace de l’arrière-dune
Neuf relevés nouveaux de la végétation vivace arrière-dunaire sont réunis dans le tableau 2.
Globalement, les relevés 1 à 7 se rattachent bien au Lobulario maritimae-Ephedretum distachyae B. Foucault & Julve 1991 [syn. : Diantho catalaunici-Ephedretum distachyae Géhu & Bournique 1992 (Colloq. Phytosoc. XIX : 138) nom. illeg.] ; on ne retient pas ici la référence à Hekking (1959) dont les données se rattachent plutôt au Malcolmio littoreae-Ephedretum distachyae (Hekking 1959) Géhu & Bournique 1992 des plages de sable coquillier (Géhu & Bournique, 1992). Par rapport aux données collectées en 2017 sur les plages plus septentrionales (colonnes A, B et C, respectivement matthioletosum sinuatae, typicum et corrigioletosum telephiifoliae), manquent ici (ou du moins n’ont pas été vus) Teucrium dunense, Ononis ramosissima, Ruta angustifolia, Crucianella maritima (mais présent dans l’Echinophoro–Elytrigietum junceae) ; il semble donc bien y avoir une différenciation nord-sud de cette association, peut-être associée à une différence de richesse chimique du sable. Cette association est par ailleurs reconnaissable dans le tableau V de Baudière et Simonneau (1974).
Les relevés 8 et 9, où manquent de nombreux taxons neutrophiles, se rattachent plutôt à l’Alkanno matthioli-Corynephoretum canescentis Braun-Blanq. ex B. Foucault 2017 acidiphile (photo 2), dans lequel persistent les taxons du Lobulario-Ephedretum qui supportent l’acidification de l’arrière-dune. Notons encore l’absence ici de Lavandula stoechas subsp. s., rencontré de temps à autre dans les arrière-plages plus septentrionales ; mais Baudière et Simonneau (1974, tableau I) le connaissaient au nord de l’embouchure de la Têt. Le nom de ce Corynephoretum arrière-dunaire rappelé en tête de ce paragraphe est en fait illégitime, car ce syntaxon avait déjà été validement publié sous le nom de Diantho catalaunici–Corynephoretum canescentis par Baudière et Simonneau (1974, Vie & Milieu, série C, XXIV (1) : 39), muté en Diantho pyrenaici–Corynephoretum canescentis nom. mut. hoc loco, lectotypus nominis : relevé 17 du tableau II in Baudière et Simonneau (1974, Vie & Milieu, série C, XXIV (1) : 28) ; la validation par Géhu & Bournique (1992 : 138) est donc illégitime. Les données de Baudière et Simonneau (leurs tableaux II et III) sont compilées dans la colonne D de notre tableau 2. Précisons encore que ce Diantho-Corynephoretum avait déjà été remarqué par Braun-Blanquet qui en publia une liste floristique (in Braun-Blanquet et al., 1940 : 36, colonne C).

La présence discrète de Glaucium flavum et Scolymus hispanicus indique une potentialité de dérive vers le Glaucio flavi-Scolymetum hispanici Bartolo et al. 1990 sous l’effet de perturbations d’origine anthropique, comme dans les arrière-dunes plus septentrionales.
3. La végétation thérophytique de l’arrière-dune
Le tableau 3 ordonne les dix-huit relevés de pelouse thérophytique effectués sur les arrière-dunes du littoral roussillonnais. À plusieurs reprises, Polycarpon tetraphyllum s. l. y a été rencontré ; bien que Baudière et Simonneau (1974) l’aient dénommé « P. alsinaefolium », il ne semble pas que ce soit vraiment le taxon actuellement dénommé P. tetraphyllum subsp. alsinifolium (voir discussion in Tison & de Foucault, 2014 : 650).
Les relevés 1 à 10 se rattachent bien à l’Erodio bipinnati-Vulpietum fasciculatae B. Foucault 2017, bien que E. cicutarium subsp. bipinnatum soit un peu moins fréquent que dans les dunes plus septentrionales. On y rencontre en revanche Hypochaeris glabra. La sous-association typicum, différenciée par Limonium echioides… (colonne A), paraît bien appauvrie ici (relevés 1 à 6) ; on y relève 9,3 taxons par relevé en moyenne. La sous-association tolpidetosum umbellatae B. Foucault 2017 (relevés 7 à 10, colonne C) y est par contre bien représentée, bien qu’il y manque Corynephorus divaricatus, Tolpis umbellata et Filago arvensis ; les relevés accueillent en moyenne 13 taxons.
Le reste du tableau 3, soit les relevés 11 à 18, décrit un syntaxon dans lequel de nouveaux taxons acidiphiles apparaissent, surtout Silene gallica, Malcolmia ramosissima, Aira caryophyllea, Trifolium arvense, Sedum caespitosum, Crassula tillaea. Bien que plusieurs espèces liées à l’Erodio-Vulpietum fasciculatae persistent, il s’agit d’un syntaxon original différent de ce dernier, étant plus acidiphile, le Malcolmio ramosissimae-Trifolietum cherleri ass. nov. hoc loco, typus nominis hoc loco : relevé 17 du tableau 3 hoc loco (photo 3). La richesse floristique est ici supérieure puisqu’on compte environ 19 taxons par relevé en moyenne. Les données de Baudière et Simonneau (1974, tableaux II et III) confirment ce syntaxon, en dépit de quelques différences floristiques (tableau 3, colonne D).


4. Comparaison avec un système arrière-dunaire atlantique
Un séjour sur le littoral du Morbihan (La Trinité-sur-Mer, le « Men Du », N 47° 34’ 45,6’’, E 3° 02’ 41,4’’, et « Poulbert », N 47° 34’ 35,9’’, E 3° 02’ 15,1’’ ; Plouharnel, N 47° 35’ 31,3’’, E 3° 07’ 41,9’’ ; juillet 2019) a été l’occasion de parcourir un système d’arrière-dune dont la comparaison avec celui du Roussillon est instructive.
On peut y distinguer :
- une pelouse psammophile vivace à Carex arenaria, Herniaria ciliolata robusta, Cynodon dactylon, Plantago coronopus subsp. c., P. lanceolata, Ephedra distachya subsp. d., Euphorbia segetalis subsp. portlandica, Eryngium campestre, Sedum acre, Allium sphaerocephalon subsp. s., Koeleria arenaria, Poterium sanguisorba subsp. s., Himantoglossum hircinum, Ononis spinosa subsp. procurrens, Dianthus gallicus, Tragopogon pratensis subsp. p., Silene otites, Convolvulus soldanella, Asparagus officinalis subsp. prostratus, Matthiola sinuata, Medicago lupulina;
- une pelouse psammophile annuelle à Silene conica, Vulpia membranacea/fasciculata, Trifolium scabrum s., T. campestre, T. subterraneum, Lagurus ovatus subsp. o., Linaria arenaria, Erodium cicutarium subsp. bipinnatum, E. lebelii, Omphalodes littoralis subsp. l., Hedypnois rhagadioloides, Medicago littoralis, Parentucellia latifolia, Leontodon saxatilis subsp. s., Arenaria serpyllifolia, Phleum arenarium, Cerastium semidecandrum, Hypochaeris glabra, Rumex bucephalophorus subsp. gallicus.
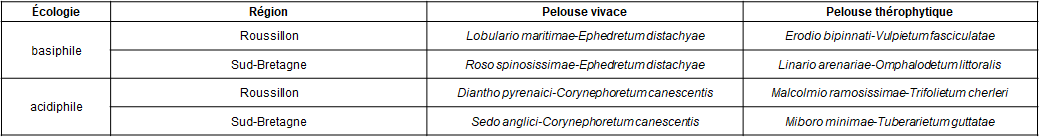
La première se rattache bien au Roso spinosissimae-Ephedretum distachyae Kuhnholtz-Lordat 1931 (Kuhnholtz-Lordat, 1931 ; Vanden Berghen, 1962-63, 1965 ; Géhu, 1964 ; le nom donné par le premier auteur est parfaitement valide). Ici, Rosa spinosissima subsp. s. manque, mais a été rencontré non loin dans l’Ephedretum riverain du musée des guerres de l’Ouest (N 47° 35’ 45,6’’, E 3° 08’ 01,1’’), toujours à Plouharnel (photo 4). Des observations plus étendues montrent qu’en fait ce petit rosier paraît plutôt être un taxon d’ourlet et donc que cet Ephedretum est « ourlifié ». Cet ourlet est alors caractérisé par R. spinosissima, Iris foetidissima, Carex arenaria, Dianthus gallicus, Salix repens subsp. r. var. dunensis et semble proche de l’Euphorbio portlandicae-Rosetum spinosissimae nom. mut. hoc loco décrit du Cotentin occidental (de Foucault, 1995, sub Euphorbio portlandicae-Rosetum pimpinellifoliae ; photo 5) et dont Kuhnholtz-Lordat (1931 : 17) donne aussi un relevé.

Cet ourlet se rattache bien au Galio veri-Geranion sanguinei Géhu & Géhu-Franck 1983 au sein des Trifolio-Geranietea sanguinei mais a été omis dans la synthèse récente de Royer (2015 ; voir alors fiche en annexe comme complément à cette synthèse). Dans la pelouse vivace du « Men Du », s’y ajoutent Helichrysum stoechas subsp. s., Galium arenarium et surtout Coincya monensis subsp. cheiranthos, Rumex acetosella, Jasione montana qui témoignent d’une acidification du système sableux arrière-dunaire. Cette évolution fait dériver le Roso-Ephedretum vers un Corynephoretum canescentis décrit par Géhu (1964, tableau 4) sous le nom de Corynephoro canescentis-Helianthemetum guttati (inversé en Tuberario guttatae-Corynephoretum canescentis par Glemarec, 2013), nom précisé ultérieurement en Sedo anglici–Corynephoretum canescentis (de Foucault, 1999 ; la dominance du second taxon éponyme impose cette forme plutôt que Corynephoro canescentis-Sedetum anglici).
La pelouse annuelle basiphile peut être dégagée des données de Géhu (1964, tableau 3 : relevés 1 à 18), de Vanden Berghen (1965, tableau V) et de Sissingh (1974, tableau 1), qui font apparaître une association à Omphalodes littoralis subsp. l., Linaria arenaria, Phleum arenarium, Arenaria serpyllifolia, Silene conica…, correspondant parfaitement au Linario arenariae-Omphalodetum littoralis défini par Neto et al. (2015).
On peut par ailleurs dégager une pelouse thérophytique acidiphile des données de Géhu (1964, tableau 3 : relevés 19 à 22, tableau 4) et Vanden Berghen (1965, tableau VI), soit 17 relevés, pour décrire une association distincte : Tuberaria guttata V, Mibora minima IV, Teesdalia nudicaulis IV, Aira praecox III, Logfia minima III, Leontodon saxatilis subsp. s. II, Aira caryophyllea II, Erodium « dunense » (E. cicutarium subsp. bipinnatum ?) II, Trifolium arvense I, Cerastium diffusum I, Vulpia membranacea I, Linaria arenaria I, Viola kitaibeliana I, Aira multiculmis I, Hypochaeris glabra I, Anthoxanthum aristatum I, Arenaria serpyllifolia +, Medicago littoralis +, Ornithopus perpusillus +, Silene conica +, Phleum arenarium +, Omphalodes littoralis +, Lagurus ovatus subsp. o. +. À La Trinité-sur-Mer (« Poulbert », à l’est du « Men Du »), il s’y ajoute Vulpia bromoides, Silene gallica, Ornithopus pinnatus, Sedum caespitosum, Lotus angustissimus, Senecio vulgaris subsp. denticulatus. Cette association du Tuberario guttatae-Airion praecocis B. Foucault 1999 (de Foucault, 1999) peut être nouvellement décrite sous le nom de Miboro minimae-Tuberarietum guttatae ass. nov. hoc loco, typus nominis hoc loco : relevé 19 du tableau 3 in Géhu (1964, Bull. Soc. Bot. N. France 17 (4), hors texte).
Cette acidification édaphique pourrait conduire au développement d’une lande arrière-dunaire (de Foucault, 2017) qui n’est cependant pas toujours exprimée. Sur le littoral morbihannais, on peut en observer des lambeaux caractérisés par Calluna vulgaris et Erica ciliaris, avec Danthonia decumbens subsp. d., menacés par les boisements progressifs de pins. Selon Glemarec (2013), elle semble pouvoir se rattacher au Carici arenariae-Ulicetum maritimi Bioret & Géhu 2008 ericetosum ciliaris décrit à la suite des données préliminaires de Wattez & Wattez (1995). Signalons en outre, en le réinterprétant, que le groupement à Corema album de l’Espagne du Nord-Ouest décrit par Losa Quintana (1975) paraît être une lande à C. album, Cistus salviifolius, Ulex gallii, Erica cinerea (Festuco-Corematetum albi), dérivant d’une pelouse arrière-dunaire vivace relevant de l’Iberidion procumbentis. Sur la façade atlantique de l’Amérique du Nord, les dunes acidifiées sont colonisées par des landes à Hudsonia, famille des Cistaceae. C’est le cas au Québec, où H. tomentosa est associé à Vaccinium vitis-idaea, V. angustifolium, Arctostaphylos uva-ursi, Empetrum nigrum, Juniperus horizontalis, Sibbaldia tridentata, Kalmia angustifolia, Diphasiastrum tristachyum, Ammophila breviligulata, au contact de fourrés oligotrophiles à Myrica pensylvanica, Comptonia peregrina, Diervillea lonicera, Aralia hispida (Petitclerc & Dignard, 2004). De même à Saint-Pierre et Miquelon, Hudsonia ericoides constitue des landes de basse altitude avec des Ericaceae, Diapensia lapponica… (Muller, 2008). Cette combinaison de Cistaceae et Ericaceae rappelle la structure formelle systématique S(Lamiaceae-Cistaceae) qui recouvre les Cisto-Lavanduletea stoechadis et les Cisto-Micromerietea julianae méditerranéens (de Foucault, 1987) ; ici la tonalité est nettement plus boréale (présence de Diapensiaceae, Lycopodiaceae…) et les Lamiaceae manquent. On peut alors définir la nouvelle S(Cistaceae-Ericaceae) pour de telles landes.
Le tableau 4 dresse les homologies entre le système du Roussillon et celui du Morbihan.
5. Synthèse des Malcolmietalia Rivas Goday 1958
La place synsystématique de l’Erodio bipinnati-Vulpietum fasciculatae a fait l’objet d’une discussion dans l’article de 2017 : il se place dans les Ononido variegatae-Cutandietea maritimae B. Foucault & Géhu in B. Foucault ex B. Foucault 2017, les Cutandietalia maritimae Rivas Mart. et al. ex Rivas Mart. et al. 2002 et le Laguro ovati-Vulpion fasciculatae Géhu & Biondi 1994. Le présent paragraphe va maintenant s’attacher à fixer celle du Malcolmio-Trifolietum cherleri.
Au sein de la classe des pelouses thérophytiques xéro-acidiphiles, les Tuberarietea guttatae (Rivas Goday 1958) Rivas Goday & Rivas Mart. 1963, on distingue deux ordres, les Tuberarietalia guttatae Braun-Blanq. in Braun-Blanq. et al. 1940 et les Malcolmietalia Rivas Goday 1958 (nom précisé en Malcolmietalia ramosissimae par Bardat et al., 2004) ; cette distinction d’ordre est confirmée dans le tableau synthétique 2 in de Foucault (1999). Ce dernier ordre rassemble des pelouses thérophytiques sabulicoles littorales dérivant souvent de pelouses neutro-basiphiles sous l’effet d’une acidification édaphique (voir des aspects systémiques et structuralistes in de Foucault, 2017) ; il est caractérisé par Malcolmia triloba (= M. patula, M. lacera ; incl. M. broussonetii selon Euro+Med Plantbase, mais ce dernier distingué sous M. lacera subsp. broussonetii par Fennane et al., 1999), Linaria spartea, Ornithopus sativus subsp. isthmocarpus, Loeflingia baetica, Cladanthus mixtus, Brassica barrelieri, Filago astericiflora, Brassica oxyrrhina, Avellinia festucoides, Corynephorus divaricatus et différencié des Tuberarietalia guttatae par des taxons caractérisant les Ononido-Cutandietea maritimae résistant à l’acidification édaphique et donc persistant dans les pelouses sabulicoles dérivées, tels que Medicago littoralis, Vulpia membranacea, Silene portensis, Pseudorlaya pumila, Erodium cicutarium subsp. bipinnatum. Ce cortège floristique mixte a amené plusieurs auteurs à rattacher aux Malcolmietalia des pelouses thérophytiques psammophiles quasiment dépourvues de taxons acidiphiles et donc mieux placées dans les Ononido-Cutandietea maritimae.
Selon Rivas Goday (1957), cet ordre se déclinerait en quatre alliances. Mais l’Alkanno–Malcolmion ramosissimae, renommé ultérieurement Maresio nanae-Malcolmion ramosissimae (Rivas Mart. 1978) Rivas Mart., M.J. Costa & Loidi 1992, est typifié par l’Erodio laciniati-Malcolmietum parviflorae Rivas Goday 1958 ; ce nom a été modifié ultérieurement (Rivas-Martínez et al., 2002) en Erodio laciniati-Maresietum nanae Rivas Goday 1958 corr. M.J. Costa, Díez-Garretas, P. Soriano & Pérez-Badia in Pérez-Badia 1997 en incluant le Loeflingio pentandrae-Maresietum nanae Alcaraz, Mateo, Figuerola, Díez-Garretas & Asensi 1987 (Alcaraz et al., 1987) et le Sileno ramosissimae-Laguretum ovati Rivas Goday & Rigual 1959 (Rivas Goday & Rigual, 1959). Pourtant tous ces syntaxons se rattachent clairement aux Ononido-Cutandietea maritimae. On ne gardera donc pas ici l’Alkanno–Malcolmion ramosissimae actualisé en Maresio nanae-Malcolmion ramosissimae. Les associations retenues au sein des Malcolmietalia sont classées selon l’ordination obtenue dans le tableau 5, qui s’écarte notablement de celle déclinée par Rivas-Martínez (1978).
Malcolmietalia ramosissimae Rivas Goday 1958 (Anales Inst. Bot. Cavanilles 15 : 549)
Typus allianciae : Anthyllido hamosae-Malcolmion trilobae Rivas Goday 1958 (Anales Inst. Bot. Cavanilles 15 : 564, sub Anthyllido hamosae-Malcolmion lacerae), lectotype désigné par Rivas-Martínez (1978 : 62).
Sileno gallicae-Malcolmion ramosissimae all. nov. hoc loco (tableau 5 : colonnes en SM)
Typus nominis hoc loco : Malcolmio ramosissimae-Trifolietum cherleri B. Foucault hoc loco.
Pelouses surtout nord-méditerranéennes à Malcolmia ramosissima, Lagurus ovatus subsp. o., Silene gallica, Polycarpon tetraphyllum subsp. diphyllum, Erodium laciniatum, Aira cupaniana, Plantago coronopus subsp. c., P. bellardii, Corynephorus articulatus, Vulpia fasciculata. Seule alliance de l’ordre présente en France.
SM1. Malcolmio ramosissimae-Trifolietum cherleri B. Foucault (tableau 3 hoc loco : relevés 11 à 18).
SM2. Malcolmietum ramosissimae Molin. 1954 (Loisel, 1976, tableau XI) ; lectotypus nominis hoc loco : composante thérophytique du relevé 8 du tableau I in Molinier (1954, Vegetatio V-VI : 258).
SM3. Malcolmio ramosissimae-Linarietum sardoae Bartolo, Brullo, De Marco, Dinelli, Signorello & Spampinato 1992 (Bartolo et al., 1992, tableau 8).
SM4. Sileno nicaeensis-Cladanthetum mixti sensu Bartolo, Brullo, De Marco, Dinelli, Signorello & Spampinato 1992 (Colloq. Phytosoc. XIX : 257) nom. mut. hoc loco, non Brullo in Brullo, De Santi, Furnari, Longhitano & Ronsisvalle 1988 (Bartolo et al., 1992, tableau 9, sub Sileno nicaeensis-Chamaemeletum mixti) ; le Sileno nicaeensis-Cladanthetum mixti Brullo in Brullo, De Santi, Furnari, Longhitano & Ronsisvalle 1988 nom. mut. hoc loco (Brullo et al., 1988) relève plutôt des Ononido-Cutandietea maritimae.
SM5. Sileno gallicae-Corynephoretum articulati Géhu & Biondi 1994 (Paradis & Piazza, 1993, tableau 18 ; Géhu & Biondi, 1994, tableau 18 ; Piazza & Paradis, 1995, tableau 8).
SM7. Sileno nummicae–Malcolmietum ramosissimae Biondi & Bagella 2005 (Biondi & Bagella, 2005, tableau 41).
SM6. Malcolmio ramosissimae-Vulpietum membranaceae O. Bolòs, Molin. & P. Monts. 1970 (de Bolòs et al., 1970, tableau 25).
Corynephoro divaricati-Malcolmion trilobae Rivas Goday 1958 (Anales Inst. Bot. Cavanilles 15 : 550, sub Corynephoro-Malcolmion) (tableau 5 : colonnes en CM)
Typus allianciae : Loeflingio hispanicae-Malcolmietum trilobae Rivas Goday 1958 (Anales Inst. Bot. Cavanilles 15 : 551, sub Loeflingio hispanicae-Malcolmietum patulae), lectotype désigné par Rivas-Martínez (1978 : 63).
Cœur de l’ordre, partageant avec les deux alliances suivantes des taxons considérés comme caractéristiques ou différentiels de l’ordre, Malcolmia triloba… ; pelouses plutôt ibéro-marocaines, surtout décrites par Rivas Goday (1958).
CM1. Loeflingio hispanicae-Malcolmietum trilobae Rivas Goday 1958 nom. mut. hoc loco (Rivas Goday, 1957, tableau 1, sub Loeflingio hispanicae-Malcolmietum patulae).
CM2. Ornithopodo sativi–Malcolmietum trilobae Rivas Goday 1958 nom. mut. hoc loco (Ibid., tableau 2, sub Ornithopodo macrorrhynchi–Malcolmietum patulae).
CM3. Loeflingio baeticae-Erodietum cicutarii Rivas Goday 1958 nom. mut. hoc loco (Ibid., tableau 3, sub Loeflingio baeticae-Erodietum aethiopici).
CM4. Leucanthemopsio pulverulentae-Malcolmietum trilobae Rivas Goday 1958 nom. mut. hoc loco (Ibid., tableau 4, sub Chrysanthemo pulverulenti-Malcolmietum patulae).
CM5. Linario donyanae-Loeflingietum baeticae Rivas Mart. et al. in M.J. Costa et al. 1978 (Costa et al., 1978, tableau 1).
CM6. Cladantho mixti-Malcolmietum trilobae Nègre 1978 nom. mut. hoc loco (Nègre, 1978, tableau 2 : « Ormenieto-Malcolmietum »).
CM7. Wahlenbergio nutabundae-Loeflingietum pentandrae Alcaraz, Díez-Garretas & Asensi in Ferre, Díez-Garretas & Asensi 1985 (Ferre et al., 1985, tableau 1).
Anthyllido hamosae-Malcolmion trilobae Rivas Goday 1958 nom. mut. hoc loco (Anales Inst. Bot. Cavanilles 15 : 564, sub Anthyllido hamosae-Malcolmion lacerae) (tableau 5 : colonnes en AM)
Typus allianciae : Arenario emarginatae–Hymenostemmatetum pseudanthemidis Rivas Goday 1958 (Anales Inst. Bot. Cavanilles 15 : 570), lectotype désigné par Rivas-Martínez (1978 : 63, ‘… Hymenostemmetum pseudanthemis’ art. 41b).
Pelouses à Anthyllis hamosa, Ononis subspicata, O. serrata, Anthoxanthum ovatum, Vulpia alopecuros, Coronilla repanda.
AM1. Anthyllido hamosae-Malcolmietum trilobae (Rivas Goday & Borja1948) Rivas Goday 1958 nom. mut. hoc loco (Rivas Goday, 1957 : 565, sub Anthyllido hamosae-Malcolmietum patulae).
AM2. Malcolmio lacerae–Anthyllidetum hamosae Rivas Goday 1958 (Ibid., tableau 6) ; après application de la synonymie actuelle au sein du groupe de Malcolmia triloba, ce nom s’avère homonyme du précédent ; il devient alors nécessaire de créer un nouveau nom (art. 44) : Ornithopodo isthmocarpi-Malcolmietum trilobae (Rivas Goday 1958) nom. nov. hoc loco ; typus nominis hoc loco : relevé 5 du tableau 6 in Rivas Goday (1957, Anales Inst. Bot. Cavanilles 15 : 568).
AM3. Arenario emarginatae–Hymenostemmatetum pseudanthemidis Rivas Goday 1958 (Ibid., tableau 7).
AM4. Anthylllido hamosae-Senecionetum gallici Rivas Goday 1958 nom. mut. hoc loco (Ibid., tableau 8, sub Anthylllido hamosae-Senecionetum exsquamei).
AM5. Corynephoro macrantheri-Arenarietum algarbiensis P. Silva & Telès in Rivas Mart. et al. 2002 (Rivas-Martínez et al., 2002 : 83).
AM6. Tolpido barbatae-Tuberarietum commutatae J.C. Costa, Lousã & Espírito-Santo 1997 nom. mut. hoc loco (Costa et al., 1997, tableau 22, sub Tolpido barbatae-Tuberarietum bupleurifoliae).
Cladantho mixti-Malcolmion broussonetii Braun-Blanq. 1940 nom. mut. hoc loco (Prodrome Group. Vég. 7 : 48, sub Ormenido multicaulis-Malcolmion broussonetii) (tableau 5 : colonne OM)
Typus allianciae : Ammochloo involucratae– Cladanthetum mixti Braun-Blanq. 1940 (Prodrome Group. Vég. 7 : 49).
Pelouses nord-ouest-marocaines à Filago carpetana subsp. maroccana, Hedypnois arenaria, Heteranthemis viscidehirta, Ammochloa involucrata, Tolpis nemoralis, Silene gracilis, Ononis cossoniana ; alliance actuellement connue uniquement par son association type décrite de la Maâmora, près de Rabat (Maroc). D’après notre tableau 5, cette alliance et la précédente pourraient être réunies, le Cladantho mixti-Malcolmion broussonetii étant alors prioritaire.
OM. Ammochloo involucratae–Cladanthetum mixti Braun-Blanq. 1940 nom. mut. hoc loco (Braun-Blanquet, 1940 : 49, sub Ammochloo involucratae–Anthemidetum multicaulis).
6. Synthèse du Corynephorion canescentis Klika 1931
Parallèlement à la synthèse précédente sur les Malcolmietalia thérophytiques, une autre peut être aussi menée, celle des pelouses vivaces ouvertes sur sables acides à Corynephorus canescens, classiquement placées dans le Corynephorion canescentis Klika 1931 (Sb. Česk. Akad. Zemed. 89 : 295). Cette synthèse s’appuie sur notre tableau 6 élaboré sur les syntaxons suivants.
* Pelouses ibériques
Cc1. Groupement à Plantago radicata-Corynephorus canescens (Rivas Goday & Borja Carbonell, 1961, tableau 51).
Cc2. Herniario unamunoanae–Corynephoretum canescentis Pinto Gomes et al. 2006 (Pinto Gomes et al., 2006, tableau III), en lien avec une lande à Lavandula pedunculata subsp. lusitanica et Ulex genistoides ; dans cette même publication, deux autres Corynephoreta sont aussi décrits ; cependant, bien moins acidiphiles, ils ne relèvent pas du Corynephorion canescentis mais plutôt du Crucianellion maritimae.
Cc3. Rumici acetosellae-Neoschischkinietum delicatulae (Rivas Goday 1958) ass. nov. hoc loco (Sánchez Mata, 1989, tableaux 48 et 49, sub Triseto ovati-Agrostietum truncatulae), typus nominis hoc loco : composante vivace du relevé 8 du tableau 48 in Sánchez Mata (1989, Flora y vegetacion del macizo… : 160…).
* Pelouses franco-méditerranéennes, souvent à Alkanna matthioli, Poterium verrucosum, Paronychia argentea
Cc4. Poo bulbosae-Corynephoretum canescentis (Aubert & R.J. Loisel 1971) ass. nov. hoc loco (Aubert & Loisel, 1971, tableau IX, sub Teesdalio-Corynephoretum canescentis) des sables siliceux du bassin d’Apt, à Corynephorus canescens, Poa bulbosa, Arabis hirsuta, Sedum ochroleucum, Alkanna matthioli, Poterium verrucosum ; typus nominis hoc loco : composante vivace du relevé 5 du tableau IX in Aubert & Loisel (1971, Ann. Univ. Provence XLV, sci., hors texte).
Cc5. Groupement basal à Corynephorus canescentis (Loisel, 1976, tableau XVII : relevés 265 à 273).
Cc6. Diantho pyrenaici-Corynephoretum canescentis Baudière & Simonneau 1974 (tableau 2 hoc loco : relevés 8 et 9 et colonne D) du littoral languedocien.
Cc7. Corrigiolo telephiifoliae-Jasionetum montanae B. Foucault 1999 (Géhu & Biondi, 1994, tableau 17 ; d’autres relevés reconnaissables chez Paradis & Piazza, 1991, tableau 1 : relevé 29, tableau 5 : relevé 28 ; chez Paradis & Piazza, 1993, tableau 18) du littoral corse.
* Pelouses plutôt atlantiques à Luzula campestris, Potentilla neglecta, Armeria arenaria subsp a., Anthoxanthum odoratum
Cc8. Groupement à Luzula campestris-Corynephorus canescens (Lemée, 1937, tableau 6, sub Corynephoretum occidentale ; Wattez et al., 1978, tableau 4) du Perche et de Brenne ; assez basal.
Cc9. Koelerio macranthae-Corynephoretum canescentis P. Allorge 1922 nom. mut. hoc loco (Allorge, 1922, tableau XV, sub Koelerio gracilis-Corynephoretum canescentis) du Vexin français ; chez l’auteur, le nom est souvent donné sous la forme de « pelouse siliceuse à… », en principe invalide (art. 3c) ; dans le texte, on trouve cependant les formes « association à… » ou « Coynephoretum », valides, d’où le choix de ne pas enlever l’autorité d’Allorge ; lectotypus nominis hoc loco : composante vivace du relevé 5 du tableau XV in Allorge (1922, Les associations végétales du Vexin français : 194).
Cc10. Groupement à Corynephorus canescens de la vallée de la Seine (Frileux, 1978, tableau 3), forme appauvrie du précédent.
Cc11. Sesamoido purpurascentis-Corynephoretum canescentis Braun-Blanq. 1967 nom. mut. hoc loco (Géhu, 1974, sub Astrocarpo sesamoidis-Corynephoretum canescentis) de Sologne et des Landes de Gascogne.
* Pelouses eu- à nord-atlantiques arrière-dunaires à Carex arenaria, Koeleria arenaria, Viola canina (incl. var. dunensis)
Cc12. Violo dunensis-Corynephoretum canescentis V. Westh. et al. ex Géhu & B. Foucault 1978 forme du littoral néerlandais (Boerboom, 1960, tableau 4, dans lequel manque le premier taxon éponyme).
Cc13. Violo dunensis-Corynephoretum canescentis V. Westh. et al. ex Géhu & B. Foucault 1978 forme du littoral du nord de la France (Géhu & de Foucault, 1978, tableau I), lectotypus nominis hoc loco : composante vivace du relevé 9 du tableau I in Géhu & de Foucault (1978, Colloq. Phytosoc. VI : 321).
Cc14. Sedo anglici-Corynephoretum canescentis (Géhu 1964) B. Foucault 1999 nom. inv. (Géhu, 1964, tableau 4) des îles morbihanaises.
Cc15. Groupement à Carex trinervis-Corynephorus canescens (Géhu & de Foucault, 1978, tableau II).
* Pelouse à Carex arenaria non arrière-dunaire
Cc16. Groupement à Agrostis vinealis-Rumex acetosella (Depasse et al., 1970, tableau 3) ; Corynephorus canescens fut présent dans le site étudié par les auteurs, mais en était disparu lors de leur étude.
* Pelouses continentales à Potentilla verna, Sedum rupestre, Scleranthus perennis, Potentilla neglecta, Armeria arenaria subsp. a.
Cc17. Anarrhino bellidifolii-Corynephoretum canescentis J.-M. Royer & B. Foucault ass. nov. hoc loco, à Corynephorus canescens, Sesamoides purpurascens, Anarrhinum bellidifolium, Scleranthus perennis, Rumex acetosella, typus nominis hoc loco : composante vivace du relevé 14 du tableau I in Royer (1971, Ann. Sci. Univ. Besançon 10) ; a – race différenciée négativement (Loiseau & Felzines, 2007, tableau 1, sub Astrocarpo purpurascentis-Corynephoretum canescentis) ; b – race à Armeria arenaria, Festuca pallens, Potentilla verna (Royer, 1971, tableau I, sub Asterocarpo-Corynephoretum).
Cc18. Corynephoretum canescentis Braun-Blanq. 1915 (Loiseau & Felzines, 2007, tableau 7 : colonne 13), des Cévennes ; étude à reprendre plus complètement.
Cc19. Groupement à Corynephorus canescens des monts d’Ardèche (Choisnet & Mulot, 2008, tableau E : colonne 3).
Cc20. « Tuberario guttatae-Corynephoretum canescentis sensu Loiseau & Felzines 2007 » (Loiseau & Felzines, 2007, tableau 2).
Cc21. « Micropyro tenelli-Corynephoretum canescentis Loiseau & Felzines 2007 » (Loiseau & Felzines, 2007, tableau 3).
Cc22. Sedo albi-Corynephoretum canescentis Loiseau & Felzines 2007 (Loiseau & Felzines, 2007, tableau 4, sub Sedo micranthi-Corynephoretum canescentis).
Cc23. Pilosello ligericae-Corynephoretum canescentis Loiseau & Felzines 2007 nom. mut. hoc loco (Loiseau & Felzines, 2007, tableau 5, sub Hieracio ligerici-Corynephoretum canescentis), qui peut se réduire à une variation à Pilosella peleteriana subsp. ligerica (= Hieracium peleterianum subsp. ligericum) et Sedum sexangulare du précédent.
Cc24. « Sileno armeriae-Corynephoretum canescentis Loiseau & Felzines 2007 » (Loiseau & Felzines, 2007, tableau 6).
Cc25. Corynephoretum canescentis Tüxen 1928 [syn. : Corniculario aculeatae-Corynephoretum canescentis Steffen 1931] (Hohenester, 1967, tableau hors texte, sub Androsaco-Corynephoretum, Veronico-Corynephoretum et Spergulo-Corynephoretum), continental.
Habituellement, toutes ces pelouses acidiphiles à Corynephorus canescens sont rattachées aux Corynephoretalia canescentis Klika 1934 (Beih. Bot. Centralbl. 52 : 14, typifié par le Corynephorion canescentis Klika 1931) et à une classe autonome, celle des Koelerio glaucae-Corynephoretea canescentis Klika in Klika & V. Novak 1941 (Praktikum rostlinné sociologie… : 59), incluant par ailleurs diverses alliances de pelouses basiphiles (Bardat et al., 2004). En 1999, j’ai proposé de ne pas conserver cette classe et de verser le Corynephorion dans l’ordre lui-même acidiphile des Sedo albi-Scleranthetalia biennis Braun-Blanq. 1955 repris par Royer & Ferrez (2018) dans leur synthèse des Sedo albi-Scleranthetea biennis Braun-Blanq. 1955. Le tableau 6 présenté ici confirme ce fort lien synsystématique par la présence de Jasione montana et Rumex acetosella. Selon le paradigme suivi ici, il n’est en outre pas possible de suivre Loiseau et Felzines (2007) qui définissent, en plus de l’alliance de Klika, le « Miboro minimae-Corynephorion canescentis » et le Sedo micranthi-Corynephorion canescentis, ce dernier incluant selon le tableau 6 hoc loco le type du Corynephorion canescentis classique.
Le Corynephorion canescentis Klika 1931 concerne la partie Cc17a à Cc25 du tableau 6, incluant le type (le Corynephoretum canescentis Tüxen 1928, syntaxon Cc25). Étant donné la forte différenciation des syntaxons au sein de ce tableau, on peut se demander s’il ne serait pas utile de distinguer d’autres alliances voisines. La mieux définie rassemble les syntaxons Cc12 à Cc16, caractérisée par Carex arenaria, Koeleria arenaria et correspondant aux pelouses eu- à nord-atlantiques arrière-dunaires, plus rarement non arrière-dunaires : Carici arenariae-Corynephorion canescentis all. nov. hoc loco, typus nominis hoc loco : Violo dunensis-Corynephoretum canescentis V. Westh. et al. ex Géhu & B. Foucault 1978 (Colloq. Phytosoc. VI : 320). Une seconde bien caractérisée est représentée par les syntaxons Cc4 à Cc6, des arrière-dunes acidifiées du littoral franco-méditerranéen avec Alkanna matthioli, Poterium verrucosum, Paronychia argentea : Alkanno matthioli-Corynephorion canescentis all. nov. hoc loco, typus nominis hoc loco : Diantho pyrenaici-Corynephoretum canescentis Baudière & Simonneau 1974 (Vie & Milieu, série C, XXIV (1) : 39). Par enchaînement, on pourrait y inclure provisoirement le syntaxon corse Cc7 ; mais Corynephorus canescens et Alkanna matthioli manquent en Corse ; il est alors préférable d’isoler une alliance originale, le Sesamoido spathulifoliae-Jasionion montanae all. nov. hoc loco, typus nominis hoc loco : Corrigiolo telephiifoliae-Jasionetum montanae B. Foucault 1999 (Doc. Phytosoc., NS, XIX : 81).
Un des problèmes subséquents à cette position est que les Corynephoretalia canescentis Klika 1934 sont prioritaires sur les Sedo albi-Scleranthetalia biennis Braun-Blanq. 1955 et les Koelerio glaucae-Corynephoretea canescentis Klika in Klika & V. Novák 1941 sur les Sedo albi-Scleranthetea biennis Braun-Blanq. 1955. Ce point a été aussi relevé par Dengler et al. (2003) qui réalisent effectivement cette inclusion et définissent deux sous-classes, les Koelerio-Corynephorenea (Klika in Klika & V. Novák) Dengler in Dengler et al. 2003, plutôt à dénommer maintenant Eu–Koelerio glaucae-Corynephorenea canescentis, et les Sedo-Scleranthenea (Braun-Blanq. 1955) Dengler in Dengler et al. 2003. On en restera ici à cette simple remarque d’ordre synsystématique sans prétendre reprendre toute la classe.
7. Conclusion
Cette nouvelle étude de la végétation arrière-dunaire du Roussillon confirme donc plusieurs associations rencontrées un peu plus au nord (Lobulario maritimae-Ephedretum distachyae ; Alkanno matthioli-Corynephoretum canescentis, nom toutefois illégitime à remplacer par Diantho pyrenaici–Corynephoretum canescentis ; Erodio bipinnati-Vulpietum fasciculatae). Elle met aussi en évidence une association nouvelle, le Malcolmio ramosissimae-Trifolietum cherleri qui s’intègre en la typifiant dans une alliance originale des Malcolmietalia ramosissimae, le Sileno gallicae-Malcolmion ramosissimae. D’un autre côté, la synthèse du Corynephorion canescentis a permis la définition du Poo bulbosae-Corynephoretum canescentis et de l’Anarrhino bellidifolii-Corynephoretum canescentis, ainsi que la reconnaissance des nouveaux Carici arenariae-Corynephorion canescentis, Sesamoido spathulifoliae-Jasionion montanae et Alkanno matthioli-Corynephorion canescentis.
Bibliographie
Alcaraz F., Mateo G., Figuerola R., Díez-Garretas B. & Asensi A., 1987. El orden Malcolmietalia Rivas Goday 1957 en el litoral mediterráneo-ibérico. Studia Botanica 6 : 47-51.
Allorge P., 1922. Les associations végétales du Vexin français. Nemours, 336 p.
Aubert G. & Loisel R., 1971. Contribution à l’étude des groupements végétaux des Isoeto-Nanojuncetea et des Helianthemetea annua dans le Sud-Est méditerranéen. Annales de l’université de Provence XLV, sciences : 203-241.
Bardat J., Bioret F., Botineau M., Boullet V., Delpech R., Géhu J.-M., Haury J., Lacoste A., Rameau J.-C., Royer J.-M., Roux G. & Touffet J., 2004. Prodrome des végétations de France. Patrimoines naturels 61 : 1-171.
Bartolo G., Brullo S., De Marco G., Dinelli A., Signorello P. & Spampinato G., 1992. Studio fitosociologico sulla vegetazione psammofila della Sardegna meridionale. Colloques phytosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 251-273.
Baudière A. & Simonneau P., 1974. Les groupements à Corynephorus canescens (L.) P. Beauv. et à Ephedra distachya L. du littoral roussillonnais. Vie & Milieu, série C, XXIV (1) : 21-42.
Biondi E. & Bagella S., 2005. Vegetazione e paesaggio vegetale dell’arcipelago di La Maddalena (Sardegna nord-orientale). Fitosociologia 42 (2), suppl. 1 : 3-99.
Bolòs O. (de), Molinier R. & Montserrat P., 1970. Observations phytosociologiques dans l’île de Minorque. Acta Geobotanica Barcinonensia 5 : 1-150.
Boerboom J.H.A., 1960. De plantegemeenschappen van de Wassenaarse duinen. Thèse, Wageningen, 135 p. et annexes.
Braun-Blanquet J. (et collaboration Molinier R. & Wagner H.), 1940. Classe Cisto-Lavanduletea (landes siliceuses à cistes et lavandes). Prodrome des groupements végétaux 7 : 1-51.
Brullo S., De Santi C., Furnari F., Longhitano N. & Ronsisvalle G.A., 1988. La vegetazione dell’Oasi della Foce del Simeto (Sicilia orientale). Braun-Blanquetia 2 : 165-188.
Choisnet G. & Mulot P.-E., 2008. Catalogue des végétations du Parc naturel régional des Monts d’Ardèche. Conservatoire botanique national du Massif central/Conseil régional Rhône-Alpes, Chavaniac-Lafayette, 263 p.
Costa J.C., Lousã M. & Espírito-Santo M.D., 1997. A vegetação do Parque Natural da Ria Formosa (Algarve, Portugal). Studia Botanica 15 : 69-157.
Costa M., Castroviejo S., Rivas-Martínez S. & Valdés-Bermejo E., 1978. Sobre la vegetación de terófitos efímeros de las dunas fósiles del Coto de Doñana. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 101-108.
Dengler J., Berg C., Eisenberg M., Isermann M., Jansen F., Koska I., Löbel S., Manthey M., Päzolt J., Spangenberg A., Timmermann T. & Wollert H., 2003. New descriptions and typifications of syntaxa within the project ‘Plant communities of Mecklenburg-Vorpommern and their vulnerability’, part I. Feddes Repertorium 114 (7-8) : 587-631.
Depasse S., Duvigneaud J. & De Zuttere Ph., 1970. Le site de la Mer de sable à Stambruges (province de Hainault, Belgique). Lejeunia, NS, 54 : 1-36.
Fennane M., Ibn Tattou M., Mathez J., Ouyahya A. & El Oualidi J., 1999. Flore pratique du Maroc, manuel de détermination des plantes vasculaires, 1. Institut scientifique, Université Mohammed V, Rabat, 558 p.
Ferre E., Díez-Garretas B. & Asensi A., 1985. Relaciones geomorfológicas-vegetación en el litoral sudeste de la provincia de Almería (España). Documents phytosociologiques, NS, IX : 445-454.
Foucault B. (de), 1987. Nouvelles recherches sur les structures systématiques végétales : caractérisation, ordination, signification. Phytocoenologia 15 (2) : 159-199.
Foucault B. (de), 1995. Synthèse phytosociologique sur la végétation observée dans le Cotentin (Manche, France). Bulletin de la Société botanique du nord de la France 48 (4) : 29-44.
Foucault, B. (de), 1999. Nouvelle contribution à une synsystématique des pelouses sèches à thérophytes. Documents phytosociologiques, NS, XIX : 47-105.
Foucault B. (de), 2017. Contribution à la connaissance phytosociologique du littoral sableux du Languedoc-Roussillon (Pyrénées-Orientales et Aude, France) ; essai structuraliste sur les systèmes sableux arrière-littoraux. Botanique 3 : 21-37.
Frileux P.-N., 1978. Aperçu de la végétation des pelouses sèches à thérophytes de Haute-Normandie (basses vallées de la Seine et de l’Eure). Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 169-175.
Géhu J.-M., 1964. La végétation psammophile des îles de Houat et Hoëdic. Bulletin de la Société botanique du nord de la France 17 (4) : 238-266.
Géhu J.-M., 1974. L’Astrocarpo–Corynephoretum canescentis Br.-Bl. 1967 dans les Landes de Gascogne. Documents phytosociologiques 6 : 43-45.
Géhu J.-M. & de Foucault B., 1978. Les pelouses siliceuses à thérophytes de la zone littorale du nord de la France. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 319-327.
Géhu J.-M. & Bournique C.-P., 1992. Interprétation phytosociologique actualisée et comparative des vestiges de végétation du cordon littoral entre Sète et Agde (Languedoc). Colloques phytosociologiques XIX, Végétation et qualité de l’environnement côtier en Méditerranée : 133-146.
Géhu J.-M. & Biondi E., 1994. Végétation du littoral de la Corse. Essai de synthèse phytosociologique. Braun-Blanquetia 13 : 1-149.
Glemarec E., 2013. Deux phytocénoses à forte valeur patrimoniale sur les dunes de Kervillen-Kervourden (Morbihan). Bulletin de la Société botanique du Centre-Ouest, NS, 44 : 583-595.
Hekking W.H.A., 1959. Un inventaire phytosociologiques des dunes à la côte méditerranéenne française entre Carnon et le Grau du Roi (département de l’Hérault). Mededelingen van het Botanisch Museum en Herbarium van de Rijksuniversiteit te Utrecht 161 : 518-532.
Hohenester A., 1967. Silbergrasfluren in Bayern. Mitteilungen der Floristisch-soziologischen Arbeitsgemeinschaft 11-12 : 13-21.
Klika J., 1931. Die Pflanzengesellschaften und ihre Sukzession auf den entblössten Sandböden in dem mittleren Elbetale. Sborník Československé Akademie Zemědelské 89 : 277-302.
Klika J., 1934. Studien über die xerotherme Vegetation Mitteleuropas. III – Die Pflanzengesellschaften auf Sandböden des Marchfelds in der Slowakei. Beihefte zum botanischen Centralblatt 52 : 1-16.
Klika J. & Novák J. (eds), 1941. Praktikum rostlinné sociologie, půdoznalství, klimatologie a ekologie. Praha (in Czech).
Kuhnholtz-Lordat G., 1931. L’association à Rosa pimpinellifolia L. et Ephedra distachya L. de la presqu’île de Quiberon (Morbihan). Annales de l’ École nationale d’agriculture de Montpellier 20 : 1-20.
Lemée G., 1937. Recherches écologiques sur la végétation du Perche. Thèse de doctorat, Paris, 387 p.
Loiseau J.-E. & Felzines J.-C., 2007. Les groupements des pelouses à Corynephorus canescens des vallées de l’Allier et du cours moyen de la Loire (Auvergne, Bourgogne, Centre ; France) Nouvelle composition des Corynephoretalia canescentis. Journal de botanique de la Société botanique de France 39 : 55-77.
Loisel R., 1976. La végétation de l’étage méditerranéen dans le Sud-Est continental français. Thèse de doctorat d‘État, Aix-Marseille, 384 p.
Losa Quintana J.M., 1975. Étude de l’association Festuco-Coremetum albae (Gimenez & Losa) Losa par la méthode computationnelle de Ceska et Roemer. Documents phytosociologiques 9-14 : 197-201.
Molinier R., 1954. Observations sur la végétation de la zone littorale en Provence. Vegetatio V-VI : 257-267.
Muller S., 2008. La lande basse à Diapensia lapponica de l’archipel de Saint-Pierre et Miquelon et sa signification biogéographique. Revue d’écologie (Terre et Vie) 63 : 391-396.
Nègre R., 1978. Données phytosociologiques sur les associations thérophytiques du Maroc aride. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 23-32.
Neto C., Fonseca J.P., Costa J.C. & Bioret F., 2015. Ecology and phytosociology of endangered psammophytic species of the Omphalodes genus in western Europe. Acta Botanica Gallica: Botany Letters 162 (1) : 37-54.
Paradis G., & Piazza C., 1991. Contribution à l’étude de la végétation des dunes du site classé de l’Ostriconi (Corse). Bulletin de la Société botanique du Centre-Ouest, NS, 22 : 149-182
Paradis G. & Piazza C., 1993. Description phytosociologique et cartographique de la végétation des dunes de Tizzano, de Tralicetu et de la plage d’Argent (sud-ouest de la Corse). Bulletin de la Société botanique du Centre-Ouest, NS, 24 : 219-266.
Petitclerc P. & Dignard N., 2004. La situation de l’hudsonie tomenteuse (Hudsonia tomentosa Nuttall) au Québec. Ministère des Ressources naturelles, de la Faune et des Parcs, direction de l’Environnement forestier et direction de la Recherche forestière, Québec, 23 p.
Piazza C. & Paradis G., 1995. Description phytosociologique et cartographique de la végétation du site protégé de Roccapina (Corse, France) : dune et zone humide. Documents phytosociologiques, NS, XV : 211-233.
Pinto Gomes C., Paiva-Ferreira R., Cano E. & Mendes S., 2006. Pelouses psammophiles à Corynephorus canescens var. maritimus Godr. du centre et du sud du Portugal. Acta Botanica Gallica 153 (3) : 341-354.
Rivas Goday S., 1957 (publié 1958). Nuevos órdenes y alianzas de Helianthemetea annuae Br. Bl. Anales del Instituto Botánico A.J. Cavanilles 15 : 539-651.
Rivas Goday S. & Rigual A., 1959. Algunas asociaciones de la provincia de Alicante. Anales del Instituto Botánico A.J. Cavanilles 16 : 533-548.
Rivas Goday S. & Borja Carbonell J., 1961. Estudio de vegetación y flórula del macizo de Gúdar y Jabalambre. Anales del Instituto Botánico A.J. Cavanilles 19 : 3-350.
Rivas-Martínez S., 1978. Sur la syntaxonomie des pelouses thérophytiques de l’Europe occidentale. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 55-69.
Rivas-Martínez S., Díaz T.E., Fernández-González F., Izco J., Loidi J., Lousã M. & Penas A., 2002. Vascular plant communities of Spain and Portugal; addenda to the syntaxonomical checklist of 2001. Itinera Geobotanica 15 : 5-922.
Royer J.-M., 1971. À propos de quelques observations phytosociologiques sur le sud du département de la Nièvre (régions de Decize et Nevers). Annales scientifiques de l’université de Besançon 10 : 117-131.
Royer J.-M., 2015. Contribution au prodrome des végétations de France : les Trifolio medii-Geranietea sanguinei T. Müll. 1962. Documents phytosociologiques, 3e série, 2 : 4-150.
Royer J.-M. & Ferrez Y., 2018. Contribution au prodrome des végétations de France : les Sedo albi-Scleranthetea biennis Braun-Blanq. 1955. Documents phytosociologiques, 3e série, 7 : 180-281.
Sánchez Mata D., 1989. Flora y vegetación del macizo oriental de la Sierra de Gredos (Avila). Diputación Provincial de Avila, Institución Gran Duque de Alba, 440 p.
Sissingh G., 1974. Comparaison du Roso-Ephedretum de Bretagne avec des unités de végétation analogues (contribution à la systématique des associations de dunes grises atlantiques et méditerranéennes). Documents phytosociologiques 7-8 : 95-106.
Tison J.-M. & de Foucault B., 2014. Flora Gallica, Flore de France. Biotope, Mèze, xx + 1196 p.
Tüxen R., 1928. Vegetationsstudien im nordwestdeutschen Flachlande, I – Über die Vegetation der nordwestdeutschen Binnendünen. Jahrbuch der geographischen Gesellschaft zu Hannover : 71-93.
Tüxen R., 1937. Die Pflanzengesellschaften Nordwestdeutschlands. Mitteilungen der Floristisch-soziologischen Arbeitsgemeinschaft in Niedersachsen 3 : 1-170.
Vanden Berghen C., 1962-63. L’association à Helichrysum stoechas dans les dunes du littoral du sud-ouest de la France (Roseto-Ephedretum Kuhnholtz-Lordat). Vegetatio 11 : 317-324.
Vanden Berghen C., 1965. La végétation de l’île Hoëdic (Morbihan, France). Bulletin de la Société royale de botanique de Belgique 98 : 275-294.
Wattez J.-R., Géhu J.-M. & de Foucault B., 1978. Les pelouses à annuelles des buttons de la Brenne. Colloques phytosociologiques VI, La végétation des pelouses sèches à thérophytes : 191-199.
Wattez J.-R. & Wattez A., 1995. Les landes à Éricacées et les formations landicoles annexes subsistant dans la région alréenne (département du Morbihan). Documents phytosociologiques, NS, XV : 153-181.
Remerciements
à J.-M. Lewin, qui m’a permis de bien préparer ma campagne de terrain dans les Pyrénées-Orientales, et à Jean-Roger Wattez, qui m’a fait découvrir les arrière-dunes de Plouharnel et de la Trinité-sur-Mer (Morbihan) fin juin/début juillet 2019.


